
Abstract
In Southeast Asian forests, there are dramatic supra-annual peaks in fruit availability known as mast fruiting, followed by low-fruit periods. Gibbons are frugivorous small apes. In this study, we investigated how gibbons varied their social and calling behaviors in response to changes in fruit availability. Activity budget, travel distance, sleeping time, and song duration of two wild Northern Gray gibbon (Hylobates funereus) groups were investigated during both mast and non-mast fruiting periods at the Danum Valley Conservation Area, Sabah, Malaysia. Gibbons traveled longer, entered the sleeping tree later, and spent more time engaged in singing, playing, and grooming behaviors during the mast fruiting period. Playing and grooming are intra-group social interactions. Gibbon songs were sung antiphonally within and among groups, indicating that singing is both an intra- and inter-group social interaction. Furthermore, copulatory behaviors that occurred during female pregnancy were observed during the mast fruiting period, suggesting this might be a social interaction behavior rather than a reproductive activity. Our results show that gibbons extended their daily activity schedules during the mast fruiting period and spent this extra time on social interactions. We examined whether male song complexity increased during the mast fruiting period by using the Levenshtein distance method, but we did not find such a trend. In the mast fruiting period, gibbons may consume more energy than in the non-mast fruiting period. Social and calling behaviors are energetically costly. Our results show that there is a link between energy intake and social and calling behaviors.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In the South-east Asian tropical rain forest region, many families of trees flower and fruit almost simultaneously, approximately every 2–10 years (Appanah 1985, 1993; Ashton et al. 1988). This fruit-rich event is known as mast fruiting (hereinafter abbreviated as “MF”) event. Between MF events, frugivorous animals must survive a long fruit-scarce (non-MF) period. Wong et al. (2005) observed famine-like emaciation or death in the sun bear (Helarctos malayanus) and in bearded pigs (Sus barbatus) during the non-MF period in the lowland tropical rainforest of Sabah, Malaysia. Variation in food availability may cause behavioral changes in animals. Frugivorous animals are free from hunger during the MF period, which often continue for several months. Many studies have indicated that primate activity budgets vary seasonally in response to changes in the abundance, quality, or distribution of food resources (Altmann and Muruthi 1988; Doran 1997; Estrada et al. 1999; Hanya 2004; Isbell and Young 1993; Passamani 1998; Robinson 1986; Vasey 2005).
Gibbons (Family Hylobatidae) are arboreal apes restricted to the tropical and sub-tropical forests of Southeast Asia, South China, Bangladesh, and Northeast India. Gibbons usually live in a monogamous social system consisting of a mated pair and their offspring. Gibbons are frugivorous (Gittins and Raemaekers 1980; Leighton 1987). They eat mostly ripe fruits, including large amounts of figs, which provide a stable food source to meet their basic energy requirements (Elder 2009). Northern Gray gibbons are found in the northern and eastern regions of Borneo (Groves 1972; Marshall and Marshall 1976). The diet of Northern Gray gibbon (Hylobates funereus) in Kalimantan comprised 62% fruits (Leighton 1987), while that of Northern Gray gibbon in the Danum Valley Conservation Area comprised 60% fruits (Inoue et al. 2016). Gibbons eat more fruits when it is available (Chivers 1974; Ahsan 1994; Bricknell 1999), although they also consume young leaves, flowers, insects (Ahsan 1994; Chivers 1984; Bartlett 2007), and bird eggs (Carpenter 1940). Previous studies have shown that in Southeast Asia, figs fruit at regular intervals independent of mast-fruiting, and are often used as a fallback food for primates (e.g., Marshall et al. 2009). Figs are included in the percentage values of fruits shown here.
The daily path lengths of gibbons vary seasonally in response to the abundance of preferred food resources (Raemaekers 1980; Bricknell 1999; Bartlett 2009; Inoue et al. 2016). Gibbons arrive at the sleeping trees several hours before nightfall (Chivers 1974; Reichard 1998; Ahsan 2001; Cheyne 2010; Inoue et al. 2016).
When gibbons ate nutritionally high-quality fruit, their feeding time decreased and traveling time increased because fruit is sparsely distributed (Bach et al. 2017). When gibbons ate energy-poor leaves, they may spend less time singing and playing to conserve energy (Bartlett 1999; Bach et al. 2017). Social interactions such as grooming, playing, and singing increase when ripe fruit was abundant (Bartlett 2003; Fan et al. 2008). Social behavior, especially singing, requires energy. Studies in birds have shown that muscles associated with breathing control, the syrinx, the upper vocal tract, and bill movement are all used during singing (Goller and Larsen 1997; Suthers et al. 1999; Larsen and Goller 2002). Field observations of birds suggest that singing has an energy cost. Birds sing frequently when they are in good physical condition (Lambrechts 1996). Fruits are rich in sugar and gibbons get their energy quickly by eating fruits (Raemaekers 1978). Knott (1998) reported that the foods most commonly eaten by orangutans during the mast-fruiting season were high in calories. Gibbons may take in more energy during the MF period than during the non-MF period and use the extra energy for social interactions that require high energy.
We collected behavioral and acoustic data from two groups of gibbons at the Danum Valley Conservation Area (DVCA) from 2002 to 2020. We collected behavioral data and acoustic recordings while following the gibbon groups. During the study period, MF occurred in 2005, 2007, 2010, and 2019. Based on previous studies showing the difference in activity budget between high and low fruit abundance (Bartlett 1999, 2003; Fan et al. 2008; Bach et al. 2017), we hypothesized that Northern Gray gibbons would show seasonal variation in activity patterns according to the change in fruit abundance between specifically fruit-rich MF and non-MF periods. In addition, we hypothesized gibbons spend more time on energetically costly activities such as grooming, playing, and calling behaviors during the MF period than during the non-MF period. To test these hypotheses, we investigated their activity budget, daily travel distance, daily sleeping time, and daily song duration during both MF and non-MF periods.
Previous bird studies showed that the performance of the bird’s song reflects food availability. Supplementary fed birds increased song rate (Berg et al. 2005). Less-fed birds reduced song rate, an increased latency to sing, and a lowered song amplitude and fundamental frequency (Ritschard and Brumm 2012). Birds with a larger repertoire size have a higher field metabolic rate, which may indicate that a more complex song reflects an energetic cost (Garamszegi et al. 2006). Furthermore, some studies show that domesticated songbirds develop more complex songs compared to their wild conspecifics, because of the relaxing of selection pressures associated with domestication (Takahasi and Okanoya 2010; Kagawa et al. 2014; Okanoya 2017). As it is assumed that domestication and fruit-rich environments are similar in terms of a stable food supply, we hypothesized that gibbon songs become complex when fruits are abundant. To test this hypothesis, we examined whether song complexity changed between MF and non-MF periods.
The goal of this study was to examine how gibbons change activity patterns, especially social and calling behaviors, in response to the amount of energy obtained from abundant fruit or poor fruit. The DVCA in Sabah, Malaysian Borneo, where Northern Gray gibbons occur, provides an opportunity to assess this problem.
Methods
Study area and subjects
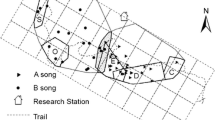
Gibbons are arboreal rainforest apes. Gibbons usually live in nuclear family groups, occupying a territory of between 20 to 100 hectares that they defend with vigorous visual and vocal displays (Cunningham and Mootnick 2009). Our study was conducted around the Borneo Rainforest Lodge (BRL) and the Danum Valley Field Center (DVFC) in the DVCA, located in Sabah, Malaysian Borneo (Fig. 1). This area consists mainly of primary rainforest. The Northern Gray gibbons we studied were members of two groups identified as SAPA and FC groups. The territory of the SAPA group is located near the Borneo Rainforest Lodge (BRL; 5° 01′ N, 117° 44′ E; elevation: ~ 190 m) and the territory of the FC group is located near the Danum Valley Field Center (DVFC; 5° 01′ N, 117° 48′ E; elevation: ~ 170 m). The DVFC is located 10 km southeast from the BRL (Fig. 1). Home range sizes of SAPA and FC groups were 34 ha and 35 ha, respectively. Boundary of home range was estimated from the cumulative day-range map of each group (Bartlett 2008). When we followed gibbon groups, we traced subject’s travel route on a map of the study site every 10 min, using a handheld global positioning system unit (Garmin GPSMAP 60CSx, Kansas City, USA). As neighbor groups were not habituated, we could not follow them. So, it was not clear how much home range overlapped with the neighboring groups. Thus, this home range approximation shows a total home range size including the overlap. The home ranges were estimated by using square-grid method. A grid with 100 m2 (10 m × 10 m) square cells was superimposed over the home range map, and the number of cells included in the area was counted. If more than one-half of any square was within the home range, we counted it as a full square, and if less than one-half of any square was within the home range, we ignored it. We collected activity data for three adult gibbons and two young gibbons: the SAPA adult male, the SAPA adolescent male, the FC adult male, the FC adult female, and the FC juvenile male. All gibbons were well habituated to humans. During our study period from 2002 to 2020, the SAPA group initially consisted of four individuals: an adult male, an adult female, a juvenile female, and an infant male. However, the adult female disappeared in April 2005, the subadult female left the SAPA group in October 2006, and the adolescent male died in April 2008. The SAPA adult male disappeared in August 2009; therefore, we used only using data from 2002 to 2009 for this group (Table 1). It was unclear why family members disappeared or died. Mast fruiting events occurred in 2005 and 2007 in this area (Kanamori et al. 2010). We studied the FC group from 2010 to 2020, which initially consisted of two individuals: an adult male and an adult female. During the study period, the pair had three offspring. However, two infants born in 2011 (January) and 2013 (June) disappeared in the successive years, in 2012 (August) and in 2014 (June) respectively. A third infant born in December 2015 was present until 2020 (Table 2). Mast fruiting events occurred in 2010 and 2019 in this area (Kanamori et al. 2017; Matsuzawa 2019).

Location of the Borneo Rainforest Lodge (BRL) and the Danum Valley Field Center (DVFC) in the Danum Valley Conservation Area, Sabah, Malaysia
Behavioral data collection
We collected activity data while following the two groups. With the aid of a field assistant, we conducted each survey over 5–9 successive days, 2–3 times a year. Behavioral observations were taken throughout full day follows by two people (Yoichi Inoue and a field assistant). Activity was focally observed. A scan sampling method at 10-min intervals was used to record the activity of gibbons from 0530 to 1600 h (Altmann 1974). For each observation sample, a 1-min scan was made recording the behavior of all visible members. Each individual was observed for 5 s, and its predominant behavior recorded. We recorded activity as either singing, moving, feeding (fig), feeding (non-fig fruit), feeding (leaves/flowers/others), playing, grooming, or resting. Infants and juveniles sometimes played by themselves, but we classified that as moving behavior. We classified playing behavior as that which occurred when two or more gibbons played together. Gibbons usually played by chasing or wrestling. We classified grooming behavior as that which occurred when two or more gibbons groomed each other. We classified self-grooming as that which occurred while at rest. The Wilcoxon signed-rank test was used to compare the proportion of playing and grooming behavior between MF and non-MF periods. We traced each focus subject’s travel route on a map of the study site every 10 min when following them, using a handheld global positioning system unit (Garmin GPSMAP 60CSx, Kansas City, USA). Travel distance and sleeping time (time of night-tree entry) were recorded as often as possible. Group travel routes were logged on a map of the study site by using a handheld GPS unit when following groups, and travel distances were measured using a digital curvimeter (COMCURVE-9JR Koizumi Sokki Mfg. Co., Ltd. Japan). Sleeping time was defined as the time at which the sleeping tree was entered. We recorded the time the subjects entered the sleeping tree and then continued to observe the group for at least 30 min until it went to sleep. At 0500 h the next morning, we went under the sleeping tree and observed the animals waking up to confirm their sleeping tree. Two-tailed t-tests were used to compare daily travel distance and daily sleeping time between MF and non-MF periods. We accumulated over 733 h (18 days during the MF period and 89 days during the non-MF period) of observations from 2002 to 2009 of the SAPA group and over 1085 h (30 days during the MF period and 139 days during the non-MF period) of observation from 2010 to 2020 of the FC group. Total observation days for activity budget, travel distance, and sleeping time are shown in Tables 1 and 2.
Acoustic data collection and statistical analysis
We started recording gibbon songs as soon as they started singing during the observation period. It allowed us to collect almost the complete song data. Gibbons sing species- and sex-specific songs (Marshall and Marshall 1976; Haimoff 1984). In the case of Northern Gray gibbons, males sing solo songs and mated pairs sing duets in a coordinated fashion (Haimoff 1985). We placed the recording device under the singing tree and attempted to record their songs as clearly as possible. We recorded songs using a digital audio recorder (R-09; Roland, Hamamatsu, Japan) with a microphone (ATM57; Audio-Technica, Tokyo, Japan). The recorder was set at a 44.1-kHz sampling rate and had 16-bit resolution. We measured the song duration through the recorded sound data. We converted the recorded sounds to spectrograms using Avisoft-SAS Lab Pro software (Avisoft, Berlin, Germany).
First, the sampling frequency was converted to 4000 Hz to allow focusing on the fundamental frequency for the on-screen measurements described below. Next, to remove ambient noise, we processed the sound through a high-pass filter to cut off sound at 500 Hz. Finally, spectrograms were created for on-screen measurements (settings: 256-point fast Fourier transformation and Hamming windows). We calculated daily male song duration and duet song duration from the spectrograms for each survey period (for example, survey periods in 2002 are August 2002 and December 2002), which is conducted 2–3 times a year, and used them for analysis. The Wilcoxon signed-rank test was used to compare daily male song duration and daily duet song duration between MF and non-MF periods. The number of male songs and duet songs recorded are shown in Tables 1 and 2.
A male song is a succession of phrases (Haimoff 1984), and a phrase is a succession of notes. A note is the minimum unit of song. Most of the note intervals in our subject males’ songs were <2 s (90.7%). Therefore, we identified different phrases within a song as being separated by pauses greater than 2 s. Haimoff (1985) classified notes of male Northern Gray gibbon songs into five types: “wa,” “oo,” “oo-wa,” “quaver-type notes,” and “trill” notes. However, neither our subject male (the SAPA male) nor observed neighboring males sang “oo-wa” or “quaver-type” notes during our study. As the trills comprised a short set of “wa” notes, we classified male songs into two note types: “wa” and “oo” (Inoue et al. 2017). We classified two notes, wa” and “oo,” through not only spectrogram figures but also acoustic characteristics (Inoue et al. 2020). The mean start and end frequencies of “wa” notes were higher than those of “oo” notes. The start frequency and minimum frequency always coincided in “wa” notes. The end frequency and maximum frequency also always coincided in “wa” notes. The duration of “wa” notes was shorter than that of “oo” notes. The frequency range of “wa” notes was wider than that of “oo” notes. “Wa” notes were characterized by a fast frequency increase. In contrast, the frequency fluctuation of “oo” notes was relatively small. Phrases of the SAPA male songs had various types of note combination (Fig. 2), and they had combinatory rules which might relate to behavioral contexts (Inoue et al. 2017). Inoue et al. (2017) investigated whether two successive notes in the SAPA male songs were emitted following any syntactical rules. As a result, frequencies of the two successive note types differed from expected prevalence in 45 songs out of 68 songs. This result indicated that male gibbon songs have combinatory rules. There were 2205 patterns of note combinations in phrases of 68 songs. The number of phrases in the SAPA male songs ranged between 6 and 702. The Levenshtein distance (LD) method provides a robust analytical tool for the study of animal vocal sequences (Kershenbaum and Garland 2015; Zanoli et al. 2020). The LD between two strings is the minimum number of substitution, insertion, or deletion operations required to turn one string into the other. We assigned one character (kanji; one of three Japanese alphabets) to each note combination, and then used a single string to represent the phrase of the SAPA male’s song. We calculated the LD for each pair of strings (package stringdist 0.9.6.3 in R; Van der Loo 2014) within MF and non-MF songs, normalized by dividing by the length of the longer of the two strings and then averaged LD values. The Wilcoxon signed-rank test was used to compare LD values between MF and non-MF periods.

Spectrograms of three phrases composed of “wa” and “oo” notes in the SAPA male songs. “w” denotes the “wa” note, and “o” denotes the “oo” note. Patterns of note combination composing three phrases were as follows: Phrase 1: wwwwwwwowoowwowwwwwww; Phrase 2: wwwwwwwwwowooowwwwwwwwww; Phrase 3: wwwwwwwwowwwwwwwwww
Results
Differences in activity budgets between MF and non-MF periods
Comparison of the activity budget during the MF period with that during the non-MF period of five gibbons revealed significant differences (Fig. 3; Supplementary Appendix Table 1). Gibbons spent more time singing, moving, feeding on fruits, playing, and grooming, and spent less time resting during the MF period. In particular, the proportion of playing and grooming behavior increased significantly during the MF period except for the FC adult female (Table 3). Adult males played with young gibbons in most cases. Copulation in the FC group was observed 5 times over 7 days in June 2019 and 17 times over 7 days in August–September in 2019. Since the FC female had a baby on October 31, 2019, she must have conceived in March and was pregnant from April to October (MF period). Our observations show that the FC female copulated during pregnancy in the MF period.

Activity budget of the SAPA adult male, FC adult male, FC adult female, SAPA adolescent, and FC juvenile during MF and non-MF periods
Difference in daily travel distance and sleeping time between MF and non-MF periods
Travel distance of the two groups was significantly different between MF and non-MF periods (Table 4). Sleeping time of the two groups was also significantly different between MF and non-MF periods (Table 4). In our study area (time zone; UTC/GMT + 8 h), sunrise time ranged from 0554 to 0625 h and sunset time ranged from 1750 to 1827 h.
Difference in daily song duration and song complexity between MF and non-MF periods
Daily male song duration during the MF period of the two groups was significantly longer than that during the non-MF period (Fig. 4; Wilcoxon signed-rank test, W = 39, p = 0.029), as was daily duet song duration of the FC group in the MF period although it was not significant (Fig. 5; Wilcoxon signed-rank test, W = 13, p = 0.053).

Box plots of daily male song duration in both MF and non-MF periods

Box plots of daily duet song duration in both MF and non-MF periods
Using LD method, we compared the complexity of note combinations between 18 songs during the MF period and 50 songs during the non-MF period of the SAPA male’s songs. There were 821 patterns of note combinations in 1817 phrases in 18 MF songs. On the other hand, there were 1906 patterns of note combinations in 6074 phrases in 50 non-MF songs. The mean LD value within songs of MF period was 0.968 (SD = 0.023, n = 153), and the mean LD value within songs of non-MF period was 0.971 (SD = 0.026, n = 1225). There was no significant difference in LD values within songs between MF and non-MF periods (Wilcoxon signed-rank test, W = 102,189, p = 0.068).
Discussion
In Southeast Asia, there is a special event called mast fruiting when food is extremely abundant. In this study, we demonstrated how gibbons varied their behaviors in response to changes in fruit availability. Daily travel distance during the MF period was longer, and daily sleeping time initiated later than that during the non-MF period. Gibbons were more active during the MF period. Gibbons might increase activity time and travel distance during the MF period because they presumably have sufficient nutrition intake from fruits during the MF period. Gibbons showed seasonal differences between MF and non-MF periods in their activity budgets. They spent more time engaged in singing, moving, feeding on fruits, playing, and grooming and less time in resting during the MF period. Gibbons spent a great deal of time playing and grooming significantly during the MF period. Playing and grooming were social interactions within a group. Daily male song duration and duet song duration increased during the MF period. Most of the male songs and duet songs in Northern Gray gibbons are sung antiphonally within and among groups (Mitani 1985; Inoue et al. 2013). In other words, singing is both an intra- and inter-group social interaction behavior. As a result, we conclude that gibbons extended their total daily activity schedules during the MF period and spent the extra time on both intra- and inter-group social interactions. Gibbons might have taken in more energy during the MF period than during the non-MF period and used the abundant energy for social interactions. Our results show that there is a link between energy intake and social and calling behaviors.
However, our results may be limited by the small sample size. We took data from only two groups. Gibbons increased singing, moving, feeding on fruits, playing, and grooming, and decreased resting when they ate more fruit in the MF period. This suggests a relationship between fruit intake and gibbon behaviors. Although our sample size was small, our results are consistent with previous studies on the link between energy intake and social and calling behaviors (Bartlett 1999, 2003; Fan et al. 2008, 2012; Bach et al. 2017). When ripe fruit was abundant, gibbons exhibited very high rates of social activities such as grooming, playing, and social contact (Bartlett 2003). When gibbons consumed less fruit and more buds, they decreased their level of social activities such as grooming, playing, and singing (Fan et al. 2012). The novelty of this study is that we compared gibbon behaviors between MF and non-MF periods. Because fruits were much more abundant during the MF period than during the non-MF period, we could see more clearly the differences in gibbon behaviors between the two periods. Future research on a large number of groups over a wide area will improve our conclusions.
In this study, there were interesting observations in the behavior of the FC adult female that has not yet been reported. She did not exhibit much playing behavior during both MF and non-MF periods. She conceived in March 2019 and was pregnant from April to October (MF period) in 2019, giving birth on October 31, 2019, after seven months of pregnancy. This might explain why she did not show much play behavior during both MF and non-MF periods. Copulations in the FC group were observed during the MF period even though the female was pregnant. In this case, copulations were non-reproductive sexual behavior and can be considered a type of social interaction.
Daily male song duration and duet song duration increased during the MF period. Our results are consistent with the previous report that male and female gibbons in 21 populations, especially males, showed reduced singing performance when the availability of high-energy foods was reduced (Cowlishaw 1996). Clink et al. (2020) showed that male solo songs in the DVCA appear to be influenced by environmental factors such as rain in the preceding night. Rain in the preceding night led to fewer calling events and led to a later start time of male solos. In this study, rainfall in the preceding night (rain that fell between 18:00 the night before and 06:00 in the morning) occurred on 15 days (31.3%) in 48 following days during the MF period and 68 days (29.8%) in 228 following days during the non-MF period. The percentage of days when it rained on the preceding night to the total number of following days was almost the same between the MF and non-MF periods, suggesting that the effect of rain on male songs was not different between the two periods. We counted how many times male gibbons sang songs after rainfall in the preceding night. They sang songs 8 times (53.3%) in 15 days during the MF period and 13 times (19.1%) in 68 days during non-MF period. Male gibbons sang songs more frequently during the MF period after rainfall in the preceding night, suggesting that they might have enough energy to sing despite the energy loss from the rain.
Among social behaviors, singing especially requires energy. Okanoya (2017) indicated that courtship songs of Bengalese finches (Lonchura striata var. domestica) became complex during the course of domestication. In the wild, gibbons vary their behaviors in response to the danger of the predator (Uhde and Sommer 2002). For example, gibbons’ choice of sleeping tree is driven primarily by predator avoidance (Cheyne et al. 2012). Although an MF event is not the same as the domestication process because predators exist in nature and MF does not continue throughout the year, an abundant food supply is a common character shared between MF and domestication. We hypothesized that gibbons might have developed a complex song during the fruit-rich MF period. However, there was no significant difference between MF and non-MF periods in the number of patterns of note combinations in the SAPA adult male’s songs, suggesting that male gibbon songs did not become more complex when fruits are abundant. The evolution of song complexity may not be achieved by temporary fruit abundance alone, but may require many more factors. Future research will be needed to clarify this issue.
In the present study, we found that gibbons increased social interactions such as playing, grooming, and singing in the MF period, although male song complexity did not increase. In addition, non-reproductive sexual behavior which could be considered as social interaction was observed in the MF period. Our results suggest that social interaction requires high energy cost and may develop in a fruit-rich environment.
References
Ahsan MF (1994) Behavioural ecology of the hoolock gibbon (Hylobates hoolock) in Bangladesh, PhD Dissertation, Cambridge University, Cambridge
Ahsan MF (2001) Socio-ecology of the hoolock gibbon (Hylobates hoolock) in two forests in Bangladesh. The apes: challenges for the 21st century. Conference proceedings. Chicago Zoological Society, Brookfield, Illinois, pp 286–299
Altmann J (1974) Observational study of behavior: sampling methods. Behaviour 49(3–4):227–266
Altmann J, Muruthi P (1988) Differences in daily life between semiprovisioned and wild-feeding baboons. Am J Primatol 15(3):213–221
Appanah S (1985) General flowering in the climax rain forests of South-east Asia. J Trop Ecol 1:225–240
Appanah S (1993) Mass flowering of dipterocarp forests in the aseasonal tropics. J Biosci 18:457–474
Ashton PS, Givnish TJ, Appanah S (1988) Staggered flowering in the Dipterocarpaceae: new insights into floral induction and the evolution of mast fruiting in the seasonal tropics. Am Nat 132:44–66
Bach TH, Chen J, Hoang MD, Beng KC, Nguyen VT (2017) Feeding behavior and activity budget of the southern yellow-cheeked crested gibbons (Nomascus gabriellae) in a lowland tropical forest. Am J Primatol 79:1–14
Bartlett TQ (1999) Feeding and ranging behavior of the white-handed gibbon (Hylobates lar) in Khao Yai National Park,Thailand. Ph.D. thesis, Washington University, St. Louis, MO
Bartlett TQ (2003) Intragroup and intergroup social interactions in white-handed gibbons. Int J Primatol 24(2):239–259
Bartlett TQ (2007) The Hylobatidae, small apes of Asia. In: Campell CJ, Fuentes A, Mackinnon KC, Panger M, Bearder SK (eds) Primates in perspective. Oxford University Press, New York, pp 274–289
Bartlett TQ (2008) The Gibbons of Khao Yai: seasonal variation in behavior and ecology. Pearson Prentice Hall, Upper Saddle River, NJ
Bartlett TQ (2009) Seasonal home range use and defendability in white-handed gibbons (Hylobates lar) in Khao Yai National Park, Thailand. In: Whittaker DJ (ed) Lappan S. The gibbons:, new perspective on small ape socioecology and population biology. Springer, pp 265–275
Berg ML, Beintema NH, Welbergen JA, Komdeur J (2005) Singing as a handicap: the effects of food availability and weather on song output in the Australian Reed Warbler Acrocephalus australis. J Avian Biol 36:102–109
Bricknell S (1999) Hybridisation and behavioural variation: a socio-ecological study of hybrid gibbons (Hylobates agilis albibarbis and H. muelleri) in Central Kalimantan, Indonesia, PhD Dissertation, Australian National University, Canberra
Carpenter CR (1940) A field study in Siam of the behaviour and social relations of the gibbon (Hylobates lar). Comp Psychol Monogr 16:1–212
Cheyne SM (2010) Behavioural ecology and socio-biology of gibbons (Hylobates albibarbis) in a degraded peat-swamp forest. In: Supriatna J, Gursky SL (eds) Indonesian Primates. Springer, New York, USA, pp 121–156
Cheyne SM, Höing A, Rinear J, Sheeran LK (2012) Sleeping site selection by agile gibbons: the influence of tree stability, fruit availability and predation risk. Folia Primatol 83(3–6):299–311
Chivers DJ (1974) The siamang in Malaya: a field study of primate in tropical rain forest. Contributions to Primatology, Karger, Basel 4:1–335
Chivers DJ (1984) Feeding and ranging in gibbons: a summary. In: Presuchoft H, Chivers DJ, Brockelman WY, Creel N (eds) The lesser Apes: evolutionary and behavioural biology. Edinburgh University Press, Edinburgh, pp 74–80
Clink DJ, Ahmad AH, Klinck H (2020) Gibbons aren’t singing in the rain: presence and amount of rainfall influences ape calling behavior in Sabah. Malaysia Sci Rep 10(1):1–13
Cowlishaw G (1996) Sexual selection and information content in Gibbon song bouts. Ethology 102(4): 272–284
Cunningham C, Mootnick A (2009) Gibbons. Curr Biol 19:R543–R544
Doran D (1997) Influence of seasonality on activity patterns, feeding behavior, ranging, and grouping patterns in Tai chimpanzees. Int J Primatol 18(2):183–206
Elder AA (2009) Hylobatid diets revisited: the importance of body mass, fruit availability, and interspecific competition. In: Whittaker DJ (ed) Lappan S. The gibbons:, new perspective on small ape socioecology and population biology. Springer, pp 133–159
Estrada A, Juan-Solano S, Martínez TO, Coates-Estrada R (1999) Feeding and general activity patterns of a howler monkey (Alouatta palliata) troop living in a forest fragment at Los Tuxtlas, Mexico. Am J Primatol 48(3):167–183
Fan PF, Ni QY, Sun GZ, Huang B, Jiang XL (2008) Seasonal variations in the activity budget of Nomascus concolor jingdongensis at Mt. Wuliang, Central Yunnan, China: effects of diet and temperature. Int J Primatol 29(4):1047
Fan PF, Fei HL, Ma CY (2012) Behavioral responses of Cao Vit gibbon (Nomascus nasutus) to variations in food abundance and temperature in Bangliang, Jingxi, China. Am J Primatol 74:632–641
Garamszegi LZ, Moreno J, Moller AP (2006) Avian song complexity is associated with high field metabolic rate. Evol Ecol Res 8:75–90
Gittins SP, Raemaekers JJ (1980) Siamang, lar and agile gibbons. In: Chivers DJ (ed) Malayan forest primates: ten years’ study in tropical rain forest. Plenum Press, New York, pp 63–105
Goller F, Larsen ON (1997) A new mechanism of sound generation in songbirds. Proc Natl Acad Sci USA 94:14787–14791
Groves CP (1972) Systematics and phylogeny of gibbons. In: Rumbaugh DM (ed) Gibbon and Siamang, vol 1. Karger, Basel, pp 1–89
Haimoff EH (1984) Acoustic and organizational features of gibbon songs. In: Chivers DJ, Brockelman WY, Creel N (eds) Preuschoft H. Edinburgh University Press, The lesser apes. Evolutionary and behavioural biology. Edinburgh, pp 333–353
Haimoff EH (1985) The organization of song in Mueller’s gibbon (Hylobates muelleri). Int J Primatol 6:173–192
Hanya G (2004) Seasonal variations in the activity budget of Japanese macaques in the coniferous forest of Yakushima: effects of food and temperature. Am J Primatol 63(3):165–177
Inoue Y, Sinun W, Yosida S, Okanoya K (2013) Intergroup and intragroup antiphonal songs in wild male Mueller’s gibbons (Hylobates muelleri). Interact Stud 14:24–43
Inoue Y, Sinun W, Okanoya K (2016) Activity budget, travel distance, sleeping time, height of activity and travel order of wild Mueller’s gibbons (Hylobates funereus) in Danum Valley Conservation Area. Raffles Bull Zool 64
Inoue Y, Sinun W, Yosida S, Okanoya K (2017) Combinatory rules and chunk structure in male Mueller’s gibbon songs. Interact Stud 18(1):1–25
Inoue Y, Sinun W, Yosida S, Okanoya K (2020) Note orders suggest phrase-inserting structure in male Mueller’s gibbon songs: a case study. Acta Ethol 23:89–102
Isbell LA, Young TP (1993) Social and ecological influences on activity budgets of vervet monkeys, and their implications for group living. Behav Ecol Sociobiol 32(6):377–385
Kagawa H, Suzuki K, Takahasi M, Okanoya K (2014) Domestication changes innate constraints for birdsong learning. Behav Processes 106:91–97
Kanamori T, Kuze N, Bernard H, Malim TP, Kohshima S (2010) Feeding ecology of Bornean orangutans (Pongo pygmaeus morio) in Danum Valley, Sabah, Malaysia: a 3-year record including two mast fruiting. Am J Primatol 72:820–840
Kanamori T, Kuze N, Bernard H, Malim TP, Kohshima S (2017) Fluctuations of population density in Bornean orangutans (Pongo pygmaeus morio) related to fruit availability in the Danum Valley, Sabah, Malaysia: a 10-year record including two mast fruitings and three other peak fruitings. Primates 58(1):225–235
Kershenbaum A, Garland EC (2015) Quantifying similarity in animal vocal sequences: which metric performs best? Methods Ecol Evol 6:1452–1461
Knott CD (1998) Changes in orangutan caloric intake, energy balance, and ketones in response to fluctuating fruit availability. Int J Primatol 19(6):1061–1079
Lambrechts MM (1996) Organization of bird song and constraints on performance. In: Kroodsma DE, Miller EH (eds) Ecology and evolution of acoustic communication in birds. Cornell University Press, Ithaca, New York, pp 305–320
Larsen ON, Goller F (2002) Direct observation of syringeal muscle function in songbirds and a parrot. J Exp Biol 205:25–35
Leighton DR (1987) Gibbons: territoriality and monogamy. In: Smuts BB, Cheney DL, Seyfarth RM, Wrangham RW, Struhsaker TT (eds) Primate societies. University of Chicago Press, Chicago, pp 135–145
Marshall J, Marshall F (1976) Gibbons and their territorial songs. Science 193:235–237
Marshall AJ, Boyko CM, Feilen KL, Boyko RH, Leighton M (2009) Defining fallback foods and assessing their importance in primate ecology and evolution. Am J Phys Anthropol 140(4):603–614
Matsuzawa T (2019) Bornean orangutans: primary forest in Danum Valley and rehabilitation program on Orangutan Island. Primates 60:477–483
Mitani JC (1985) Gibbon song duets and intergroup spacing. Behaviour 92:59–96
Okanoya K (2017) Sexual communication and domestication may give rise to the signal complexity necessary for the emergence of language: an indication from songbird studies. Psychon Bull Rev 24(1):106–110
Passamani M (1998) Activity budget of Geoffroy’s marmoset (Callithrix geoffroyi) in an Atlantic forest in southeastern Brazil. Am J Primatol 46(4):333–340
Raemaekers J (1978) Changes through the day in the food choice of wild gibbons. Folia Primatol 30(3):194–205
Raemaekers J (1980) Causes of variation between months in the distance traveled daily by gibbons. Folia Primatol 34:46–60
Reichard U (1998) Sleeping sites, sleeping places, and presleep behavior of gibbons (Hylobates lar). Am J Primatol 46:35–62
Ritschard M, Brumm H (2012) Zebra finch song reflects current food availability. Evol Ecol 26:801–812
Robinson JG (1986) Seasonal variation in use of time and space by the wedge-capped capuchin monkey, Cebus olivaceus: implications for foraging theory. Smithsonian Contrib Zool 431:1–60
Suthers RA, Goller F, Pytte C (1999) The neuromuscular control of birdsong. Philos Trans R Soc Lond B Biol Sci 354:927–939
Takahasi M, Okanoya K (2010) Song learning in wild and domesticated strains of white-rumped munia, Lonchura striata, compared by cross-fostering procedures: domestication increases song variability by decreasing strain-specific bias. Ethology 116:396–405
Uhde NL, Sommer V (2002) Anti-predatory behaviour in gibbons. In: Miller LE (ed) Eat or be eaten: predator sensitive foraging among primates. Cambridge University Press, Cambridge, pp 268–292
Van der Loo MP (2014) The stringdist package for approximate string matching. R J 6(1):111–122
Vasey N (2005) Activity budgets and activity rhythms in red ruffed lemurs (Varecia rubra) on the Masoala Peninsula, Madagascar: seasonality and reproductive energetics. Am J Primatol 66(1):23–44
Wong ST, Servheen C, Ambu L, Norhayati A (2005) Impacts of fruit production cycles on Malayan sun bears and bearded pigs in lowland tropical forest of Sabah, Malaysian Borneo. J Trop Ecol 21:627–639
Zanoli A, De Gregorio C, Valente D, Torti V, Bonadonna G, Randrianarison RM, Giacoma C, Gamba M (2020) Sexually dimorphic phrase organization in the song of the indris (Indri indri). Am J Primatol 82(6):e23132
Acknowledgements
We thank the Economic Planning Unit of the Malaysia Federal Government, the Economic Planning Unit of Sabah State Government, Danum Valley Management Committee, and Sabah Biodiversity Centre for permission to conduct this study in Sabah. We are grateful to Etsuko Inoue, Shigeto Yosida, and Mike Bernadus for their dedicated help with our research. We also thank Donny Sah Itin, Isnadil Mohd, and Dennysius Aloysius for assistance with gibbon tracking and data collection.
Funding
This study was partly supported by Heisei Foundation for Basic Science.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Inoue, Y., Sinun, W. & Okanoya, K. Increase in social interactions of wild Northern Gray gibbons (Hylobates funereus) during the mast fruiting period in the Danum Valley Conservation Area, Sabah, Malaysia. acta ethol 24, 153–163 (2021). https://doi.org/10.1007/s10211-021-00370-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10211-021-00370-1