
Abstract
Worldwide, there are at least 12 ILTER sites with an emphasis on karst, landforms arising from the combination of high rock solubility and well-developed solutional channel porosity underground, but the study of cave ecosystems has been largely neglected. Only two ILTER sites, both in Slovenia, are primarily caves. Caves are under-represented for several reasons, but especially because of the overall difficulty of access and the lack of a clear research agenda for cave ecosystem studies. We review several aspects of long-term studies in Postojna Planina Cave System (PPCS), proposing our approach as a model for ILTER research in caves. In PPCS, analysis of short-term temperature data shows a muted daily cycle and seasonality, and analysis of long-term temperature data shows an increase, largely the result of climate change. Changes in drip rate of epikarst aquifers above the cave are correlated with rainfall but with lags and complications resulting from differences in longer term rainfall patterns. Analysis of discharge rates indicates a rapid response to precipitation not only in the Pivka River at its sinking, but also at Unica Spring, where discharge is augmented from other parts of the aquifer, including epikarst. Quantitative analysis of the obligate epikarst-dwelling copepod community shows that, unlike most cave communities, complete sampling of the fauna is possible. Finally, organic carbon levels in PPCS indicate likely carbon limitation in the system. These five factors (temperature, drip rate, river discharge, epikarst copepod fauna, and organic carbon) are the appropriate variables for capturing the essential long-term trends in cave ecosystems and their causes.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Background
Caves are widely recognized as a common feature of the Earth’s landscape. Caves occur in a wide variety of geologic settings, including limestone, lava, quartzite, and even iron deposits (Auler 2012; Palmer 2007; Parker et al. 2013), but the vast majority of caves occur in soluble sedimentary rocks in karst landscapes. The term “karst” has its origins as a particular type of landscape found in the Dinaric Mountains of Slovenia and the northeast corner of Italy. As a geologic term, it refers to both a terrain and a process (Klimchouk 2015). For example, Monroe (1972) defines karst as “a terrain, generally underlain by limestone or dolomite, in which the topography is chiefly formed by the dissolving of rock, and which may be characterized by sinkholes, sinking streams, closed depressions, subterranean drainage, and caves.” Russian researchers have used a more process-oriented definition, such as Sokolov’s 1962 definition (in Klimchouk 2015) as “a process of destruction and obliteration of permeable soluble rocks mainly through their leaching by moving water”. In an essay (Worthington et al. 2017), five definitions of karst aquifers found in the literature are compared and discussed. They are based upon the presence of solution channel networks, hydraulic conductivities > 10–6 m/s, karst landscapes, channels with turbulent flow, and caves. By these and other definitions, caves are part, but not necessarily a critical part, of the karst landscape.
Karst landscapes are relatively well represented among the International Long Term Ecological Research (ILTER) sites including at least 12 of 1030 sites (ILTER 2018): Alta Murgin Point (Italy); Baget (France); Cerknica Lake (Slovenia); Fontaine de Vaucluse (France); Huanjiang (China); Jurassic (Dorvan) karst (France); Koliaris (Greece); Larzac (France); MEDYCYSS (France); Puding Karst Ecosystem (China); Sumaria/White Mountains (Greece); and Zöbelboden (Austria).
The focus is on karst, not caves, especially the subterranean drainage basin. Rouch (1977) championed the appropriateness of the karst basin as the unit of analysis in his work in the Baget basin in southern France, now an ILTER site. In other sites, the focus of research has been on the aboveground system, rather than the subterranean system (e.g., Gaberščik 2003). In the USA, karst terrains are almost completely absent from ILTER sites. The only exceptions are the Konza Prairie (Kansas) and the Florida Coastal Everglades, where the focus is on the prairie and the swamp, not the underlying rock.
There are only two sites globally where the focus is on a cave or cave system—both in Slovenia. Postojna Planina Cave System (PPCS), the birthplace of biospeleology (Romero 2009), has a long history of biological study, dating back to the early nineteenth century. Škocjan caves are better known for their spectacular hydrological features, including a large underground channel of the Reka River. Biological research in the cave has focused on the epikarst fauna (Pipan 2005). It is also a UNESCO World Heritage site.
There is no doubt that a karst basin/terrain approach to ecosystem studies has been a fruitful one, especially Rouch’s long-term study of Baget, with more than 20 papers (Danielopol and Rouch 1991), and Gibert’s extensive study of the Dorvan karst basin in the Jura Mountains (Gibert 1986, see also Simon 2013). Caves can also be a useful unit of analysis, as shown by Simon and his colleagues (Simon and Benfield 2001, 2002; Simon et al. 2003, 2007, 2010). Furthermore, Klimchouk (2015) argues, from a geological perspective, that formation of caves (speleogenesis) is the primary process in the formation of karst. Because of the emerging ideas that initiation of cave development is at depth (hypogene) and unconnected with surface features, surface features of karst are not central. In his view, speleogenesis is essentially a coupled mass-transfer/mass-transport process that depends on both the aggressiveness of groundwater and its flow.
According to the speleological definition of karst aquifers (Worthington et al. 2017), the presence of caves formed by dissolution is a diagnostic feature of karst aquifers, and the presence of extensive caves with underground streams is generally considered to characterize a well-developed karst aquifer (Ford and Williams 2007; Kresic 2013). Although caves are only small pieces of a larger master drainage network, important information about the characteristics of percolation (Hartmann and Baker 2017; Kogovšek 2010; Lange et al. 2010) and conduit flow (Genthon et al. 2005; Kaufmann et al. 2016) can be obtained by hydrological and hydrochemical monitoring in the caves. Comparison with precipitation and spring discharge data improves the general understanding of flow processes within karst aquifers (Mayaud et al. 2013; Worthington and Soley 2017).
Most of the existing ecological work on the subsurface of karst is at the scale of caves, even though it is recognized that there are other components, particularly epikarst (Pipan and Culver 2013; Williams 2008). There is also a burgeoning literature on band thickness and stable isotopic analysis of oxygen in stalagmites and other speleothems as a proxy for paleo-climate (Fairchild and Baker 2012). In part, stalagmites are useful climate proxies because cave temperatures are approximately the mean annual temperature of the surface (Covington and Perne 2015). Hydrological variability is one of many uncertainties in paleo-climatic reconstruction with speleothems that should be considered thoroughly (Hartmann and Baker 2017; Lange et al. 2010).
Nevertheless, long-term ecological monitoring in caves is uncommon, aside from speleothem analysis, which requires only a single visit to a cave to obtain a sample. This deficiency has several causes. Caves are usually difficult to access, certainly more so than surface habitats. As Engel (2015) points out, caves are extreme environments, not necessarily in the sense of average conditions, but in the possibilities of flooding and other extreme events. Fluctuating water levels and high relative humidity make for a challenging monitoring environment. Finally, it has not always been apparent exactly what new information would be gained by monitoring in caves.
In this contribution, we propose that the measurement of five factors is both practical and appropriate for the analysis of cave systems: temperature, epikarst discharge, discharge rates from exiting cave water, epikarst fauna, and organic carbon. First, we make the arguments that these are the appropriate parameters, outline their measurement in caves, and review results from one ILTER cave site in Slovenia–Postojna Planina cave system.
Ecosystem parameters
Temperature is a key parameter for environmental fluctuation in general in caves (Badino 2010). With the advent of small, waterproof data loggers, thousands of measurements of temperature over the course of time are feasible (see Pipan et al. 2011). Temperature is controlled by the average temperature of the fluids that flow through the cave, both air and water, and is equilibrated to the average temperature at the surface at the same altitude (Badino 2010; Covington and Perne 2015). Temperature throughout the day and year is not constant. Shallow caves have a daily fluctuation (Pipan et al. 2011), and even deep, isolated caves can have a yearly cycle of oscillation (Cigna 2002). Such oscillations are proxies for changes in other parameters, and offer the possibility that organisms limited to caves can detect daily and annual cycles. As an example, the persistence of circadian clocks in some but not all cave dwellers is an active and puzzling topic of research (see Friedrich 2013), and the presence of daily cycles in caves makes their persistence easier to explain. Temperature records also can document climate change over the past several decades (Domínguez-Villar et al. 2015; Šebela et al. 2015), and long-term stable isotope proxies of temperature from speleothems provide millennial records of temperature (Fairchild and Baker 2012).
While water enters caves through sinking streams, not all caves have sinking streams. However, except for unusual situations, water enters all caves in soluble rock as vertically percolating water as well. Dissolved carbon present in percolating water forms the basis for biofilms and in turn is the major carbon source for the animal community living in caves (Simon et al. 2003, 2007). Common sites of percolating water are ceiling drips, some of which are associated with stalactites. Drip rates are correlated with rainfall but with considerable local variation, such that two nearby drips may show very different patterns of discharge (Kogovšek 2010; Kogovšek and Urbanc 2007). Drip rate is relatively easy to measure, using automated infrared sensors (McDonald and Drysdale 2007) and tipping rainfall gauges.
Discharge (output) from a cave, from both sinking streams and percolating water, can typically be measured at the resurgence of the stream to the surface, by gauging stations, weirs, and the like (Kresic and Stevanovic 2010). It is ecologically important as a summary of hydrological inputs and a proxy for flooding events.
Hydrological features and ecosystem features are linked by the dynamics of organic carbon (Simon et al. 2007). Caves are likely to be carbon limited (Simon and Benfield 2002; Venarsky et al. 2014), and its measurement has proven to give important insights into not only sources, but also utilization and temporal variation (Simon 2013; Simon et al. 2007). Sources of organic carbon in caves include chemoautotrophy, percolating water, flowing water, wind and gravity, and the active movement of animals (Culver and Pipan 2009). Of these, percolating water and flowing water are the most important carbon sources for aquatic cave ecosystems. Chemoautotrophy is limited to a few caves (Engel 2012), and the others are only important in terrestrial systems. In a recent symposium on “Carbon and Boundaries in Karst,” only one paper was devoted to terrestrial habitats (Novak et al. 2013). Measurement of organic carbon typically requires automated measurement using the persulfate digestion method (Rice et al. 2017).
One of the most vexing problems in monitoring caves is a quantitative and comparable census of the cave fauna. The macroscopic fauna, both aquatic and terrestrial, is often rare and difficult to find, making even a complete list of species problematic (see Zagmajster et al. 2008). Pitfall traps, both baited and unbaited, can be used as a quantitative sample (Schneider et al. 2011), but they differentially attract species and may have a negative impact on species abundance. A more unbiased and less destructive method is to sample the aquatic fauna that exits the epikarst via ceiling drips. The technique involves continuous filtration of drip water, with retention of the organisms in the sampling device, and periodic collecting of the individuals (Pipan 2005). If five or more drips are sampled for a year, with monthly removal of specimens, nearly all species are found, according to analyses that estimate missing species (Pipan and Culver 2007). The epikarst fauna, especially copepods, is a major component of species richness in many caves (Pipan and Culver 2013). Furthermore, the geographic pattern of epikarst species richness is at least roughly concordant with patterns of other components of the cave fauna.
Methods and materials
Study site
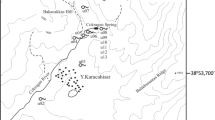
The Postojna Planina Cave System (PPCS) consists of two caves and their confluence. Postojna Cave, more than 24 km long, is hydrologically connected to Planina Cave, more than 6 km long, and the streams of the two caves merge and emerge at Unica Spring in Planinsko polje (Fig. 1). In Planina Cave, this spring gets additional recharge from the karst massif and from the Rak River which sinks in the area of Rakov Škocjan. The average flow of the Pivka River at its swallow point in Postojna Cave is 5.26 m s−1 (Šebela 2012), and the average flow of the Rak River at its swallow point is 11.68 m s−1 (ARSO 2017). During the late 1950s, the Pivka River carried a heavy load of organic pollutants into Postojna Cave, reducing oxygen concentrations by 50% up to 1 km into the cave (Sket and Velkovrh 1981). Major sources of organic pollutants, especially from the village of Pivka, disappeared by the 1980s. Postojna Cave has a long history as a commercial cave, and first opened to visitors in 1819, with a total of more than 37 million visitors. Currently, between 500,000 and 630,000 tourists visit the cave annually (Šebela et al. 2015). However, the commercial portion of the cave is less than 5 km, including a train that carries visitors into the cave. The land above the cave is forested, with various owners, and was largely pasture in the 1800s (Šebela 2012). Biologically, PPCS has more obligate cave dwelling terrestrial species (troglobionts) and more obligate cave dwelling aquatic species (stygobionts) than any other cave in the world (Culver and Pipan 2013).

Map of Postojna Planina Cave System. Temperature sampling points (Postojna 2 and Razpotje), hydrological sampling points (Unica and Pivka), epikarst hydrology (Kristalni rov), and epikarst fauna sampling area (Postojnska jama, Pivka jama, and Črna jama) are shown. Organic carbon was sampled at most of the above points as well
PPCS has a long history of monitoring and study, both of biological and physico-chemical parameters. Among physico-chemical parameters, these include a 20-year-long study of percolating water in the Kristalni Rov passage (Kogovšek 2010; Kogovšek and Šebela 2004), detailed temperature data from the 1930s (Crestani and Anelli 1939) and 2000s (Šebela and Turk 2011), and even earth tides since 1932 (Šebela 2012). Discharges of the Pivka River and Unica spring were regularly measured in the past by the Environmental Agency of the Republic Slovenia, but in the recent years, only measurements of water level at the Pivka River have been carried out (ARSO 2017). At several points within PPCS, detailed measurements of water level and discharges, temperature, and conductivity were done in the last decade (Kaufmann et al. 2016; Petrič 2011; Turk 2010). Biological parameters include species lists (Sket 1979), as well as several decades of monitoring of the terrestrial and aquatic fauna of Planina Cave by the SubBio Lab of the Department of Biology of the University of Ljubljana. The epikarst fauna was collected since 2000 (Culver and Pipan 2013; Pipan 2005; Pipan et al. 2006).
Temperature
Air temperature was measured at two cave locations (Lepe Jame-Postojna 2 and Razpotje) every hour with Baro-Diver® (Schlumberger Water Services) data loggers with an accuracy of ± 0.1 °C and resolution of 0.01 °C. To compare cave air temperature with outside cave conditions, additional air temperature measurements were made in the forest above Postojna Cave. The Postojna 2 monitoring site was in a narrow side passage about 15 m distant from the main tourist passage, Lepe Jame (Fig. 1). The Razpotje monitoring site was located at the intersection of the Male Jame and Stara Jama passages (Fig. 1). The Razpotje site was selected because temperature measurements were made at the same place from October 1933 to December 1937 (Crestani and Anelli 1939). Spectral analysis was done on the 2000’s data to detect 24-h cycles in the modern data. Spectral analysis as one of statistical techniques was used for characterizing and analyzing sequenced air temperature data. The spectral analysis aims to identify the seasonal or daily peaks in time series using some spectrum estimator.
Discharge of percolation water
Kogovšek (2010) did a detailed analysis of three drips in Postojna Planina Cave System over several decades, including drip rate and precipitation for the period from August 2005 to July 2006. Drip rate was measured by an evaporimeter and mechanical clock. A container with a siphon was attached to the drip. A larger container was used to collect the water each time it flowed from the siphon. Precipitation was measured on the surface above the study area at edge of the Postojna military barracks with a rain gauge fitted with an Onset™ Hobo Event Logger RG2-M, which recorded every 0.2 mm of precipitation. The drip was located in Kristalni rov.
Discharge of springs
Discharge rates, measured since 2007 (Petrič 2011; Turk 2010), were computed for the sinking point and the spring based on the stage-discharge curves. At the swallow point of the Pivka River water, levels were measured by Logotronic Gealog S and at the Unica spring by Onset™ Hobo Water Level Logger. An Onset™ Hobo Event Logger RG2-M was used for measurement of precipitation. Data were collected at 30-min intervals. The stage-discharge curve for the Pivka River was defined based on several parallel measurements of discharge with dilution method (Turk 2010) and for Unica it was prepared by the Environmental Agency of the Republic of Slovenia.
Organic carbon
Filtered water samples were collected (with three replicates) from a sinking of the Pivka River, two cave streams (the underground Pivka River and the underground Rak River), 33 epikarst drips, and the resurgence of the cave streams at the Unica River (Simon et al. 2007). Dissolved organic carbon (DOC) was determined using the persulfate digestion method on an OI Analytical Total Organic Carbon Analyzer Model 1010.
Epikarst fauna
Epikarst drip sampling devices (Pipan 2005) continuously sampled drips, by filtering drip water, and retaining the organisms in the sampling device until they are removed. These devices were deployed in 20 drips, 10 is Postojnska jama, five in Črna jama, and five in Pivka jama (Fig. 1). Using species accumulation curves and missing species estimators (e.g., abundance based Chao measures), it is possible to determine sampling completeness. In particular, we used Mao-tau procedures for species abundance curves and Chao’s abundance-based estimate (Colwell 2013):
where Sobs is the observed number of species, F1 is the number of species known from a single individual (singleton), and F2 is the number of species known from two individuals (doubleton). It was possible to analyze species richness patterns at a series of scales (Pipan and Culver 2007):
-
1.
Individual drips, with replicates consisting of bimonthly sampling periods;
-
2.
Individual sections of Postojna Planina Cave System, with replicates consisting of individual drips, summed over all months;
-
3.
Postojna Planina Cave System, with replicates consisting of cave sections, summed over all drips and all months.
Cave sections were Črna jama, Pivka jama, and Postojnska jama.Footnote 1 Mean distance between drips was 77 m for Črna jama, 99 m for Pivka jama, and 413 m for Postojnska jama. Overall, the distance between drips for PPCS was 1048 m (Pipan and Culver 2007).
Results
Temperature
Air temperatures for the period 22 November 2012 to 5 January 2017 are shown for two cave locations and one outside location (Fig. 2). Postojna 2 site showed the lowest values in summer and highest values in winter (Šebela and Turk 2011). Overall, temperature ranged from 10.1 to 11.1 °C, while surface temperature ranged from − 15 °C to 35 °C. Summer air temperature was less variable than winter temperature. In winter, cave air temperature at the Postojna 2 site showed larger daily oscillations than in summer, probably due to stronger ventilation with exchange of cave air masses with surface air masses. The reversal of seasonal temperatures at the Postojna 2 site relative to both the surface and Razpotje suggests that there is winter temperature exchange, not only with the surface, but also with cave passages.

Temperature profiles (air temperature measured every hour) for surface, Postojna 2 site, and Razpotje (see Fig. 1)
Razpotje data, from 19 March 2015 to 4 January 2017, has a significantly different pattern than the Postojna 2 site (Fig. 2). Highest temperatures were in mid-September (10.6 °C) and the lowest in January to March (9.6 °C). In winter, cold outside air penetrated deep into the cave system and cooled the air at the Razpotje site, resulting in an overall seasonal pattern similar to the surface, albeit with greatly reduced amplitude. Considered together, the two cave sites, while very close to each other (Fig. 1), have very different daily and seasonal temperature regimes.
No spectral analysis of Razpotje data was done, but differences in temperature pattern between the Lepe jame passage (similar to Razpotje) and the Postojna 2 site were obvious in spectral analysis (Fig. 3). The Postojna 2 site had a very weak 24-h signal (Šebela and Turk 2011), which indicated its relatively higher isolation from the rest of the cave, as did the reversal of times of maximum and minimum temperatures.

In 2016, mean annual air temperature at the Postojna 2 site was 10.63 °C, 10.19 °C at Razpotje, and 9.97 °C outside the cave. Historical cave air temperature data for 1933–1937 (Crestani and Anelli 1939) at Razpotje showed a temperature range of 7.5 to 8.4 °C. This was 1.74–2.64 °C lower than measurements taken from 2015 to 2017, with an average of 10.14 °C.
Discharge of percolation water
The relationship between precipitation and epikarst drip discharge is shown in Fig. 4 for a drip in the Kristalni passage (see Fig. 1). It is clear from Fig. 4 that there is not a simple lagged response of drip discharge to precipitation. After an extensive period of little or no response to precipitation (August 2005 to May 2006), there was a rapid increase in drip discharge, nearly simultaneously with a precipitation event of approximately 100 mm. The analog of epikarst to a sponge that first fills up before there is discharge is a useful one. Although we only show one temporal record here, Kogovšek (2010) showed that different drips also have different responses, presumably because of different pathways from the surface to the drip, and different sized storage reservoirs.

Discharge of springs
The relationship between discharges of the Pivka River, which sinks into PPCS and Unica spring at the outflow from the same system, shows a fast response of discharge to rainfall events for both streams, much faster than the drip response (Fig. 4, Supplement 1). Significant differences in the flow rates and shape of the recession part of the hydrographs indicate additional recharge of Unica spring from other parts of the karst aquifer. Overall, discharge of the Pivka River is more variable, indicating the buffering effects of the karst system between the insurgence (Pivka River) and resurgence (Unica spring). Other physico-chemical parameters (water temperature and conductivity) provided more insights into the complex nature of this karst aquifer (Petrič 2011; Petrič 2017).
Organic carbon
DOC concentrations in the sinking Pivka River were approximately the same as DOC concentrations in the underground rivers (Table 1). In contrast, concentrations of DOC in epikarst were four times lower (Fig. 5). DOC at the resurgence is intermediate, with values of 2.67 mg L−2 (Table 1). The apparent anomaly of higher concentrations of DOC in cave streams relative to their resurgence is because all measurements in the cave included water from the eutrophic Pivka River. The larger Rak River is less eutrophic. A schematic diagram of the standing stock of organic carbon is shown in Fig. 5.

Conceptual model of dissolved organic carbon (DOC) and particulate organic carbon (POC) in the Postojna Planina Cave System’s (PPCS) basin. Modified from Simon et al. (2007)
Epikarst fauna
Unlike many species accumulation curves for cave fauna (e.g., Zagmajster et al. 2010), species accumulation curves for the epikarst fauna reached asymptotes, indicating sampling completeness (Pipan and Culver 2007). The number of species of stygobiotic copepods found in individual drips was highly variable (Table 1), ranging from zero to seven. Species richness was much less in the Postojnska jama section of the cave (four, with a Chao estimate of total species of 4.5) than in either Črna jama (eight, with a Chao estimate of eight) or Pivka jama (eight, with a Chao estimate of nine). The thicker ceilings in the Postojnska jama section provide a partial explanation of the difference (Pipan 2005; Pipan and Culver 2013). At the scale of the entire cave, the estimate of species richness of 12 is well supported, both by accumulation curves and a Chao estimate of total richness of 12.
Some drips had a very high percentage of the total number of species found in a cave section (one drip in Pivka jama had 87% of the species known from the entire section of the cave). In general, about half of the species found in a cave section were found in each drip. Postojnska jama had a number of drips without copepods (Table 1).
Discussion
Approaches to the study of karst and cave ecosystems
Odum’s classic study of Silver Springs (Odum 1957) was a study of a karst system. His perspective was different from the one proposed here because it was the spring itself that was the ecosystem, i.e., the “black box,” with inputs being the boils of water emerging at the spring. This is the conceptual equivalent of making Unica spring in PPCS the black box ecosystem, with the PPCS as the input. Rouch (1970, 1977) shifted the perspective to the subsurface basin, rather than the spring, as the black box to be analyzed. For Rouch, inputs were sinking streams and infiltrating water. This approach in PPCS would focus on the recharges in the swallets and resurgence, and organic carbon at these two sites to the exclusion of epikarst drip hydrology and fauna.
Klimchouk (2015) provided geomorphological reasons why caves rather than karst should be central, and there are biological reasons as well. Caves are either the primary habitat for or the primary sampling point for the obligate subterranean fauna. We have shown that the epikarst fauna has many technical advantages as a focus of biological sampling (e.g., complete and quantitative). It is not part of the cave fauna in a strict sense, but it can only be sampled in caves. Epikarst drips are the source of the fauna found in pools that are situated so high that they never flood (Pipan et al. 2010). Epikarst drips are also a major source of organic carbon for caves (Simon et al. 2007).
Long-term studies in caves
Because of the presence of speleothems, which continue to grow by the deposition of calcium carbonate, it is possible to obtain long-term records of temperature and hence climate change (Fairchild and Baker 2012). Depending on their geographic location, they may be useful as proxies of local or of regional climate (Horvatinčić et al. 2003). These data afford the opportunity to connect contemporary studies of the fauna with long records of environmental conditions. Now that the threat of extinction of cave populations due to global warming has been recognized (Mammola et al. 2017), long-term records, both for millennia and for decades, will become increasingly interesting. While caves are quite stable with respect to many environmental variables, most caves, including PPCS, show daily and seasonable variation, as well as fine scale spatial variation, as was seen in the two temperature sites (Fig. 3). All of the analyses proposed here are designed to be repeated over the course of a multi-year study.
Conclusion
The five variables highlighted in this study are ones that encapsulate the dynamics of a cave ecosystem. Organic carbon, epikarst hydrology, and stream discharge are, together, the inputs and outputs of the system. The epikarst fauna is a proxy for the cave fauna, one that is relatively easy to quantitatively measure. And finally, temperature is a proxy for environmental cyclicity and variability in general.
The results we report here are only a sample of these studies, and there are more extensive results with respect to temperature (Šebela and Turk 2011), hydrology (Petrič 2011), and epikarst hydrology (Kogovšek 2010), as well as other environmental parameters (e.g., Gams 1974). Our intent was not to review this work in detail but to point to it as an example and a model for the long-term study of caves as ecosystems. The study of cave ecosystems must remain profoundly interdisciplinary and multidisciplinary. Perhaps it is because of the absence of vegetation in caves that makes the connection between the fauna on the one hand, and geology and hydrology on the other, so obvious and important.
Notes
We use the Slovenian names for the sections of the cave, including Postojnska jama, but use the English name for the cave system (Postojna Planina Cave System), in order to use the name of the ILTER site.
References
ARSO (2017) http://vode.arso.gov.si/hidarhiv/pov_arhiv_tab.php. Accessed 24 Apr 2017. https://doi.org/10.1016/j.pharep.2017.12.005
Auler A (2012) Quartzite caves in South America. In: White WB, Culver DC (eds) Encyclopedia of caves, 2nd edn. Academic/Elsevier Press, Amsterdam, pp 635–639. https://doi.org/10.1016/b978-0-12-383832-2.00093-1
Badino G (2010) Underground meteorology—what’s the weather underground? Acta Carsologica 39:427–448. https://doi.org/10.3986/ac.v39i3.74
Cigna AA (2002) Modern trend[s] in cave monitoring. Acta Carsologica 31:35–54. https://doi.org/10.3986/ac.v31i1.402
Colwell RK (2013) EstimateS: statistical estimation of species richness and shared species from samples. Version 9. User’s Guide and application http://purl.oclc.org/estimates. Accessed 25 Apr 2017
Covington M, Perne M (2015) Consider a cylindrical cave: a physicist’s view of cave and karst science. Acta Carsologica 44:363–380 https://doi.org/10.3986/ac.v44i3.1925
Crestani G, Anelli F (1939) Ricerche di meteorologia ipogea delle grotte di Postumia. Istitutopolografico dello stato Libreria, Roma
Culver DC, Pipan T (2009) The biology of caves and other subterranean habitats. Oxford Univ Press, Oxford. https://doi.org/10.1097/GIM.0b013e3181b2e5eb
Culver DC, Pipan T (2013) Subterranean ecosystems. In: Levin SA (ed) Encyclopedia of biodiversity, vol 7, 2nd edn. Academic Press, Waltham, pp 49–62. https://doi.org/10.1016/B978-0-12-384719-5.00224-0
Danielopol DL, Rouch R (1991) L'adaptation des organismes au milieu aqautique souterrain. Reflexionssur l'apport des recherches ecologiques recentes. Stygologia 3:129–142
Domínguez-Villar D, Lojen S, Krklec K, Baker A, Fairchild IJ (2015) Is global warming affecting cave temperatures? Experimental and model data from a paradigmatic case study. Clim Dyn 45:569–681. https://doi.org/10.1007/s00382-014-2226-1
Engel AS (2012) Chemoautotrophy. In: White WB, Culver DC (eds) Encyclopedia of caves, 2nd edn. Academic/Elsevier Press, Amsterdam, pp 125–134. https://doi.org/10.1016/b978-0-12-383832-2.00019-0
Engel AS (2015) Preface. In: Engel AS (ed) Microbial life of cave systems. De Gruyter, Berlin, pp v–vi. https://doi.org/10.1515/9783110339888-001
Fairchild IJ, Baker A (2012) Speleothem science. From process to past environments. Wiley-Blackwell, Chichester. https://doi.org/10.1002/9781444361094
Ford DC, Williams PW (2007) Karst hydrogeology and geomorphology. Wiley, Chichester
Friedrich M (2013) Biological clocks and visual systems in cave-adapted animals at the dawn of speleogenomics. Integr Comp Biol 53:50–67. https://doi.org/10.1093/icb/ict058
Gaberščik A (2003) Jezero, ki izginja. Društvo Ekologov Slovenije, Ljubljana
Gams I (1974) Concentration of CO2 in caves in relation to the air circulation (in the case of the Postojna Cave). Acta Carsologica 6:183–192
Genthon P, Bataille A, Fromant A, D’Hulst D, Bourges F (2005) Temperature as a marker for karstic waters hydrodynamics. Inferences from 1 year recording at La Peyrére cave (Ariège, France). J Hydrol 311:157–171. https://doi.org/10.1016/j.jhydrol.2005.01.015
Gibert J (1986) Ecologie d’un systeme karstique jurassien. Hydrogéologie, derive animale, transits de matières, dynamique de la population de Niphargus (Crustacé Amphipode). Mém Biospéol 13:1–379
Hartmann A, Baker A (2017) Modelling karst vadose zone hydrology and its relevance for paleoclimatereconstruction. Earth Sci Rev 172:178–192. https://doi.org/10.1016/j.earscirev.2017.08.001
Horvatinčić N, Bronić IK, Obelić B (2003) Differences in the 14C age, δ13C and δ18O of Holocene tufa and speleothem in the Dinaric Karst. Palaeogeogr Palaeoclimatol Palaeoecol 193:139–157. https://doi.org/10.1016/S0031-0182(03)00224-4
ILTER (2018) https://data.ilter-europe.net/deims/. Accessed 24 Jan 2018. https://doi.org/10.1111/add.14244
Kaufmann G, Gabrovšek F, Turk J (2016) Modelling flow of subterranean Pivka river in Postojnska jama, Slovenia. Acta Carsologica 45:57–70. https://doi.org/10.3986/ac.v45i1.3059
Klimchouk A (2015) The karst paradigm: changes, trends, and perspectives. Acta Carsologica 44:289–313. https://doi.org/10.3986/ac.v44i3.2996
Kogovšek J (2010) Characteristics of percolation through the karst vadose zone. ZRC Publishing, Ljubljana
Kogovšek J, Šebela S (2004) Water tracing through the vadose zone above Postojnska Jama, Slovenia. Environ Geol 45:992–1001. https://doi.org/10.1007/s00254-003-0958-z
Kogovšek J, Urbanc J (2007) Ocena dinamike premikajoče vode skozi vadozno cono Postojnske jame na osnovi izotopskih značilnosti. Geologija 56:477–486. https://doi.org/10.5474/geologija.2007.033
Kresic N (2013) Water in karst: management, vulnerability, and restoration. McGraw-Hill, New York
Kresic N, Stevanovic Z (2010) Groundwater hydrology of springs. Elsevier, Amsterdam
Lange J, Arbel Y, Grodek T, Greenbaum N (2010) Water percolation process studies in a Mediterranean karst area. Hydrol Process 24:1866–1879. https://doi.org/10.1002/hyp.7624
Mammola S, Goodacre SL, Isaia M (2017) Climate change may drive cave spiders to extinction. Ecography 40. https://doi.org/10.7717/peerj.3972
Mayaud C, Wagner T, Benischke R, Birk S (2013) Understanding changes in the hydrological behaviour within a karst aquifer (Lurbach system, Austria). Carbonates Evaporites 31:357–365. https://doi.org/10.1007/s13146-013-0172-3
McDonald J, Drysdale R (2007) Hydrology of cave drip waters at varying bedrock depth from a karst system in southeastern Australia. Hydrol Process 21:1737–1748. https://doi.org/10.1002/hyp.6356
Monroe WH (1972) A glossary of karst terminology. U.S. Geol Survey Water Supply Paper 1899-K. U.S. Geol Survey, Washington. https://doi.org/10.1172/JCI107074
Novak T, Janžekovič F, Lipovšek S (2013) Contribution of non-troglobiotic terrestrial invertebrates to carbon input in hypogean habitats. Acta Carsologica 42:301–309. https://doi.org/10.3986/ac.v42i2-3.669
Odum HT (1957) Trophic structure and productivity of Silver Springs, Florida. Ecol Monogr 27:55–112. https://doi.org/10.2307/1948571
Palmer AN (2007) Cave geology. Cave Books, Dayton
Parker CW, Wolf JA, Auler AS, Barton HA, Senko JM (2013) Microbial reducibility of Fe (III) phases associated with the genesis of iron ore caves in the Iron Quadrangle, Minas Gerais, Brazil. Minerals 3:395–411. https://doi.org/10.3390/min3040395
Petrič M (2011) Spatial and temporal variations of natural traces in a karst spring catchment at the Planina polje (SW Slovenia). In: Bertrand, C. (ed) Proceedings of the 9th conference on limestone hydrogeology = Actes du 9ème congrès d’hydrogéologie en pays calcaire, Besançon, France, September 1–3, 2011. [S. l.]: UMR 6249 Chrono-Environnment. pp 399–402
Petrič M (2017) Flow dynamics in a karst aquifer studied by means of natural and artificial tracers: a case study of the Malenščica and Unica karst springs. Geologija 60:211–221. https://doi.org/10.5474/geologija.2017.015
Pipan T (2005) Epikarst – a promising habitat. Založba ZRC, ZRC Publishing, Ljubljana
Pipan T, Culver DC (2007) Regional species richness in an obligate subterranean dwelling fauna—epikarst copepods. J Biogeogr 34:854–861. https://doi.org/10.1111/j.1365-2699.2006.01667.x
Pipan T, Culver DC (2013) Forty years of epikarst: what biology have we learned? Int J Speleol 42:225–233. https://doi.org/10.5038/1827-806X.42.3.5
Pipan T, Christman MC, Culver DC (2006) Dynamics of epikarst communities: microgeographic pattern and environmental determinants of epikarst copepods in Organ Cave, West Virginia. Am Midl Nat 156:75–87. https://doi.org/10.1674/0003-0031(2006)156[75:DOECMP]2.0.CO;2
Pipan T, Holt N, Culver DC (2010) How to protect a diverse, poorly known, inaccessible fauna: identification of source and sink habitats in the epikarst. Aquat Conserv Mar Freshwat Ecosyst 20:748–755. https://doi.org/10.1002/aqc.1148
Pipan T, López H, Oromí P, Polak S, Culver DC (2011) Temperature variation and the presence of troglobionts in shallow subterranean habitats. J Nat Hist 45:253–273. https://doi.org/10.1080/00222933.2010.523797
Rice EW, Baird RB, Eaton AD (2017) Standard methods for the examination of water and wastewater, 23rd edn. Am Pub Health Assoc, Washington, DC. https://doi.org/10.1097/INF.0000000000001875
Romero A (2009) Cave biology. Life in darkness. Cambridge Univ Press, Cambridge. https://doi.org/10.2174/157340309789317887
Rouch R (1970) Recherches sure les eaux souterraines—12—Le système karstique du Baget. I. Lephénoméne d’ “hémorragie’ au niveau de l’exutoire principal. Ann Spéléol 25:665–709
Rouch R (1977) Considérations sur l’écosystème karstique. Comp Rend Acad Sci, Paris 284:1101–1103
Schneider K, Christman MC, Fagan WF (2011) The influence of resource subsidies on cave invertebrates: results from an ecosystem-level manipulation experiment. Ecology 92:765–776. https://doi.org/10.1890/10-0157.1
Šebela S (2012) Postojna-Planina cave system. In: White WB, Culver DC (eds) Encyclopedia of caves, 2nd edn. Academic/Elsevier Press, Amsterdam, pp 618–624. https://doi.org/10.1016/B978-0-12-383832-2.00091-8
Šebela S, Turk J (2011) Local characteristics of Postojna Cave climate, air temperature, and pressure monitoring. Theor Appl Climatol 105:371–386. https://doi.org/10.1007/s00704-011-0397-9
Šebela S, Turk J, Pipan T (2015) Cave micro-climate and tourism: towards 200 years (1819-2015) atPostojnska jama (Slovenia). Cave Karst Sci 42:78–85
Simon KS (2013) Organic matter flux in the epikarst of the Dorvan karst. Acta Carsologica 42:237–244. https://doi.org/10.3986/ac.v42i2-3.665
Simon KS, Benfield EF (2001) Leaf and wood breakdown in cave streams. J N Am Benthol Soc 20:550–563. https://doi.org/10.2307/1468087
Simon KS, Benfield EF (2002) Ammonium retention and whole-stream metabolism in cave streams. Hydrobiologia 482:31–39. https://doi.org/10.1023/A:1021257822591
Simon KS, Benfield EF, Macko SA (2003) Food web structure and the role of epilithic films in cave streams. Ecology 84:2395–2406. https://doi.org/10.1094/PHYTO.2003.93.11.1422
Simon KS, Pipan T, Culver DC (2007) A conceptual model of the flow and distribution of organic carbon incaves. J Cave Karst Stud 69:279–284
Simon KS, Pipan T, Ohno T, Culver DC (2010) Spatial and temporal patterns in abundance and character of dissolved organic matter in two karst aquifers. Fundam Appl Limnol 177:81–92. https://doi.org/10.1127/1863-9135/2010/0177-0081
Sket B (1979) Jamska favna notranjskega trikotnika (Cerknica-Postojna-Planina), njena ogroženost in naravovarstveni pomen. Varstvo Narave 12:45–59
Sket B, Velkovrh F (1981) Postojnsko-Planinski jamski sistem kot model za preučevanje onesnaženjapodzemeljskih voda. Naše Jame 22:27–44
Turk J (2010) Hydrological role of large conduits in karst drainage system: examples from the Ljubljanica River catchment area. Dissertation, University of Nova Gorica
Venarsky MP, Huntsman BM, Huryn AD, Benstead JP, Kuhajda BR (2014) Quantitative food web analysis supports the energy-limitation hypothesis in cave stream ecosystems. Oecologia 176:859–869. https://doi.org/10.1007/s00442-014-3042-3
Williams PW (2008) The role of the epikarst in karst and cave hydrogeology: a review. Int J Speleol 37:1–10. https://doi.org/10.5038/1827-806X.37.1.1
Worthington RH, Soley RVN (2017) Identifying turbulent flow in carbonate aquifers. J Hydrol 552:70–80. https://doi.org/10.1016/j.jhydrol.2017.06.045
Worthington SRH, Jeannin P-Y, Alexander EC, Davies GJ, Schindel GM (2017) Contrasting definitions for the term ‘karst aquifer’. Hydrogeol J 25:1237–1240. https://doi.org/10.1007/s10040-017-1628-7
Zagmajster M, Culver DC, Sket B (2008) Species richness patterns of obligate subterranean beetles in a global biodiversity hotspot—effect of scale and sampling intensity. Divers Distrib 14:95–105. https://doi.org/10.1111/j.1472-4642.2007.00423.x
Zagmajster M, Culver DC, Christman MC, Sket B (2010) Evaluating the sampling bias in pattern of subterranean species richness—combining approaches. Biodivers Conserv 19:3035–3048. https://doi.org/10.1007/s10531-010-9873-2
Acknowledgments
We thank Wolfgang Cramer, Editor-in-Chief, Thomas Dirnböck, Guest Editor, Christopher Reyer, and two anonymous reviewers for their valuable comments and suggestions.
Funding
This work was supported by grants from the European Commission (GA: 654359 - H2020 INFRAIA, eLTER and GA: 739558 - H2020 INFRADEV Advance_eLTER), and from the Slovenian Research Agency (ARRS projects “Natural resources of karst show caves: a balance among protection, exploitation, and promotion [no. J7-7100,]” and “Karst research for sustainable use of Škocjan Caves as World Heritage [(no. L7-8268)].
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Supplement 1
Discharge from the sinking of the Pivka River and the emergence at Unica Spring, and precipitation. (GIF 30 kb)
Rights and permissions
About this article

Cite this article
Pipan, T., Petrič, M., Šebela, S. et al. Analyzing climate change and surface-subsurface interactions using the Postojna Planina Cave System (Slovenia) as a model system. Reg Environ Change 19, 379–389 (2019). https://doi.org/10.1007/s10113-018-1349-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10113-018-1349-z