
Abstract
A taxonomic review of the Sebastes vulpes complex (S. vulpes, S. zonatus and S. ijimae) established the existence of two valid species, Sebastes vulpes Döderlein in Steindachner and Döderlein 1884 and Sebastes zonatus Chen and Barsukov 1976, despite evidence of hybridization between them. Similarities between the species include the following: top of cranium armed with robust nasal, preocular, postocular, tympanic and parietal spines; interorbital space flat; anterior and posterior lacrimals without distinct spines, forming two blunt lobes; thickened rays in ventral half of pectoral fin; dorsal fin usually with 13 spines and 13 soft rays; caudal fin rounded; and pored lateral-line scales usually 30–35. However, S. zonatus is distinguishable from S. vulpes in usually having distinct vertical dark bands on the dorsum (vs. usually lacking), minute cycloid scales usually present posteriorly on the lower jaw (vs. usually absent) and present on the entire basal portion of the spinous dorsal-fin membrane (vs. absent below first to ninth or to last dorsal-fin spine). Based on specimen and literature records, S. vulpes inhabits depths of 0–50 m, ranging from Hokkaido southward to Shimane and Sagami Bay, Japan, and along the southern coast of the Korean Peninsula, whereas S. zonatus inhabits depths of 50–100 m, from Hokkaido southward to Shimane and Tosa Bay, including the Seto Inland Sea, and along the southern coast of the Korean Peninsula. Sebastodes ijimae Jordan and Metz 1913 is considered to be a junior synonym of S. vulpes, based on examinations of type and other genetically assigned specimens. A lectotype is designated for S. vulpes.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The live-bearing rockfish genus Sebastes Cuvier 1829 (Scorpaenoidei: Sebastidae), the most speciose in the family, comprises ca. 110 species worldwide (Kai et al. 2003; Hyde and Vetter 2007; Nelson et al. 2016). The greater part of such diversity is attributable to recently diverged species pairs, indicative of ongoing diversification processes (Kai et al. 2002, 2011, 2012; Narum et al. 2004; Burford 2009). Some of these species pairs have been shown to hybridize in nature, possibly generating novel genotypes and thereby facilitating further evolutionary changes via natural and/or sexual selection (Seeb 1998; Roques et al. 2001; Buonaccorsi et al. 2005, 2011; Hawkins et al. 2005; Hyde et al. 2008; Stefánsson et al. 2009; Kai et al. 2011; Muto et al. 2013). Such species pairs have provided valuable opportunities for evolutionary studies, shedding light on the origin of and diversification within the genus (e.g., Johns and Avise 1998; Rüber and Zardoya 2005; Ingram 2010; Ingram and Kai 2014). However, they have also caused considerable confusion from a taxonomic perspective; recently diverged species pairs are typically morphologically similar to each other and subsequent hybridization generates intermediate individuals. Notwithstanding, such species pairs have recently been studied using advanced genetic analyses that have successfully clarified the species status of the pairs (e.g., Orr and Hawkins 2008; Kai and Nakabo 2008, 2013; Frable et al. 2015).
Three commercially important western North Pacific species, viz., Sebastes vulpes Döderlein in Steindachner and Döderlein 1884, Sebastes ijimae (Jordan and Metz 1913) and Sebastes zonatus Chen and Barsukov 1976, have been the subject of taxonomic confusion. Döderlein in Steindachner and Döderlein (1884) first described S. vulpes, subsequently listed as valid by Jordan and Metz (1913), who described a second species, Sebastodes ijimae. Matsubara (1943) recognized both as valid, noting two color variants within S. vulpes. Later, Chen and Barsukov (1976) described one of the latter color variants as a separate new species, S. zonatus. Subsequently, the three species have been variously considered as either valid (e.g., Lindberg and Krasyukova 1987; Amaoka 1988; Nakabo 1993, 2000, 2002; Barsukov 2003; Kim IS et al. 2005; Nakabo and Kai 2013) or as intraspecific variations due to their morphological similarity or the presence of morphologically intermediate individuals (e.g., Kanayama and Kitagawa 1983; Kitagawa et al. 2008; Ishida 2009). Furthermore, the apparent rarity of S. ijimae has occasionally resulted in descriptions of the species based on a limited number of specimens (e.g., Matsubara 1943; Lindberg and Krasyukova 1987).
Recently, Muto et al. (2011) used comprehensive genetic and morphological analyses to demonstrate that S. vulpes and S. zonatus were separate species, characterized mainly by body coloration and gill-raker numbers, thereby generally agreeing with Chen and Barsukov (1976). The former subsequently showed that the two species hybridize in nature, nevertheless maintaining distinct gene pools in sympatry (Muto et al. 2013). However, they did not refer to S. ijimae, its relationships with the other two species thus remaining unclear.
Nominal species herein collectively referred to as the S. vulpes complex include Sebastes vulpes Döderlein in Steindachner and Döderlein 1884, Sebastodes ijimae Jordan and Metz 1913 and Sebastes zonatus Chen and Barsukov 1976. The present study first delineated separate species within the complex by a genetic assignment analysis. Once a genetic identification was established, a morphological analysis was conducted to clarify their morphological differences. Critical examination of type specimens and comparisons with genetically examined specimens confirmed the validity of S. vulpes and S. zonatus. Sebastes ijimae was regarded as a junior synonym of S. vulpes. Detailed redescriptions of S. vulpes and S. zonatus are provided and a lectotype designated for S. vulpes, in order to avoid further taxonomic confusion. A brief description of hybrids between the two species is also given.
Materials and methods
Genetic analysis. Amplified fragment length polymorphism (AFLP) analysis, which is useful for discriminating among Sebastes vulpes, Sebastes zonatus and their hybrids (Muto et al. 2011, 2013), was undertaken. A total of 216 specimens were subjected to the analysis. These included nine referable to Sebastes ijimae (uniformly black body) and all of the specimens examined by Muto et al. (2011, 2013), plus additional specimens representing wide geographic areas within the distributional range of the S. vulpes complex. Genomic DNA for each specimen was extracted from preserved muscle tissue using the DNeasy Tissue Kit (Qiagen), according to the manufacturer’s protocol. AFLP profiles were generated with the AFLP Plant Mapping Kit (Applied Biosystems), following the manufacturer’s protocol slightly modified by Kai et al. (2002). For selective amplification, 12 primer pairs were used following Muto et al. (2011, 2013); Mse I + Eco RI (ACA + CAA, AAG + CAG, ACA + CTT, ACA + CAC, ACT + CTA, ACA + CTG, AAG + CAC, AGG + CAT, ACC + CTT, ACG + CAC, AAG + CAT, AGG + CTT). Selective amplification products were analyzed on an ABI PRISM 310 genetic analyzer (Applied Biosystems), together with a GeneScan-500 Rox size standard (Applied Biosystems). Fragment data were collected using Peak Scanner software, vers. 1.0 (Applied Biosystems). Electropherograms were scored for the presence or absence of fragments between 90 base pairs (bp) to 450 bp in size, so as to create binary matrices. Fragments were inferred as homologous if they differed by ≤ 0.5 bp from the median.
Bayesian-based individual assignment was conducted using the program STRUCTURE 2.3.3 (Pritchard et al. 2000), in order to differentiate between genetically isolated species among the specimens, based on AFLP genotypes. The analysis was conducted assuming admixture and correlated allele frequency models, and a recessive genotype mode without any prior population information. Ten replicates for each predefined number of genetic groups (K) (= 1–10) were obtained with runs of 100 000 burn-in and 250 000 Markov chain Monte Carlo (MCMC) iterations. To determine the most likely value of K, the delta K approach according to Evanno et al. (2005), implemented in the program STRUCTURE HARVESTER (Earl and vonHoldt 2012), which determines the second-order rate of change in the distribution of L (K), was used. Each specimen was then assigned to an inferred group/s based on the individual proportion of membership (qi). Individuals were assigned to one group if qi ≥ 0.90 (purebred individuals) or jointly to two or more groups if the proportion of membership to each was qi < 0.90 (hybrid individuals). This threshold was used to give an optimal balance between the efficiency and accuracy of assigning individuals as purebreds or hybrids (Vähä and Primmer 2006), the value having been widely adopted in previous studies seeking to separate hybrids from purebreds (e.g., Burgarella et al. 2009; Sanz et al. 2009). CLUMPP v. 1.1.2 (Jakobsson and Rosenberg 2007) was used to summarize parameters across ten replicate runs and DISTRUCT v.1.1 (Rosenberg 2004) to produce the corresponding graphical output.
Morphological analysis. Morphological analysis was conducted on a total of 317 specimens, including all 216 specimens examined genetically. Measurements and counts followed Muto et al. (2011). Standard length is abbreviated as SL. All measurements are given as percentages of SL. The last two soft rays of the dorsal and anal fins were counted as single rays, each pair being associated with a single pterygiophore. Counts of gill rakers were taken from the first gill arch on the right side. Head spine terminology followed Randall and Eschmeyer (2001). To provide an objectively defined score that summarized the major components of variable measurements between the specimens, principal component analysis (PCA) (based on 13 measurements) was conducted for every specimen used in the morphological analysis via a correlation matrix. Raw morphometric data were standardized by log transformation prior to PCA. Subsequently, PCs that were considered shape components (see “Results” section) were used to perform canonical discriminant analysis (CDA). The CDA was initially applied to only purebreds of the two species, as determined by the STRUCTURE analysis, yielding a linear function (CDF) that maximized the variance between group centroids in multivariate space. The CDF was then used to calculate a single variable (CDF value) for each specimen, which described the morphology of the specimen relative to a set of predefined groups (in the present case, purebreds of the two species as assigned by the STRUCTURE analysis). Both PCA and CDA were conducted using R 3.0.3 (R Core Team 2014).
Because of the lack of a single diagnostic morphological character, specimens not examined genetically were identified primarily from the combination of body coloration, CDF value derived from 13 measurements, and presence/absence of minute scales basally on the dorsal-fin membrane and posteriorly on the lower jaw. Differences in these characters were clarified following comparisons with genetically examined specimens. Preserved color descriptions were based on type and non-type specimens, with fresh color descriptions being restricted to non-types only. Institutional codes followed Sabaj (2016), with one addition: “PKU”, Pukyong National University, Busan, Korea. In addition to literature records, the online database “FishPix” (available at http://fishpix.kahaku.go.jp/fishimage/index.html), jointly managed by KPM and NSMT, was accessed for distributions of each species. Characters given in the description of the complex were not repeated in individual species’ descriptions.
Results
Genetic analysis. The STRUCTURE analysis based on AFLP yielded a highest delta K value of 65.36 for K = 2 (delta K values for K = 3 to 9 ranged from 0.03 to 13.47), indicating that two genetic groups (species) were most likely represented (Evanno et al. 2005). Each specimen was then identified as a purebred or hybrid of the two groups, based on individual proportion of membership (qi). Sixty-seven specimens were assigned to one of the purebred groups (qi ≥ 0.90) and 70 to the other (qi ≤ 0.10) (Fig. 1), indicating two separate species within the Sebastes vulpes complex, corresponding to S. vulpes and Sebastes zonatus (sensu Muto et al. 2011, 2013), respectively. All nine specimens with a uniformly black body, referable to Sebastes ijimae, were included in the former species and treated as conspecific with it in the following description. The remaining 79 specimens were recognized as hybrids (0.10 < qi < 0.90), being excluded from the following species’ descriptions.

Model-based individual genetic assignment by STRUCTURE v. 2.3.3 based on AFLP with predefined number of genetic groups (K) = 2. Vertical bars represent individual probabilities of assignment to each group (= species). Individuals ranked from highest to lowest qi, with qi ≥ 0.90, qi ≤ 0.10 and 0.10 < qi < 0.90 representing Sebastes vulpes, S. zonatus and hybrids, respectively. Arrows indicate specimens with uniformly black body, previously recognized as “S. ijimae”
Morphological analysis. Distributions of selected counts of genetically examined specimens are shown in Table 1. In the PCA based on 13 measurements, the first principal component (PC) accounted for 95.6 % of the variation [Electronic supplementary material (ESM) Table S1]. Because all loadings were negative, it was considered as a size component. Principal components 2 to 13 were shape components, with both positive and negative loadings (ESM Table S1). These components were then used to perform a CDA, revealing a single function (CDF) separating genetically examined specimens of S. vulpes and S. zonatus [CDF = 0.0308 × PC2 + 0.3061 × PC3 + 1.0567 × PC4 + 1.4217 × PC5 + (-0.0125) × PC6 + 1.0655 × PC7 + (-4.5481 × PC8) + 4.1405 × PC9 + 1.1028 × PC10 + (-3.0283) × PC11 + 5.2715 × PC12 + 5.0000 × PC13]. That function was then used to calculate a CDF value for each specimen, ranging from -3.73 to 2.13, -1.14 to 2.88 and -1.96 to 2.49 in genetically examined specimens of S. vulpes, S. zonatus and hybrids, respectively (Fig. 2).

Distributions of canonical discriminant function (CDF) values derived from 13 morphometric characters of genetically examined specimens of Sebastes vulpes, S. zonatus and their hybrids. Data is presented as box plots with medians represented by horizontal lines with the 75th and 25th percentiles at the top and bottom of the boxes, respectively. The bar extends to 1.5 times the inter-quartile range (equivalent to box length). Outliers are represented as open circles that extend beyond 1.5 times the inter-quartile range
The Sebastes vulpes complex
Diagnosis. A species complex within the genus Sebastes from the western North Pacific with the following combination of characters: top of cranium armed with robust nasal, preocular, postocular, tympanic and parietal spines; interorbital space flat; anterior and posterior lacrimals without distinct spines, forming two blunt lobes; dorsal fin usually with 13 spines and 12–13 soft rays; pectoral-fin rays usually 17, ventral 7–9 rays unbranched and somewhat thickened; caudal fin rounded; pored lateral-line scales usually 30–35.
Description. Body oblong, relatively high. Nape rounded. Dorsal profile of head straight or slightly convex. No tentacles on head and body, except for nostril tentacle.
Mouth terminal, large, slightly oblique. Posterior margin of maxilla variable, extending from just short of to beyond level of posterior margin of orbit. Maxilla covered with thick skin. Lower jaw may or may not protrude beyond tip of upper jaw, with weak symphysial knob in larger specimens (over ca. 250 mm SL). Upper and lower jaws with bands of villiform teeth. Palatines and vomer with villiform teeth, latter forming V-shaped patch.
Nasal spine simple, sharp, directed dorsoposteriorly. Preocular spine simple, directed dorsoposteriorly, embedded, except for tip, which may or may not reach level with anterior margin of pupil. Postocular spine well developed, directed dorsoposteriorly, embedded, except for tip. Tympanic spine simple, directed dorsally. Interorbital space flat with weak ridge. Parietal spine well developed, somewhat divergent posteriorly in dorsal view. Supracleithral spine flattened, moderate or weak, directed posteriorly. Upper posttemporal spine well developed, embedded, except for dorsoposteriorly directed tip. Lower posttemporal, sphenotic, supraocular and pterotic spines absent. Lacrimals without distinct spines, forming a single or two rounded lobes (if latter, posterior lobe below anterior part of orbit). Suborbital without spines or ridge. Preopercle with five spines; upper two spines large, directed dorsoposteriorly; third and fourth moderate, blunt, directed posteriorly; fifth small, blunt, directed ventroposteriorly. Opercle with two simple flattened spines; upper spine larger, directed dorsoposteriorly, lower spine directed posteriorly.
Dorsal-fin origin slightly anterior to anterodorsal portion of gill slit. Posterior margin of soft dorsal fin rounded. Anal-fin origin below soft-rayed portion of dorsal fin, slightly anterior to midpoint. Posterior margins of anal, caudal and pectoral fins rounded.
Sebastes vulpes Döderlein in Steindachner and Döderlein 1884
(Standard Japanese name: Kitsune-mebaru) (Figs. 1, 2, 3a–e, 4a, 5a, b, 6a; Tables 1, 2)

Photographs of Sebastes vulpes (a–e) and S. zonatus (f–j) when fresh. a FAKU 96156, 191.0 mm SL, Miyako, Iwate, Japan; b FAKU 97669, 203.1 mm SL, Tomakomai, Hokkaido Japan; c FAKU 96099, 195.3 mm SL, Noto, Ishikawa, Japan; d FAKU 96100, 170.0 mm SL, Noto, Ishikawa, Japan; e FAKU 130099, 182.1 mm SL, Noto, Ishikawa, Japan; f FAKU 97079, 174.2 mm SL, Noto, Ishikawa, Japan; g FAKU 96275, 139.5 mm SL, Miyako, Iwate, Japan; h FAKU 97538, 198.1 mm SL, Tomakomai, Hokkaido, Japan; i FAKU 97671, 217.5 mm SL, Tomakomai, Hokkaido, Japan; j FAKU 97541, 288.6 mm SL, Otaru, Hokkaido, Japan

Schematic diagrams of Sebastes vulpes and S. zonatus showing dorsal-fin membrane and lower jaw squamation patterns. aS. vulpes, FAKU 96075, 218.5 mm SL, Ushitsu, Noto, Ishikawa, Japan; bS. zonatus, FAKU 97084, 154.1 mm SL, Noto, Ishikawa, Japan

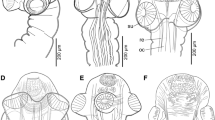
Type specimens of nominal species of Sebastes vulpes complex. a Lectotype of Sebastes vulpes (designated herein), NMW 74386, 256.8 mm SL; b holotype of Sebastodes ijimae, FMNH 55437, 157.5 mm SL; c holotype of Sebastes zonatus, SIO 75-475, 166.1 mm SL

Distributions of Sebastes vulpes, S. zonatus and their hybrids. Closed circles based on specimens examined in this study; open circles based on unequivocal literature records. aSebastes vulpes; bS. zonatus; c hybrids. Gray arrows indicate major ocean current routes
Sebastes vulpes Döderlein in Steindachner and Döderlein 1884: 203 (type locality: Tokyo, Japan); Steindachner and Döderlein 1887: pl. 2; Kanayama and Kitagawa 1983: 39, unnumbered fig. (upper color photo) (in part; Iwate, Japan); Lindberg and Krasyukova 1987: 70, fig. 29 [illustration after Steindachner and Döderlein (1887)] (Japan and Busan, Korea); Nakabo 1993: 516 (in part; Japan); Nakabo 2000: 593 [in part; followed Nakabo (1993)]; Nakabo 2002: 593 [in part; followed Nakabo (1993)]; Kitagawa et al. 2008: 59, unnumbered fig. of blackish individual (in part; Pacific coast of Tohoku district, Japan); Muto et al. 2011: 431, fig. 2-A (Ishikawa, Japan); Muto et al. 2013: 1755 (Hokkaido, Iwate and Ishikawa, Japan); Nakabo and Kai 2013: 679 (in part; Japan and southern and eastern Korean Peninsula).
Sebastes (Neohispaniscus) ijimae: Matsubara 1943: 231, fig. 81 (in part; Busan, Korea).
Sebastes ijimae: Chen and Barsukov 1976: 4, fig. 1-B (in part; Japan and Busan, Korea); Lindberg and Krasyukova 1987: 67, fig. 28 [illustration after Jordan and Metz (1913)] (Japan and Busan, Korea); Amaoka 1988: 298, pl. 360-A [illustration after Jordan and Metz (1913)] (Japan and southern Korean Peninsula); Nakabo 1993: 516, unnumbered fig. (Japan and southern Korean Peninsula); Nakabo 2000: 593, unnumbered fig. [followed Nakabo (1993)]; Nakabo 2002: 593, unnumbered fig. [followed Nakabo (1993)]; Youn 2002: 250 (Korea); Kim IS et al. 2005: 220, unnumbered fig. (eastern and southern coasts of Korea and Japan); Nakabo and Kai 2013: 679, unnumbered fig. (Japan and Busan, Korea).
Sebastes (Sebastodes) vulpes: Barsukov 2003: 260 (in part; 35º32’N, 140º50’E).
Sebastes zonatus: Chen and Barsukov 1976: 3 (in part; Japan).
Sebastichthys vulpes: Jordan and Starks 1904: 113 (in part; Japan).
Sebastodes vulpes: Jordan and Evermann 1898: 1835 (followed original description).
Sebastodes ijimae Jordan and Metz 1913: 49, pl. IX-1 (type locality: Busan, Korea).
Lectotype (new designation; Fig. 5 a). NMW 75386, 256.8 mm SL, Tokyo, Japan (purchased at Tokyo Fish Market), January 1883, collected by Franz Steindachner.
Paralectotypes. NMW 58146, 162.9 mm SL, collected with lectotype; ZMB 12069, 224.7 mm SL, Tokyo, Japan, collected by Ludwig Döderlein.
Paralectotype (Sebastes vulpes or Sebastes zonatus: excluded from following description). NMW 5878, 158.7 mm SL, Tokyo, Japan, 1885, collected by Franz Steindachner.
Other material: 95 specimens (including holotype of Sebastodes ijimae), comprising 67 subjected to meristic, morphometric and genetic examination [indicated by asterisk (*)], plus 28 subjected to meristic and morphometric examination only [indicated by double asterisk (**)]. JAPAN: FAKU 50136–50138**, 96152*, 96156*, 96161*, 96163*, 96164*, 96238*, 96239*, 96257*, 157.6–199.7 mm SL, Miyako, Iwate; FAKU 59316–59318**, 154.0–194.8 mm SL, Enome, Ishikawa; FAKU 74454**, 82514*, 83188–83197*, 151.1–243.2 mm SL, Noto, Ishikawa; FAKU 84026*, 84027*, 84029*, 84033**, 84035**, 158.7–199.2 mm SL, Otaru, Hokkaido; FAKU 84101*, 150.7 mm SL, Taro, Iwate; FAKU 96073*, 96074*, 96075**, 96076–96078*, 96097*, 96099*, 96100*, 131566*, 131567*, 131569*, 156.4–249.9 mm SL, Ushitsu, Ishikawa; FAKU 96082–96088*, 96090–96094*, 159.8–230.9 mm SL, Matsunami, Ishikawa; FAKU 97669*, 97672*, 199.9–203.1 mm SL, Tomakomai, Hokkaido; FAKU 97679–97681*, 239.6–262.3 mm SL, Rausu, Hokkaido; FAKU 98758–98760*, 206.6–216.3 mm SL, Shakotan, Hokkaido; FAKU 103040**, 204.5 mm SL, locality unknown; FAKU 130099*, 182.1 mm SL, Ukawa, Ishikawa; FAKU 130347*, 130348*, 165.2–179.1 mm SL, Fujinami, Ishikawa; FAKU 131533–131535*, 161.0–175.4 mm SL, Ogi, Ishikawa; FAKU 131536–131538*, 170.6–192.4 mm SL, Hime, Ishikawa; NSMT-P 74206**, 141.6 mm SL, locality unknown; NSMT-P 74458**, 94.4 mm SL, Aomori; NSMT-P 78914**, 126.3 mm SL, Sawara, Hokkaido; SIO 03-90** (1 of 2 specimens), 140.2 mm SL, Tokyo; SIO 75-481** (2 of 3), 171.5–198.8 mm SL, Hakodate, Hokkaido; UMMZ 142653**, 102.7 mm SL, Otaru, Hokkaido; UMMZ 191954**, 203.7 mm SL, Oki Island, Shimane; UMMZ 191982** (1 of 2), 192.2 mm SL, Obama, Fukui; UMMZ 197588**, 197.4 mm SL, Asamushi Marine Biology Station, Aomori; UMMZ 197589**, 220.6 mm SL, Setogashima, Hamada, Shimane; UMMZ 191984** (2 specimens), 218.5–233.7 mm SL, Tottori. KOREA: FMNH 55437**, 157.5 mm SL, Busan (holotype ofSebastodes ijimae); PKU 2492*, 193.1 mm SL, Wando, Jeollanamdo; PKU 4070*, 4072*, 143.3–160.3 mm SL, Tongyeong, Gyeongsangnamdo; PKU 4314*, 159.7 mm SL, Uljin, Gyeongsangbukdo; UMMZ 191951**, 238.9 mm SL, Kori Bay; UMMZ 197590** (2 of 4), 225.2–241.3 mm SL, Busan.
Diagnosis. Posterior part of lower jaw usually without scales; gill rakers usually 25–27, spinous dorsal-fin membrane usually naked basally from first to ninth spine, sometimes with minute scales from first to last spine; body and head dark brown but varying from light to dark gray dorsally, usually with white mottling dorsally and laterally, sometimes uniformly dark gray or black laterally, whitish or light gray ventrally; rarely two indistinct dark brown or dark gray saddles on body, enclosing small white mottles; dorsal, anal, caudal and pectoral fins dark brown, light or dark gray, bluish-gray or black, with small irregular white markings on membranes; pelvic fin brown or light gray.
Description. Counts and proportional measurements given in Table 2. Body relatively wide, usually greater than 50 % of head length at uppermost point of pectoral-fin base. Head covered with ctenoid scales, except for snout tip, chin, maxilla, upper and lower lips, and branchiostegal rays. Posterior part of maxilla usually with minute cycloid scales (including lectotype). Posterior part of lower jaw usually naked (including lectotype), sometimes with patches of minute cycloid scales. Body covered with ctenoid scales with some accessory scales posteriorly; embedded cycloid scales covering thorax. Spinous dorsal-fin membrane usually naked basally from first to ninth spine, sometimes with minute cycloid scales from first to last spine (naked from below first to 11th spine in lectotype), posteriorly gradually spreading distally. Entire soft dorsal-fin membrane covered with minute cycloid scales. Spinous anal-fin membrane covered basally with minute cycloid scales, sometimes spreading distally. Entire soft anal-fin membrane covered with minute cycloid scales. Pectoral-fin rays covered with minute cycloid scales, except for tips of lowermost 7–9 rays; membrane naked. Pelvic-fin rays covered with minute cycloid scales, fin base and membrane naked.
Dorsal fin with 13 (rarely 14) spines and 12–13 (rarely 14) soft rays (13 spines and 13 soft rays in lectotype); all soft rays branched. Dorsal-fin spines gradually increasing in length to fourth spine, fourth and fifth spines subequal, thereafter shortening to penultimate spine; last spine longer than penultimate spine (shorter in lectotype), forming anterior support of soft-rayed portion of dorsal fin. Anal fin with 3 spines and 6 (rarely 5 or 7; 6 in lectotype) soft rays. Second and third anal-fin spines usually subequal in length (including lectotype), second spine sometimes longer than third spine. Second anal-fin soft ray longest, posterior rays gradually shortening. Tip of pectoral fin reaching or nearly reaching (rarely extending beyond; nearly reaching in lectotype) anus; uppermost ray unbranched, next 7–9 (usually 8; 9 in lectotype) rays on upper half branched, lowermost 7–9 (usually 8, including lectotype) rays unbranched and somewhat thickened. Posterior tip of depressed pelvic fin slightly anterior to tip of pectoral fin, usually not reaching anus (including lectotype).
Gill rakers on ceratobranchial 6–8 (usually 7, including lectotype), on hypobranchial usually 18–20 (rarely 17 or 21; 23 in lectotype). Rakers on angle between ceratobranchial and hypobranchial slender, longish; adjacent rakers progressively shorter, blunter; two uppermost and lowermost rakers sometimes united.
Color when fresh (Fig. 3a–e). Body and head dark brown but varying from light to dark gray dorsally, usually with white mottling dorsally and laterally, sometimes uniformly dark gray or black laterally, whitish or light gray ventrally. Head with two indistinct irregular dark gray or dark brown bands radiating from eye; upper band extending onto subopercle. Lower part of maxilla margined with dark brown or dark gray. Lower jaw, preopercle, interopercle and pectoral-fin base light or dark gray. Thorax and pectoral-fin base sometimes with irregular dark brown or dark gray markings. Two indistinct dark brown or dark gray saddles sometimes present on body, enclosing small white mottles, extending onto dorsal-fin membrane; anterior saddle under fourth to 12th dorsal-fin spine, posterior saddle under soft dorsal fin; when saddles present, a third indistinct dark brown or dark gray saddle often apparent on caudal peduncle. Dorsal fin dark brown, light or dark gray, bluish-gray or black, with small irregular white markings. Anal fin dark brown, light gray, dark gray, bluish-gray or black, with white mottling. Caudal fin dark brown, light gray, dark gray, bluish-gray or black, with white mottling. White margins of soft-rayed portions of dorsal and anal fins, and caudal fin usually narrow or absent. Pectoral fin dark brown, dark gray, bluish-gray or black; ventroposterior part sometimes pale. Pelvic fin brown or light gray; membrane darker.
Color in preserved specimens (Fig. 5a, b). Head and body dark brown dorsally and laterally, lighter ventrally, uniformly dark brown or with light mottling laterally (with light mottling in lectotype). Indistinct saddles on body and caudal peduncle when fresh remain darker following preservation (absent in lectotype). Lower jaw light or dark brown (dark brown in lectotype), thorax light brown. Head sometimes with two faintly darker irregular bands radiating from eye (absent in lectotype). Lower part of maxilla sometimes with darker margin. Dorsal, anal and caudal fins dark brown with irregular lighter markings. White margins of soft-rayed portions of dorsal, anal, and caudal fins when fresh remain lighter following preservation (only caudal fin with narrow light margin in lectotype). Pectoral fin dark brown, sometimes lighter posteroventrally (including lectotype). Pelvic fin light brown; membrane darker.
Distribution and habitat.Sebastes vulpes is presently known from Japanese waters, from coastal Hokkaido southward to Shimane and Sagami Bay, and off the southern and eastern coasts of the Korean Peninsula (Fig. 6a), most specimens having been collected from 0–50 m depth. Two records from Akita, Japan (KPM-NR 0047234 and KPM-NR 0047235, both registered as S. ijimae) are available on FishPix.
Remarks. The above description was based on 67 specimens genetically assigned by the STRUCTURE analysis based on AFLP, plus additional specimens not subjected to the genetic analysis. The former included examples of S. vulpes examined by Muto et al. (2011, 2013), with the latter being regarded as conspecific due to having one or more of the following characters: lower CDF value (< -1.85); body with grayish to black ground color, without dark saddles on dorsum; minute scales absent posteriorly on lower jaw and basally on spinous dorsal-fin membrane from first to ninth spine.
Döderlein in Steindachner and Döderlein (1884) originally described S. vulpes based on several specimens from Tokyo, Japan, the number of specimens being unknown. Because a holotype was not designated at that time, all of the specimens used in the description are automatically considered to be syntypes of S. vulpes, according to Article 73.2 of ICZN (1999).
In the ZMB type catalog, Paepke and Fricke (1992) listed ZMB 12069 as one of the syntypes of S. vulpes. Subsequently, Amaoka and Fujii (1999) conducted a preliminary taxonomic and historical study on Döderlein’s collection of Japanese animals deposited at several European museums, noting as follows: “ZMB 12069: Sebastes vulpes Döderlein, Yedo, Döderl., syntype?”. Eschmeyer et al. (2016) listed four specimens as syntypes of S. vulpes, as follows: “Syntypes: (several) NMW 5878 (1), 58146 (1), 75386 (1); ?ZMB 12069 (1)”, all of which were available for morphological examination during the present study. These specimens, deposited as syntypes in their respective collections, are herein considered to have been used in the original description of S. vulpes by Steindachner and Döderlein (1884), and are accordingly regarded as syntypes.
The body coloration of the four syntypes has faded during their long period of preservation and the CDF values for these specimens were equivocal with respect to the two species recognized here. Nevertheless, three of the four syntypes (NMW 58146, NMW 75386 and ZMB 12069) lacked minute scales posteriorly on the lower jaw and basally on the spinous dorsal-fin membrane from the first to 11th spines, characters agreeing closely with the genetically assigned specimens used for the present description. Steindachner and Döderlein’s (1887) illustration of S. vulpes, in the succeeding part of the article in which S. vulpes was originally described, indicated the absence of dark saddles on a specimen of ca. 260 mm SL, [which clearly matched NMW 75386 (256. 8 mm SL)], dorsal fin with 13 spines and 13 soft rays, anal fin with three spines and six soft rays, pectoral fin with nine unbranched rays and nine distally branched rays, and the last dorsal-fin spine shorter than the penultimate dorsal-fin spine. Although the last-mentioned character of NMW 75386 is rather unusual in the S. vulpes complex, the other characters and body coloration are consistent with other specimens identified as that species. In order to forestall any further taxonomic problems, NMW 75386 is designated as the lectotype of S. vulpes, in accordance with Recommendation 74B (ICZN 1999). Although the number of hypobranchial gill rakers (23) in the lectotype was greater than in other specimens (17–21), such was considered as an intraspecific variation. The remaining syntypes become paralectotypes.
Among the paralectotypes, NMW 58146 and ZMB 12069 are most likely S. vulpes, based on the dorsal-fin and lower-jaw squamation patterns. However, NMW 5878 was damaged and cannot be identified with certainty as either of the species recognized here. Nonetheless, it was clearly a member of the S. vulpes complex, having the head with nasal, preocular, postocular, tympanic and parietal spines; interorbital space flat; lacrimal without a distinct spine; dorsal fin with 12 spines and 13 soft rays; anal fin with three spines and six soft rays; pectoral fin with nine unbranched rays and nine distally branched rays; pelvic fin with one spine and five soft rays; and lateral-line pores 33.
Jordan and Metz (1913) originally described Sebastodes ijimae from a single specimen (FMNH 55437, formerly CM 4172) from Busan, Korea, designating it as the “type” [now “holotype” of S. ijimae, according to Article 73.1 (ICZN 1999)]. Although the specimen had an equivocal CDF value with respect to the two species recognized here, it possessed 25 gill rakers and lacked minute scales posteriorly on the lower jaw, significant features of S. vulpes recognized here (Table 1). In addition, Jordan and Metz (1913) described the coloration of S. ijimae as follows: “color dusky, lighter below; fins all black, tipped with black, except the lower half of pectoral which has a broad, white margin; snout and top of head almost black”. The accompanying illustration (pl. IX-1) also clearly showed a uniformly dark body color, falling within the range of color variation of S. vulpes as presently recognized. Accordingly, S. ijimae (Jordan and Metz 1913) is considered a junior synonym of S. vulpes Döderlein in Steindachner and Döderlein 1884, the holotype of the former being conspecific with the latter.
Although Okada and Matsubara (1938), followed by Matsubara (1955), discriminated between S. vulpes and S. ijimae (placed in genus Sebastichthys by the former) on the basis of head length relative to longest dorsal-fin spine and pelvic-fin length, these characters were not found to be significant in this study.
Sebastes (Neohispaniscus) vulpes (sensu Matsubara 1943) was characterized on the basis of 31 specimens (no catalog numbers given) from Hakodate (5 specimens), Miyako (1), Fukushima (4) and Misaki (17) (Japan), and Wonsan (1) and Busan (3) (Korea). He suggested that the species included two color morphotypes, one having a light brown, light gray or dark brown ground color with three irregular blackish-brown cross bands (pl. I-1 of Matsubara 1943), and the other, a light brown ground color with many small white blotches, but lacking blackish bands (pl. I-2). The former morphotype was clearly consistent with specimens identified herein as S. zonatus, whereas the latter was equivocal, specimens lacking dark bands but having small white blotches being found in both S. vulpes and S. zonatus in the present study, although much more commonly in the former. Matsubara (1965) continued to recognize two morphotypes within S. vulpes, but provided an illustration for only the saddled form, reproduced from pl. I-1 of Matsubara (1943).
Matsubara’s (1943) redescription of Sebastes (Neohispaniscus) ijimae was based on four specimens (catalog numbers not given) from Busan, Korea, the color description stating “in formalin dark brown with 3 indefinite darker blotches, the first of which is under the base of the fourth to the ninth dorsal spines, the second below the entire base of the soft dorsal, and the last on the caudal peduncle”. A specimen with “indefinite darker blotches” (= indistinct dark saddle, as recognized here) was largely consistent with S. zonatus, although that condition is sometimes found in S. vulpes. On the other hand, the illustration of the species (fig. 81 of Matsubara 1943) is like S. vulpes as presently recognized, showing a black ground body color with a small number of somewhat lighter blotches and no saddles.
Chen and Barsukov (1976) recognized three species within the S. vulpes complex: S. vulpes, S ijimae and their new species, S. zonatus. They characterized S. vulpes based on 25 specimens as having a grayish body with dense lighter spots and 28–30 gill rakers. However, our examination of 14 of those specimens confirmed that nine represented S. zonatus as presently recognized (ESM Table S2). These include CAS-SU 53416, one of two specimens of SIO 75-478 (302.7 mm SL), two of four specimens of SIO 75-480 (156.6 mm and 198.8 mm SL), UMMZ 191983, UMMZ 191985, UMMZ 197585, and one of three specimens of USNM 71787 (155.4 mm SL), having two distinct dark saddles on the dorsum: and UMMZ 191953, having minute scales on the entire basal part of the spinous dorsal-fin membrane and posteriorly on the lower jaw (see also Remarks under S. zonatus for identification of UMMZ 197585). On the other hand, five specimens could not be identified to either species due to a lack of definitive characters or character inconsistency. These include one of two specimens of SIO 75-479 (281.6 mm SL), two of four specimens of SIO 75-480 (215.0 mm and 191.2 mm SL), USNM 48143, and one of three specimens of USNM 71787 (151.0 mm SL). The remaining 11 specimens, viz., one of two specimens of SIO 75-479, ZIN 37490, ZIN 37491, ZIN 42100, and seven specimens of ZIN 42099, were not available here. Chen and Barsukov’s (1976) photograph of S. vulpes, presently unidentifiable due to the equivocal body coloration, lacked a specimen registration number.
Chen and Barsukov (1976) characterized S. ijimae as having a uniformly blackish or grayish body, dark saddles indistinct if present, and 25–27 gill rakers, based on 25 specimens including the holotype. Examination of 20 of the non-type specimens showed 11 to be S. vulpes (ESM Table S2). These include one of three specimens of SIO 75-481 (198.8 mm SL: miswritten as “SIO 57-481” in Chen and Barsukov 1976), UMMZ 191954, one of two specimens of UMMZ 191984 (218.5 mm SL), UMMZ 197588, and two of four specimens of UMMZ 197590 (225.2 mm and 241.3 mm SL), having a uniformly dark body; UMMZ 191951, one of two specimens of UMMZ 191984 (233.7 mm SL), and UMMZ 197589, having a CDF value < -1.85; and one of three specimens of SIO 75-481 (171.5 mm SL) and one of two specimens of UMMZ 191982 (192.2 mm SL), having a relatively low CDF value and minute scales absent posteriorly on the lower jaw. Six specimens were identified as S. zonatus as follows (ESM Table S2): two specimens of UMMZ 191947, one of two specimens of UMMZ 191982 (230.7 mm SL), two of four specimens of UMMZ 197590 (144.1 mm and 155.0 mm SL), having two distinct dark saddles on the dorsum; and UMMZ 191948, having minute scales basally on the spinous dorsal-fin membrane and posteriorly on the lower jaw. The remaining three specimens, viz., one of three specimens of SIO 75-481 (182.6 mm SL), UMMZ 191950 and UMMZ 191952, were unidentifiable due to a lack of definitive characters. ZIN 42401 and 42402 (three specimens) were unavailable for examination.
Nakabo (1993, 2000, 2002) also recognized S. vulpes, S. ijimae and S. zonatus, but noted that their taxonomy should be reviewed. He characterized S. vulpes as having 26–29 gill rakers, indistinct vertical dark bands on the body, an extremely narrow (or absent) white margin on the caudal fin and a moderately compressed body (based on three FAKU specimens). One of these (FAKU 103040, locality unknown) was herein identified as S. vulpes, having a CDF value < -1.85. FAKU 42115, on which an illustration by Nakabo (1993, 2000, 2002) was based, could not be identified unequivocally as either S. vulpes or S. zonatus, being characterized by faint dark saddles on the body with numerous white blotches. FAKU 103039 was also unidentifiable.
Nakabo’s (1993, 2000, 2002) description of S. ijimae was based on three specimens (FAKU 59316–59318), each having a uniformly dark body and identified herein as S. vulpes. Diagnostic characters for S. ijimae proposed by Nakabo (1993, 2000, 2002), including 24–26 gill rakers and almost uniformly black body, match those established here for S. vulpes.
Sebastes zonatus (sensu Nakabo 1993, 2000, 2002), redescribed on the basis of FAKU 36310, FAKU 59312–59315, FAKU 58088–58090, FAKU 58206 and FAKU 103038, was characterized as having 26–29 gill rakers, usually distinct vertical dark bands on the body, a relatively broad white margin on the caudal fin and a strongly compressed body. Color variants (including an almost uniformly light body) were also noted and illustrated (FAKU 59313 and FAKU59315, almost uniformly light body and distinct dark lateral bands, respectively). Nakabo’s specimens (except FAKU 36310 and FAKU 58206) were herein identified as S. zonatus owing to their light ground body color with distinct dark saddles (indistinct saddles in FAKU 59313, which also had minute scales posteriorly on the lower jaw and on the entire basal portion of the spinous dorsal-fin membrane). Identifications of FAKU 36310 and FAKU 58206 were uncertain due to a lack of definitive characters.
Subsequently, Nakabo and Kai (2013) recharacterized S. vulpes, S. zonatus and S. ijimae, generally following Nakabo (1993, 2000, 2002), but modifying the gill raker counts following Muto et al. (2011), whose specimens are also applicable to the present study.
Youn’s (2002) account of S. vulpes, S. ijimae and S. zonatus from Korean waters apparently followed Nakabo (1993, 2000, 2002), with slight modifications. Sebastes ijimae was described as having a uniformly dark grayish body and maxilla extending beyond the posterior margin of the orbit. Sebastes vulpes and S. zonatus were distinguished from each other by aspects of the white margin on the caudal fin, only the upper and lower tips of the caudal fin being white in the former, compared with the entire posterior margin white in the latter. However, the caudal-fin margin varies considerably within the species recognized here. Accordingly, S. vulpes and S. zonatus (sensu Youn 2002) cannot be identified with certainty, whereas S. ijimae (sensu Youn 2002) body coloration is consistent with the present specimens of S. vulpes.
Sebastes (Sebastodes) ijimae (sensu Barsukov 2003) was described as having 25–27 gill rakers and uniformly dark body color, agreeing well with S. vulpes as presently recognized. However, the accompanying illustration based on a specimen collected from Tokyo, Japan, is problematic, showing several characters not attributable to the S. vulpes complex. These include lateral-line pores 37 (vs. 28–36 in S. vulpes complex), a distinct symphysial knob (vs. indistinct) and the soft dorsal-fin posterior margin almost straight (vs. rounded). Therefore, S. ijimae of Barsukov (2003) is unidentifiable here.
Kanayama and Kitagawa (1983), Kitagawa et al. (2008) and Ishida (2009) recognized only S. vulpes as valid, regarding S. zonatus and S. ijimae as conspecific with the former owing to the existence of equivocal specimens. Of the two photographs provided by Kanayama and Kitagawa (1983), the upper one is referable to S. vulpes as recognized here, owing to the black body with no saddles on the dorsum, and the lower one to S. zonatus, showing distinct saddles. Similarly, one of the two photographs in Kitagawa et al. (2008), labeled “blackish individual” by the authors, was identifiable as S. vulpes, the other one (“brownish individual”), reproduced from Kanayama and Kitagawa (1983), being identified as S. zonatus. Ishida (2009) provided three photographs, one (an adult) being referable to S. zonatus as presently recognized, and the others (juveniles) being unidentifiable. He also noted variations in body coloration, ranging from distinct dark saddles on the dorsum to a complete absence of such.
The figures and descriptions of S. vulpes by Amaoka et al. (1983, 2011), Amaoka (1988), Sasaki (2003), Kim IS et al. (2005) and Kim YU et al. (2005) are all of uncertain identity, due to equivocal body coloration and meristic counts with respect to S. vulpes and S. zonatus as recognized here. Similarly, the photo of S. vulpes by Masuda and Kobayashi (1994) was also unidentifiable due to the equivocal body coloration.
Amaoka’s (1988) description of S. ijimae was also equivocal, although the accompanying photograph was identifiable as S. vulpes, based on body coloration (uniformly dark). Kim IS et al.’s (2005) S. ijimae was identified as S. vulpes, judging from both the description and coloration (photo), the body being uniformly charcoal gray.
Sebastes vulpes has also been reported from the following coastal Japanese localities: Oshima, Yamaguchi, Seto Inland Sea (Katayama and Fujioka 1958), Ibaraki (Funabashi 1998), Sea of Japan coast of Fukuoka (Nishida et al. 2005), Choshi, Chiba (Aizawa 2003), Kasumi, Hyogo (Mori 1956), Toyama Bay (Katayama 1940) and Aomori (Shiogaki et al. 2004). However, the true taxonomic status of such references to S. vulpes remain equivocal, due to the lack of description. Accordingly, the listed localities have not been included in the present distribution of S. vulpes or S. zonatus. Similarly, Sebastichthys vulpes [from Hakodate, Japan (Snyder 1912); Busan, Korea (Jordan and Metz 1913)], Sebastodes vulpes [Japan (Jordan et al. 1913)] and Sebastodes (Pteropodus) vulpes [Otaru market, Hokkaido, Japan (Jordan and Hubbs 1925)] were listed without descriptions and remain of unknown status.
Sebastes zonatus Chen and Barsukov 1976
(Standard Japanese name: Tanuki-mebaru) (Figs. 1, 2, 3f–j, 4b, 5c, 6b)
Sebastes zonatus Chen and Barsukov 1976: 6, fig. 1-C (type locality: off Furube, near Usujiri, Hakodate, Hokkaido, Japan); Amaoka et al. 1983: 87, unnumbered fig. (Japan); Lindberg and Krasyukova 1987: 66, fig. 27 (Japan); Amaoka 1988: 298, pl. 278-D [photo reproduced from Amaoka et al. (1983)] (Japan); Nakabo 1993: 516, unnumbered figs. (Japan); Masuda and Kobayashi 1994: 84, fig. 3 (Japan); Nakabo 2000: 593, unnumbered figs. [followed Nakabo (1993)]; Nakabo 2002: 593, unnumbered figs. [followed Nakabo (1993)]; Kim IS et al. 2005: 225, unnumbered fig. (southern coast of Korean Peninsula and Japan); Amaoka et al. 2011: 186, unnumbered fig. (Hokkaido, Japan); Muto et al. 2011: 431, fig. 2-B (Ishikawa, Japan); Muto et al. 2013: 1755 (Hokkaido, Iwate, and Ishikawa, Japan); Nakabo and Kai 2013: 679, unnumbered figs. (Japan and southern and eastern coasts of Korean Peninsula).
Sebastes ijimae (not of Jordan and Metz): Chen and Barsukov 1976: 4 (in part; Japan and Busan, Korea).
Sebastes (Neohispaniscus) ijimae (not of Jordan and Metz): Matsubara 1943: 231 (in part; Busan, Korea).
Sebastes (Neohispaniscus) vulpes (not of Döderlein): Matsubara 1943: 230, pl. I-1 (in part; Japan).
Sebastes vulpes (not of Döderlein): Okada 1955: 314, unnumbered fig. [illustration after pl. I-1 of Matsubara (1943)] (Japan and Korea); Matsubara 1965: 426, unnumbered fig. [illustration after pl. I-1 of Matsubara (1943)] (Japan and Korea); Chen and Barsukov 1976: 2 (in part; Japan); Kanayama and Kitagawa 1983: 39, unnumbered fig. (lower color photo) (in part; Iwate, Japan); Nakabo 1993: 516 (in part; Japan); Nakabo 2000: 593 [in part; followed Nakabo (1993)]; Nakabo 2002: 593 [in part; followed Nakabo (1993)]; Kitagawa et al. 2008: 59, unnumbered fig. of brownish individual [photo reproduced from Kanayama and Kitagawa (1983)] (in part; Pacific coast of Tohoku district, Japan]; Ishida 2009: 193, unnumbered fig. of adult (Hokkaido, Japan); Nakabo and Kai 2013: 679 (in part; Japan and southern and eastern Korean Peninsula).
Sebastes (Sebastodes) vulpes (not of Döderlein): Barsukov 2003: 260, fig. 117 (in part; 35º32’N, 140º50’E).
Sebastes (Sebastodes) zonatus: Barsukov 2003: 266, fig. 121 [illustration after Lindberg and Krasyukova (1987)] (Japan).
Sebastichthys vulpes (not of Döderlein): Jordan and Starks 1904: 113 (in part; Japan); Okada et al. 1935: 205, pl. 114 (Japan and southern Korea).
Materials examined. 140 specimens, including the holotype and five paratypes, comprising 70 specimens subjected to meristic, morphometric and genetic examination [indicated by asterisk (*)], plus 70 subjected to meristic and morphometric examination only [indicated by double asterisk (**)]. JAPAN: BSKU 4996**, 179.8 mm SL, Kochi; BSKU 38831**, 131.0 mm SL, Yawatahama, Ehime; CAS 34077** (2 specimens), 177.7–194.1 mm SL, off Furube near Usujiri, Hakodate, Hokkaido (paratypes ofSebastes zonatus); FAKU 50126**, 50127**, 50129–50133**, 50139**, 84086*, 96143–96148*, 96149**, 96150*, 96154*, 96158*, 96159**, 96234–96237*, 96260*, 96261*, 96263–96270*, 96273–96276*, 96283*, 98032*, 98042*, 98043*, 131.2–359.5 mm SL, Miyako, Iwate; FAKU 58088–58090**, 271.4–305.8 mm SL, off San-in; FAKU 59312–59315**, 145.4–217.8 mm SL, Enome, Ishikawa; FAKU 82516–82519*, 82520**, 82521*, 97078*, 97079*, 97081**, 97084*, 137.3–174.2 mm SL, Wajima, Ishikawa; FAKU 83185–83187*, 252.1–286.4 mm SL, Noto, Ishikawa; FAKU 85799*, 257.5 mm SL, No, Niigata; FAKU 96095*, 129995*, 130103–130105*, 211.3–234.4 mm SL, Toyama Bay, Toyama; FAKU 97538–97540*, 97671*, 162.2–217.5 mm SL, Tomakomai, Hokkaido; FAKU 97541*, 288.6 mm SL, Otaru, Hokkaido; FAKU 97678*, 236.1 mm SL, Rausu, Hokkaido; FAKU 97981*, 274.6 mm SL, Soma, Fukushima; FAKU 98755*, 98757*, 98767–98769*, 98776*, 98782*, 98786*, 203.1–233.1 mm SL, Shakotan, Hokkaido; FAKU 103038**, 208.8 mm SL; FAKU 130100–130102*, 160.1–176.2 mm SL, Mawaki, Ishikawa; HUMZ 123423**, 123436**, 172.6–199.8 mm SL, Kinaoshi, Hokkaido; NSMT-P 73629**, 143.3 mm SL locality unknown; NSMT-P 73685**, 172.9 mm SL, Motoyoshi, Miyagi; NSMT-P 74275**, 175.0 mm SL, Natori, Miyagi; NSMT-P 74598**, 202.8 mm SL, Miyagi; NSMT-P 74625** (2), 238.4–239.9 mm SL, off Kamo, Yamagata; NSMT-P 74773**, 257.3 mm SL, Higashitsugaru, Aomori; NSMT-P 92096**, 148.4 mm SL, Usujiri, Hakodate, Hokkaido; SIO 75-475**, 166.1 mm SL, female (holotype ofS. zonatus), SIO 75-476** (3), 145.8–195.4 mm SL (paratypes ofS. zonatus), off Furube near Usujiri, Hakodate, Hokkaido; SIO 75-477**, 206.1 mm SL, 35º52’–35º48’N, 141º04’–141º08’E; SIO 75-478** (2), 217.8–302.7 mm SL, 35º32’–35º31’N, 140º50’–140º49’E; SIO 75-480** (2 of 4), 156.6–198.8 mm SL, Tokyo; UMMZ 191947** (2), 123.6–150.3 mm SL, off Kuro-zaki, Ojika Peninsula, Miyagi; UMMZ 191948**, 174.4 mm SL, Namerikawa, Toyama; UMMZ 191949**, 250.6 mm SL, off ?Usetsu; UMMZ 191982** (1 of 2), 230.7 mm SL, Wakasa bay, Coast of Oshima, near Obama, Fukui; UMMZ 191983**, 261.5 mm SL, Sendai, Miyagi; UMMZ 191985**, 198.0 mm SL, off No, Niigata; UMMZ 197585**, 178.0 mm SL, Choshi, Chiba; UMMZ 197586**, 214.3 mm SL, off Kyogamisaki, Kyoto; UMMZ 197587**, 208.1 mm SL, Toyama Bay, Toyama; UMMZ 212640**, 146.9 mm SL, Hakodate, Hokkaido; UMMZ 212687**, 148.0 mm SL, Kesen-numa, Miyagi; CAS-SU 53416**, 242.1 mm SL, Tokyo market; USNM 51314**, 71787** (2), 113.0–181.8 mm SL, Hakodate, Hokkaido; USNM 51369–51370**, 239.3–291.3 mm SL, locality unknown; USNM 205460**, 91.2 mm SL, Aomori. KOREA: FAKU 99752–99755**, 99756*, 148.1–193.8 mm SL, Busan; PKU 2490*, 147.7 mm SL, Wando, Jeollanamdo; PKU 4315*, 117.9 mm SL, Uljin, Gyeongsangbukdo; UMMZ 197590** (2 of 4), 144.1–155.0 mm SL, Busan.
Diagnosis. Posterior part of lower jaw usually with patches of minute scales; gill rakers usually 26–28, entire basal portion of spinous dorsal-fin membrane usually covered with minute scales, sometimes naked from first to last spine; body white, pinkish, light brown or gray dorsally and laterally, whitish ventrally, mottled with brown, dark brown or dark gray laterally; two distinct reddish-brown, dark brown or dark gray saddles with small white mottles on dorsum usually present, sometimes indistinct; dorsal, anal and caudal fins reddish-brown, light brown or light gray, mottled with light gray or white; pectoral fin pinkish, reddish-brown, dark brown or dark gray; pelvic fin light brown, reddish-brown or light gray.
Description. Counts and proportional measurements of examined specimens given in Table 2. Body relatively narrow. Head covered with ctenoid scales, except for snout tip, chin, maxilla, upper and lower lips, and branchiostegal rays. Posterior part of maxilla usually with minute cycloid scales (including holotype). Patches of minute cycloid scales usually present posteriorly on lower jaw (absent in holotype). Body covered with ctenoid scales, some accessory scales in posterior field; embedded cycloid scales covering thorax. Entire basal portion of spinous dorsal-fin membrane usually covered with minute cycloid scales, posteriorly gradually spreading distally (including holotype); sometimes naked from first to last spine (naked from first to seventh spine in holotype). Entire soft dorsal-fin membrane covered with minute cycloid scales. Basal portion of spinous anal-fin membrane covered with minute cycloid scales, sometimes spreading distally. Entire soft anal-fin membrane covered with minute cycloid scales. Pectoral-fin rays covered with minute cycloid scales, except for tips of lowermost 7–9 (8 in holotype) rays; membrane naked. Pelvic-fin rays covered with minute cycloid scales, fin base and membrane naked.
Dorsal fin with 13 (rarely 12) spines and 12–13 (rarely 11) soft rays (13 spines and 13 soft rays in holotype); all soft rays branched. Dorsal-fin spines gradually increasing in length to fourth spine, fourth and fifth spines subequal, thereafter shortening to penultimate spine; last spine longer than penultimate spine, forming anterior support of soft-rayed portion of dorsal fin. Anal fin with 3 spines and 6 (rarely 5 or 7; 6 in holotype) soft rays. Second anal-fin spine relatively long, robust, its length usually equal to or greater than third spine. Second anal-fin soft ray longest, posterior rays gradually shortening. Tip of pectoral fin reaching or nearly reaching (rarely extending beyond; reaching in holotype) anus; uppermost ray unbranched, next 7–9 (usually 8; 9 in holotype) rays on upper half branched, lowermost 7–9 (usually 8; 7 in holotype) rays unbranched, somewhat thickened. Posterior tip of depressed pelvic fin slightly anterior to tip of pectoral fin, usually not reaching anus (including holotype).
Gill rakers on ceratobranchial 7–8 (rarely 6 or 9), on hypobranchial 18–20 (rarely 21, 22 or 23) (7 and 20 in holotype). Rakers on angle between ceratobranchial and hypobranchial slender, longish; adjacent rakers progressively shorter, blunter; two uppermost or lowermost rakers sometimes united.
Color when fresh (Fig. 3f–j). Body whitish, pinkish, light brown or gray dorsally and laterally, whitish ventrally, mottled with brown, dark brown or dark gray laterally. Head brown, dark brown or dark gray with two dark brown or dark gray irregular narrow bands radiating from eye; upper band extending onto subopercle. Lower part of maxilla margined with dark brown or dark gray. Tip of snout, upper and lower jaws, preopercle, interopercle and pectoral-fin base pinkish, light brown or light gray. Thorax and pectoral-fin base sometimes with irregular dark brown or dark gray markings. Two distinct reddish-brown, dark brown or dark gray saddles with small white mottles usually present on dorsum, sometimes indistinct, rarely absent; anterior saddle under fourth to 12th dorsal-fin spine, posterior saddle under soft-rayed portion of dorsal fin. Caudal peduncle with indistinct reddish-brown, dark brown or dark gray saddle. Head and pectoral-fin base usually with dark brown or dark gray vermiculations. Spinous dorsal fin white, reddish-brown or light brown or light gray, mottled with light gray or white, margined with white or pink in lighter specimens; dark saddle on dorsum extending onto membrane. Soft-rayed portion of dorsal fin brown, light brown or gray, mottled with white or light gray. Anal fin brown, light brown or gray, mottled with white or light gray; membrane between second and third spines dark gray or black. Caudal fin brown, gray or dark gray, mottled with white or light gray. White margins of soft-rayed portions of dorsal and anal fins and caudal fin usually broad, sometimes narrow, rarely absent. Pectoral fin pinkish, reddish-brown, dark brown or dark gray, dorsoposterior portion usually darker, ventroposterior portion usually lighter; uniformly pink in lighter specimens. Pelvic fin light brown, reddish-brown or light gray; membrane darker.
Color in preserved specimens (Fig. 4c). Head and body light brown laterally and ventrally, mottled with dark brown laterally. Two distinct saddles on dorsum when fresh remain darker than surrounding area following preservation (including holotype). Head with two darker irregular narrow bands radiating from eye. Lower part of maxilla margined with dark brown. Spinous dorsal fin light brown with irregular darker markings; dark saddle on dorsum extending onto membrane. Soft dorsal fin dark brown with white mottling basally. Anal fin dark brown with irregular light mottling; membrane between second and third spines darker. Caudal fin dark brown with irregular light mottling. White margins of soft-rayed portions of dorsal and anal fins and caudal fin when fresh remain lighter following preservation. Pectoral fin light brown. Pelvic fin light brown; membrane darker.
Distribution and habitat.Sebastes zonatus is presently known from Japanese waters, from coastal Hokkaido southward to Tosa Bay, Yamaguchi and the Seto Inland Sea, and off the southern Korean Peninsula (Fig. 6b). The following records are available on FishPix: one specimen each of KPM-NR 0000457 and KPM-NR 0001021 (from Kasumi, Hyogo, Japan), KPM-NR 0090941 (Kashiwazaki, Niigata, Japan), KPM-NR 0090942 (Sakata, Yamagata, Japan) and KPM-NR 0106833 (Pacific coast of Hokkaido, Japan), all registered as S. zonatus. Usually collected from 50–150 m depth.
Remarks. The present description is based on 70 specimens [including those examined by Muto et al. (2011, 2013)] genetically assigned by STRUCTURE analysis, plus additional specimens. The latter were considered conspecific with the former, having the following features: high (> 2.49) CDF value; pinkish to light brown body color with two distinct dark saddles on dorsum; and minute scales posteriorly on the lower jaw or on the entire basal portion of the spinous dorsal-fin membrane.
Chen and Barsukov (1976) considered the two color morphotypes within Sebastes vulpes (sensu Matsubara 1943) (see “Remarks” under S. vulpes) as representing separate species, describing one of them as new, viz., S. zonatus. The holotype of S. zonatus (SIO 75-475), collected off Furube, near Usujiri, Hakodate, Hokkaido, Japan, exhibited distinct dark saddles on a light ground color (Fig. 5c), clearly agreeing with one of the two species genetically assigned by the STRUCTURE analysis, on which the present description is based. Therefore, the species described here is identified as S. zonatus. Paratypes of S. zonatus, CAS 34077 (two specimens) and SIO 75-476 (three specimens), collected together with the holotype, were also considered as conspecific with the species herein described, owing to their body coloration. Among the other specimens examined and identified by Chen and Barsukov (1976) as S. zonatus, seven also agreed with specimens identified here as S. zonatus, all having distinct dark saddles on the dorsum (ESM Table S2). These include one of two specimens of SIO 75-477 (206.1 mm SL), one of two specimens of SIO 75-478 (217.8 mm SL), UMMZ 191949, UMMZ 197586, UMMZ 197587, USNM 51314, and one of three specimens of USNM 71787 (181.8 mm SL). However, UMMZ 142653 was identified as S. vulpes owing to the absence of minute scales on the lower jaw and basal portion of the spinous dorsal-fin membrane from the first to ninth spine, although the body color was not definitive. The second specimen of SIO 75-477 and material registered as ZIN 37492, ZIN 42096, ZIN 42099 a, ZIN 42093, ZIN 42094, ZIN 42095 and ZIN 42098, plus three additional specimens listed by Chen and Barsukov (1976) (catalog numbers not given) were unavailable for examination.
According to Chen and Barsukov (1976), UMMZ 197585 (identified therein as S. vulpes) and UMMZ 197587 (S. zonatus) were 208 mm SL and 184 mm SL, respectively. However, their actual lengths (178.0 mm SL and 208.1 mm SL, respectively) suggest that the specimens had been confused. Both were identified herein as S. zonatus, having distinct dark saddles on the dorsum.
Lindberg and Krasyukova (1987) recognized S. vulpes, Sebastes ijimae and S. zonatus following examination of specimens deposited in ZIN, a subset of those examined by Chen and Barsukov (1976) and not available here. The former reproduced illustrations of S. vulpes and S. ijimae from those of the lectotype (designated here) (NMW 75386; Steindachner and Döderlein 1887) and holotype (FMNH 55437; Jordan and Metz 1913), respectively. They also provided an illustration of S. zonatus (based on ZIN 42095, 193 mm SL, Pacific coast of Japan), which clearly agreed in body coloration with that species as presently recognized. Because their accompanying descriptions of the three species generally followed Chen and Barsukov (1976), S. zonatus (sensu Lindberg and Krasyukova 1987) was most likely that species as presently recognized, whereas the remaining two probably included both S. vulpes and S. zonatus [as did those of Chen and Barsukov (1976); see “Remarks” for S. vulpes].
Barsukov (2003) reproduced an illustration in Lindberg and Krasyukova (1987) to accompany his description of S. zonatus in a worldwide review of rockfishes, noting that it was based on a 160 mm SL specimen, collected from 33º11’N, 133º31’E (Tosa Bay, Pacific coast of Japan), but without providing the registration number.
Barsukov (2003) also noted Sebastes (Sebastodes) vulpes as having 28–30 gill rakers and reticulated markings on the body sometimes aggregating to form vertical bands (= dark saddles). However, the supporting illustration, based on a specimen collected from 35º32’N, 140º50’E (off Chiba, Pacific coast of Japan), is attributable to S. zonatus as recognized here, owing to the presence of distinct saddles. Therefore, S. vulpes (sensu Barsukov 2003) most likely comprised both S. vulpes and S. zonatus.
The photographs of S. zonatus shown in Amaoka et al. (1983, 2011), Amaoka (1988) and Kim IS et al. (2005) are identifiable as that species, based on their body coloration. Those authors described S. zonatus as having distinct dark saddles on the dorsum, with the gill-raker count and body coloration of Amaoka’s (1988) S. zonatus apparently following Chen and Barsukov (1976). A photograph of S. zonatus in Masuda and Kobayashi (1994) also matches the species as presently recognized, unlike a photograph labeled S. zonatus by Kim YU et al. (2005), which was unidentifiable owing to indistinct saddles on the dorsum.
Jordan and Starks’ (1904) Sebastichthys vulpes, described as having a “sometimes distinct broad 1/2 bar of dark brown under middle of spinous dorsal and a less distinct one under soft dorsal”, should be re-identified as S. zonatus, as should Okada et al.’s (1935) Sebastichthys vulpes, having a “yellowish light-purplish-brown body with broad brown bands on lateral body”. Okada’s (1955) illustration of Sebastichthys vulpes is identified as S. zonatus, having been reproduced from Matsubara (1943: pl. I-1).
Hybrids between Sebastes vulpes and Sebastes zonatus
(Figs. 1, 2, 6c, 7; Tables 1, 2)

Photographs of hybrids between Sebastes vulpes and S. zonatus when fresh. a FAKU 98033, 199.2 mm SL, Miyako, Iwate, Japan; b FAKU 98030, 184.2 mm SL, Miyako, Iwate, Japan; c FAKU 97673, 199.1 mm SL, Tomakomai, Hokkaido, Japan
Specimens examined (as inferred by genetic assignment). 79 specimens. JAPAN: FAKU 82517, 97077, 97080, 97082, 97083, 162.0–189.0 mm SL, Wajima, Noto, Ishikawa; FAKU 84028, 84034, 97542, 162.9–236.1 mm SL, Otaru, Hokkaido; FAKU 96096, 96098, 131568, 184.8–210.3 mm SL, Ushitsu, Noto, Ishikawa; FAKU 96151, 96153, 96155, 96157, 96160, 96162, 96240, 96241, 96258, 96259, 96262, 96271, 96272, 96277–96282, 98030, 98031, 98033–98037, 136.6–362.8 mm SL, Miyako, Iwate; FAKU 97670, 97673, 199.1–225.5 mm SL, Tomakomai, Hokkaido; FAKU 97682, 245.1 mm SL, Rausu, Hokkaido; FAKU 97980, 274.5 mm SL, Soma, Fukushima; FAKU 98752–98754, 98756, 98761–98766, 98770–98775, 98777–98781, 98783–98785, 200.7–257.1 mm SL, off Shakotan, Hokkaido; FAKU 98787, 220.6 mm SL, off Tango, Kyoto; FAKU 130235, 130236, 154.8–202.8 mm SL, Noto, Ishikawa; FAKU 130349–130351, 198.6–207.9 mm SL, Toyama Bay, Toyama. KOREA: PKU 2489, 2491, 184.4–207.7 mm SL, Wando, Jeollanamdo; PKU 4066, 4069, 4071, 152.1–170.4 mm SL, Tongyeong, Gyeongsangnamdo; PKU 4895–4897, 153.5–230.7 mm SL, Donghae, Gangwondo.
Description. Counts and proportional measurements given in Table 2. Morphological variations in hybrids widely overlapped characteristics of S. vulpes and S. zonatus: maxilla and lower jaw sometimes with minute cycloid scales posteriorly; basal portion of spinous dorsal-fin membrane sometimes with minute cycloid scales from first to last spine; gill rakers on ceratobranchial 6–8 (usually 7), on hypobranchial 18–21 (usually 19); body and head of fresh specimens pinkish, light brown, dark brown, light gray or dark gray dorsally, usually mottled with white dorsally and laterally, whitish or light gray ventrally; head with two dark gray or dark brown irregular bands radiating from eye; two reddish-brown, dark brown or dark gray saddles on body usually indistinct, sometimes distinct, rarely absent; reddish-brown, dark brown or dark gray saddle often present on caudal peduncle in specimens with similar saddles on body; dorsal, anal, and caudal fins reddish-brown, dark brown, light gray, dark gray, bluish-gray or black, with small irregular white markings; soft-rayed portions of dorsal, anal and caudal fins usually with white margins; pectoral fin pinkish, reddish-brown, dark brown, dark gray, bluish-gray or black, ventroposterior part usually lighter; pelvic fin reddish-brown, light brown or light gray, membrane darker; head and body of preserved specimens light or dark brown dorsally and laterally, lighter ventrally; dark saddles on body and caudal peduncle, and two dark bands radiating from eye when fresh remain darker than surrounding area following preservation; dorsal, anal and caudal fins dark brown with lighter mottling; white margins of soft-rayed portions of dorsal, anal and caudal fins when fresh remain light following preservation; pectoral fin light or dark brown, sometimes lighter posteroventrally; pelvic fin light brown, membrane darker.
Distribution. Hybrids between S. vulpes and S. zonatus are known from coastal Hokkaido southward to Fukushima and Kyoto, Japan, and the southern Korean Peninsula (Fig. 6c).
Remarks. The present study found hybrids of S. vulpes and S. zonatus from almost the entire area of overlap between the two species (Fig. 6). Morphological variations of the hybrids overlapped those of S. vulpes and S. zonatus, including the diagnostic characters of each species (Figs. 2, 7; Table 2). Therefore, discrimination of purebreds from naturally occurring hybrids, based on morphological examination alone, is not completely free from error; the morphologically equivocal specimens that remained unidentified after the present study are possibly of hybrid origin. Nevertheless, the study satisfactorily established that pure S. vulpes could be distinguished from pure S. zonatus.
Interspecific hybridization between closely related species has often been accompanied by taxonomic confusion (e.g., Orr and Hawkins 2008; Kai and Nakabo 2013; Frable et al. 2015), owing to its propensity to produce somewhat continuous entities among the participating taxa. The S. vulpes complex may be among the most difficult cases, extensive hybridization between S. vulpes and S. zonatus resulting in a considerable proportion of admixed and morphologically similar individuals. Nevertheless, the present study successfully overcame such difficulties by employing a combination of individual-based genetic assignment and multivariate morphological analyses, such being commonly used in evolutionary studies, but less often in a taxonomic context.
References
Aizawa M (2003) Myxini, Cephalaspidomorphi, Chondrichthyes, Osteichthyes. In: The foundation of Chiba Prefecture for the study of historical materials (ed) Checklist of animals known from Chiba Prefecture. Chiba Prefecture, Chiba, pp 304–335
Amaoka K (1988) Genus Sebastes. In: Masuda H, Amaoka K, Araga C, Uyeno T, Yoshino T (eds) The fishes of the Japanese Archipelago, 2nd edn. Tokai University Press, Tokyo, pp 310–313
Amaoka K, Fujii R (1999) Fishes of the Döderlein Collection. In: Nishikawa T (ed) Preliminary taxonomic and historical studies on prof. Ludwig Döderlein’s Collection of Japanese animals made in 1880–1881 and deposited at several European museums. Rep Act 1997–8 Grant-in-Aid Int Sci Res (Field Res) No. 090411155, pp 147–165
Amaoka K, Nakaya K, Yabe M (2011) Fishes of Hokkaido. Hokkaido Shimbun Sha, Sapporo
Amaoka K, Nakaya K, Yabu H, Yamamoto H (1983) Fishes and marine algae of northern Japan. Kitanihon Kaiyo Center Co Ltd, Sapporo
Barsukov VV (2003) Annotated and illustrated check-list of rockfishes of the world. In: Dorofeyeva E, Sideleva V (eds) Proc Zoo Inst Russ Acad Sci, vol 95, pp 1–320
Buonaccorsi VP, Kimbrell CA, Lynn EA, Vetter RD (2005) Limited realized dispersal and introgressive hybridization influence genetic structure and conservation strategies for brown rockfish, Sebastes auriculatus. Conserv Genet 6:697–713
Buonaccorsi VP, Narum SR, Karkoska KA, Gregory S, Deptola T, Weimer AB (2011) Characterization of a genomic divergence island between black-and-yellow and gopher Sebastes rockfishes. Mol Ecol 20:2603–2618
Burford MO (2009) Demographic history, geographical distribution and reproductive isolation of distinct lineages of blue rockfish (Sebastes mystinus), a marine fish with a high dispersal potential. J Evol Biol 22:1471–1486
Burgarella C, Lorenzo Z, Jabbour-Zahab R, Lumaret R, Guichoux E, Petit RJ, Soto Á, Gil L (2009) Detection of hybrids in nature: application to oaks (Quercus suber and Q. ilex). Heredity 102:442–452
Chen LC, Barsukov VV (1976) A study of the western North Pacific Sebastes vulpes species complex (Scorpaenidae), with description of a new species. Japan J Ichthyol 23:1–8
Cuvier G (1829) Des Sébastes (Sebastes, nob.). In: Cuvier G, Valenciennes A (eds) Histore naturelle des poissons, vol 4. Chez F. G. Levrault, Paris-Strasbourg, pp 326–350
Earl DA, vonHoldt BM (2012) STRUCTURE HARVESTER: a website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv Genet Resour 4:359–361
Eschmeyer WN, Fricke R, van der Laan R (2016) Catalogue of Fishes: genera, species, references. http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp. Accessed 28 Feb 2018
Evanno G, Regnaut S, Goudet J (2005) Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study. Mol Ecol 14:2611–2620
FishPix. see http://fishpix.kahaku.go.jp/fishimage-e/search.html. Accessed 28 Feb 2016
Frable BW, Wagman DW, Frierson TN, Aguilar A, Sidlauskas BL (2015) A new species of Sebastes (Scorpaeniformes: Sebastidae) from the northeastern Pacific, with a redescription of the blue rockfish, S. mystinus (Jordan and Gilbert, 1881). Fish Bull 113:355–377
Funabashi M (1998) Checklist of marine fishes from the coastal region of Ibaraki Prefecture. Bull Ibaraki Nat Mus 1:75–96
Hawkins SL, Heifetz J, Kondzela CM, Pohl JE, Wilmot RL, Katugin ON, Tuponogov VN (2005) Genetic variation of rougheye rockfish (Sebastes aleutianus) and shortraker rockfish (S. borealis) inferred from allozymes. Fish Bull 103:524–535
Hyde JR, Kimbrell CA, Budrick JE, Lynn EA, Vetter RD (2008) Cryptic speciation in the vermilion rockfish (Sebastes miniatus) and the role of bathymetry in the speciation process. Mol Ecol 17:1122–1136
Hyde JR, Vetter RD (2007) The origin, evolution, and diversification of rockfishes of the genus Sebastes (Cuvier). Mol Phylogenet Evol 44:790–811
ICZN (1999) International code of zoological nomenclature, 4th edn. International Trust of Zoological Nomenclature, London
Ingram T (2010) Speciation along a depth gradient in a marine adaptive radiation. Proc R Soc Lond B Biol Sci 278:613–618
Ingram T, Kai Y (2014) The geography of morphological convergence in the radiation of Pacific Sebastes rockfishes. Am Nat 184:E115–E131
Ishida M (2009) Scorpaenidae. In: Okamura O and Amaoka K (eds) Sea fishes of Japan, 3rd edn. Yama to Keikoku Sha, Tokyo, pp 189–209
Jakobsson M, Rosenberg NA (2007) CLUMPP: a cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics 23:1801–1806
Johns GC, Avise JC (1998) Tests for ancient species flocks based on molecular phylogenetic appraisals of Sebastes rockfishes and other marine fishes. Evolution 52:1135–1146
Jordan DS, Evermann BW (1898) The fishes of North and Middle America: a descriptive catalogue of the species of fish-like vertebrates found in the waters of North America, north of the Isthmus of Panama, part II. Bull US Nat Mus 47:i–xxx, 1241–2183
Jordan DS, Metz CW (1913) A catalog of the fishes known from the waters of Korea. Mem Carnegie Mus 6:1–65
Jordan DS, Hubbs CL (1925) Record of fishes obtained in Japan, 1922. Mem Carnegie Mus 10:93–346
Jordan DS, Starks EC (1904) A review of the Scorpaenoid fishes of Japan. Proc US Nat Mus 27:91–175
Jordan DS, Tanaka S, Snyder JO (1913) A catalogue of the fishes of Japan. J Coll Sci Imp Univ Tokyo 33:1–497
Kai Y, Nakabo T (2008) Taxonomic review of the Sebastes inermis species complex (Scorpaeniformes: Scorpaenidae). Ichthyol Res 55:238–259
Kai Y, Nakabo T (2013) Taxonomic review of the Sebastes pachycephalus complex (Scorpaeniformes: Scorpaenidae). Zootaxa 3637:541–560
Kai Y, Nakayama K, Nakabo T (2002) Genetic differences among three colour morphotypes of the black rockfish, Sebastes inermis, inferred from mtDNA and AFLP analyses. Mol Ecol 11:2591–2598
Kai Y, Nakayama K, Nakabo T (2003) Molecular phylogenetic perspective on speciation in the genus Sebastes (Scorpaenidae) from the Northwest Pacific and the position of Sebastes within the subfamily Sebastinae. Ichthyol Res 50:239–244
Kai Y, Nakayama K, Nakabo T (2011) Genetic and morphological divergence within the Sebastes pachycephalus complex (Scorpaeniformes: Scorpaenidae). Ichthyol Res 58:333–343
Kai Y, Park KD, Nakabo T (2012) The incomplete history of mitochondrial lineages between two rockfishes, Sebastes longispinis and Sebastes hubbsi (Scorpaeniformes: Scorpaenidae). J Fish Biol 81:954–965
Kanayama T, Kitagawa D (1983) Fishes of Iwate II, Soi-Menukerui. Iwate Fisheries Experimental Station, Kamaishi
Katayama M (1940) A catalogue of the fishes of Toyama Bay. Toyama J Nat His 3:1–28
Katayama M, Fujioka Y (1958) Fishes of Oosima-gun, Yamaguti Prefecture. Bull Fac Agr Yamaguchi Univ 9:1147–1168
Kim IS, Choi Y, Lee CL, Lee YJ, Kim BJ, Kim JH (2005) Illustrated book of Korean fishes. Kyohak Publishing Co Ltd, Seoul
Kim YU, Myoung JG, Kim YS, Han KH, Kang CB, Kim JK, Ryu JH (2005) Marine fishes of Korea, 2nd edn. Hanguel Graphics Co Ltd, Busan
Kitagawa D, Imamura H, Goto T, Ishito Y, Fujiwara K, Ueda Y (2008) Field guide of the fishes from the Tohoku district, north-eastern waters of Japan. Tokai University Press, Hadano
Lindberg GU, Krasyukova ZV (1987) Fishes of the Sea of Japan and adjacent parts of Okhotsk and Yellow Sea. Part 5. Nauka Publishers, Leningrad
Masuda H, Kobayashi Y (1994) Grand atlas of fish life modes. Tokai University Press, Tokyo
Matsubara K (1943) Studies on the scorpaenid fishes of Japan. Anatomy, phylogeny and taxonomy II. Trans Shigenkagaku Kenkyusho 2:171–486
Matsubara K (1955) Fish morphology and hierarchy, part II. Ishizakishoten, Tokyo
Matsubara K (1965) Scorpaenidae. In: Okada K, Uchida K, Uchida R (eds) New Illustrated Encyclopedia of the Fauna of Japan. Hokuryu-kan, Tokyo, pp 422–435
Mori T (1956) Fishes of San-in District including Oki Islands and its adjacent waters (Southern Japan Sea). Mem Hyogo Univ Agr 2:1–62
Muto N, Kai Y, Nakabo T (2011) Genetic and morphological differences between Sebastes vulpes and S. zonatus (Teleostei: Scorpaeniformes: Scorpaenidae). Fish Bull 109:429–439
Muto N, Kai Y, Noda T, Nakabo T (2013) Extensive hybridization and associated geographic trends between two rockfishes Sebastes vulpes and S. zonatus (Teleostei: Scorpaeniformes: Sebastidae). J Evol Biol 26:1750–1762
Nakabo T (1993) Scorpaenidae. In: Nakabo T (ed) Fishes of Japan with pictorial keys to the species. Tokai University Press, Tokyo, pp 491–518, 1295–1298
Nakabo T (2000) Scorpaenidae. In: Nakabo T (ed) Fishes of Japan with pictorial keys to the species, 2nd edn. Tokai University Press, Tokyo, pp 565–595, 1524–1528
Nakabo T (2002) Scorpaenidae. In: Nakabo T (ed) Fishes of Japan with pictorial keys to the species, English edn. Tokai University Press, Tokyo, pp 565–595, 1524–1528
Nakabo T, Kai Y (2013) Sebastidae. In: Nakabo T (ed) Fishes of Japan with pictorial keys to the species, 3rd edn. Tokai University Press, Hadano, pp 668–681, 1933–1934
Narum SR, Buonaccorsi VP, Kimbrell CA, Vetter RD (2004) Genetic divergence between gopher rockfish (Sebastes carnatus) and black and yellow rockfish (Sebastes chrysomelas). Copeia 4:926–931
Nelson JS, Grande TC, Wilson MVH (2016) Fishes of the world, 5th edn. John Wiley & Sons, Hoboken
Nishida T, Nakazono A, Oikawa S, Matsui S (2005) Changes of coastal fish fauna in the Chikuzen sea according to rise of sea water temperature in recent years. Sci Bull Fac Agr Kyushu Univ 60:187–201
Okada Y (1955) Fishes of Japan. Maruzen Co Ltd, Tokyo
Okada Y, Matsubara K (1938) Keys to the fishes and fish-like animals of Japan. Sanseido Co Ltd, Tokyo
Okada Y, Uchida K, Matsubara K (1935) Color atlas of fishes of Japan. Sanseido Co Ltd, Tokyo
Orr JW, Hawkins S (2008) Species of the rougheye rockfish complex : resurrection of Sebastes melanostictus (Matsubara, 1934) and a redescription of Sebastes aleutianus (Jordan and Evermann, 1898) (Teleostei: Scorpaeniformes). Fish Bull 106:111–134
Paepke HJ, Fricke R (1992) Kritischer Katalog der Typen der Fischsammlung des Zoologischen Museums Berlin. Teil 4: Scorpaeniformes. Mitt Zool Mus Berlin 68:267–293
Pritchard JK, Stephens M, Donnelly P (2000) Inference of population structure using multilocus genotype data. Genetics 155:945–959
R Core Team (2014) R: a language and environment for statistical computing. R foundation for Statistical Computing, Vienna. http://www.R-project.org/. Accessed 17 May 2018
Randall JE, Eschmeyer WN (2001) Revision of the Indo-Pacific scorpionfish genus Scorpaenopsis, with descriptions of eight new species. Indo-Pacific Fishes 34:1–79
Roques S, Sévigny JM, Bernatchez L (2001) Evidence for broadscale introgressive hybridization between two redfish (genus Sebastes) in the North-west Atlantic: a rare marine example. Mol Ecol 10:149–165
Rosenberg NA (2004) Distruct: a program for the graphical display of population structure. Mol Ecol Notes 4:137–138
Rüber L, Zardoya R (2005) Rapid cladogenesis in marine fishes revisited. Evolution 59:1119–1127
Sabaj MH (2016) Standard symbolic codes for institutional resource collections in herpetology and ichthyology: an Online Reference. Version 6.5 (16 August 2016). American Society of Ichthyologists and Herpetologists, Washington DC. http://www.asih.org/resources/standard-symbolic-codes-institutional-resource-collections-herpetology-ichthyology. Accessed 28 Feb 2018
Sanz N, Araguas RM, Fernández R, Vera M, García-Marín J (2009) Efficiency of markers and methods for detecting hybrids and introgression in stocked populations. Conserv Genet 10:225–236
Sasaki M (2003) Kitsune mebaru Sebastes vulpes Döderlein. In: Ueda Y, Maeda K, Shimada H, Takami T (eds) Fisheries and Aquatic Life in Hokkaido, Hokkaido Shimbun Sha, Sapporo, pp 194–195
Seeb LW (1998) Gene flow and introgression within and among three species of rockfishes, Sebastes auriculatus, S. caurinus, and S. maliger. J Hered 89:393–403
Shiogaki M, Ishito Y, Nomura Y, Sugimoto T (2004) Revised catalogue of the fishes collected from the waters of Aomori Prefecture. Bull Aomori Pref Fish Res Cent 4:39–80
Snyder JO (1912) Japanese shore fishes collected by the United States Bureau of Fisheries Streamer “Albatross” expedition of 1906. Proc US Nat Mus 42:399–450
Stefánsson MO, Sigurdsson T, Pampoulie C, Daníelsdóttir AK, Thorgilsson B, Ragnarsdóttir A, Gíslason D, Coughlan J, Cross TF, Bernatchez L (2009) Pleistocene genetic legacy suggests incipient species of Sebastes mentella in the Irminger Sea. Heredity 102:514–524
Steindachner F, Döderlein L (1884) Beiträge zur kenntniss der fische Japan’s. (III.). Denkschr Akad Wiss Wien 49:171–212, pls 1–7
Steindachner F, Döderlein L (1887) Beiträe zur Kenntniss der Fische Japan’s. (IV.). Denkschr Akad Wiss Wien 49:257–295, pls 1–4
Vähä JP, Primmer CR (2006) Efficiency of model-based Bayesian methods for detecting hybrid individuals under different hybridization scenarios and with different numbers of loci. Mol Ecol 15:63–72
Youn CH (2002) Fishes of Korea with pictorial key and systematic list. Acad Pub Com, Seoul
Acknowledgements
We thank K. Nakayama and M. Tagawa (FAKU) for helpful comments, G. Hardy (Ngunguru, New Zealand) for English corrections and valuable comments on the manuscript, and K. Sakai (Noto Marine Center) and T. Noda (Seikai National Fisheries Research Institute) for help in collecting specimens and valuable discussion. We also thank L. Parenti and J. Williams (USNM), W. Smith and K. Swagel (FMNH), W. Fink and D. Nelson (UMMZ), P. Hastings, H. Walker and C. Klepadlo (SIO), P. Bartsch and C. Lamour (ZMB), E. Mikschi and H. Wellendorf (NMW), M. Yabe, H. Imamura and T. Kawai (HUMZ), K. Matsuura, G. Shinohara and M. Nakae (NSMT), M. Motokawa (The Kyoto University Museum) and N. Nakayama (formerly The Kyoto University Museum), who facilitated the authors’ visits to examine specimens, J.-K. Kim (PKU), H. Endo (BSKU) and D. Catania (CAS) for loans of specimens and H.-J. Kwun (formerly PKU) for translating Korean literature. Our appreciation is also extended to the following who helped the collection of specimens and literature: Y. Koizumi and K. Koizumi (Otaru city), S. Matsui and K. Takahashi (Tomakomai city), C.-H. Jeong (Inha University) and N. Yoshimura (formerly HUMZ). This study was supported in part by JSPS KAKENHI Grant Numbers 19770063 and 20370034, the Mikimoto Fund for Marine Ecology and the Sasakawa Scientific Research Grant from the Japan Science Society.
Author information
Authors and Affiliations
Corresponding author
Additional information
This article was registered in the Official Register of Zoological Nomenclature (ZooBank) as 6E610D75-840F-43C9-A640-01179703E406.
This article was published as an Online First article on the online publication date shown on this page. The article should be cited by using the doi number.
Electronic supplementary material
Below is the link to the electronic supplementary material.
About this article

Cite this article
Muto, N., Kai, Y. & Nakabo, T. Taxonomic review of the Sebastes vulpes complex (Scorpaenoidei: Sebastidae). Ichthyol Res 66, 9–29 (2019). https://doi.org/10.1007/s10228-018-0641-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10228-018-0641-8