
Abstract
Changes in vegetation conditions are induced both by climatic and human factors. Knowledge about how changes in vegetation conditions relate to its degradation is often lacking, although needed to sustainably manage rangeland resources in drylands. Contributions made to develop land management options require assessing vegetation dynamics and degradation in Palapye, an agro-pastoral region of high economic and biodiversity importance in eastern Botswana. The Vegetation Degradation Index (VDI) was applied to 18-year (1986–2016) time series of annual Normalized Difference Vegetation Index (ANDVI) images to establish vegetation degradation levels, i.e. relatively undegraded, low degradation, medium degradation, and high degradation. Plant species metrics and biophysical variables were then examined at nine sites with the aim of explaining variations in the vegetation degradation levels. Approximately 95% of the study area experienced low degradation and 11% had significant negative trends in ANDVI. Species richness, diversity, and rainfall explained 49% of the variance in degradation levels. Species diversity and richness were highest in Moeng (a relatively undegraded site), whereas they were lowest in Lesenopole (a low degradation site). A probable reason being that Colophospermum mopane contributed 87% of individual trees and shrubs at Lesenopole. Species diversity and richness were also negatively associated with VDI. The non-occurrence of some species such as Sclerocarya birrea and Lonchocarpus capassa on highly degraded sites suggests a link between species composition and the level of degradation. This study combined the use of theremote sensing-based VDI with field-based plant species data for validating vegetation degradation levels in a dryland context.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Land degradation is one of the twenty-first century environmental issues at global, regional, and local scales (Orr et al. 2017). Land degradation is heightened in African drylands due to their sensitivity to changes in the natural and human systems (Rufino et al. 2013). Particularly vulnerable are the agro-pastoral systems of these dryland areas, partly due to the practice of subsistence livestock farming and rainfed agriculture by the majority of the people. Degradation of vegetation and soil, indicative of the state of the land, jeopardises the livelihood of poor smallholder households in agro-pastoral regions (Safriel 2007; Sallu et al. 2010). Ecosystems supporting vital ecological processes are also vulnerable to land degradation. The combined effects of land degradation and climate change exacerbate the vulnerability of social-ecological systems and undermine adaptation efforts in various contexts (Webb et al. 2017). Drylands amount to 43% of Africa’s land area and are projected to increase between 5 and 8% by 2080 (FAO 2017).
Vegetation degradation is a biological type of land degradation related to a reduction in biomass, diversity, and/or reduced nutritional value of vegetation for livestock and wildlife (Eswaran et al. 2001). Vegetation monitoring for rangeland management is necessary as vegetation plays an important regulatory role in both the biosphere and the atmosphere (Miao et al. 2013; Belayneh and Tessema 2017; Treydte et al. 2017). Earth observation (EO)-based vegetation indices are increasingly used for analysing and monitoring vegetation change as these are timely and cost-efficient. As ground-based data collection is expensive, it is conducted intermittently resulting in data gaps. Often, ground data points are not well distributed and inconsistency in data collection methods and definitions make comparison and replication difficult. Without remote sensing, we can rarely gain the historical perspective of vegetation change. Moreover, the availability of images for Africa is now sufficiently long enough for examining trends in time series (Vlek et al. 2010).
Although vegetation degradation is a major process of environmental change, its dynamics, drivers, and impacts in Botswana are not well understood except in the cattle-based systems. There is currently a very limited amount of research analysing changes in vegetation conditions and degradation outside the Kalahari rangelands of southwest Botswana. Much of what is known relate to bush encroachment whereby bush cover and density are increasing at the expense of grass production, particularly around cattle watering points (Reed et al. 2015; Dougill et al. 2016). Bush encroachment is associated with reduced species diversity and forage resources in cattle-based systems (Ward 2005; Thomas et al. 2018). Reed et al. (2015) examined the impacts of bush encroachment on ecosystem services under different land tenure types. Studies have also argued whether bush encroachment that provides ground cover should be considered as land degradation (Abel 1997; O’Connor et al. 2014; Stevens et al. 2016).
Over an 18-year period (1998–2016), this study assessed changes in vegetation conditions and related these to vegetation degradation in Palapye, an agro-pastoral region of high economic and biodiversity importance in eastern Botswana (Akinyemi and Mashame 2018). Knowledge about vegetation degradation is lacking in this context, hence the need to fill this gap. This study examined how plant species diversity and richness related to varying vegetation degradation levels using remote sensing-based vegetation indices. Most studies comparing species diversity in natural and degraded woodlands do so without explicitly establishing the degradation levels. This study examined how plant species metrics and biophysical variables relate to the measured degradation levels. By incorporating data from satellite image analysis with detailed field-based observation, mechanisms driving the assessed patterns and trends in vegetation degradation can be better deciphered and understood (Brandt et al. 2017). It is the first study relating quantitatively assessed vegetation degradation with species composition in this relatively unknown region. Questions answered relate to the degree of vegetation degradation and variables explaining variations in vegetation degradation levels were identified. This study contributes to the limited literature on EO use and provides empirical evidence of vegetation degradation in Botswana. Overall, it aims to improve our understanding of the drivers of vegetation degradation in agro-pastoral systems of Southern Africa.
Materials and methods
Description of the study location
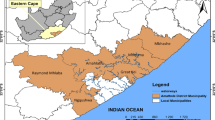
Our study area comprising the Palapye settlement with some 26 surrounding villages covers an area of 4479 km2 (Mashame and Akinyemi 2016), henceforth referred to as Palapye (Fig. 1). Administratively, the study location covers the jurisdiction of the Palapye Administrative Authority in the Botswana Central District. As a coal mining town, Palapye is the site of the only coal combustion power station in Botswana. Land is mostly under communal land tenure and used mainly for agriculture. Palapye is urbanising and tourism is an important economic sector (Akinyemi and Mashame 2018).

The study location. a Palapye in eastern Botswana. b Palapye settlement and the sites where vegetation survey was conducted
Palapye’s climate is semi-arid, hot steppe (Köppen’s BSh classification) as in most parts of Botswana. The summer months are October to March with temperature ranging from 32 to 39 °C, whereas winter is from April to September. Rainfall occurs during summer with an annual average of 351 mm over Palapye (Akinyemi 2017). Soils are mostly Arenosols and Luvisols and are predominantly sandy in texture (Kebonye et al. 2017). Palapye is dominated by low relief with the Tswapong Hills to the southeast and the Matepilepi Hills to the northwest. The Tswapong hills stretch over a 70-km range, 15 km wide, and rise up to 1380 m.
Data sources
Remote sensing data
Normalised Difference Vegetation Index (NDVI) is widely used to monitor vegetation greenness, plant phenology, and net primary productivity as induced by both natural and human factors (Vlek et al. 2010; Fensholt et al. 2013). Dekadal (10-daily composite) NDVI 1-km time series (July 1998 to June 2016) were analysed in this study, making 648 NDVI images. Datasets are accessible from the EUMETCast reception station of the Monitoring the Environment and Security for Africa SADC Thema (MESA-SADC) project at the Botswana International University of Science and Technology (BIUST). NDVI datasets from July 1998 to May 2014 are from the SPOT – VGT, whereas datasets from June 2014 to June 2016 are from the PROBA-V. These datasets are pre-processed at source to ensure the comparativeness of the long-term NDVI statistics and continuity of the time series derived from both sensors. Pre-processing includes atmospheric correction of the surface reflectance in the RED and NIR bands from the PROBA-V images to the SPOT – VGT. Adjustments were required because of minor sensor differences such as in their spectral response functions, overpass time, and radiometric calibration (Vito 2015). Sterckx et al. (2014) give an extensive description of the corrections applied to the PROBA-V images.
Field data
Tree and shrub species data were collected from nine sites representing vegetation degradation levels between 6 and 9 December 2016. This survey was needed to validate the vegetation degradation levels (this procedure is described in the subsections “Vegetation Degradation Index” and “Degradation classes”). Stratified random sampling was employed to ensure that each degradation level as a stratum had an equal chance of being sampled and that vegetation sampling plots are not poorly distributed to minimise over- or under-representing certain species. According to Noori et al. (2014), the choice of an appropriate sampling technique depends upon the type of data needed. Referring to Fig. 1, the nine sites are Mogapinyana and Moeng (relatively undegraded), Lesenepole and Mahibitswana (low degradation), Malaka, Maunatlala and BIUST (medium degradation), Lecheng and Morupule (high degradation). Coordinates of site centroids obtained in a GIS were entered in a Leica Zeno field Global Positioning System (GPS) receiver and located on the field. We established two quadrats (50 by 50 m) per site and each quadrat was further divided into two equal halves for ease of identifying and counting the species.
Methods
Annual NDVI difference
NDVI images for the Southern African Development Community (SADC) were clipped to the boundary of the study area for use in creating the NDVI difference maps in the Drought Management System (DSM) software. NDVI differencing is widely used as a technique to identify deviation (anomaly) in vegetation state for a certain period in comparison with the long-term average for that period, or between two periods (Rudy et al. 2013; Meroni et al. 2019). NDVI difference (NDVIdiff) was calculated annually for December in relation to the long-term average from 1998 to 2016 (Eq. 1). The 3rd dekad of December was selected as this period marks the peak of the growing season in the study area.
where i represents the year of interest, a is a specific 10-day period, i.e. a dekad, and n represents the total number of years in the entire series.
Vegetation Degradation Index
The extent of vegetation degradation is analysed with the Vegetation Degradation Index (VDI), a regression-based method. As a first step, the VDI requires establishing the linear relationship between ANDVI and time. We estimated the slope of the trend line fitted to each pixel using the ordinary least square method (Eq. 2) following Li et al. (2015).
Where i represents a particular year and n represents the total number of years in the entire series. ANDVIi is the annual NDVI for each year, composited as the maximum value of NDVI per pixel between the first dekad in July to the third dekad in June of the following year. The maximum measurable NDVI over a period indicates the vegetation greenness peak, which usually occurs during the growing season (Li et al. 2015). Afterwards, the VDI was evaluated by separating change in ANDVI from no change at the 0.05 significance level using the F-test (Eq. 3).
Any reduction in ANDVI is significant if the VDI is < 0 and F > F0.05 (Li et al. 2015). Therefore, if VDI is < 0, i.e. negative, there is a decreasing trend of ANDVI. These negative values were multiplied by − 100 to derive positive values, which were then categorised into degradation classes. Otherwise, ANDVI is not changed or an increasing trend (regeneration) if VDI is > 0. Pixels with an increasing trend were reclassified to zero as only negative changes in ANDVI are associated with a decreasing trend to depict degradation in vegetation.
Degradation classes
The VDI values formed the basis for classifying the entire study area into four vegetation degradation levels using the natural break in the data as follows: 0–1 = relatively undegraded, 1–5 = low degradation, 5–10 = medium degradation, and 10–25 = high degradation. These categories were adopted in order to largely correspond to the previous legend utilised in mapping land degradation in Palapye (Akinyemi 2015).
Plant species metrics
Species occurrence, richness, density, and diversity were computed at the quadrat level and their mean values were calculated for each site in the study area. For each species identified in a quadrat, the number of individuals was counted and recorded. Species occurrence in the entire study area was derived as the average of occurrences in all sites. Species richness (S) is the total number of species present in each site (Whittaker 1972). The density of each species is calculated by dividing the number of individual species by the area of the quadrat as in Eq. 4.
where Di is the density of species i, Ni is the number of occurrences of species i in a quadrat, and Qa is the area of the quadrat. This measure was used because each tree and shrub species could be consistently recognised and counted per quadrat. The density of individual species in metre squared (m2) was extrapolated to the hectare (ha). Species diversity was computed per site using the Shannon Diversity Index (SDI, Shannon 1948) as in Eq. 5.
where Pi is the fraction of the population of species i identified divided by the total number of species found, s is the number of species found, and LN is the natural log.
Spearman’s correlation and principal component analysis
Variables of interest in explaining the variations in degradation levels that were identified from the literature and field observation are species richness, SDI, NDVI, VDI, rainfall (total annual average), the Standardized Precipitation Index (SPI), elevation, and slope aspect. The SPI is a probability-based drought index that quantifies the deviation of the total rainfall from the mean for a certain period (Du et al. 2012). Rainfall and SPI (a proxy for drought) are major climatic factors affecting plant growth in semi-arid contexts. Spearman’s coefficient (r2) was used to correlate VDI with the ecologic, climatic, and terrain variables in our case study. Principal component analysis (PCA) was conducted for its utility in variable reduction into the underlying, uncorrelated components (PCs). PCA as a statistical method, with minimal information loss, is capable of reducing a large number of correlated variables into a smaller number of uncorrelated components (Silva-Flores et al. 2014). A varimax rotation was used to simplify the structure and interpretation was based on the variable with the highest loading on each PC (Barik et al. 2019). The aim is to understand how variables contributed to the degradation levels on the basis of their loadings in the PCs.
Results
Vegetation conditions
Depicted as NDVI anomaly maps, changes in vegetation conditions in Palapye between 1998 and 2016 reveal high temporal and spatial variability (Fig. 2). In relation to the long-term average which is the expected condition, vegetation performance improved in some years such as 1999–2000, 2010–2011, 2011–2012, 2013–2014, and 2014–2015, whereas it declined in other years such as 2001–2002, 2004–2005, 2005–2006, 2009–2010, 2012–2013, and 2015–2016.

NDVI anomaly from 1998 to 2016
Vegetation degradation
VDI values for the 18-year period (1998–2016) were mapped at the pixel level to depict varying vegetation degradation levels. 44.3% of the study area was classified as relatively undegraded, 51.3% as low degradation, 4.35% as medium degradation, and 0.05% as highly degraded (Fig. 3a, Supplementary Fig. 1). Figure 3b shows statistically significant negative trends in ANDVI (approximately 11% of the study area) with p values (F-test) ranging from 0.02 to 0.15, whereas Fig. 3c depicts VDI classes where trends are significant.

Vegetation degradation. a Vegetation Degradation Index (VDI) map, b significant negative trends in ANDVI by p value, and c by VDI
Areas classified as medium to highly degraded, such as in the north-west and south of Palapye, are mostly erosion sites with exposed rock outcrops and/or burrow pits. These areas tend to have negative trends signifying decreasing ANDVI (Fig. 3a). Only areas where these trends are significant are shown in Fig. 3b, whereas the VDI categories under which they fall are depicted in Fig. 3c. These findings are consistent with Akinyemi and Mashame (2018) which found a 26% loss of natural land cover in this region between 1986 and 2014. Savanna and shrubland losses were largely attributed to increasing rocky surfaces (48 to 426 km2) and grasslands (245 to 891 km2) as consequences of recurrent droughts and deforestation. Other factors are agriculture with croplands increasing from 71 to 221 km2 and settlement, i.e. artificial surfaces and built-up areas, increasing from 10 to 72 km2 over the 28-year period. As the region is also important for mining, there are environmental impacts. For example, coal mining and coal-fired electricity generation are ongoing at Morupule and clay brick manufacturing near Lecheng (Mashame and Akinyemi 2016, see Fig. 1).
Species composition and density
Thirty-three (33) tree and shrub species with a total of 3444 individuals occurred in the entire nine sites (Table 1). Species occurrence and average density in the study sites are shown in Table 1. The average densities of all species are further disaggregated according to the level of vegetation degradation at the nine sites (Table 2). The densest species isFootnote 1Colophospermum mopane (Mopane, Mophane) which occurred 1474 times (43%) on all nine sites at a rate of 527 per ha. Dichrostachys cinerea (Sickle bush, Moselesele) also occurred on all nine sites with 631 (16%) individuals at a rate of 269 per ha. Grewia flava (Grey/silver raisin, Moretwa) occurred on eight sites at a rate of 65 per ha, Ximenia Americana caffra (Blue sour plum, Moretologa) on seven sites at a rate of 89 per ha, Combretum apiculatum (Red bush willow, Mohudiri) on six sites at a rate of 94 per ha, Gymnosporia buxifolia (African spike thorn, Mothono) and Grewia monticola (Sand paper raisin, Mokgomphatha) on five sites at rates of 68 and 23 per ha respectively, Peltophorum africanum (African wattle, Mosetha) on three sites at 60 per ha, Sclerocarya birrea (Marula, Morula), and Lonchocarpus capassa (Apple leave rain tree, Mopororo) occurred only on the two relatively undegraded sites at rates of 16 and 10 per ha respectively.
The presence of the most occurring species at multiple sites is depicted in Fig. 4. To identify some factors influencing the different levels of vegetation degradation, the VDI was correlated with plant species (e.g. plant density, diversity) and biophysical variables at the nine sites representing degradation levels (Table 3).

Presence of the densest plant species on multiple sites
SDI (species diversity) and richness were highest in Moeng (relatively undegraded site), BIUST and Mahibitswana, which are both medium degradation sites, whereas these were lowest in Lesenepole (a low degradation site) and Lecheng (a high degradation site). SDI is significantly positively correlated with richness but both variables are negatively correlated with SPI. Rainfall also has a significant positive association with species richness but its positive relationship to species diversity was not significant. VDI has a positive relationship with slope aspect and SPI. Analysis from PCA showed the combined influence of variables as three components explained 85% of the total variance in vegetation degradation levels. PC1 accounted for 49% of the total variance with high positive loadings from species diversity, richness, and rainfall, whereas it had a very high negative loading from SPI. PC2 with high positive loading from NDVI accounted for 20% of total variance and PC3 accounted for 17% of total variance with high positive loadings from elevation (terrain) (Table 4).
Discussion
The use of the VDI in this study demonstrates a means for measuring and monitoring vegetation degradation based on freely available remote sensing vegetation indices time series. The method is quantitative making its replication in similar contexts feasible and the results reproducible.
Vegetation conditions
Vegetation indices are useful for assessing vegetation conditions despite their limitation in distinguishing between vegetation cover types (Karnieli et al. 2013). For example, bush encroachment is undesirable in cattle-based systems as it degrades the quality of rangelands for grazing but vegetation indices associate it with increasing trends of vegetation greening in drylands as it appears as an increase in vegetation production (Brandt et al. 2017). Assessing and interpreting degradation from images require in situ data for ground truthing. It is essential to interpret vegetation indices as proxies in the context of changing land uses and land management practices of an area (Reed et al. 2015). The VDI was validated in our context by relating plant species metrics and biophysical variables to vegetation degradation in nine sites representing four degradation levels.
High variability in vegetation performance over the 18-year study period reflects the non-equilibrium, dynamic nature of dryland ecosystems (Mogotsi et al. 2011; Dougill et al. 2016). Vegetation degradation in Palapye is attributable to the impacts of both climatic and anthropogenic factors. Vegetation growth decreased mostly during years of drought as confirmed by the drought and household security outlook report (Rural Development Council, RDC 2017). The report noted that vegetation growth declined in most parts of Botswana due to drought’s negative impacts on vegetation, whereas there was improvement when rainfall was abundant. For example, vegetation condition improved in 1999–2000 as the 649 mm rainfall recorded during this period was well above the long-term average of 351 mm (1960–2015) for Palapye (Akinyemi 2017). Studies such as Rutherford and Powrie (2013) examined the varying influence of rainfall gradients and grazing on changing species richness in Mopane woodlands in South Africa.
The negative climatic impacts on vegetation are further accentuated by heightened human activities in the region. Most of the places in the southern part of Palapye where vegetation degraded are areas on the footslopes of the Tswapong Hills. Apart from the environmental consequences of mining in the region, soil erosion is most pronounced in the cultivated and settled areas where the deep, freely drained and medium-textured Luvisols occur such as around Malaka (Paris et al. 1995).
Relating species composition and diversity to vegetation degradation levels
Mechanisms underlying vegetation degradation and its relationships with environmental and anthropogenic factors in dryland contexts are varied and complex (Waide et al. 1999; Isbell et al. 2011). With plant’s strong links to climatic and soil conditions as well as reflecting the impacts of human activities, they serve as useful indicators to infer rangeland degradation (Pärt and Söderström 1999; Fensholt et al. 2013; Treydte et al. 2017). It is noted that the VDI was calculated based on 18-year NDVI data, whereas the field-based plant species data was available only for 1 year. However, since plant species composition evolved over time as a consequence of the interaction between climatic and human factors at the various sites, it is assumed to be a reflection of the conditions captured in the NDVI time series. Notwithstanding, developing multi-temporal plant species data for future monitoring is essential.
Mopane, a very valuable economic tree species, occurred on all sites as it is well adapted to this region (Timberlake 1995). Findings relating decreasing species diversity and richness to degradation levels are in line with Bruschi et al. (2014) which found more plant species in undegraded sites in the Miombo woodlands of Mozambique. However, lower species diversity and richness in Lesenepole (a low degradation site) is probably attributable to the dominance of Mopane, which contributed 87% of individuals. Timberlake (1995, 1999) noted the comparatively low alpha and gamma diversities of mopane woodlands. Sclerocarya birrea and Lonchocarpus capassa did not occur in any of the degraded sites probably due to their vulnerability to overutilisation because these species have multiple, competing uses by rural communities (Muok et al. 2009 and Mudzengi et al. 2017). As a dioecious plant, Sclerocarya birrea is particularly vulnerable to soil degradation. Muok et al. (2009) found that changes in the biotic properties of soil such as decreasing density of the Arbuscular mycorrhizal fungi hampered the reestablishment of the plant. Thus, only land management options that improve the biotic soil properties should be adopted to prevent sites from degrading.
The result suggesting that vegetation degradation is linked to species composition and diversity in this region should be further explored as results from previous studies are mixed, particularly regarding the influence of varying levels of degradation. Rutherford and Powrie (2013) found that differences in the influence of land utilisation intensity on plant diversity, richness, and structure between commercial and communal rangelands in Mopane woodlands were negligible. Wen et al. (2013) examined how ecosystem services of alpine grasslands varied along the degradation gradient in China. They found that ecosystem services were lower in degraded grasslands in comparison with non-degraded ones. In other contexts, the effect of grazing, which often is implicated as driving land degradation, on plant composition and diversity were not always negative. Komac et al. (2014) found greater species diversity in areas of high grazing pressure in the eastern Pyrenees (Andorra) but with lower forage quality. Luo et al. (2018) found deterioration in soil conditions, vegetation cover but improved species diversity, and increased productivity in cushion plants and shrubs in extremely degraded areas. The foregoing could help to further explain why in our study area some medium degraded sites were high in species diversity. For example, the occurrence of Dichrostachys cinerea, an unpalatable species, on all sites is evidence of bush encroachment in Palapye. This species encroaches and dominates with increasing grazing intensity, while nutritious grass species disappear (Fensholt et al. 2013). This finding that bush encroachment is occurring in the region was confirmed by farmers in an earlier socio-economic survey conducted among land users in Palapye (Akinyemi 2017). Based on field observation, unpalatable species such as Datura stramonium L. (Jimsonweed, Mokhure) and Striga asiatica (Witchweed, Molelwana) were found growing on some croplands.
The relationship of slope aspect with VDI is positive and significant suggesting it has an influence on vegetation degradation. This is in line with the findings of Mohr (2004) and Nadal-Romero et al. (2014). Nadal-Romero et al. (2014) found that larger angles existing on south-facing slopes determined the extent of plant colonisation on Mediterranean badlands. That species diversity and richness are positively associated with rainfall, whereas they are negatively associated with drought agree with the findings of Liu et al. (2019). In contrast, VDI’s positive relationship with SPI implies that vegetation degrades under drought conditions due to moisture constraints.
Conclusion
The Vegetation Degradation Index (VDI) was applied to assess vegetation degradation in Palapye, an agro-pastoral region in eastern Botswana. The index was calculated by means of regression slopes from annual Normalized Difference Vegetation Index (ANDVI) images between 1998 and 2016. Based on the VDI values, four degradation classes were derived (relatively undegraded, low, medium, and high degradation). Approximately 95% of the study area experienced low degradation and 11% of the study area had significant negative trends in ANDVI. Much of the variation in vegetation degradation levels was explained by rainfall, species richness, and diversity. Species richness and diversity were found to be negatively associated with drought, whereas vegetation degradation was positively related to drought.
Of the 33 species found, Mopane and Dichrostachys cinerea were densest, occurring on all sites. Mopane as an economic tree is well adapted to the region, whereas Dichrostachys cinerea is an unpalatable species signifying bush encroachment. Results suggest a link between species composition and vegetation degradation levels. For example, Sclerocarya birrea and Lonchocarpus capassa did not occur in any of the degraded sites. This link should be further explored in order to decipher how vegetation degradation influences species composition, diversity, etc. within the context of the local land management practices. A better understanding of such influences could be of importance in developing suitable land management options in this and similar dryland regions.
Although the level of vegetation degradation was found to be relatively low in this region, it is essential to curtail the negativity of human activities on the environment as these have the potential to exacerbate negative climatic impacts, particularly from drought. Incorporating remote sensing and geostatistical modelling with plant species metrics enabled vegetation degradation assessment in an African dryland context. Its findings provide future research with input for identifying probable factors driving vegetation degradation in drylands in and beyond Africa.
Notes
When known, the plant’s scientific name, common name, and local name (underlined) are given.
References
Abel N (1997) Mis-measurement of the productivity and sustainability of African communal rangelands: a case study and some principles from Botswana. Ecol Econ 23:113–133
Akinyemi FO (2015) Land degradation in Palapye. Available online at http://www.arcgis.com/apps/MapSeries/index.html?appid=47b31ecd5930432fb4efa182b30608a0. Accessed 15 June 2019
Akinyemi FO (2017) Climate change and variability in semi-arid Palapye, eastern Botswana: an assessment from smallholder farmers’ perspective. Weather Clim Soc 9(3):349–365. https://doi.org/10.1175/WCAS-D-16-0040.1
Akinyemi FO, Mashame G (2018) Analysis of land change in the dryland agricultural landscapes of eastern Botswana. Land Use Policy 76:798–811. https://doi.org/10.1016/j.landusepol.2018.03.010
Barik SK, Bramha S, Behera D, Bastia TK, Cooper G, Rath P (2019) Ecological health assessment of a coastal ecosystem: case study of the largest brackish water lagoon of Asia. Mar Pollut Bull 138:352–363. https://doi.org/10.1016/j.marpolbul.2018.11.056
Belayneh A, Tessema ZK (2017) Mechanisms of bush encroachment and its inter-connection with rangeland degradation in semi-arid African ecosystems: a review. J Arid Land 9(2):299–312. https://doi.org/10.1007/s40333-016-0023-x
Brandt M, Tappan G, Diouf AA, Beye G, Mbow C, Fensholt R (2017) Woody vegetation die off and regeneration in response to rainfall variability in the West African Sahel. Remote Sens 9(1):39. https://doi.org/10.3390/rs9010039
Bruschi P, Mancini M, Mattioli E, Morganti M, Signorini MA (2014) Traditional uses of plants in a rural community of Mozambique and possible links with Miombo degradation and harvesting sustainability. J Ethnobiol Ethnomed 10:59. https://doi.org/10.1186/1746-4269-10-59
Dougill AJ, Akanyang L, Perkins JS, Eckardt FD, Stringer LC, Favretto N, Atlhopheng J, Mulale K (2016) Land use, rangeland degradation and ecological changes in the southern Kalahari, Botswana. Afr J Ecol 54(1):59–67. https://doi.org/10.1111/aje.12265
Du J, Fang J, Xu W, Shi P (2012) Analysis of dry/wet conditions using the standardized precipitation index and its potential usefulness for drought/flood monitoring in Hunan Province, China. Stoch Env Res Risk A 27(2):377–387. https://doi.org/10.1007/s00477-012-0589-6
Eswaran H, Lal R, Reich PF (2001) Land degradation: an overview. In: Bridges EM, Hannam ID, Oldeman LR, Pening de Vries FWT, Scherr SJ, Sompatpanit S (eds) Responses to land degradation, Proc. 2nd International Conference on Land Degradation and Desertification. Oxford Press, New Delhi
FAO (2017) Regional overview of food security and nutrition in Africa 2016. The challenges of building resilience to shocks and stresses. FAO, Accra, Ghana. Available online at http://www.fao.org/publications/rofsn-africa/en/ Accessed 07 December 2017
Fensholt R, Rasmussen K, Kaspersen P, Huber S, Horiton S, Swinner E (2013) Assessing land degradation/recovery in the African Sahel from long-term earth observation based productivity and precipitation relationships. Remote Sens 5(2):664–686. https://doi.org/10.3390/rs502066
Isbell F, Calcagno V, Hector A, Connolly J, Harpole WS, Reich PB, Scherer-Lorenzen M, Schmid B, Tilman D, van Ruijven J, Weigelt A, Wilsey BJ, Zavaleta ES, Loreau M (2011) High plant diversity is needed to maintain ecosystem services. Nature 477:199–202. https://doi.org/10.1038/nature10282
Karnieli A, Bayarjargal Y, Bayasgalan M, Mandakh B, Dugarjav C, Burgheimer J, Khudulmur S, Bazha SN, Gunin PD (2013) Do vegetation indices provide a reliable indication of vegetation degradation? A case study in the Mongolian pastures. Int J Remote Sens 34(17):6243–6262. https://doi.org/10.1080/01431161.2013.793865
Kebonye NM, Eze PN, Akinyemi FO (2017) Long-term treated wastewater impacts and source identification of heavy metals in semi-arid soils of Central Botswana. Geoderma Reg 10:200–214. https://doi.org/10.1016/j.geodrs.2017.08.001
Komac B, Domènech M, Fanlo R (2014) Effects of grazing on plant species diversity and pasture quality in subalpine grasslands in the eastern Pyrenees (Andorra): implications for conservation. J Nat Conserv 22(3):247–255. https://doi.org/10.1016/j.jnc.2014.01.005
Li YL, Shuang SC, Yue Y, Cheng Y (2015) Effects of urbanisation on vegetation degradation in the Yangtze River Delta of China: assessment based on SPOT-VGT NDVI. J Urban Plann Dev 141:05014026. https://doi.org/10.1061/(ASCE)UP.1943-5444.0000249
Liu J, Lindenmayer DB, Yang W, Ren Y, Campbell MJ, Wu C, Luo Y, Zhong L, Yu M (2019) Diversity and density patterns of large old trees in China. Sci Total Environ 655:255–262. https://doi.org/10.1016/j.scitotenv.2018.11.147
Luo J, Liu X, Yang J, Liu Y, Zhou J (2018) Variation in plant functional groups indicates land degradation on the Tibetan Plateau. Sci Rep 17606, 8(1). https://doi.org/10.1038/s41598-018-36028-5
Mashame G, Akinyemi FO (2016) Towards a remote sensing based assessment of land susceptibility to degradation: examining seasonal variation in land use-land cover modeling land degradation in a semi-arid context. ISPRS J. Photogramm. Remote Sens 3(8):137–144. https://doi.org/10.5194/isprs-annals-III-8-137-2016
Meroni M, Fasbender D, Rembold F, Atzberger C, Klisch A (2019) Near real-time vegetation anomaly detection with MODIS NDVI: timeliness vs. accuracy and effect of anomaly computation options. Remote Sens Environ 221(1):508–521. https://doi.org/10.1016/j.rse.2018.11.041
Miao L, Luan Y, Luo X, Liu Q, Moore JC, Nath R, He B, Zhu F, Cui X (2013) Analysis of the phenology in the Mongolian Plateau by inter-comparison of global vegetation datasets. Remote Sens 5:5193–5208. https://doi.org/10.3390/rs5105193
Mogotsi K, Nyangito MM, Nyariki DM (2011) Drought management strategies among agro-pastoral communities in non-equilibrium Kalahari ecosystems. Environ Res J 5(4):156–162. https://doi.org/10.3923/erj.2011.156.162
Mohr D (2004) Soil degradation in simple oak coppice forests of the Ahr-Eifel: implications for forest management derived from soil ecological studies. Cuvillier Verlag, Göttingen
Muok BO, Matsumura A, Ishii T, Odee DW (2009) The effect of intercropping Sclerocarya birrea (A. Rich.) Hochst, millet and corn in the presence of arbuscular mycorrhizal fungi. Afr J Biotechnol 8(5):807–812
Mudzengi CP, Murwira A, Zengeya FM, Murungweni C, González-Redondo P (2017) Screening key browse species in a semi-arid rangeland. Cogent Food Agric 3(1). https://doi.org/10.1080/23311932.2017.1285854
Nadal-Romero E, Petrlic K, Verachtert E, Bochet E, Poesen J (2014) Effects of slope angle and aspect on plant cover and species richness in a humid Mediterranean badland. Earth Surf Process Landf 39:1705–1716. https://doi.org/10.1002/esp.3549
Noori F, Gholinejad B, Jonaidi H (2014) Comparing efficiency of Whittaker and Stoglern design for measuring species richness. Bull Environ Pharmacol Life Sci 3(3):69–74
O’Connor TG, Puttick JR, Hoffman MT (2014) Bush encroachment in southern Africa: changes and causes. Afr J Range Forage Sci 31:67–88. https://doi.org/10.2989/10220119.2014.939996
Orr BJ, Cowie AL, Castillo Sanchez VM, Chasek P, Crossman ND, Erlewein A, Louwagie G, Maron M, Metternicht GI, Minelli S, Tengberg AE, Walter S, Welton S (2017) Scientific conceptual framework for land degradation neutrality: a report of the Science-Policy Interface. United Nations Convention to Combat Desertification (UNCCD), Bonn, German. Available online at http://www2.unccd.int/sites/default/files/documents/2017-08/LDN_CF_report_web-english.pdf Accessed 04 January 2018
Paris S, Moahi TS, Venema JH. 1995. Proposed agricultural land use plan for Ratholo agricultural extension area, Central region. FAO/UNDP/Government of Botswana Project BOT/91/001 Land use planning for sustainable agricultural development. field document 3. 88pp. Available online at http://www.fao.org/docrep/field/009/ar784e/ar784e.pdf Accessed 04 August 2017
Pärt T, Söderström B (1999) Conservation value of semi-natural pastures in Sweden: contrasting botanical and avian measures. Conserv Biol 13:755–765
Reed MS, Stringer LC, Dougill AJ, Perkins JS, Atlhopheng JR, Mulale K, Favretto N (2015) Reorienting land degradation towards sustainable land management: linking sustainable livelihoods with ecosystem services in rangeland systems. J Environ Manag 151:472–485. https://doi.org/10.1016/j.jenvman.2014.11.010
Rudy A, Lamoureux SF, Treitz P, Collingwood A (2013) Identifying permafrost slope disturbance using multi-temporal optical satellite images and change detection techniques. Cold Reg Sci Technol 88:37–49. https://doi.org/10.1016/j.coldregions.2012.12.008
Rufino MC, Thornton PK, Ng’ang’a SK, Mutie I, Jones PG, van Wijk MT, Herrero M (2013) Transitions in agro-pastoralist systems of East Africa: impacts on food security and poverty. Agric Ecosyst Environ 179:215–230. https://doi.org/10.1016/j.agee.2013.08.019
Rutherford MC, Powrie LW (2013) Impacts of heavy grazing on plant species richness: a comparison across rangeland biomes of South Africa. S Afr J Bot 87:146–156. https://doi.org/10.1016/j.sajb.2013.03.020
Rural Development Council (2017) Drought and household food security outlook 2016/17. Ministry of Local Government and Rural Development, Gaborone
Safriel UN (2007) The assessment of global trends in land degradation. In: Sivakumar MVK, Ndiang’ui N (eds) Climate and land degradation. Springer Verlag, Berlin, pp 1–38
Sallu SM, Twyman C, Stringer LC (2010) Resilient or vulnerable livelihoods? Assessing livelihood dynamics and trajectories in rural Botswana. Ecol Soc 15(4):3 http://www.ecologyandsociety.org/vol15/iss4/art3/
Shannon CE (1948) A mathematical theory of communication. Bell Syst Tech J 27(3):379–423 623–656
Silva-Flores R, Perez-Verdin G, Wehenkel C (2014) Patterns of tree species diversity in relation to climatic factors on the Sierra Madre Occidental, Mexico. PLoS One 9(8):e105042. https://doi.org/10.1371/journal.pone.0105034
Sterckx S, Benhadj I, Duhoux G, Livens S, Dierckx W, Goor E, Adriaensen S, Heyns W, Van Hoof K, Strackx G, Nackaerts K, Reusen I, Van Achteren T, Dries J, Van Roey T, Mellab K, Duca R, Zender J (2014) The PROBA-V mission: image processing and calibration. Int J Remote Sens 35(7):2565–2588. https://doi.org/10.1080/01431161.2014.883094
Stevens N, Erasmus BFN, Archibald S, Bond WJ (2016) Woody encroachment over 70 years in South African savannahs: overgrazing, global change or extinction aftershock? Philos Trans R Soc Lond Ser B Biol Sci 371(1703):20150437. https://doi.org/10.1098/rstb.2015.0437
Thomas AD, Elliott DR, Dougill AJ, Stringer LC, Hoon SR, Sen R (2018) The influence of trees, shrubs, and grasses on microclimate, soil carbon, nitrogen, and CO2 efflux: potential implications of shrub encroachment for Kalahari rangelands. https://doi.org/10.1002/ldr.2918
Timberlake J (1995) Colophospermum mopane: annotated bibliography and review. Zimbabwe Bulletin of Forestry Research No.11. Forestry Commission Zimbabwe, Harare, 49 p. ISBN 0797414207, 9780797414204.
Timberlake J (1999) Colophospermum mopane: an overview of current knowledge. In: Timberlake J, Kativu S (eds) African plants. Biodiversity, taxonomy and uses, Proceedings 1997 AETFAT Congress, Zimbabwe. Royal Botanic Gardens, Kew, pp 565–571
Treydte AC, Schmiedgen A, Berhane G, Tarekegn KD (2017) Rangeland forage availability and management in times of drought – a case study of pastoralists in Afar, Ethiopia. J Arid Environ 139:67–75. https://doi.org/10.1016/jaridenv.2016.12.014
Vito (2015) Operation of the Global Land Component: algorithm theoretical basis document. Issue I2.11 GIO-GL Lot1 consortium. Available online at https://land.copernicus.eu/global/sites/cgls.vito.be/files/products/GIOGL1_ATBD_PROBA2VGT_I1.10.pdf Accessed 24 January 2019
Vlek PLG, Le QB, Tamene L (2010) Assessment of land degradation, its possible causes and threat to food security in Sub-Saharan Africa. In: Lal R, Stewart BA (eds) Food security and soil quality. CRC Press, New York, pp 57–86
Ward D (2005) Do we understand the causes of bush encroachment in African savannas? Afr J Range Forage Sci 22:101–105
Waide RB, Willing MR, Steiner CF, Mittelbach G, Gough L, Dodson SI, Juday GP, Parmenter R (1999) The relationship between productivity and species richness. Annu Rev Ecol Syst 30:257–300
Webb NP, Marshall NA, Stringer LC, Reed MS, Chappell A, Herrick JE (2017) Land degradation and climate change: building climate resilience in agriculture. Front Ecol Environ 15(8):450–459. https://doi.org/10.1002/fee.1530
Wen L, Dong S, Li Y, Li X, Shi J, Wang Y, Ma Y (2013) Effect of degradation intensity on grassland ecosystem services in the alpine region of Qinghai-Tibetan Plateau, China. PLoS One 8(3):e58432. https://doi.org/10.1371/journal.pone.0058432
Whittaker RH (1972) Evolution and measurement of species diversity. Taxon 21:213–251
Acknowledgements
We are grateful to MESA-SADC thema for the provision of NDVI images and the DSM software used for analysis. We thank Prof. Gizaw Tsidu for assistance with F-test, Ndiye Kebonye for field assistance, and Dr. Lijuan Miao and Dr. Peter Eze for corrections on initial drafts. Anonymous reviewers provided constructive feedback that improved the paper’s quality.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Editor: Xiangzheng Deng
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Supplementary Figure 1
VDI validation sites showing vegetation type and cover (PNG 6747 kb)
Rights and permissions
About this article

Cite this article
Akinyemi, F.O., Kgomo, M.O. Vegetation dynamics in African drylands: an assessment based on the Vegetation Degradation Index in an agro-pastoral region of Botswana. Reg Environ Change 19, 2027–2039 (2019). https://doi.org/10.1007/s10113-019-01541-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10113-019-01541-4