
Abstract
Analyses of acoustic variation between and within populations can help to clarify the evolution and diversification of cicada calling songs. In this study, we analyzed the acoustic variation in the calling song of Mogannia formosana within Taiwan and between Taiwan and Green Island to assess the effects of geographic locations. Furthermore, chorusing males in the Green Island population were recorded and collected from the same habitat site during the same time period to investigate the relationship between individual body size and the acoustic features of calling songs. Among populations of M. formosana, we found that most of the acoustic variation in M. formosana calling songs was associated with frequency parameters, in which six frequency parameters changed significantly with latitude on the island of Taiwan. In contrast, temporal parameters, which were associated with principal components corresponding to less acoustic variation than that of the first principal component, were also found to be significant among populations but did not show consistent trends of difference with latitude. However, the geographically isolated Green Island population exhibited the lowest number of short echemes in segment B, which is the diagnostic structure employed to separate M. formosana from other M. species. This finding suggests that the temporal pattern of segment B in the calling songs of M. formosana might be important for both population differentiation and interspecific recognition. In a chorus of the Green Island population, the sound frequency of the last short echeme was found to be significantly correlated with individual body size. The possible role played by sexual selection in shaping sound frequency as a result of its reliable association with body size was discussed. We suggest that, in comparison with temporal elements, the frequency elements of segment B in calling songs of M. formosana in chorus can serve as a more reliable indicator of body size for female mate choice.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Male cicadas produce loud calling songs to attract females (Claridge 1985). The calling songs of cicadas are well known for their functional role in mate attraction (Claridge 1985; Marshall and Cooley 2000). Every species has a distinct calling song for species recognition, whether the species are closely related or are unrelated sympatric species (Villet 1989; Marshall and Cooley 2000; Sanborn and Phillips 2001; Sueur 2002). The calling songs of cicadas can thus be used to resolve taxonomic problems. This includes the identification of sibling or cryptic species that do not differ in morphology (e.g., Sueur and Puissant 2007). Nevertheless, the calling songs of male cicadas also show considerable intra-specific variation, as documented by several studies (e.g., Pinto-Juma et al. 2005, 2008; Quartau and Simões 2006; Popple et al. 2008). Intra-specific variation consists of both variation between populations and variation within a population. One of the most important sources of variation between populations is geographic variation, which is most informative for understanding the ecological factors that shape the evolution and diversification of communication systems (Wilczynski and Ryan 1999). Therefore, the comparisons of populations that display geographic variation in calling songs can offer substantial insight into the mechanisms of adaptive divergence and speciation (Foster 1999).
Regarding the variation within a population, individual body size has been suggested to be the most informative trait associated with variation in the acoustic structures of animals as a result of sexual selection (Gerhardt and Huber 2002). In particular, the sound frequency characteristics of male anurans have been found to be negatively correlated with body size and thus serve as reliable cues for females to assess a mate’s quality/body size or for males to assess an opponent’s fighting ability/body size (Gerhardt 1994; Wang et al. 2012). In the cicadas Magicicada cassini and Magicicada sependecim, males in copula have been found to be larger than males from the same population that are not in copula (Karban 1983; English 2001); that is, female cicadas prefer to mate with larger-sized males in a population. Because the size of the sound-producing system determines the song frequency and is closely related to overall body size (Bennet-Clark and Young 1994), the song frequency could be selected as a reliable indicator of body size through female choice in cicadas. Thus, within a cicada population, we would expect a negative relationship between individual body size and song frequency as a result of sexual selection. However, in addition to social sources of selection on body size, such as female choice, that may directly select the song frequency as a result of its reliable association with body size, ecological sources, such as habitats and climate temperatures, can also influence the evolution of communication systems through correlated effects on the song frequency (Cocroft and De Luca 2006). Therefore, without controlling ecological factors by investigating the signaling of male cicadas at the same habitat site during the same time period, the relationship between individual body size and song frequency as a result of sexual selection may not be revealed. Few studies aimed at investigating the relationship between individual body size and song frequency-controlling ecological factors within a cicada population have been conducted.
Four cicada species of the genus Mogannia have been identified in Taiwan, and the males of each species produce species-specific calling songs (Lin 2005). Among the four species, M. formosana Matsumura is the only endemic species that inhabits both Taiwan and Green Island, which is located 33 km from the southeastern coast of Taiwan. The other three species exhibit wide distributions throughout mainland China, Okinawa, and Taiwan (Lin 2005). In this study, we analyzed the variation in the calling song structure of M. formosana within Taiwan and between Taiwan and Green Island to assess the effect of geography. Furthermore, chorusing males from the Green Island population were recorded and collected at the same habitat site during the same time period to investigate the relationship between individual body size and song frequency. Studies on the effects of individual body size and geography on the acoustic variation within a cicada species are scarce, and this study will shed light on the diversification and evolution of acoustic communication between populations as well as within a population of cicadas.
Materials and methods
Song recording
We recorded M. formosana calling songs in the field between 2009 and 2015 in three regions of Taiwan (northern region: E121° 34′–121° 38′, N25° 10′–25° 18′, elevation <300 m; central region: E 120° 40′–120° 42′, N 22° 57′–22° 59′-, elevation 600–700 m; and southern region: E 120° 50′–120° 51′, N 21° 54′–21° 56′, elevation <200 m) and on Green Island (E 121° 28′–121° 30′, N 22° 38′–22° 41′). Each sampled region was at least 80 km away from the other sampled regions, and each cicada was recorded until it stopped singing. The air temperature was noted during each recording session, with a temperature range of 31–34 °C being observed in the northern region, 27–30 °C in the central region, 33–36 °C in the southern region, and 25–29 °C on Green Island. The calling songs were recorded using a Denon portable IC recorder (DN-F20R) connected to a Sennheiser ME67 unidirectional microphone (mounted with foam windscreen) with a 48 kHz sampling frequency. The sound recordings were analyzed using Avisoft-SASLab Pro software (version 4.40) at a resolution of 16 bits. Because the calling songs of M. formosana were observed to occur at above 5 kHz in the spectrograms, a high-pass filter was used to remove noise components below 5 kHz from the recordings. According to the spectrograms of the calling songs, four orderly segments were identified: (A) clicks, (B) series of short echemes, (C) one long echeme, and (D) clicks (Fig. 1). Because the number of clicks in segments A and D were highly variable, only segments B and C were used for further analyses. We produced spectrograms with the following settings: FFT = 512, frame size = 100%, hamming window, frequency resolution = 94 Hz, overlap = 87.5%, and time resolution = 1.33 ms. We measured 13 variables (Table 1) on the spectrograms using the Automatic Parameter Measurements function in Avisoft-SASLab Pro.

Spectrograms and corresponding oscillograms showing (a) the four segments of the M. formosana calling song and (b) the last short echeme in segment B. A: clicks; B: a series of short echemes; C: a long echeme; and D: clicks
Individual body size measurements and near distance recording experiment
A total of 45 M. formosana cicadas were caught on Green Island and brought back to the laboratory in April of 2015. The body length from the tip of head to tip of the abdomen was measured for each individual to the nearest 0.01 mm using a digital caliper (CT200-197). Males exhibited similar body lengths as females (male: mean ± SE = 16.67 ± 0.17 mm, n = 32; female: 16.12 ± 0.29 mm, n = 13; t = 1.62, P > 0.05) in the Green Island population. Because of the need to record the calling songs of males individually while they were aggregated in chorusing groups on small bushes, the microphone was placed less than 10 cm from the target male to reduce the masking effect of neighbors’ singing. Eighteen males in chorusing groups within a distance of 10 m were recorded during a 2-h period with a temperature range of 25–26 °C on April 26 of 2015 on Green Island. After recording, the males were immediately caught and brought back to the laboratory for body measurements; however, only 11 males among the 18 recorded individuals were successfully caught, and 10 of the 11 caught individuals showed good-quality recordings for the acoustic analysis. For body size measurements of those 10 calling males, wing-spread specimens of them were photographed in a dorsal aspect by a digital camera (Nikon Coolpix P900). The body length (from the tip of head to tip of the abdomen) and the pronotum width (maximum width of pronotal collar) were measured from the images of the specimens in pixel units using the software Adobe Illustrator CS2 12.0.0. All measurements in pixel units were converted to data in millimeters (mm) according to the ratio of pixel distance to actual distance.
To investigate the possible effect of near distance recording on frequency measurements, we generated 5-s pure tones of 3, 4, 5, and 9 kHz using Audacity software (sine waveform, amplitude = 0.8, rate = 48 kHz). In the laboratory, those source sounds (pure tones) were broadcast 10 times each through an Acer PC multimedia speaker (LS-21 T3A233) at distances of 3, 6, 10, and 30 cm from the tip of the shotgun microphone (Sennheiser ME67 with foam windscreen and a total length of 37 cm), recorded with the same equipment and settings as in the field, and analyzed using Avisoft-SASLab Pro software at a resolution of 16 bits. Both spectrographic cross-correlation (SPCC) and parametric methods were used to compare the source and recorded sounds. The spectrographic cross-correlation (SPCC) coefficients were computed using Avisoft-CORRELATOR (FFT = 512, frequency tolerance deviation = 0); a SPCC coefficient of 1 indicates that the two spectrograms are identical (Specht 2002). The mean peak frequency of an entire sound was measured on the spectrogram using the Automatic Parameter Measurements function in Avisoft-SASLab Pro. SPCC coefficients between the source and recorded sounds were greater than 0.95 for all pure tones at a recording distance less than 10 cm (Online Resource 1: Appendix 1). Furthermore, the mean peak frequency measurements were all similar regardless of the recording distance for each source pure tone; i.e., the mean peak frequency was 3000 Hz for the 3 kHz source pure tone at all recording distances, 4030 Hz for the 4 kHz source pure tone, 4960 Hz for the 5 kHz source pure tone, and 9000 Hz for the 9 kHz source pure tone (Online Resource 1: Appendix 2). No frequency distortion was observed in recorded sounds as close as 3 cm with our recording equipment.
Statistical analyses
We used individuals as our sample units and chose individuals with good-quality recordings for acoustic measurements (202 calling songs of 83 individuals). Two approaches were employed to assess song variation in a geographic context. The first approach examined the geographic patterns of the calling songs through principal component analysis (PCA) of the averaged measurements from the songs of each male cicada. PCA was performed on 13 variables, applying a normalized transformation to each variable to obtain a zero mean and unit standard deviation. We retained all components with eigenvalues greater than one and interpreted each component based on its correlations with the original variables (coefficients/eigenvectors). The second approach used the variables with absolute values of coefficients greater than 0.3 in PC1, 0.5 in PC2, and 0.6 in PC3 and PC4 to analyze population differences. We assessed population differences by performing Kruskal-Wallis tests (two-tailed, with a significance level of 0.05) and then compared pairs of populations using Mann-Whitney tests (two-tailed, with a significance level of 0.01 to adjust for multiple comparisons). The relationships between body size and the acoustic variables were analyzed through Spearman’s rank correlation (two-tailed, with a significance level of 0.05). The PCAs were performed using PRIMER 6.0, and nonparametric tests were performed using SYSTAT 11.
Results
Geographic variation in calling song structures
Four principal components were derived from the correlation matrix of 13 acoustic variables through PCA (Table 2). The first two components (PC1 and PC2) accounted for 58.9% of the total variation. When PC1 was plotted against PC2, the songs of individuals from Green Island were separated from the songs of individuals from southern Taiwan but overlapped with the songs of individuals from central and northern Taiwan (Fig. 2). Furthermore, along PC1, there was an increasing trend from the southern songs to the northern songs (Fig. 2).

A bidimensional diagram of the principal component analysis using the 13 acoustic variables from the spectrograms of M. formosana calling songs. N: northern region; C: central region; S: southern region; and G: Green Island
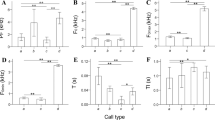
PC1, which accounted for 45.1% of the total variation, was highly correlated with frequency variables such as BFPFMEAN, BLPFMEAN, BLPFMIN, CPFMEAN, CPFMIN, and CPFMAX (Table 2). There were significant differences in these variables among the four populations (Kruskal-Wallis test, n = 83, df = 3, P < 0.01; BFPFMEAN: H = 47.34; BLPFMEAN: H = 60.1; BLPFMIN: H = 44.23; CPFMEAN: H = 54.68; CPFMIN: H = 41.17; CPFMAX: H = 44.25). Additionally, within Taiwan, these frequency variables showed an increasing trend from north to south, and the frequency variables of songs from southern Taiwan showed significantly higher values compared with the other populations (Mann-Whitney tests, P < 0.01, Fig. 3).

Box-plots of the acoustic variables with absolute values of coefficients (eigenvectors) greater than 0.3 for PC1. Different letters indicate significant differences (Mann-Whitney, p < 0.01) between two populations. Circles and asterisks represent extreme values. Descriptions of the variables are provided in Table 1
PC2 accounted for 13.8% of the total variation, with a higher value of PC2 corresponding to a longer duration of the first and last echemes in segment B (BFDUR and BLDUR) (Table 2). PC3 and PC4 accounted for 9.4 and 8.8% of the total variation, respectively (Table 2). PC3 was highly correlated with the number of echemes in segment B (BAMO), while PC4 was highly correlated with the duration of segment C (CDUR). There were significant differences observed among the four populations for the duration variables (Kruskal-Wallis test, n = 83, df = 3, P < 0.01; BFDUR: H = 26.93; BLDUR: H = 29.17; CDUR: H = 15.31); however, no common trend was detected (Fig. 4). The number of echemes in segment B (BAMO) was significantly different among populations (Kruskal-Wallis test, n = 83, df = 3, P < 0.01, H = 48.13) and was lowest in the Green Island population (Fig. 4).

Box-plots of the acoustic variables with absolute values of coefficients (eigenvector) greater than 0.5 for PC2 (BFDUR, BLDUR), 0.6 for PC3 (BAMO), and PC4 (CDUR). Different letters indicate significant differences (Mann-Whitney, p < 0.01) between two populations. Circles and asterisks represent extreme values. Descriptions of the variables are provided in Table 1
Body size and acoustic structures in chorus
In chorus, male body sizes did not significantly correlate with the duration variables of segment B or C in calling songs (n = 10,∣rs∣ < 0.648, P > 0.05 for BAMO, BFDUR, BLDUR, CDUR). However, for the frequency variables examined, the mean peak frequency of the last echeme in segment B (BLPFMEAN) showed significantly negative relationships with pronotum width (r s = −0.732, P < 0.05) and body length (r s = −0.677, P < 0.05). Larger males exhibited lower frequency of the last echeme in segment B in calling song (Fig. 5).

Plots of the mean peak frequency of the last echeme in segment B (BLPFMEAN) against (a) body length and (b) pronotum width in M. formosana from Green Island
Discussion
Acoustic variation between populations
In this study, we observed significant acoustic variation in the calling songs of M. formosana among four populations, and most of the acoustic variation was associated with frequency parameters. In particular, we found that six frequency parameters of the M. formosana calling songs were not only significantly different among populations but also decreased significantly from the southern population to the northern population of Taiwan. A significant decrease in frequency parameters with latitude was also reported in Cicada barbara Stål (Pinto-Juma et al. 2008). Latitudinal clines, which are one form of geographic variation, have received attention in investigations of climate change (Gardner et al. 2011; Sheridan and Bickford 2011). The latitudinal clines in the acoustic frequency of cicada songs could be due to the direct or indirect effects of environmental factors such as temperature (Wilczynski and Ryan 1999). Ambient temperature has been shown to directly influence the temporal parameters but not the frequency parameters of cicada songs (Quartau et al. 2000; Fonseca and Revez 2002a; Sanborn et al. 2011). Therefore, if temperature plays an important role in shaping the latitudinal clines in the acoustic frequency of M. formosana, it most likely results from indirect effects acting on morphological traits such as body size, which has been found to be negatively correlated with the dominant song frequency in cicadas (Bennet-Clark and Young 1994). We would then predict that the body size of male M. formosana would increase from southern populations to northern populations with the decreasing annual mean temperature from southern to northern Taiwan. A further study of the collections of male cicadas from populations in each region is needed to verify this prediction.
The songs of Green Island cicadas tended to group separately from those of southern Taiwan cicadas but overlapped with those of the cicadas from central and northern Taiwan. In fact, the songs of males from Green Island showed a similar frequency range to those of the northern population, but not to those of the southern population, though Green Island is latitudinally farther from northern Taiwan than it is from southern Taiwan. This result is not concordant with our expectations based on the latitudinal clines of frequency parameters observed on the island of Taiwan. Geographic isolation caused by the oceanic barrier could be a major factor accounting for the discordant findings in the Green Island population.
For temporal parameters associated with the second, third, and fourth principal components (PC2–4), although significant differences were found among populations in the durations of the first short echeme (BFDUR), the last short echeme (BLDUR), the long echeme (CDUR), and the number of short echemes (BAMO), no consistent trends of difference from north to south were found. However, the geographically isolated Green Island population had a significantly smaller number of short echemes (BAMO) and longer durations of the last short echeme (BLDUR) in segment B in calling songs than those of populations on the main island of Taiwan. Segment B, which consists of short echemes, is used as the diagnostic acoustic feature of M. formosana, separating the calling songs of this species from the calling songs of the other three M. species found in Taiwan (Lin 2005). Based on those findings, we therefore suggest that the temporal pattern of segment B in the calling songs of M. formosana is important not only for population differentiation but also for interspecific recognition; the evolution of short echemes might have played a role in the speciation of M. species, particularly for the endemic species M. formosana. The significance of temporal patterns of calling songs in species recognition has also been found for C. barbara, in which males discriminate conspecific songs from heterospecific songs by the temporal pattern of the songs and use the temporal song structure for long-range communication (Fonseca and Revez 2002b).
Individual body size and acoustic variation in chorus
We found that in chorus, larger males exhibited significantly lower frequency features of the last short echeme in segment B of calling songs in the Green Island population. In cicadas, although studies involving interspecific comparisons have revealed a negative relationship between body size and the dominant song frequency (Bennet-Clark and Young 1994; Oberdörster and Grant 2007), few studies have found significant relationships between sound frequency and the individual body size of male cicadas within a population in the field (Mehdipour et al. 2015). Moreover, a positive correlation between individual body weight and the dominant song frequency was found in the cicada Chloropsalta smaragdula (Mohammadi-Mobarakeh et al. 2014), contradicting what might be expected according to interspecific comparative studies. The lack of evidence concerning negative correlations between individual body size and frequency features within a cicada population in the field might be attributed to two factors. First, the song samples analyzed in previous studies might have been recorded from different chorusing groups in various ecological contexts, including diverse habitats and different time periods. Thus, without recording male cicadas in the same chorusing group to ensure that the same ecological factors are at play; the frequency features of calling songs can be confounded with factors other than individual body size and the relationship between individual body size and frequency features cannot be revealed. Second, because of the first issue, male calling songs must be individually recorded within the same chorusing group during the same time period to ensure that they are subject to the same ecological context, which might result in sampling difficulties in the field. In chorus, few cicada populations allow researchers to place a microphone sufficiently close to them to avoid the masking effect of neighbors’ singing to obtain good-quality recordings of individual cicadas for acoustic analysis. It is even more difficult for researchers to capture individual males immediately after sound recording to perform body size measurements. Our finding that individual body size was significantly correlated with the sound frequency of the last short echeme of the calling songs of M. formosana therefore provides valuable field evidence to support that song frequency can be a reliable indicator of male body size in cicadas.
However, being a reliable indicator of male body size does not ensure the possible role song frequency may play in sexual selection, specifically, female mate choice. Previous studies provide evidence to support the significant roles played by sexual selection in shaping the sound frequency as a result of its reliable association with body size. First, female cicadas do prefer to mate with larger-sized males in a population (Karban 1983; English 2001). Second, female cicadas are well tuned and capable of discriminating fine frequency differences and have many receptors involved in frequency discrimination as do vertebrates (Fonseca et al. 2000). Furthermore, female cicadas have good directional hearing at the main frequencies of the calling song of its own species (Fonseca and Popov 1997). Therefore, with good frequency resolution maintained in the CNS and good directional hearing ability in the tympanum, it is reasonable to conclude that female cicadas can discriminate fine frequency differences of calling songs in a chorus without difficulty. Third, compared to temporal elements, song frequency is less temperature dependent (Quartau et al. 2000; Fonseca and Revez 2002a; Sanborn et al. 2011) and thus more reliable as a criterion for female mate choice.
In this study, we found that frequency features of the last short echeme of segment B but not the other parts of calling songs correlated significantly with male body size. We speculated that the production of the last short echeme might rely primarily on the tymbal plate; therefore, frequency features of the last short echeme are mass-sensitive and less affected by temperature (Bennet-Clark 1997), which resulted in a significant relationship between the frequency features of the last short echeme and body size. As our results of population comparisons on acoustic variation suggest, segment B (a series of short echemes) in calling songs of M. formosana is important for species recognition and population differentiation. If female M. formosana do prefer larger males and use acoustic cues as criteria for mate choice, temperature-independent features such as frequency elements in the species-specific component, segment B, would be more reliable than the temperature-dependent features (temporal elements), even though temperature increase might also lead to an increase in sensitivity of the auditory response (Fonseca and Correia 2007). As to other components of the calling songs in M. formosana such as segment C (a long echeme), we found no significant relationship between frequency features and individual body size. In addition to the problem of small sample size, temperature-dependent production of segment C might account for the lack of a negative relationship between frequency features of segment C and individual body size. That is, the production of segment C in M. formosana might rely primarily on the tensor muscle (Fonseca and Hennig 1996) or abdomen cavity (Bennet-Clark and Young 1992), and frequency features of segment C are thus easily affected by changing individual body postures, which not only affect the volume of the abdomen resonator but also increase exposure to sunlight and elevate body temperatures (Fonseca and Revez 2002a). Nevertheless, a further experiment on the female preference of the frequency of various components of calling songs in M. formosana is needed to verify the roles played by sexual selection in shaping the song frequency as a result of its reliable association with body size, which has been suggested in vertebrates (Morton 1977; Gerhardt and Huber 2002), but not in cicadas until now.
Between populations on the island of Taiwan, we found that most of the acoustic variation in M. formosana calling was associated with frequency parameters, in which six frequency parameters of songs decreased significantly from the southern population to the northern population of Taiwan. In contrast, temporal parameters, which were highly associated with the second, third, and fourth principal components (PC2–4) but not PC1, showed no consistent trends from north to south. However, the temporal pattern of segment B in the calling songs of M. formosana is important not only for population differentiation but also for interspecific recognition, given that the geographically isolated Green Island population exhibiting the lowest number of short echemes in segment B, which is the diagnostic structures employed to separate M. formosana from other M. species. Furthermore, in a Green Island population chorus, the sound frequency of the last short echeme in segment B was found to be significantly correlated with individual body size, and we suggest that, in comparison with temporal elements, frequency elements can serve as a reliable indicator of body size for female mate choice.
References
Bennet-Clark HC (1997) Tymbal mechanics and the control of song frequency in the cicada Cyclochila australasiae. J Exp Biol 200:1681–1694
Bennet-Clark HC, Young D (1992) A model of the mechanism of sound production in cicadas. J Exp Biol 173:123–153
Bennet-Clark HC, Young D (1994) The scaling of song frequency in cicadas. J Exp Biol 191:291–294
Claridge MF (1985) Acoustic signals in the Homoptera: behavior, taxonomy, and evolution. Annu Rev Entomol 30:297–317
Cocroft RB, De Luca PA (2006) Size-frequency relationships in insect vibrational signals. In: Drosopoulos S, Claridge MF (eds) Insect sounds and communication, physiology, behaviour, ecology and evolution. CRC Press Taylor & Francis Group, Boca Raton, pp 109–120
English JE (2001) Sexual selection in the lek-mating system of periodical cicadas Magicicada cassini and M. decim; selection for male body size, dominant courtship call and pathogen avoidance. Univ. Arkansas, Fayetteville
Fonseca PJ, Correia T (2007) Effects of temperature on tuning of the auditory pathway in the cicada Tettigetta josei (Hemiptera, Tibicinidae). J Exp Biol 210:1834–1845
Fonseca PJ, Hennig R (1996) Phasic action of the tensor muscle modulates the calling song in cicadas. J Exp Biol 199:1535–1544
Fonseca PJ, Popov AV (1997) Directionality of the tympanal vibrations in a cicada: a biophysical analysis. J Comp Physiol A180:417–427
Fonseca PJ, Revez MA (2002a) Temperature dependence of cicada songs (Homoptera, Cicadoidea). J Comp Physiol A 187:971–976
Fonseca PJ, Revez MA (2002b) Song discrimination by male cicadas Cicada barbara lusitanica (Homoptera, Cicadidae). J Exp Biol 205:1285–1292
Fonseca PJ, Munch D, Hennig RM (2000) How cicadas interpret acoustic signals. Nature 405:297–298
Foster SA (1999) The geography of behavior: an evolutionary perspective. Trends Ecol Evol 14:190–195
Gardner JL, Peters A, Kearney MR, Joseph L, Heinsohn R (2011) Declining body size: a third universal response to warming? Trends Ecol Evol 26:285–291
Gerhardt HC (1994) The evolution of vocalization in frogs and toads. Annu Rev Ecol Syst 25:293–324
Gerhardt HC, Huber F (2002) Acoustic communication in insects and anurans: common problems and diverse solutions. The University of Chicago Press, Chicago
Karban R (1983) Sexual selection, body size and sex-related mortality in the cicada Magicicada cassini. Am Midl Nat 109:324–330
Lin Y-D (2005) Taxonomy of the genus Mogannia Amyot & Serville (Hemiptera: Cicadidae) of Taiwan. Master thesis. National Taiwan University, Taipei
Marshall DC, Cooley JR (2000) Reproductive character displacement and speciation in periodical cicadas, with description of a new species, 13-year Magicicada neotredecim. Evolution 54:1313–1325
Mehdipour M, Sendi JJ, Zamanian H (2015) External morphology and calling song characteristics in Tibicen plebejus (Hemiptera: Cicadidae). C R Biol 338:103–111
Mohammadi-Mobarakeh A, Hatami B, Zand AJ, Fattahi RA, Zamanian H (2014) Dominant frequency characteristics of calling songs in cicada Chloropsalta smaragdula Haupt, 1920 (Hem: Cicadidae) and its relationship with body length, width, and weight. J Entomol Zool Stud 2:330–332
Morton ES (1977) On the occurrence and significance of motivation—structural rules in some bird and mammal species. Am Nat 11:855–869
Oberdörster U, Grant PR (2007) Acoustic adaptations of periodical cicadas (Hemiptera: Magicicada). Biol J Linn Soc 90:15–24
Pinto-Juma GA, Simões PC, Seabra SG, Quartau JA (2005) Calling song structure and geographical variation in Cicada orni L. (Hemiptera: Cicadidae). Zool Stud 44:81–94
Pinto-Juma GA, Seabra SG, Quartau JA (2008) Patterns of acoustic variation in Cicada barbara Stål (Hemiptera, Cicadoidea) from the Iberian Peninsula and Morocco. Bull Entoml Res 98:1–14
Popple LW, Walter GH, Raghu S (2008) The structure of calling songs in the cicada taxon Pauropsalta annulata Goding and Froggatt (Hemiptera: Cicadidae)—evidence of diverging populations? Evol Ecol 22:203–215
Quartau JA, Simões PC (2006) Acoustic evolutionary divergence in cicadas: the species of Cicada L. in southern Europe. In: Drosopoulos S, Claridge MF (eds) Insect sounds and communication, physiology, behaviour, ecology and evolution. CRC Press Taylor & Francis Group, Boca Raton, pp 227–237
Quartau JA, Seabra S, Sanborn A (2000) Effect of ambient air temperature on some temporal parameters of the calling song of Cicada orni Linnaeus, 1758 (Hemiptera: Cicadidae) in Portugal. Acta Zool Craco 43:193–198
Sanborn AF, Phillips PK (2001) Re-evaluation of the Diceroprocta delicata (Homoptera: Cicadidae) species complex. Ann Entomol Soc Am 94:159–165
Sanborn AF, Simöes PC, Phillips PK, Quartau JA (2011) Thermoregulation and the influence of body temperature on calling song parameters in Cicada orni (Hemiptera: Cicadidae). Eur J Entomol 108:365–369
Sheridan JA, Bickford D (2011) Shrinking body size as an ecological response to climate change. Nat Clim Chang 1:401–406
Specht R (2002) Avisoft-SASLab Pro sound analysis and synthesis laboratory. Avisoft Bioacoustics, Berlin
Sueur J (2002) Cicada acoustic communication: potential sound partitioning in a multispecies community from Mexico (Hemiptera: Cicadomorpha: Cicadidae). Biol J Linn Soc 75:379–394
Sueur J, Puissant S (2007) Similar look but different song: a new Cicadetta species in the montana complex (Insecta, Hemiptera, Cicadidae). Zootaxa 1442:55–68
Villet MH (1989) Systemic status of Platypleura stridula L. and Platypleura capensis L. (Homoptera: Cicadidae). S Afr J Zool 24:329–332
Wang JC, Cui JG, Shi HT, Brauth SE, Tang YZ (2012) Effects of body size and environmental factors on the acoustic structure and temporal rhythm of calls in Rhacophorus dennysi. Asian Herpetol Res 3:205–212
Wilczynski W, Ryan MJ (1999) Geographic variation in animal communication systems. In: Foster SA, Endler JA (eds) Geographic variation in behavior. Oxford University Press, New York, pp 234–261
Acknowledgments
We thank the administration of the Shanping Forest Ecological Garden of the Taiwan Forestry Research Institute for their support during the study. Hsiang-Hao Loa and Szu-Ying Lin are acknowledged for their help with the fieldwork. Dr. Hong-Young Yan is acknowledged for his suggestion on conducting the near distance recording experiment. We thank Matija Gogala for his valuable comments and suggestions on previous drafts of the manuscript. This work was supported by the Ministry of Science and Technology, Taiwan (MOST 105-2311-B-037-002, NSC 98-2313-B-037-002-MY3).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Shieh, BS., Liang, SH., Liao, CY. et al. Song frequency correlates with latitude and individual body size in the cicada Mogannia formosana Matsumura (Hemiptera: Cicadidae). acta ethol 20, 147–155 (2017). https://doi.org/10.1007/s10211-017-0258-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10211-017-0258-3