
Abstract
We evaluated the fourth stage of the “Conservation and Management Plan for Sika Deer (Cervus nippon) in Hokkaido, Japan (CMPS4)”, focusing on its cost-effectiveness and sika deer migration between two management areas of eastern and western Hokkaido. To clarify these factors, we constructed a stochastic matrix population model that accounts for deer migration and several uncertainties. We assumed four different budget scenarios and simple rules regarding nuisance control, and simulated four alternative management strategies. In the stochastic simulation, we calculated the probability of successfully satisfying the population target given by the CMPS4, an average total actual management cost, and a cost-effectiveness index given four budget conditions of migration rate and budget allocation ratio. The simulation results suggest the following. First, the current management budget is so small that the probability of successfully satisfying the population targets in both areas is only 26–30 %. If the total budget remains small, it should be almost entirely invested in one area, regardless of migration situation, to maximize the probability of successfully meeting the target density in at least that area. However, these probabilities of success decrease with greater migration rate. Second, when the government invests more of its budget in the early management stage, the expected total actual cost decreases and the probability of management success increases. These findings represent cost-effective management strategies for satisfying the CMPS4 targets.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The biological and economic implications of migration effects on wildlife management for metapopulations in which subpopulations are connected have been addressed (Sanchirico and Wilen 1999, 2005). However, population is frequently divided by administrative boundaries and wildlife moves between management areas (Clutton-Brock et al. 2002; Bhat and Huffaker 2007). In the context of theoretical analysis of adaptive management for wildlife, these migration effects on management are often ignored (Yamamura et al. 2008).
In 2000, the government of Hokkaido began implementation of the “Conservation and Management Plan for Sika Deer (Cervus nippon)” (CMPS) owing to increased agricultural damage with increase of the sika deer population (Hokkaido Government 2000). The CMPS uses feedback control as a management policy, aimed at the sustainable use of natural resources at an optimal level under uncertain parameters (Kaji et al. 2010), making this policy useful for wildlife management. Feedback control for wildlife management was originally developed by Tanaka (1982) for implementation in commercial whaling. To apply the feedback program to deer management, the Hokkaido government developed an adaptive management model based on relative population size, accounting for uncertainties (Matsuda et al. 1999), and divided Hokkaido into eastern, western, and southern management areas (Fig. 1) according to administrative boundaries (Hokkaido Government 2012). In this management program, deer are harvested by game hunting and nuisance control in the eastern and western areas. In the present study, we did not consider the southern area, where the deer population density remains low.

Management areas for sika deer on Hokkaido Island, Japan (eastern, western, and southern management areas). The Hidaka subpopulation is divided into the eastern and western management areas. Source Japanese National Digital Information Data base (administrative district)
In Japan, hunters must pay an annual registration fee to prefectural governments to participate in game hunting. Under normal management plans, game hunting is regulated by the open season period and bag limits, in daily deer management units per hunter. In the case of the CMPS in Hokkaido, there is no bag limit on the number of female deer per day, but there is a bag limit of one male deer per hunter per day. In many regions of Hokkaido, the open season for deer is from October through February.
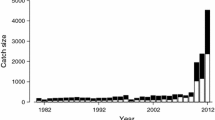
The number of game hunters can be estimated from the number of hunters registered in previous years. Hunters are paid rewards from their municipal government for each deer culled during nuisance control periods, which are mainly during the closed season from March through October. The allowable number of individuals culled so depends on the fixed and limited budget of each municipality, with support from the prefectural government. In recent years, the total annual deer harvest has been rapidly increasing (Fig. 2). However, deer population density has not yet declined within the target range specified by the CMPS (Hokkaido Government 2012). Considering this situation, the Hokkaido prefectural government has published the fourth stage of the CMPS (CMPS4) to adequately control the Hokkaido deer population (Hokkaido Government 2012). However, the government overlooked two fundamental problems when drafting the CMPS4. First, the plan does not describe the cost-effectiveness of deer management; second, it does not account for deer migration between management areas.

Total number of deer harvested in the a eastern and b western management areas during 1993–2013 in Hokkaido (K. Yamamura et al., unpublished data)
In an economic context, the CMPS4 does not explicitly calculate the average management cost to meet the CMPS4 target or cost-effectiveness under various possible scenarios. Estimating these pieces of economic information is difficult, but it is important to wildlife management (Hughey et al. 2003; Baxter et al. 2006; Naidoo et al. 2006).
Because of migration, the distribution of deer across Hokkaido has expanded rapidly since the 1950s (Kaji et al. 2010). Seasonal deer migration has recently been observed in each Hokkaido subpopulation (Uno and Kaji 2000; Igota et al. 2004). In their study on the effects of migration in Hokkaido deer management, Ou et al. (2014) indicated culling effect of one subpopulation on an adjacent subpopulation is not large because deer do not significantly change their breeding habitat. However, the boundary defined by the CMPS4 splits the Hidaka subpopulation, resulting in their habitat to lie within both the eastern and western areas (Fig. 1). The Hidaka subpopulation is one of the main subpopulations of sika deer in Hokkaido (Kaji et al. 2010). Hence, it is very important to consider the migration between management areas in the CMPS4.
Analyses of Hokkaido deer have been previously carried out using matrix population models (Matsuda et al. 1999, 2002; Yamamura et al. 2008; Ueno et al. 2010); however, these models assumed no migration between the eastern and western areas and economical effect was not evaluated for deer management.
In this study, we constructed a stochastic matrix population model that accounts for migration between the management areas, uncertainties under insufficient information, and the limited total budget for deer management. The probability of achieving the CMPS4 target density depends on the available budget. The total actual cost is not determined because of data uncertainties, especially those resulting from measurement error of the initial population size. To obtain an efficient strategy for implementing the CMPS4, we: (1) devised fixed budget management options that reflected actual management conditions; (2) performed stochastic simulation to calculate the probability of meeting the CMPS4 target population and the total actual cost; and (3) evaluated the effects of deer migration on cost-effectiveness of the CMPS4.
Methods
Matrix population model
To evaluate the CMPS4 performance, we constructed a stochastic matrix population model that includes uncertainties. Individuals within the Hokkaido deer population were considered three categories (calf, adult female, or adult male), and these categories were considered in the matrix. Biological and management events were sorted into four phases. In the first phase, deer are harvested through game hunting (late October through February) or die of natural causes (mainly January through April). In the second phase, we presumed a migration event; in Hokkaido, deer migrate seasonally between wintering and breeding habitats (Uno and Kaji 2000; Sakuragi et al. 2003; Igota et al. 2004). Those two habitats exist in both the eastern and western management areas. We defined “migration” as a shift of breeding site from that of the previous year between the eastern and western areas, which does not include the seasonal migration between wintering and breeding habitats. In the third phase, adult female deer give birth to their calves, and calves born in the previous year mature into adults (Kaji et al. 2010). In the final phase, the Hokkaido government implements nuisance control after the breeding season (March through late October).
We accounted for population structure, and the following management and biological events between census periods: winter survival, migration between management areas, reproduction and maturation, and nuisance control. The survival rates during wintering are given by matrix L t . The migration event is given by matrix M. Reproduction and maturation events are given by matrix G t . The nuisance control vector is given by C t . Overall transitions of individuals within the year are obtained by these three component matrices and one vector. Then, we have the stochastic matrix population model:
This model begins in October and divides Hokkaido into two areas with elements of the three population categories; thus, the population vector in year t is
where N t,i,j is the number of deer in year t, area i (i = e or w), and category j (j = c, f, m); T indicates matrix transposition, and area divisions (e, w) are the eastern and western management areas, respectively, as defined by the CMPS4; categories c, f, and m are calf, adult female, and adult male, respectively.
We constructed a winter survival matrix in each area. When a mother dies in winter, her calf cannot survive until spring (Matsuda et al. 1999). The winter survival matrix \({\mathbf{L}}_{t}\) can be expressed as
where \(L_{t,i,j}\) is the winter survival rate in year t in area i of category j. In winter, deer die from both game hunting and natural causes. We assumed that the survival rate under natural conditions is the same in both management areas. Therefore, the survival rate \(L_{t,i,j}\) can be expressed as
where \(\rho_{j} S_{t}\) is the survival rate under natural conditions in year t of category j. \(S_{t}\) and \(\rho_{j}\) are the adult female survival rate under natural conditions and a multiplier to convert that rate to another category, respectively; \(\rho_{j}\) was calculated from the estimated average survival rate given in a report by an investigative commission on deer (K. Yamamura et al., unpublished data). Observations to estimate \(S_{t}\) were performed on the Shiretoko Peninsula of Hokkaido (Kaji et al. 2004). However, observations from several other studies have reported mortality values varying with environment, such as those with severe winters (Takatsuki et al. 1994; Uno et al. 1998). To reflect these variations, we assumed that \(S_{t}\) follows a beta distribution with mean value and variance S and \({{S^{2} } \mathord{\left/ {\vphantom {{S^{2} } {300}}} \right. \kern-0pt} {300}}\), respectively (Yamamura et al. 2008). The parameters of the beta distribution are \(\alpha_{S}\) and \(\beta_{S}\) (\(\alpha_{S} = 17.06\), \(\beta_{S} = 1.0889\), Yamamura et al. 2008). \((1 - q_{i,j} E_{t,i,j} )\) is the survival rate against game hunting in year t in area i of category j, and \(q_{i,j}\) indicates game hunting efficiency (catchability) in area i of category j. \(E_{t,i,j}\) denotes hunting effort in year t in area i of category j. We estimated \(q_{i,j}\) by the maximum likelihood method (see below). In the estimation, we used catch per unit effort (CPUE) data and estimated deer population size as reported by the Hokkaido government and the Eastern Hokkaido Wildlife Research Station, Nature Conservation Department, Hokkaido Institute of Environmental Sciences (Table 1). CPUE is an indicator that can be used to determine hunting efficiency (Uno et al. 2006). We calculated the log-likelihood given by the relationship between CPUE and the estimated size of the deer population:
where \(\varepsilon_{t,i,j}^{CPUE}\) is observation error in year t in area i of category j, which follows a lognormal distribution with mean 0 and variance \(\sigma_{CPUE,i,j}^{2}\). Hence, log-likelihood can be expressed as a function of the number of observations n:
We used R (Ver. 3.0.1) software package “optim” to estimate parameter values. All estimated parameters were statistically significant (P < 0.001). We considered the process error in hunting effort, \(E_{t,i,j}\) as
where \(\bar{E}_{i,j}\) is the average effort in area i of category j, \(\varepsilon_{t,i,j}^{E}\) follows normal process error on a logarithmic scale in year t in area i of category j with mean 0 and variance \(\sigma_{E,i,j}^{2}\). The term \(- 0.5\sigma_{E,i,j}^{2}\) is a bias correction for lognormal errors (Haltuch et al. 2008). We calculated these average efforts and process errors from observed data for the period 2004–2008, as reported by the Institute of Environmental Sciences, Hokkaido Research Organization (unpublished data).
Many studies have provided examples of matrix population models accounting for migrations that can be used to evaluate wildlife management (e.g., Hunter and Caswell 2005; Mantzouni et al. 2007). The migration matrix \({\mathbf{\rm M}}\) is given by
where \(m\) is the probability that an individual of a given category moves to a neighboring area after the breeding season. The migration matrix describes the probabilities of individual migration to the neighboring management area. However, it is difficult to estimate the migration rate for each category and area, because of a lack of observation data regarding dispersal between the eastern and western areas. For simplicity, we used fixed migration rates with the same value for each category and area.
In Hokkaido, density dependence of the deer mortality or reproduction rate does not appear until the population is near carrying capacity (Kaji et al. 1988). Therefore, we did not include density dependence in our model. The reproduction and category transition matrix \({\mathbf{G}}_{t}\) is
The sex ratio among calves is 1:1 (Takatsuki 1998). The reproduction rate \(r_{t}\) follows a beta distribution with an expected value \(2r\) and variance \({{r^{2} } \mathord{\left/ {\vphantom {{r^{2} } {75}}} \right. \kern-0pt} {75}}\) (Yamamura et al. 2008). The beta distribution parameters are \(\alpha_{r}\) and \(\beta_{r}\) (\(\alpha_{r} = 2 9. 1\), \(\beta_{r} = 3. 2 3 3 3\)) (Yamamura et al. 2008).
We reflected realistic management by nuisance control, rather than by game hunting, in the CMPS4. The vector of the number of animals culled by nuisance control \({\mathbf{C}}_{t}\) is
where \(C_{t,i,j}\) is the nuisance control level (the number of animals culled) in year t in area i of category j. \(C_{t,i,j}\) is separated into three components:
where \(\varphi_{t}\) is the management multiplier (0 or 1) given by the nuisance control rule and \(\gamma_{i,j}\) is the allocation rate of nuisance control of category j area i. These values were calculated from observed nuisance control data from 2004–2008. \(C_{i}^{max}\) is the maximum allowable number of animals culled via nuisance control in area i. Considering category-specific nuisance control, management is usually “tailored” to the age and sex structure of the population, rather than to simple population counts (Gordon et al. 2004). The importance of harvesting female deer has been previously mentioned (Matsuda et al. 1999; Milner et al. 2006). The Hokkaido government began aggressive female deer harvesting in 1998 (Kaji et al. 2010). In recent years, the total harvest number (by both nuisance control and game hunting) of female deer has been larger than that of male deer (Fig. 2). We therefore assumed that in recent years, female deer have been sufficiently hunted. The allowable number of animals culled through nuisance control in area i depends on the governmental budget. In principle, the budget for nuisance control is interchangeable between each area, because the reward for such control is strongly supported by the government. The total number of animals culled through nuisance control in each management area is
where \(B\) is the total annual budget for nuisance control, μ is the allocation ratio of the budget for the eastern area (\(0 \le \mu \le 1\)), and \(P^{reward}\) is the reward paid to hunters for one deer. In this study, we set this reward to 5,000 Japanese yen (Hokkaido Government, unpublished data).
Simulation outline
The government of Hokkaido set targets of the CMPS4, which were measured by the population index obtained by spotlight counts (Kaji et al. 2010). In the eastern area, the target is between 25 and 50 % of the population index in 1993. In the western area, the target is under 200 % of the population index in 2000 (Hokkaido Government 2012). We used the limit of the target population \(N_{i}^{TN}\) (\(N_{\text{e}}^{TN} = 200,000\), \(N_{\text{w}}^{TN} = 110,000\)) that corresponds to the target population index.
To evaluate the feasibility of the CMPS4 targets, we simulated population by a stochastic matrix population model. We used several data sets in the stochastic simulation and parameter estimation (Table 1). Estimated parameter values were set for each area and category (Table 2). The simulation began in 2012, and the evaluated period was the same as that of CMPS4 (2013–2017). Uncertain parameters are initial population for 2012 (N 2012), female survival rate under natural conditions (S t ), reproduction rate (r t ), process error of game hunting effort (\(\varepsilon_{t,i,j}^{E}\)), and estimated population number (\(N_{t,i}^{EST}\)). The Hokkaido government estimated the deer population using Bayesian estimation, and we used posterior probability distribution for N 2012 (K. Yamamura et al., unpublished data). Using MATLAB R2009b software, we assumed four management scenarios and ran 1,000 trial simulations to calculate the management success probability and cost-effectiveness of the CMPS4.
Management scenario
In the stochastic simulation, we defined four alternative scenarios based on the fixed, limited management budget. This budget in scenario 1 is the average cost from 2004–2008 (about 147 million Japanese yen), calculated by the average number of animals culled through nuisance control and the reward per animal. The budget in scenario 2 is twice that of scenario 1. Similarly, scenarios 3 and 4 represent budgets threefold and fourfold that of scenario 1, respectively.
Rules for nuisance control
The government of Hokkaido designed an adaptive management program in which the number of animals to be culled through nuisance control is set according to predicted population indices (Kaji et al. 2010). Current management status is the emergent population phase in which control pressure is as high as possible (Hokkaido Government 2012). The number of animals allowed to be culled through nuisance control, however, depends on budgetary constraints. We therefore set two simple and realistic rules for nuisance control: (1) total management budget (\(B\)) does not change during the management period; and (2) when the estimated population number in area i in year t (\(N_{t,i}^{EST}\)) is below the upper limit of the target population (\(N_{i}^{TN}\)), the government halts nuisance control. After this halt, if \(N_{t,i}^{EST}\) exceeds \(N_{i}^{TN}\), the government re-implements nuisance control:
We did not know the actual population size. The estimated population size \(N_{t,i}^{EST}\) therefore includes estimation error
where \(\sum\nolimits_{j} {N_{t,i,j} }\) is total population in the area; \(\varepsilon_{t,i}^{N}\) is estimation error, which follows a lognormal distribution with mean 0 and variance \(\sigma_{N,i}^{2}\); \(- 0.5\sigma_{N,i}^{2}\) is a bias correction term for lognormal errors; and \(\sigma_{N,i}^{2}\) was calculated according to the 2012 estimated population in (\(\sigma_{{N , {\text{e}}}}^{2} = 0.043\), \(\sigma_{{N , {\text{w}}}}^{2} = 0.050\), K. Yamamura et al., unpublished data).
Success probability of the CMPS4
We assumed two successful situations for the CMPS4. First management success is assumed when the deer population at least one area (eastern or western) is less than the CMPS4 target for that area. The Second successful situation is when the eastern and western deer populations are simultaneously below the CMPS4 targets. According to these two situations, we calculated the probability of success of satisfying the target of at least one area (SP one ) and those of both areas (SP both ) simultaneously. We assumed that the probability of success in each management areas would vary with management strategy.
Cost-effectiveness of the CMPS4
There are many studies focusing on the costs of wildlife management and conservation (e.g., Nugent and Choquenot 2004; Hilborn et al. 2006; Ross and Pollett 2007; Yokomizo et al. 2007, 2009, 2014; Duca et al. 2009). To compare each management scenario, we analyzed cost-effectiveness (Macmillan et al. 1998; Drechsler et al. 2007), which can be used to find the least-cost means for meeting a conservation object or biological standard while tracking a single measure or effect (Hughey et al. 2003). In our management plan, the Hokkaido government may cut the management budget when the size of the deer population falls below the upper limit of the management target. Thus, total actual management cost (T cost) is given as the reward paid to hunters \(P^{reward}\) and the number of animals culled through nuisance control \(C_{t,i,j}\):
where \(\delta\) is the economic discount rate and t is the year with t = 0 in 2012. Economic discount rate is usually used in environmental economics (e.g., Yokomizo et al. 2004), and means that humans tend to perceive a future economic burden less than present expenditure. Consequently, the economic discount rate is important for analysis of long-term investigation. We assumed a present-value discount rate of 5 % annually.
Lindsey et al. (2005) defined a cost-efficiency index for wild dog conservation in South Africa. To evaluate cost-effectiveness in the present study, we defined a cost effectiveness index (CEI) that was a success probability (SP) per unit cost. In this study, SP one and SP both correspond to CEI one and CEI both , respectively. If the government does not fund the CMPS4, SP one is 0.4 % and SP both is 0 %. Therefore, we simply calculated CEI as success probability (SP one and SP both ) per T cost.
Results
Focusing on the probabilities of satisfying the target population number in at least one management area, SP one decreased with increasing migration rate (m) into that area under unbalanced budget-allocation management (Fig. 3). In scenarios 1 through 3, SP one increased with the allocated budget (Fig. 3a–c). In scenario 1, SP one was less than 20 % in every situation (Fig. 3a). In scenario 3, SP one maximized when the entire budget was allocated to one area, and it decreased as much as 25 % with increased m (Fig. 3c).

Success probabilities of meeting the target population number in at least one area (SP one ) for management scenarios 1–4 (a–d), with four conditions of migration rate m (0.000, 0.025, 0.050, and 0.075) and various eastern area budget allocation ratios μ (0–1 in steps of 0.05). The budgets of scenarios 1–4 are about 147, 293, 440, and 586 million Japanese yen
In scenario 1, SP both was less than 1 % for all migration rates and budget allocation ratios (Fig. 4a). However, in scenarios 2 through 4, SP both was high (scenario 2: 4–7 %; scenario 3: 26–34 %; scenario 4: 57–66 %) when the budget allocation ratio (μ) was 0.50–0.65 (Fig. 4). SP both in scenario 4 was much larger than that in other scenarios (Fig. 4). In scenarios 3 and 4, the maximum SP both did not differ remarkably with m (Fig. 4c, d).

Success probabilities of meeting the target population number in both management areas (SP both ) for management scenarios 1–4 (a–d), with four conditions of migration rate m (0.000, 0.025, 0.050, and 0.075) and various budget allocation ratios for the eastern area μ (0–1 in steps of 0.05). The budgets of scenarios 1–4 are about 147, 293, 440, and 586 million Japanese yen
Comparing T cost in different management scenarios, mean T cost was minimized when the entire budget was invested in the western area (Fig. 5). The maximum T cost in scenario 4 was approximately the same as the budget in scenario 3 (Fig. 5c, d). T cost was maximized when SP both maximized (Figs. 4, 5).

Predicted mean total cost during the management period 2013–2017 for scenarios 1– 4 (a–d), with four cases of migration rate m (0.000, 0.025, 0.050, and 0.075) and various budget allocation ratios for the eastern area μ (0–1 in steps of 0.05). We assumed a 5 % discount rate. Dashed line indicates the total management budget (B) for each scenario
CEI one maximized when the entire budget was invested in one area (scenario 1: 1.63–2.40 × 10−8; scenario 2: 4.92–7.89 × 10−8; scenario 3: 6.48–8.34 × 10−8; and scenario 4: 6.76–7.90 × 10−8) (Fig. 6). Maximum CEI one decreased with m under unbalanced budget-allocation management (Fig. 6). In scenarios 3 and 4, maximum CEI one values were approximately the same (Fig. 6c, d). CEI both increased with budget (scenario 1: 0.05–0.06 × 10−8; scenario 2: 0.32–0.51 × 10−8; scenario 3: 1.45–1.89 × 10−8; and scenario 4: 2.78–3.16 × 10−8) when μ values were 0.50–0.80 (Fig. 7).

Cost-effectiveness index for managing at least one area (CEI one ), calculated as the probability of success in at least one area divided by mean total cost, for scenarios 1–4 (a–d), with four cases of migration rate m (0.000, 0.025, 0.050, and 0.075) and various budget allocation ratios for the eastern area μ (0–1 in steps of 0.05). The budgets of scenarios 1–4 are about 147, 293, 440, and 586 million Japanese yen

Cost-effectiveness index for managing both areas (CEI both ), calculated as the probability of success in both areas divided by mean total cost, for scenarios 1–4 (a–d), with four cases of migration rate m (0.000, 0.025, 0.050, and 0.075) and various budget allocation ratios for the eastern area μ (0–1 in steps of 0.05). The budgets of scenarios 1–4 are about 147, 293, 440, and 586 million Japanese yen
Discussion
We explored four alternative management scenarios for the CMPS4. SP one and SP both were positively related to the total budget in scenarios 2 through 4 (Figs. 3, 4). Scenario 1 is the base case in our assessment, in which the total budget is taken as the average of the 2004–2008 budget. To increase SP one or SP both , the government must increase the budget above this average cost. Specifically, to satisfy the CMPS4 target in at least one management area, the Hokkaido government must budget more than 441 million yen (scenario 3), which should be invested in only one area (Fig. 3). To maximize SP both , the government requires over 586 million yen (scenario 4), and SP both will total about 60 % for any m (Fig. 4).
We performed stochastic simulations according to a simple management rule: when the population number satisfies the CMPS4 target, the government halts nuisance control. As a consequence, the CEI both increased with the budget (Fig. 7). This is intuitively easy to understand, because nuisance control is frequently halted, thereby removing the government’s need to fund it. Increasing the budget is a cost-effective way to satisfy the CMPS4 targets. The differences in maximum CEI one between scenarios 2, 3, and 4 were not large (Fig. 6). CEI one decreased with m (Fig. 6). These results suggest that allocating entire budgets to one area is cost-effective given small budgets, but that effectiveness will be less when migration rate m is high.
For the sake of simplicity, we assumed a fixed hunter reward for nuisance control, and the number of culled individuals was determined by the total budget. However, this reward has been altered in the past. Presumably, our model may need to incorporate the correlation between reward and effort invested in nuisance control, because a greater reward gives more incentive for nuisance control. This assumption would decrease implementation errors (Kotani et al. 2011) in the analysis.
In a previous study of deer management in Hokkaido (Yamamura et al. 2008), biological parameters (survival rate, reproduction rate, and sex ratio) were assumed equal between the eastern and western areas. We also assumed no bias in migration westward or eastward. However, in its 2013 population estimates, the Hokkaido government assumed natural mortality in the western area to be 30 % less than that in the eastern area, as indicated by different population indices estimated from spotlight counts in the Hidaka and Iburi regions (K. Yamamura et al., unpublished data). We can therefore consider several additional factors for empirical adjustment, such as observation error of population index, natural mortality, and biased migration. For example, we can define an area-dependent female survival rate under natural conditions (S t ). It is also possible that m depends on population density and habitat quality. Using an area-dependent S t and our matrix model, we can compute a future projection with assumptions similar to the population estimation in 2013, or estimate an appropriate m. However, we could not estimate m variation by year, owing to a lack of observation data regarding bias and historical change in m.
We also assumed that m was constant for each category, management area, and year. However, m may vary by category and year. Greenwood (1980) discussed the importance in promoting sex differences in mammal dispersal. For deer, Clutton-Brock et al. (2002) concluded that migration of male red deer from their natal areas increased with female density. The complex combination of m seen in sika deer would produce different results in our assessment. The m of female deer has important implications for population control, because with a focus on only one management area, the maximum probabilities differed directly by more than 25 % given different m (Fig. 3). In the future, we must therefore estimate migration between management areas by, for instance, radio tracking or mark-recapture, which have proven useful in understanding such deer migration in Hokkaido (e.g., Uno and Kaji 2000; Sakuragi et al. 2003; Coulson et al. 2004; Igota et al. 2004).
The Hokkaido government has begun implementing special management measures related to nuisance control for the period 2011–2015. In 2011, total harvests (via game hunting and nuisance control) in the eastern and western areas were about 69,000 and 60,000 deer, respectively (Fig. 2). However, during 2004–2008 these totals averaged 46,000 (eastern) and 29,000 (western) annually. If the number of deer harvested by game hunting in 2011 was equal to the average number harvested so during 2004–2008, the number of animals culled through nuisance control in 2011 would be 2.88 times larger than the average number culled so during 2004–2008. During these periods, the budget allocation ratio was approximately 0.50. Scenario 3, with μ = 0.5, corresponds to the present management situation, in which SP both was 25.7–29.5 % (Fig. 4c). Generally, culling a large number of individuals in the early management stage is more effective for reducing deer reproduction, because this number is proportional to the female deer population. When the government establishes a higher budget in the early management stage, actual costs in later management stages could be reduced. This is why T cost per budget declined as the management budget increased in our simulation (Fig. 5). These factors suggest that investing a larger budget in the early management stage is an efficient strategy for meeting the targets set by the CMPS4. We therefore recommend that the Hokkaido government invest a higher proportion of its current total budget in this stage, to manage the Hokkaido population of sika deer in a cost-efficient manner.
References
Baxter PWJ, McCarthy MA, Possingham HP, Menkhorst PW, McLean N (2006) Accounting for management costs in sensitivity analyses of matrix population models. Conserv Biol 20:893–905
Bhat MG, Huffaker RG (2007) Management of a transboundary wildlife population: a self–enforcing cooperative agreement with renegotiation and variable transfer payments. J Environ Econ Manag 53:54–67
Clutton-Brock TH, Coulson TN, Milner-Gulland EJ, Thomson D, Armstrong HM (2002) Sex differences in emigration and mortality affect optimal management of deer populations. Nature 415:633–637
Coulson T, Guinness F, Pemberton J, Clutton-Brock TH (2004) The demographic consequences of releasing a population of red deer from culling. Ecology 85:411–422
Drechsler M, Wätzold F, Johst K, Bergmann H, Settele J (2007) A model-based approach for designing cost-effective compensation payments for conservation of endangered species in real landscapes. Biol Conserv 140:174–186
Duca C, Yokomizo H, Marini MA, Possingham HP (2009) Cost-efficient conservation for the white-banded tanager (Neothraupis fasciata) in the Cerrado, central Brazil. Biol Conserv 142:563–574
Gordon IJ, Hester AJ, Festa-Bianchet M (2004) The management of wild large herbivores to meet economic, conservation and environmental objectives. J Appl Ecol 41:1021–1031
Greenwood PJ (1980) Mating systems, philopatry and dispersal in birds and mammals. Anim Behav 28:1140–1162
Haltuch MA, Punt AE, Dorn MW (2008) Evaluating alternative estimators of fishery management reference points. Fish Res 94:290–303
Hilborn R, Annala J, Holland DS (2006) The cost of overfishing and management strategies for new fisheries on slow–growing fish: orange roughy (Hoplostethus atlanticus) in New Zealand. Can J Fish Aquat Sci 63:2149–2153
Hokkaido Government (2000) Conservation and management plan for Sika deer (Cervus nippon) in Hokkaido. Hokkaido Government, Department of Health and Environment, Sapporo (in Japanese)
Hokkaido Government (2012) Conservation and management plan for Sika deer (Cervus nippon) in Hokkaido (4th stage). Hokkaido Government, Department of Health and Environment, Sapporo (in Japanese)
Hughey KFD, Cullen R, Moran E (2003) Integrating economics into priority setting and evaluation in conservation management. Conserv Biol 17:93–103
Hunter CM, Caswell H (2005) The use of the vec-permutation matrix in spatial matrix population models. Ecol Model 188:15–21
Igota H, Sakuragi M, Uno H, Kaji K, Kaneko M, Akamatsu R, Maekawa K (2004) Seasonal migration patterns of female sika deer in eastern Hokkaido, Japan. Ecol Res 19:169–178
Kaji K, Koizumi EJ, Ohtaishi N (1988) Effects of resource limitation on the physical and reproductive condition of sika deer on Nakashima Island, Hokkaido. Acta Theriol 33:187–208
Kaji K, Okada H, Yamanaka M, Matsuda H, Yabe T (2004) Irruption of a colonizing sika deer population. J Wildl Manag 68:889–899
Kaji K, Saitoh T, Uno H, Matsuda H, Yamamura K (2010) Adaptive management of sika deer populations in Hokkaido, Japan: theory and practice. Popul Ecol 52:373–387
Kotani K, Kakinaka M, Matsuda H (2011) Optimal invasive species management under multiple uncertainties. Math Biosci 233:32–46
Lindsey PA, Alexander R, Du Toit JT, Mills MGL (2005) The cost efficiency of wild dog conservation in South Africa. Conserv Biol 19:1205–1214
Macmillan DC, Harley D, Morrison R (1998) Cost–effectiveness analysis of woodland ecosystem restoration. Ecol Econ 27:313–324
Mantzouni I, Somarakis S, Moutopoulos DK, Kallianiotis A, Koutsikopoulos C (2007) Periodic, spatially structured matrix model for the study of anchovy (Engraulis encrasicolus) population dynamics in N Aegean Sea (E. mediterranean). Ecol Model 208:367–377
Matsuda H, Kaji K, Uno H, Hirakawa H, Saitoh T (1999) A management policy for sika deer based on sex-specific hunting. Popul Ecol 41:139–149
Matsuda H, Uno H, Tamada K, Kaji K, Saitoh T, Hirakawa H, Kurumada T, Fujimoto T (2002) Harvest–based estimation of population size for Sika deer on Hokkaido island, Japan. Wildl Soc Bull 30:1160–1171
Milner JM, Bonenfant C, Mysterud A, Gaillard JM, Stenseth NC, Csányi S (2006) Temporal and spatial development of red deer harvesting in Europe: biological and cultural factors. J Appl Ecol 43:721–743
Naidoo R, Balmford A, Ferraro PJ, Polasky S, Ricketts TH, Rouget M (2006) Integrating economic costs into conservation planning. Trends Ecol Evol 21:681–687
Nugent G, Choquenot D (2004) Comparing cost–effectiveness of commercial harvesting, state–funded culling, and recreational deer hunting in New Zealand. Wildl Soc Bull 32:481–492
Ou W, Takekawa S, Yamada T, Terada C, Uno H, Nagata J, Masuda R, Kaji K, Saitoh T (2014) Temporal change in the spatial genetic structure of a sika deer population with an expanding distribution range over a 15-year period. Popul Ecol 56:311–325
Ross JV, Pollett PK (2007) On costs and decisions in population management. Ecol Model 201:60–66
Sakuragi M, Igota H, Uno H, Kaji K, Kaneko M, Akamatsu R, Maekawa K (2003) Seasonal habitat selection of an expanding sika deer Cervus nippon population in eastern Hokkaido, Japan. Wildlife Biol 9:141–153
Sanchirico JN, Wilen JE (1999) Bioeconomics of spatial exploitation in a patchy environment. J Environ Econ Manag 37:129–150
Sanchirico JN, Wilen JE (2005) Optimal spatial management of renewable resources: matching policy scope to ecosystem scale. J Environ Econ Manag 50:23–46
Takatsuki S (1998) The twinning rate of sika deer, Cervus nippon, on Mt. Goyo, northern Japan. Mammal Study 23:103–107
Takatsuki S, Suzuki K, Suzuki I (1994) A mass-mortality of Sika deer on Kinkazan Island, northern Japan. Ecol Res 9:215–223
Tanaka S (1982) The management of a stock-fishery system by manipulating the catch quota based on the difference between present and target stock level. Bull Jpn Soc Sci Fish 48:1725–1729
Ueno M, Kaji K, Saitoh T (2010) Culling versus density effects in management of a deer population. J Wildl Manag 74:1472–1483
Uno H, Kaji K (2000) Seasonal movements of female sika deer in eastern Hokkaido, Japan. Mammal Study 25:49–57
Uno H, Yokoyama M, Takahashi M (1998) Winter mortality pattern of Sika deer (Cervus nippon yesoensis) in Akan National Park, Hokkaido. Honyurui Kagaku (Mamm Sci) 38:233–246 (in Japanese with English summary)
Uno H, Kaji K, Saitoh T, Matsuda H, Hirakawa H, Yamamura K, Tamada K (2006) Evaluation of relative density indices for sika deer in eastern Hokkaido, Japan. Ecol Res 21:624–632
Yamamura K, Matsuda H, Yokomizo H, Kaji K, Uno H, Tamada K, Kurumada T, Saitoh T, Hirakawa H (2008) Harvest-based Bayesian estimation of sika deer populations using state-space models. Popul Ecol 50:131–144
Yokomizo H, Haccou P, Iwasa Y (2004) Multiple-year optimization of conservation effort and monitoring effort for a fluctuating population. J Theor Biol 230:157–171
Yokomizo H, Haccou P, Iwasa Y (2007) Optimal conservation strategy in fluctuating environments with species interactions: resource-enhancement of the native species versus extermination of the alien species. J Theor Biol 244:46–58
Yokomizo H, Possingham HP, Thomas MB, Buckley YM (2009) Managing the impact of invasive species: the value of knowing the impact-density curve. Ecol Appl 19:376–386
Yokomizo H, Coutts SR, Possingham HP (2014) Decision science for effective management of populations subject to stochasticity and imperfect knowledge. Popul Ecol 56:41–53
Acknowledgments
This work was partly supported by Japan Society for the Promotion of Sciences grants to H.M. (22370009) and H.Y. (25281057) and a Grant-in-Aid from the Japan Society for the Promotion of Science Fellows (25–3477) to U.O.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Ijima, H., Fujimaki, A., Ohta, U. et al. Efficient management for the Hokkaido population of sika deer Cervus nippon in Japan: accounting for migration and management cost. Popul Ecol 57, 397–408 (2015). https://doi.org/10.1007/s10144-015-0478-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10144-015-0478-1