
Abstract
Facilitation is an ecological interaction in which the presence of one species (e.g., ecosystem engineers) alters the environment in a way that enhances growth, survival or reproduction of a neighboring species. Wood-boring insects are considered facilitators for cavities-nesting ants, which experience intense intra and interspecific competition for these sites. But how do ants find these cavities? Here, we suggest that ants could be attracted by chemical/odorous cues emitted by the frass of wood-boring insects. Using captive colonies of Cephalotes pusillus (Klug 1824) in controlled conditions, we carried out a bioassay to test the following hypotheses: (i) the frass from beetles are more attractive to ants than those from caterpillars (Cossidae: Lepidoptera), since boring beetles are more common and produce more cavities for ant nest on the studied plant, Caryocar brasiliensis (Caryocaraceae). If this first hypothesis is true, (ii) in relation to the cavity substrate type, the insect frass will be an attraction for workers from captive colonies (colonies, hereafter): both from compromised colonies (colonies without nest and in vulnerable condition) and intact colonies in tubes and (iii) and workers from compromised colonies colonize tubes with frass more quickly than those from intact colonies. The three hypotheses were confirmed, and Cephalotes pusillus ants prefer beetle frass, moving more quickly (five times faster) into tubes with frass when in vulnerable conditions. Our findings revealed that frass from beetles is a trigger to workers of C. pusillus locate and then nesting on branches of C. brasiliensis in Brazilian Cerrado. We suggest that the selection for a nesting cavity by arboreal ants is not random, but guided by chemical/odorous cues.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Plant–animal interactions are one of the main structural factors in conserved natural communities. These mutual relationships may be positive, negative or neutral to one of both partners varying since predation to mutualism (Thompson 2014). Among these interactions, facilitation has been defined as an interaction in which the presence of one species alters the environment in a way that enhances growth, survival or reproduction of neighboring species (Bronstein 2009). For instance, several ant species that nest inside plant trunks and stems need the action of facilitators (Powell 2008, 2009; Santos et al. 2017).
In terrestrial ecosystems, many facilitators species act as ecosystem engineers, which are organisms that modulate the availability of resources to other species, causing physical changes in living and non-living materials (Jones et al. 1994; Lill and Marquis 2003; Velasque and Del-Claro 2016), increasing trophic interactions and species diversity (Sanders et al. 2014; Cornelissen et al. 2016; Calixto et al. 2021). Protection against natural enemies and harsh abiotic conditions has been considered one of the main benefits produced by ecosystem engineers (see Velasque and Del-Claro 2016). For instance, building shelters (e.g., mines, galls and leaf rolls) increases the survival and growth of insects, influencing the entire associated community (Cornelissen et al. 2016 and therein).
Choosing a nesting site is pivotal for ant species which inhabit fragile, ephemeral, and pre-existing cavities, such as hollow branches (Pratt and Pierce 2001). Many ant genera (e.g., Camponotus, Cephalotes, Crematogaster and Pseudomyrmex) are specialized in nesting on cavities made by wood-boring insects, founding new colonies or expanding their existing colonies (e.g., satellite nests) (Hölldobler and Wilson 1990; Yamamoto and Del-Claro 2008; Powell 2009). Beetles of the family Cerambycidae, Buprestidae, and Curculionidae, for instance, dig galleries inside and on the dead wood surface of vegetation (Hanks et al. 2005; Satoh et al. 2016). When abandoned, these galleries and tunnels provide nesting sites for many ant species through intense intra and inter-specific competition (Powell 2009).
In the Brazilian Cerrado, the largest and most diverse savanna in the world, Cephalotes ants select the cavities according to the optimal fit between the area of the cavity entrance and the head area of the largest workers, to defend their nests against predators and other ants (De Andrade and Urbani 1999; Powell 2008). Thus, in Cerrado the nest-site is an important resource for cavity-nesting ants, what limits colony establishment and structures ant community composition. However, this ecosystem is frequently disturbed by fire, forcing compromised colonies like those of Cephalotes ssp. to move to new nests, what ants must do quickly to avoid the fire and the attack of natural enemies (Fagundes et al. 2015). Although some physical features of cavities preferred by ants are known (De Andrade and Urbani 1999; Pratt and Pierce 2001; Powell 2008), the pattern of searching (i.e., random or attraction-driven) for new nesting sites is still unknown. For instance, colonies with more workers or mass foraging such as compromised colonies (i.e., colonies without physical nest and in vulnerable condition) are expected to be more efficient at locating and founding new or satellite nests than small colonies or those that forage at a lower intensity, such as intact colonies (i.e., colonies with physical nest).
From previous observations, Cephalotes ants on branches of Caryocar brasiliensis (Caryocaraceae) have been recorded close to insect frass, a mix of feces and sawdust from the digging of logs by insects. Although studies suggest that these ants use cavities made by beetles (Andrade and Urbani 1999; Powell 2008), recently a wood-boring caterpillar (Cossidae: Lepidoptera) was described as an important pest of C. brasiliensis (Leite et al. 2011). Thus, ants may use cavities formed by the action of facilitators beetles and cossid larvae in C. brasiliensis (Fig. 1). But how do ants find these cavities? Here, we suggest that ants could be attracted by chemical/odorous cues present in the frass. For this, three hypotheses were tested: (i) the frass from beetles are more attractive to ants than those from cossid larvae, since boring beetles are more common and produce more cavities for an ant nest on the studied plant, Caryocar brasiliensis. If this first hypothesis is true, (ii) in relation to the cavity substrate type, the preferred insect frass will be an attraction for workers from captive colonies (colonies, hereafter): both from compromised colonies (i.e., colonies without physical nest and in vulnerable condition) and intact colonies (i.e., colonies with physical nest) in tubes and (iii) and workers from compromised colonies move on to tubes with preferred frass more quickly than those from intact colonies.

Stems of Caryocar brasiliensis (Caryocaraceae, popular name Pequi) with the presence of wood boring beetle (a—frass; and b—the larvae inside the stem) and Lepidoptera (Cossidae; c and d) larvae. A typical entrance nest of Cephalotes ants e, f in a hollowed trunk of Pequi
Methods
Study area
The study was conducted in a Cerrado sensu stricto area of the Brazilian savanna reserve Clube Caça e Pesca Itororó de Uberlândia (CCPIU; 48°17’W; 18°58’S), located in the municipality of Uberlândia, Minas Gerais State, Brazil. The annual mean temperature is 23 °C and mean annual rainfall is approximately 1500 mm, of which > 90% occurs during the wet season (October–April) (Vilela et al. 2014).
Studied species
Cephalotes pusillus (Klug 1824) is a polydomous species occupying many nests which are abandoned or natural cavities on vegetation, presenting small colonies (< 200). This arboreal small ant feeds mainly on nectar and pollen, and has wide distribution in the Neotropical region (Del-Claro et al. 2002; Wild 2007). Workers of C. pusillus were seen foraging close to insect frass on branches of C. brasiliensis. The ‘Pequi’, popular name of C. brasiliensis, is considered a tree symbol of the Cerrado, occurring from north to south of Brazil in this vegetation formation (Prance and Freitas de Silva 1973). This plant species has great economic, social and ecological importance in the savanna, being protected by Brazilian environmental legislation. Cephalotes pusillus as well as many others ant species forage on C. brasiliensis due to its extrafloral nectaries (Oliveira 1997).
Nesting site selection: bioassays
Colonies of C. pusillus were collected in C. brasiliensis trees and maintained in glass tubes in the laboratory under controlled conditions for 1 week before used in bioassays. A total of 30 captive ant colonies were used. We used independent colonies of similar size: from 20 up to 30 workers, plus a few larvae and eggs (< 10 in total per colonies) and without queens. We built arenas using plastic trays (35 × 25 × 5 cm), with two glass tubes (2 cm in diameter and 10 cm in length). At the edge of the arenas, polytetrafluoroethylene (Whitford, UK) was used to prevent workers from escaping. All arenas were under normal daylight cycle, air intact (24 °C) and humidity of 65–75%.
Fresh frass from beetles and cossid larvae communities were collected from C. brasiliensis and used in the bioassays. To determine the insect responsible by the frass in the field, each stem with frass was opened carefully. A teaspoon (5 ml) (~ 15 g) of frass from beetles were placed inside a tube, while the same amount of frass from cossid larvae were added to another tube, both placed on the same side of arena (spaced 20 cm apart). Since frass from beetles and cossid larvae had probably a similar composition, a full teaspoon represented similar weights (~ 15 g). To test the frass preference, in each arena one colony was placed (compromised colonies—workers scattered in the arena without a physical nest) in the opposite site of the arena (10 colonies). The bioassays started at 8 a.m. in the first day and were followed-up every 30 min. When one tube with frass from beetles/or from cossid larvae was occupied by all workers (excluding some individuals visits), the time of colonization was recorded. Each bioassay lasted 48 h. Between 9 p.m. and 6 a.m. of the next day, the colonies were monitored using a Sony handycam. The same colony was never used in the same type of laboratorial bioassays.
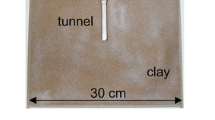
To investigate ant preference regarding the type of substrate, frass or sawdust (from the same wood, grasped with a knife), we used the same methodology mentioned above, but we used only fresh frass (15 g) from beetles, since this frass type were more available. In another tube we used 5 ml (~ 15 g) of sawdust from a dry trunk of C. brasiliensis. We weighed 15 g of sawdust on an analytic balance and put this weight on a teaspoon (full teaspoon), to view the weight estimate which was used as our standard measurement. These experiments were carried out independently with compromised colonies (N = 10) and with colonies kept in tubes (intact colonies) (N = 10) (Fig. 2a, b). The intact colonies, with ants previously conditioned in the tubes, were used to test if the frass have the attraction enough to elicit a nest move. The experiments were monitored (every 30 min) and the time of colonization was recorded as described above. After all bioassays, ants were returned to the field.

Experimental design (e.g., arenas with ants and experimental tubes) showing the preferences of compromised a and intact b colonies regarding beetle frass or sawdust nesting. c Number of colonized nests regarding the types of substrates (frass or sawdust) in compromised and d intact colonies. *Represents significant difference (p < 0.05)
Data analysis
To test the effect of frass type (from beetles or from cossid larvae) on nest colonization, we did a binomial test, considering the proportion of nest colonization as response variable, and the frass type as explanatory variable. To test whether the nest colonization is affected by type of substrate considering compromised and intact colonies, we did another binomial test. As explanatory variable, we used the type of substrate (frass or sawdust) and we used the proportion of nest colonization as response variable. Finally, we built a GLMM using Gaussian error distribution, after verifying normality (‘qqplots’ and Shapiro tests), to test whether the time of nest colonization (variable normally distributed) is affected by the type of colonies (compromised or intact colonies). We considered the time (total number of hours) of nest colonization as response variable, type of colonies as explanatory variable and experiment days as random effects. The residuals of all models were analyzed to verify the adequacy of the models (Crawley 2013). The analysis was conduct in the software R (version 4.0.0) (R Core Team 2020).
Results
The workers of C. pusillus showed a clear preference for nesting in tubes with frass from beetles than those with frass from cossid larvae (χ2 = 9.8; p < 0.01, N = 10) (Fig. 3A), being that in 90% of the bioassays the ants colonized tubes with frass from beetles, taking 11.90 ± 7.39 h (mean ± SD) to adopt the nest. Both compromised and intact colonies preferred to colonize tubes with frass (χ2 = 9.8; p < 0.01, N = 10; χ2 = 5.0; p = 0.02, N = 10, respectively) than those with sawdust (Fig. 2C, D). In both bioassays, the ants colonized 90% (compromised colonies) and 80% (intact colonies) of tubes with frass. Although both types of colony have a preference for frass, compromised colonies colonized tubes with frass five times faster than intact colonies (GLMM: F1,9 = 13.61; p < 0.01, N = 10) (Fig. 3B). Therefore, compromised colonies spent 3.00 ± 1.87 h (mean ± SD) to colonize while intact colonies spent 16.44 ± 11.80 h (mean ± SD).

a Number of colonized nests among the different types of frass (those of beetles and those from cossid larvae). b Nest colonization time (in hours) between different types of colonies (compromised and intact colonies). Boxes show the median, 25th and 75th percentiles; whiskers indicate minimum and maximum values. Points represent each of the observations of the time of colonization in both types of colonies. *Represents significant difference (p < 0.05)
Discussion
Our findings revealed workers of C. pusillus appear to use beetle frass as a cue for locating nests on branches of C. brasiliensis, although it could also simply be that the ants prefer cavities with beetle frass than with either lepidopteran frass or sawdust. Both compromised (vulnerable) and intact captive colonies were attracted to beetle frass. Moreover, the ants that do not have a physical nest (compromised colonies) colonizes more quickly (five times faster) the tubes with beetles’ frass than those from intact colonies (i.e., colonies with physical nest). For the first time, we have shown that the selection for a nesting cavity by arboreal ants can be non-random, therefore being probably guided by a chemical/odorous clue.
The preference for tubes with beetles’ frass is intriguing. Beetles often dig narrow and resistant cavities on hard branches such as dead wood (Banno and Yamagami 1991; Hanks et al. 2005; Satoh et al. 2016), while the caterpillar digs very long cavities with several lateral paths on softer branches (personal observation, but see Leite et al. 2011). We suggest that due to the fact that beetles dig in harder wood, these cavities probably have less moisture and fungi. The moisture into the nest can be positive associated with microbial activity (Coenen-Stass et al. 1980), leading to contamination of newly established ant colonies. Therefore, cavities made by beetles offer greater protection (e.g., physical structure) against invaders (e.g., Azteca ants) and pathogens (Powel 2009).
In Cerrado, the days are very hot and dry and there is also an abundance of many species of predator ants (e.g., Pseudomyrmex and Ectatomma spp.), spiders and birds (Oliveira and Marquis 2002), so it is imperative to an ant colony to find a place to hind and nest. Even workers who already had a nest (intact colonies in tubes) migrated to another tube containing frass. Likewise, workers scattered in the arena without a physical nest (compromised colonies) quickly located the tube with frass and colonized it. The higher speed of workers from compromised colonies in nesting on tube with beetle frass might be explained by the greater number of foraging workers in the arena. Many workers from intact colonies may stay for longer periods inside the nests and only few workers may go out to forage. In any case, beetle frass is clearly an attractant, and the found pattern does not reflect any deterrent properties of coccid frass.
Insects often use chemical/odorous as orientation cues. For instance, odor cues are used by ants to locate food resources such as leaves (Roces 1990) and nectar (Provecho and Josens 2009). This way, it is likely that the same can happen when these ants are looking for nesting sites, since the choice of the nesting place is as pivotal as the search for food resources, for instance. Moreover, we cannot rule out that Cephalotes ants can also detect frass through visual cues, such as C. atratus, which perceives the reflectance properties of the trunks in the canopy (Yanoviak and Dudley 2006). Finally, Cephalotes ants can also use the beetles' frass as an organic material to modify the shape of entrance of their nests, improving nest use such as defense against invaders (Priest et al. 2021).
This study is a first step to investigate how ants find their nesting sites. Our findings show that besides the beetles acting as ecosystem engineers, they leave indirect cues to facilitate colonization by ants. Facilitation like this one, is not a simple type of interaction; rather, as argued by Mathis and Bronstein (2020) it is a suite of phenomena associated with distinct ecological processes and evolutionary consequences. Studies that assess the chemical composition and odors of frass from beetles should be encouraged. Therefore, understanding whether ants use the same patterns for searching for food and nesting sites will increase our ability to understand ecological–evolutionary mechanisms of the behavior of social insects.
References
Banno H, Yamagami A (1991) Life cycle and larval survival rate of the redspotted longicorn beetle, Eupromus ruber (Dalman) (Coleoptera: Cerambycidae). Appl Entomol Zool 26:195–204
Bronstein JL (2009) The evolution of facilitation and mutualism. J Ecol 97:1160–1170
Calixto ES, Santos DFB, Anjos DV, Colberg E (2021) How plant-arthropod interactions modify the environment: concepts and perspectives. In Del-Claro K, Torezan-Silingardi (eds) Plant-Animal Interactions: Source of Biodiversity. Springer Nature, Switzerland AG. https://doi.org/10.1007/978-3-030-66877-8
Coenen-Stass D, Schaarschmidt B, Lamprecht I (1980) Temperature distribution and calorimetric determination of heat production in the nest of the wood ants Formica polyctena (Hymenoptera: Formicidae). Ecology 61:238–244
Cornelissen T, Cintra F, Santos J (2016) Shelter-building insects and their role as ecosystem engineers. Neotrop Entomol 45:1–12
Crawley MJ (2013) The R book. Wiley, Chichester
De Andrade ML, Urbani CB (1999) Diversity and adaptation in the ant genus Cephalotes, past and present. Staatliches Museum für Naturkunde.
Del-Claro K, Santos JC, Júnior ADS (2002) Etograma da formiga arborícola Cephalotes pusillus (Klug, 1824) (Formicidae: Myrmicinae). Revista De Etologia 4(1):31–40
Fagundes R, Anjos DV, Carvalho R, Del-Claro K (2015) Availability of food and nesting-sites as regulatory mechanisms for the recovery of ant diversity after fire disturbance. Sociobiology 62(1):1–9
Hanks LM, Paine TD, Millar JG (2005) Influence of the larval environment on performance and adult body size of the wood-boring beetle Phoracantha semipunctata. Entomol Exp Appl 114:25–34
Hölldobler B, Wilson EO (1990) The ants. Harvard University Press, Cambridge
Jones CG, Lawton JH, Shachak M (1994) Organisms as ecosystem engineers. Oikos 69:373–386
Leite GLD, Alves SM, Nascimento AF, Lopes PSDN, Ferreira PSF, Zanuncio JC (2011) Identification of the wood-borer and the factors affecting its attack on Caryocar brasiliense trees in the Brazilian Savanna. Acta Sci Agron 33:589–596
Lill JT, Marquis RJ (2003) Ecosystem engineering by caterpillars increases insect herbivore diversity on White Oak. Ecology 84:682–690
Mathis KA, Bronstein JL (2020) Our Current Understanding of Commensalism. Annu Rev Ecol Evol Syst, 51.
Oliveira PS (1997) The ecological function of extrafloral nectaries: herbivore deterrence by visiting ants and reproductive output in Caryocar brasiliense (Caryocaraceae). Funct Ecol 11(3):323–330
Oliveira PS, Marquis RJ (2002) The cerrados of Brazil: ecology and natural history of a neotropical savanna. Columbia University Press, New York
Powell S (2009) How ecology shapes caste evolution: Linking resource use, morphology, performance and fitness in a superorganism. J Evol Biol 22:1004–1013
Powell S (2008) Ecological specialization and the evolution of a specialized caste in Cephalotes ants. Funct Ecol 22:902–911
Prance GT, Freitas de Silva MF (1973) Caryocaraceae. Flora Neotropica 12:1–75
Pratt SC, Pierce NE (2001) The cavity-dwelling ant Leptothorax curvispinosus uses nest geometry to discriminate between potential homes. Anim Behav 62:281–287
Priest GV, Camarota F, Vasconcelos HL, Powell S, Marquis RJ (2021) Active modification of cavity nest-entrances is a common strategy in arboreal ants. Biotropica. https://doi.org/10.1111/btp.12922
Provecho Y, Josens R (2009) Olfactory memory established during trophallaxis affects food search behaviour in ants. J Experim Biol 212:3221–3227
R Development Core Team 2020. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna. http://www.R-project.org.
Roces F (1990) Olfactory conditioning during the recruitment process in a leaf-cutting ant. Oecologia 83:261–262
Sanders D, Jones CG, Thébault E, Bouma TJ, Van der Heide T, Van Belzen J, Barot S (2014) Integrating ecosystem engineering and food webs. Oikos 123:513–524
Santos L, Feitosa R, Carneiro M (2017) The role of senescent stem-galls over arboreal ant communities structure in Eremanthus erythropappus (DC.) MacLeish (Asteraceae) Trees. Sociobiology 64:7–13
Satoh T, Yoshida T, Koyama S, Yamagami A, Takata M, Doi H, Hata Y (2016) Resource partitioning based on body size contributes to the species diversity of wood-boring beetles and arboreal nesting ants. Insect Conservation and Diversity 9:4–12
Thompson JN (2014) Interaction and coevolution. University of Chicago Press, Chicago
Velasque M, Del-Claro K (2016) Host plant phenology may determine the abundance of an ecosystem engineering herbivore in a tropical savanna. Ecol Entomol 41:421–430
Vilela AA, Torezan-Silingardi HM, Del-Claro K (2014) Conditional outcomes in ant-plant-herbivore interactions influenced by sequential flowering. Flora 209:359–366
Yamamoto M, Del-Claro K (2008) Natural history and foraging behavior of the carpenter ant Camponotus sericeiventris Guérin, 1838 (Formicinae, Campotonini) in the brazilian tropical savanna. Acta Ethol 11:55–65
Yanoviak SP, Dudley R (2006) The role of visual cues in directed aerial descent of Cephalotes atratus workers (Hymenoptera: Formicidae). J Experim Biol 209:1777–1783
Wild AL (2007) A catalogue of the ants of Paraguay (Hymenoptera: Formicidae). Zootaxa 1622:1–55
Acknowledgements
We would like to thank Renan Moura for valuable suggestions on earlier versions of this manuscript and thank the “Clube de Caça e Pesca Itororó de Uberlândia” for permitting the use of its Cerrado reserve.
Funding
K. Del-Claro thanks Conselho Nacional de Ciência e Tecnologia (CNPq) (PQ grant: 301605/2013–0 and Universal 400748/2016–9) and also Fundação de Amparo à Pesquisa de Minas Gerais (Fapemig) for financial support. D. V. Anjos thanks CAPES (‘Coordenação de Aperfeiçoamento de Pessoal de Nível Superior’)—Finance code 88887.466638/2019–00 for financial support.
Author information
Authors and Affiliations
Contributions
KD-C, HMT-S: Conceptualization, methodology, data curation, visualization, investigation, writing—reviewing and editing. DVA: Software, writing—reviewing and editing.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
About this article

Cite this article
Del-Claro, K., Anjos, D.V. & Torezan-Silingardi, H.M. Ant nesting site selection mediated by insects frass. J Ethol 40, 31–36 (2022). https://doi.org/10.1007/s10164-021-00724-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10164-021-00724-z