
Abstract
Illumination at night is determined by moon phase, naturally affecting the reproductive activities of nocturnal animals. The European badger (Meles meles Linnaeus, 1758) is a large-sized mustelid that lives throughout Bulgaria. Since information on its reproductive activity is scarce for the country, a two-year camera trap survey was conducted at five main setts. It aimed to establish the mating period of the badger, as well as some behavioral activities of the observed individuals in a semi-mountainous area in Central Bulgaria (Sarnena Sredna gora Mts). In the present study, badger mating behavior was only observed in winter. Winter is the main breeding period for badgers from Central Bulgaria, which corresponds to greater activity in front of the setts. Darker lunar phases (i.e., new moon) significantly increased the badger's reproductive activities at their setts. Further research is needed to elucidate the underlying mechanism responsible for the nocturnality and preferences to darkness of reproductive badgers.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The balance between activity and rest is an important ethoecological feature for any species related to minimizing energy costs. However, the activity patterns of medium sized mammals are difficult to determine in natural conditions in a cost-effective and non-invasive manner (Rowcliffe et al. 2014). This is the reason for the growing use of camera traps in ecological studies on nocturnal mammals, specifically on mustelid species (Griffiths 2008; Yoxon 2008).
The European badger (Meles meles Linnaeus, 1758) is a large-sized mustelid with wide distribution in Europe (Prigioni 1999). In Bulgaria, the species is distributed across the whole country (Popov and Sedefchev 2003).
The badger is well known as a nocturnal animal (Harris 1982; Neal and Cheeseman 1996; Zabala et al. 2002; Kowalczyk et al. 2003; Rotenko et al. 2012; Magowan et al. 2022). Its behavioral patterns (daily and seasonal) are influenced by climatic conditions (Cresswell and Harris 1988; Slamka 2016), day length, light intensity at night (i.e. moon phase; Dixon et al. 2006), local food resources and some social factors (Neal and Cheeseman 1996; Buesching et al. 2003; Carpenter et al. 2005; Silk et al. 2017). The reproductive biology of the badger is still poorly understood because it is complex (Roper 2010). While mating behavior can happen year round, the primary period of reproductive behavior is between January and March (Mori et al. 2015). Courtship and copulation usually take place in front of the main sett (Roper 2010) preceded by increased scent-marking activity (Kruuk et al. 1984; Buesching and Macdonald 2001) and allo-grooming (Kruuk 1989; Buesching et al. 2003).
The influence of moon illumination on mammalian behavior is of scientific interest. This factor influences the activity of predators towards their prey (Cozzi et al. 2012; Penetriani et al. 2013) while for non-obligate carnivores like brown bear (Ursus arctos) this does not appear to be the case (Lamamy et al. 2022). Avoiding large predators, the European badger synchronizes its mating bevaviors with the lunar cycle, increasing its activities in low illumination conditions, i.e., around new moon phase (Cresswell and Harris 1988; Dixon et al. 2006). Therefore, the badgers’ reproductive activities could fluctuate with both short (monthly) and long (annual) term biological rhythms.
The present study aims to determine the European badger mating period and the associated behavioral activities in the conditions of a semi-mountainous area in Central Bulgaria (Sarnena Sredna gora Mts). In Bulgaria, information on the reproductive activity of the badger is scarce (Popov and Sedefchev 2003; Racheva et al. 2012). To address this information gap, we investigated free-ranging badger activities at the setts, using camera traps. First, we determined the seasonality in reproductive behaviors observed in front of the main sett, by annual camera-trapping. Second, we compared diel activity patterns between reproductive and non-reproductive seasons, to assess seasonal changes in the nocturnality. Third, we compared reproductive behavior and diel activities in different moon phases, to elucidate the effects of the lunar-cycle on mating and nocturnality in European badgers.
Materials and methods
Study area and camera trap survey
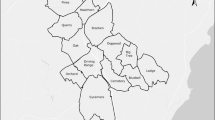
Remote recording devices are suitable for investigating behavioral activity patterns of elusive nocturnal and fossorial species such as badgers (Stewart et al. 1997). Five camera traps were placed in front of five different breeding setts, located at semi-mountainous ranges (217–503 m a.s.l.) in an agro forestry landscape, District of Stara Zagora, Central Bulgaria (42°28ʹN, 25°37ʹE; Fig. 1). Field observations were carried out for a period of up to 2 years as follows: camera site ID No.1, 20.12.2018–06.03.2020; No.2, 28.12.2018–04.02.2020; No.3, 03.01.2019–04.02.2020; No.4, 08.11.2019–06.03.2020; No.5, 01.12.2019–06.03.2020.

Location of the camera traps in a semi-mountainous region, part of Central Bulgaria
Setts large spoil heaps with obvious runs (Neal and Cheesman 1996), with more than three entrances, large debris piles with obvious marks (Weber et al. 2013), presence of footprints, fresh signs of digging (Remonti et al. 2006) and latrines were assumed to be main or breeding setts. External paths between sett entrances were considered as additional evidence for the sett usage. The terrain of these sett sites was hilly, covered with Oak (Quercus L. spp.) and Oriental Hornbeam shrubs (Carpinus orientalis Mill.) mixed with orchards, vineyards and croplands (Fig. 1). Human presence is moderate compared to that in mountains and lowlands of Central Bulgaria. It includes several villages at about 5–6 km distance from each other. The terrain consists of smooth hills and numerous ravines overgrown with dense bushes, providing suitable places for digging setts.
Five setts dug at the ravines edges were selected for reliable monitoring. All five chosen setts consisted of two entrances connected by a common path. The camera traps were placed in front of the entrances of the setts, distanced between ca. 2 and ca.5 m, according to the specifics of the terrain of each burrow and the microhabitat around. In all cases, the camera view covered the entrance and the specific resting place in front of it. The following two models of passive infrared camera traps were used: Keep Guard Cam (KG690NV) and HD camera with Black IR (SG56OK – 14 m). Both camera models provided the opportunity for the same modes of operation with the same requisites (temperature, moon phase and timing) of the obtained photos. The devices were set to take three photos with a minute delay. Memory cards and batteries were changed at 85–90 days’ intervals to minimize animals disturbance, so that at no time were the batteries dead or the cards full and no activities missed. Photographs of badgers separated by more than 30-min interval were treated as independent observation events (Meek et al. 2014).
Analysis
To address our first objective, we analyzed seasonal changes in reproductive behaviors observed at the setts. For this purpose, we counted the daily observations of the three types of mating behaviors, i.e., marking, allo-grooming and copulation, as well as observations of cubs as evidence of successful reproduction. Allo-grooming was defined when different parts of the partner’s body were touched simultaneously (Kruuk 1989; Buesching et al. 2003). The most frequently observed contacts between animals were in the muzzle and neck area in the present study. We pooled daily observations from five observed setts.
To address our second objective of comparing the diel activities between reproductive and non-reproductive seasons, we divided the datasets of badger observations into the following four seasons: spring, March–May; summer, Jun–Aug.; autumn, Sep–Nov.; winter, Dec–Feb. We first performed a kernel density interference (Ridout and Linkie 2009) for the snapshot time, i.e., 0:00–23:59, with circularly transformed data (0–2π), to assess diel activity patterns and their comparisons between different seasons, using R 3.5.1 (R Development Core Team 2018) and the package ‘overlap’ (Meredith and Ridout 2021).
We then estimated nocturnality of badgers in each season. For this analysis, we divided the seasonal datasets into four periods of the day, i.e., dawn, day, dusk and night. The crepuscular periods of dawn and dusk were defined as ± 1 h from sunrise and sunset (Theuerkauf et al. 2003; Eggernmann et al. 2009; Gerber et al. 2012). To assess statistical differences in daily activities between seasons and the four periods of the day, we performed generalized linear mixed effect model (GLMM). We used the relative frequency of badger sightings (%) in a day-period per sum of the seasonal sightings at each camera-trapping site as the response variable (using an offset term), with a log link function and a Poisson distribution. For explanatory variables, we used season, four day-periods (dawn, day, dusk and night) and their interaction as fixed effects (as categorical parameters), while camera site ID as random effects. We used maximum likelihood estimation with Wald test (z-value) for the estimated coefficients. We used the ‘glmmTMB’ package for R (Brooks et al. 2017).
To address our third objective of determining the effects of the lunar-cycle on badger activities, we divided the seasonal observation data into the following eight lunar-phases: new moon (NM); waxing crescent (+ WC); first quarter (FQ); waxing gibbous (+ WG); full moon (FM); waning gibbous ( – WG); third quarter (TQ); and waning crescent ( – WC). We then performed GLMM analyses of the relationships between badger sightings and moon phases. We used the relative frequency of badger observations (%) in moon phase per season at each camera-trapping site as the response variable (using an offset term), while moon phase (as a fixed effect) and camera station ID (as random effects) for explanatory variables (with log link function and Poisson distribution). Additionally, to determine potential brightness effects associated with moon phases, we also performed the GLMM on the relationships between badger observations and proportional lunar illuminations. For the analyses, we used the relative proportions of moon illuminations, i.e., 0, NM; 0.25, + / − WC; 0.5, FQ/TQ; 0.75, + / − WG; and 1.0, FM, as explanatory variables. We developed GLMMs in reproductive (i.e., winter) and non-reproductive seasons (i.e., spring and summer) separately. We used maximum likelihood estimation with a Wald test (z-value) for estimated coefficients for all GLMM analyses. In these analyses, we used the ‘glmmTMB’ package.
For all statistical analyses (i.e., Kernel density estimations and GLMM), seasonal sub-datasets with sample sizes less than ≤ 10 or from short sampling durations (e.g., less than 1 consecutive month of camera-trapping) were removed from the analyses; thus, the datasets from six winter, three spring, and one summer season were used (Table 1).
Results
Timing of observed reproductive behaviors
Allo-grooming was observed in early winter (Dec–Jan.) and then reproductive behaviors (i.e., copulation and marking) were mainly observed in late winter (Feb–Mar.; Fig. 2). In the present study both short-term (n = 12, mean = 1.05 min) and long-term copulations (n = 8, mean = 48.63 min) were observed (Table S1 in Suppl. Mat.). Cubs were observed between spring and early summer (Mar–Jul.; Fig. 2). In two cases, it was possible for us to observe bringing of newborns in the mouth of an adult individual (probably the mother) from one entrance of the sett to another, after the last recorded copulation (i.e., in the second half of February; Fig. 2 and Fig. S1 in Supplementary Materials).

Seasonal distribution (represented by 10-days intervals) of activities related to mating as well as cubs observations in European badger from Central Bulgaria—X-axis represents a leap year due to our total sampling periods (i.e., from 20.12.2018 to 06.03.2020)
Seasonal changes in diel activities and nocturnality
Diel activity patterns in badgers differed between winter and spring–summer seasons (Fig. 3) in proximity of setts. In winter, badger activity increased in the early hours of the night, peaking around 18:00, decreasing during the night and almost ending with the dawn onset (Figs. 3a, 4). However, in spring and summer, bimodal patterns were observed in front of setts with peaks around 19:00–20:00 and 6:00 (Fig. 3b).

Diel activity patterns of badgers in front of the dens in Central Bulgaria (used R package ‘overlap’). The different color corresponds to the different datasets (i.e., site-season)

Relative frequency of observations (%) of the European badger (Meles meles) in front of the setts at different time of the day in (a) winter and (b) spring–summer seasons in Central Bulgaria. Horizontal line, box, vertical dashed lines and open circle represent median, range of first and third quantile, min.–max. range, and outlier, respectively
The relative frequency of badger observations at setts dominated at night (median 85.9% of the total observations), over at dawn, day and dusk in winter (Fig. 4a). In spring–summer seasons, however, the differences were between the periods of the day, comprised of 19.7% frequency at dusk to 39.3% frequency at night (Fig. 4b). The results of the GLMM analysis on the relative frequency of observations among the different seasons and four periods of the day indicated that badgers were significantly more active at night than during the other periods of the day in winter (Wald test, P < 0.001 for both seasons and periods of the day as well as their interactions; Table 2). Furthermore, the statistical assessment also indicated that badgers were more active at night in winter in front of setts than in other seasons (Wald test, P < 0.001 for all periods; Table 2).
Changes in badger activities with the lunar cycle
The relative frequency of badger detections close to the setts was highest at new moon (NM) followed by crescent moons (WC + and WC −) and lowest at full moon (FM) in winter (Fig. 5a), statistically indicated by the GLMM analysis (Wald test, P < 0.001 for all moon phases; Table 3). In the spring–summer season, the relative frequency of badger observations in front of setts was also different among moon phases but slightly different from that in winter (Fig. 5a) with statistical significances for new moon (NM; Wald tests, P < 0.001), crescent moons (WC + , P < 0.001; WC − , P = 0.044) and gibbous moons (WG + , P = 0.030; WC − , P = 0.043) obtained by the GLMM analysis (Table 3). The relative frequency of badger observations decreased with increasing proportional moon illuminations in both winter and spring–summer seasons (Wald tests, P < 0.001 for both seasons; Table 4 and Fig. 5b), indicating that badgers were more active with less moon illuminations.

Relative frequency of observations (%) of the European badger (Meles meles) in front of the setts during different lunar phases (a) and proportional moon illumination (b) in winter and spring–summer seasons. Horizontal line, colored box, vertical lines and closed circle represent median, range of first and third quantile, min.–max. range, and outlier, respectively
Finally, we compared the relative frequency of observed reproductive behaviors within setts (i.e., allo-grooming, scent-marking and copulation) at different moon phases (Fig. 6). Allo-grooming and copulations were less frequent at bright nights (i.e. FM and WG), than during other moon phases (Fig. 6).

Relative frequency of observations (%) of scent-marking, copulation and allo-grooming in the European badger (Meles meles) during lunar phase
Discussion
Timing of the reproduction
The mating time frame of the badger is similar throughout its geographical range, although there is potential local variation in the exact timing of reproductive behaviors observed. Winter tracking of a badger in Poland revealed a restriction of its activity in the vicinity of the sett (Goszczynski et al. 2005). As courtship and copulation were described to occur in front of the main sett by Roper (2010), it was possible for us to observe the badgers' reproductive behavior using camera traps. The present study showed that the winter period (from the beginning of December to the end of February) was the breeding season for badgers in Central Bulgaria, taking into account their reproductive behaviors (i.e. marking, allo-grooming and copulation) observed close to the setts. Camera trapping gave us the unique opportunity to register badgers copulative behavior at the setts from mid-January to late February. In line with these results, mating is most likely to happen in February, but it can occur all year round (Roper 2010). The second half of February was also the period with reported mating in Slovakia (Slamka 2016). Increased marking with sub caudal gland secretion by both sexes is typical for badgers mating in winter (Buesching and Macdonald 2001). This type of marking left by a solitary badger in front of a sett is a reliable sign to other individuals in the context of reproduction (Marchlewska-Koj et al. 2001).
For the badger, short-duration copulations (up to 2 min; Johnson 2001, and ≤ 10 min; Lebourgeois 2021) have been described, as well as long-duration ones (from 15 min up to 90 min; Johnson 2001, and ≥ 60 min; Lebourgeois 2021). The long-duration copulations are successful in fertilizing the female and are observed most frequently in the period February–May in both free-living and captive animals (Neal and Cheeseman 1996; Johnson 2001). In the present study, the short-term and more numerous copulations were detected from late January to mid-February. Long-duration copulations were observed in two or three per pair in mid-February (Table S1 in Suppl. Mat.). The female is fertilized after the middle of winter, and the cubs are born as a subsequence of delayed implantation, before the next mating period (Roper 2010). The cubs are generally born between mid-January and mid-March (Neal and Cheesman 1996), and 76% of them in England were born in mid-February (Macdonald et al. 2015). In this study, the mother was photographed carrying the newborns in her mouth from one entrance of the sett to another from mid-February to early March (Fig. S1 in Suppl. Mat.), after the last copulation was observed. The cubs are able to move out of the sett in the period between eighth to ten weeks after birth (Yamaguchi et al. 2006). The first appearances of cubs occurred 6–8 weeks after delivery to the setts by the mother (Fig. S1 in Suppl. Mat.). These results are consistent with the first emergence of cubs in front of the setts in mid-April in France (Lebourgeois 2021), in England (Fell et al. 2006) and in Netherlands (Roper 2010).
Seasonal changes in diel activities
In general, the European badger is known as a typical nocturnal animal (Kowalczyk et al. 2003; Magowan et al. 2022). Indeed, we have already observed the annual nocturnal activities of free-living badgers in our study area, by camera trapping along trails (i.e., far from the setts) (Tsunoda et al. 2018, 2020). In the present investigation we observed predominant nocturnal activity in front of the setts only in winter; however, the animals were also active during daytime in spring–summer seasons (Fig. 3). Consistent with our findings, previous studies also reported that badgers were mostly nocturnal in winter, while their diurnal activities increased during warm seasons in Belarus (Rotenko et al. 2012), Slovakia (Slamka 2016), and Russia (Sidorchuk and Rozhnov 2018). Seasonal changes in badger activity, specifically related to social behavior (Silk et al. 2017), also demonstrate the established trend in the present study. In cool-temperate regions, badgers generally decrease their activity during winter (Kowalczyk et al. 2003), appearing on days with mild weather (Rotenko et al. 2012; Slamka 2016; Lebourgeois 2021); nevertheless, they showed predominant nocturnal sett activities (Rotenko et al. 2012; Slamka 2016). As we discussed in the previous section, mating behaviors in proximity of the setts were mainly observed in winter, and our observations indicated that badgers were more nocturnal in the mating season than in the post-mating one (i.e., cub-rearing).
The effects of moon phase on badger activities
Our surveillance indicated that European badgers were more active at their setts on nights with low lunar illumination (i.e., around new moon) in the mating season. Further, mating behaviors (copulation and allo-grooming) significantly increased from the waning crescent to the first quarter (Fig. 6). Dixon et al. (2006) also reported an increase in badger copulation in the periods from third to first quarters, consistent with our findings. Kronfeld-Schor et al. (2013) consider that lunar-cycle may induce changes in some environmental parameters (e.g., food availability or predation risk), indirectly related to reproductive success. Earthworms, the staple food for badgers in many regions (Goszczynski et al. 2003; Kowalczyk et al. 2003), decrease their activity on the soil surface around full moon (Michiels et al. 2001). In addition, bright night increases the potential risks caused by predators or competitors (Cowgill et al. 1962; Prugh and Golden 2014; Mori and Menchetti 2019). In the study area, the golden jackal (Canis aureus) could be a potential threat to badgers, leading to a separation of their spatio-temporal activities to avoid confrontations (Tsunoda et al. 2018, 2020). In our study, badgers exhibited mating and copulation above ground (i.e., in front of the setts), making them vulnerable to interference by jackals and stray dogs. Nocturnal activities in darkness may be helpful in reducing this risk and thus contribute to reproductive success.
Conclusions
In Central Bulgaria, the European badger exhibited predominantly nocturnal activities in front of the setts during the reproductive season (winter), compared to the cub-rearing season (spring and summer) when the animals were more diurnal. Moreover, our observations indicated that badgers were more active on dark nights around new moon phase and thus reproductive behavior was largely synchronized with the lunar phase. Further research is needed to elucidate the underlying mechanism responsible for the nocturnality and darkness preference of reproductive badgers.
Data availability statement
The datasets generated and/or analyzed during the present study are available upon request from the corresponding author.
References
Brooks ME, Kristensen K, van Benthem KJ, Magnusson A, Berg CW, Nielsen A, Skaug HJ, Maechler M, Bolker BM (2017) GlmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J 9:378–400
Buesching CD, Macdonald DW (2001) Scent-marking behaviour of the European badger (Meles meles): resource defense or individual advertisement? In: Marchlewska-Koj A, Lepri J, Muller-Schwarze D (eds) Chemical signals in Vertebrates 9. Springer, US, Boston, MA, pp 321–327
Buesching CD, Stopka P, Macdonald DW (2003) The social function of allo-marking in the European badger (Meles meles). Behaviour 140:965–980
Carpenter PJ, Pope LC, Greig C, Dawson DA, Rodgers LM, Erven K, Wison GJ, Delahay RJ, Cheeseman CL, Burke T (2005) Mating system of the Eurasian badger, Meles meles, in a high density population. Mol Ecol 14:273–284
Cowgill U, Bishop A, Andrew R, Hutchinson C (1962) An apparent lunar periodicity in the sexual cycle of certain prosimians. Proc Nat Acad Sci 48:238
Cozzi G, Broekhuis F, McNutt J, Turnbull L, Macdonald D, Schmid B (2012) Fear of the dark or dinner by moonlight? Reduced temporal partitioning among Africa’s large carnivores. Ecology 93:2590–2599
Cresswell W, Harris S (1988) The effects of weather conditions on the movements and activity of badgers (Meles meles) in a suburban environment. J Zool 216:187–194
Dixon D, Dixon L, Bishop J, Pettifor R (2006) Lunar-related reproductive behaviour in the badger (Meles meles). Acta Ethol 9:59–63
Eggernmann J, Gula R, Pirga B, Theuerkauf J, Tsunoda H, Brzezowska B, Rouys S, Radler S (2009) Daily and seasonal variation in wolf activity in the Bieszczady Mountains, SE Poland. Mammal Biol 74:159–163
Fell R, Buesching C, Macdonald D (2006) The social integration of European badger (Meles meles) cubs into their natal group. Behaviour 143:683–700
Gerber B, Karpanty S, Randrianantenaina J (2012) Activity patterns of carnivores in the rain forests of Madagascar: implications for species coexistence. J Mammal 93:667–676
Goszczynski J, Jedrzejewska B, Jedrzejewski W (2003) Diet composition of badgers (Meles meles) in a pristine forest and rural habitats of Poland compared to other European populations. J Zool 250:495–500
Goszczynski J, Juszko S, Pacia A, Skoczynska J (2005) Activity of badgers (Meles meles) in Central Poland. Mamm Biol 70:1–11
Griffiths L (2008) Use of a remote camera to rapidly assess Eurasian badger Meles meles occupancy within a sett to be lost due to pipeline laying near Uppingham, Rutland, England. Conserv Evid 5:92–94
Harris S (1982) Activity patterns and habitat utilization of badgers (Meles meles) in suburban Bristol: a radio tracking study. In: Cheeseman CL, Mitson RB (eds) Symposia of the Zoological Society of London 49. Academic Press, London, pp 301–323
Johnson D (2001) Models of coexistence, conflict and cooperation. PhD Thesis, University of Oxford, Oxford
Kowalczyk R, Jedrzejewska B, Zalewski A (2003) Annual and circadian activity patterns of badgers (Meles meles) in Bialowieza Primeval Forest (eastern Poland) compared with other Palaearctic populations. J Biogeogr 30:463–472
Kronfeld-Schor N, Dominoni D, de la Iglesia H, Levy O, Herzog E, Dayan T, Helfrich-Forster C (2013) Chronobiology by moonlight. Proc R Soc B 280:20123088. https://doi.org/10.1098/rspb.2012.3088
Kruuk H (1989) The social badger: ecology and behaviour of a group living carnivore (Meles meles). Oxford University Press, Oxford
Kruuk H, Gorman M, Leitch A (1984) Scent-marking with the subcaudal gland by the European badger, Meles meles L. Anim Behav 32:899–907
Lamamy C, Delgado MM, Kojola I, Heikkinen S, Penteriani V (2022) Does moonlight affect movement patterns of a non-obligate carnivore? Brown bears do not mind that the moon exists. J Zool 316:128–138
Lebourgeois F (2021) Long-term monitoring of activities of badgers (Meles meles L.) in a broadleaved forest in France. Eur J Wildl Res 67:8
Macdonald DW, Newman C, Buesching CD (2015) Badgers in the rural landscape conservation paragon of farmland pariah: lessons from the Wytham badger project. In: Macdonald D, Feber R (eds) Wildlife conservation on farmland, vol 2. conflict in the countryside. Oxford University Press, Oxford, pp 65–95
Magowan E, Maguire I, Smith S, Redpath S, Marks R, Wilson R, Menzies F, O’Hagan M, Scantlebury D (2022) Dead-reckoning elucidates fine-scale habitat use by European badgers Meles meles. Anim Biotelem. https://doi.org/10.1186/s40317-022-00282-2
Marchlewska-Koj A, Lepri J, Muller-Schwarze D (2001) Chemical signals in vertebrates 9. Springer, US, Boston, MA
Meek PD, Ballard G, Claridge A, Kays R, Moseby K, O’Brien T, O’Connell A, Sanderson J, Swan DE, Tobler M, Townsend S (2014) Recommended guiding principles for reporting on camera trapping research. Biodivers Conserv 23:2321–2343
Meredith M, Ridout M (2021) overlap: estimates of coefficient of overlapping for animal activity patterns (ver. 0.3.4), https://CRAN.R-project.org/package=overlap
Michiels NK, Hohner A, Vorndran IC (2001) Precopulatory mate assessment in relation to body size in the earthworm Lumbricus terrestris: avoidance of dangerous liaisons? Behav Ecol 12:612–618
Mori E, Menchetti M (2019) Living with roommates in a shared den: spatial and temporal segregation among semifossorial mammals. Behav Processes 164:48–53
Mori E, Menchetti M, Balestrieri A (2015) Interspecific den sharing: a study on European badger setts using camera traps. Acta Ethol 18:121–126
Neal E, Cheeseman C (1996) Badgers. T & A D Poyser, London
Penetriani V, Kuparinen A, del Mar DM, Palomares F, Lόpez-Bao J, Fedriani J, Calzada J, Moreno S, Villafuerte R, Campioni L, Lourenço R (2013) Responses of a top and a meso predator and their prey to moon phases. Oecologia 173:753–766
Popov V, Sedefchev A (2003) Mammals in Bulgaria. Geosoft EOOD, Sofia (in Bulgarian)
Prigioni C (1999) Meles meles (Linnaeus, 1758). Mitchell-Jones AJ, Amori G, Bogdanowicz W, Kryštufek B, Reijnders PJH, Spitzenberger F, Stubbe M, Thissen JBM, Vohralik V. Zima J. The atlas of European mammals. The Academic Press, London, pp 348–349
Prugh L, Golden C (2014) Does moonlight increase predation risk? Meta-analysis reveals divergent responses of nocturnal mammals to lunar cycles. J Anim Ecol 83:504–514
R Core Team (2018) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna https://www.R-project.org/ (Accessed 21 Nov. 2018)
Racheva V, Zlatanova D, Peshev D, Markova E (2012) Camera traps recorded use of sett sites by badgers (Meles meles L., Mammalia) in different habitats. Acta Zool Bulg 64:145–150
Remonti L, Balestrieri A, Prigioni C (2006) Factors determining badger Meles meles sett location in agricultural ecosystems of NW Italy. Folia Zool 55:19–27
Ridout MS, Linkie M (2009) Estimating overlap of daily activity patterns from camera trap data. J Agric Biol Environ Stat 14:322–337
Roper T (2010) Badger. Harper Collins Publishers, London
Rotenko I, Sydorovich V, Saluk S (2012) Daily activity in badgers (Meles meles) in relation to family group size in north-eastern Belarus. Proc Nat Acad Sci Belarus Biol Ser 57:101–105
Rowcliffe J, Kays R, Kranstauber B, Carbone C, Jansen P (2014) Quantifying levels of animal activity using camera trap data. Methods Ecol Evol 5:1170–1179
Sidorchuk N, Rozhnov V (2018) Daily activity of the European badger (Meles meles, Mustelidae, Carnivora) on setts in Darwin reserve and Meschera National Park (Russia) in summer and autumn. Nat Conserv Res 3:47–56
Silk M, Weber N, Steward L, Delahay R, Croft D, Hodgson D, Boots M, McDonald R (2017) Seasonal variation in daily patterns of social contacts in the European badger Meles meles. Ecol Evol 7:9006–9015
Slamka M (2016) Winter activity of European badger (Meles meles) in Slovakia. Folia Oecol 43:95–100
Stewart P, Ellwood S, Macdonald D (1997) Remote video-surveillance of wildlife – an introduction from experience with European badger Meles meles. Mammal Rev 27:185–204
Theuerkauf J, Jedrzejewski W, Schmidt K, Okarma H, Ruczynski I, Sneizko S, Gula R (2003) Daily patterns and duration of wolf activity in the Bialoweiza forest, Poland. J Mammal 84:243–253
Tsunoda H, Ito K, Peeva S, Raichev E, Kaneko Y (2018) Spatial and temporal separation between the golden jackal and three sympatric carnivores in a human-modified landscape in central Bulgaria. Zool Ecol 28:172–179
Tsunoda H, Newman C, Peeva S, Raichev E, Buesching CD, Kaneko Y (2020) Spatio-temporal partitioning facilitates mesocarnivore sympatry in the Stara Planina Mountains. Bulgaria Zoology 141:125801. https://doi.org/10.1016/j.zool.2020.125801
Weber N, Bearhop S, Dall S, Delahay R, McDonald R, Carter S (2013) Denning behaviour of the European badger (Meles meles) correlates with bovine tuberculosis infection status. Behav Ecol Sociobiol 67:471–479
Yamaguchi N, Dugdale HL, Macdonald DW (2006) Female receptivity embryonic diapauses, and superfetation in the European badger (Meles meles): Implications for the reproductive tactics of males and females. Q Rev Biol 81:33–48
Yoxon P (2008) Using remote cameras to monitor otters. Proceedings of 26th Mustelid Colloquium, 2–31 August 2008, Budapest, Hungary
Zabala J, Zuberogoitia I, Garin I, Aihartza J (2002) Seasonal activity-patterns of badgers (Meles meles) related to food availability and requirements. Est Mus Cienc Nat De Alava 17:201–207
Funding
The study was supported by the Trakia University Research Fund (Project 7AF/19).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors report no potential conflict of interest. Approval from research ethics committees is not required to achieve the objectives of this study, as the experimental work was conducted using a non-invasive method—observations using camera traps.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
About this article

Cite this article
Peeva, S., Raichev, E., Georgiev, D. et al. European badger's mating activities associated with moon phase. J Ethol 41, 15–24 (2023). https://doi.org/10.1007/s10164-022-00762-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10164-022-00762-1