
Abstract
Increased dieback and mortality of “dark needle conifer” (DNC) stands (composed of fir (Abies sibirica), Siberian pine (Pinus sibirica) and spruce (Picea obovata)) were documented in Russia during recent decades. Here we analyzed spatial and temporal patterns of fir decline and mortality in the southern Siberian Mountains based on satellite, in situ and dendrochronological data. The studied stands are located within the boundary between DNC taiga to the north and forest-steppe to the south. Fir decline and mortality were observed to originate where topographic features contributed to maximal water-stress risk, i.e., steep (18°–25°), convex, south-facing slopes with a shallow well-drained root zone. Fir regeneration survived droughts and increased stem radial growth, while upper canopy trees died. Tree ring width (TRW) growth negatively correlated with vapor pressure deficit (VPD), drought index and occurrence of late frosts, and positively with soil water content. Previous year growth conditions (i.e., drought index, VPD, soil water anomalies) have a high impact on current TRW (r = 0.60–0.74). Fir mortality was induced by increased water stress and severe droughts (as a primary factor) in synergy with bark-beetles and fungi attacks (as secondary factors). Dendrochronology data indicated that fir mortality is a periodic process. In a future climate with increased aridity and drought frequency, fir (and Siberian pine) may disappear from portions of its current range (primarily within the boundary with the forest-steppe) and is likely to be replaced by drought-tolerant species such as Pinus sylvestris and Larix sibirica.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
An increase in dark needle conifer forests (DNC: Siberian pine (Pinus sibirica (Rupr.) Mayr.), spruce (Picea obovata Ledeb.) and fir (Abies sibirica Ledeb.) dieback and mortality has been reported throughout Russia (Fig. s1). In the European part of Russia, spruce decline was observed on over 2 million ha (Zamolodchikov 2012), and in the Archangelsk region spruce mortality was observed on more than 390 thousand ha (Chuprov 2008). Potential causes of spruce mortality that were considered included over maturity of stands, drought, root fungi and insect attacks (Pavlov et al. 2008; Russian Federal Agency of Forestry 2013). In Siberia, decline and mortality of Abies sibirica and Pinus sibirica were observed in the southern Siberian Mountains where drought was observed to be a primary factor of mortality with bark beetles and fungi attack as cofactors (Kharuk et al. 2013a). Siberian pine and fir mortality was also described in the Baikal Mountains and attributed to water stress in synergy with pest attacks (Kharuk et al. accepted). Mixed spruce (Picea ajansis) and fir (Abies nephrolepis) forests mortality in the Russian Far East was considered to be caused by “unfavorable climatic factors with fungi as a cofactor” (Man’ko et al. 1998). Birch mortality, caused by severe drought, was documented within the Trans-Baikal forest-steppe (Kharuk et al. 2013b).
In Eastern Europe, spruce (Picea abies) mortality was documented in Belarus on over 300 thousand ha and was attributed to the combined effect of drought and bark beetle attacks (Sazonov et al. 2013; Kharuk et al. 2015a). Spruce decline was also reported in Latvia, Lithuania (Arhipova 2013; Vasilyauskas 2013), Ukraine and other European countries (Yousefpour et al. 2010; Martínez-Vilalta et al. 2012; Allen et al. 2015). On a global scale, forest decline and mortality were reported for every continent (Worrall et al. 2010; Allen et al. 2015) and commonly attributed to increased drought episodes in synergy with insect and fungal attacks (Breda et al. 2006; Aitken et al. 2008; Anderegg et al. 2013).
Topography has a mediating effect on the spatial pattern of drought stress and tree mortality. Spatial variations in site moisture conditions may influence the spatial pattern and severity of drought-induced tree mortality (Guarın and Taylor 2005). For example, slope aspect was found to be a key variable influencing soil moisture availability in the western mountains of North America (Stephenson 1990). Siberian pine and spruce stands climate-induced mortality was found to be related to elevation, terrain curvature, slope steepness and aspect (Kharuk et al. 2013a, 2015a). Along with droughts, insect-caused tree mortality also is dependent on relief features (Lausch et al. 2013).
The objective of this study was to analyze temporal and spatial patterns and causes of fir decline and mortality in the southern Siberian East Sayan Mountains. We hypothesize that mortality was triggered by drought and that topography features (slope steepness, aspect and elevation) influence the timing and extent of tree death. In particular, we expect that mortality is greater for topography features with a higher risk for water stress.
Materials and methods
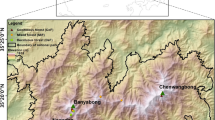
The study area was located in central Siberia. Forest decline and mortality studies were based on dendrochronology, climate variables and geospatial analysis, in situ measurements and satellite (Landsat, QuickBird and GRACE) image analysis as described below.
Study area
The studied stands covered 108,400 ha of the East Sayan Mountains (Fig. s1; inset) in Siberia, Russia. Studied stands were dominated by fir (Abies sibirica) with a mixture of Siberian pine (Pinus sibirica), spruce (Picea obovata) and birch (Betula pendula). Abies sibirica is an endemic species that ranges into the northeastern European part of Russia (Fig. s1). The area covered by fir in Russia is over 14.4 million ha including 8.8 million ha of fir-dominated stands (Fig. s1). Fir is a tree with mean height of about 20–25 m and a lifespan of about 150 years (with maximum up to 300 years). It has an extensive but shallow root system with thin, short roots. Fir is a shade-tolerant and moisture-sensitive species with optimal growth achieved with about 600–700 mm year−1 of precipitation (Krylov 1961). Regeneration of Abies sibirica is able to survive under dense upper canopy for a long time until released by any opening in the upper canopy (Krylov 1961). The study area is located within the transition between “dark needle” taiga on the northeast and forest-steppe on southwest (Fig. s1).
Climate data
Climate within the study area is continental with long cold winters and warm or hot summers (Fig. 1).

Climate variables within the East Sayan study area. a, b, c, d—mean air temperature, precipitation, drought index SPEI and vapor pressure deficit anomalies, respectively (base period 1901–2013). Significant trends are shown by solid (May–August) and dashed (annual) lines; e, f—average annual cycle of temperature and SPEI, respectively (averaged periods shown on panels). Notes 1. SPEI decrease means drought increase by definition. 2. Droughts occurs during 1990, 1998, 2008 (see also Fig. 4)
Meteorological variables (i.e., air temperature, precipitation, number of days with frosts) were obtained from the British Atmospheric Data Centre (www.badc.nerc.ac.uk); the variables were acquired as monthly mean values based on daily values. Vapor pressure deficit values were calculated. Drought index (SPEI), data were obtained from (www.sac.csic.es/spei). SPEI (the Standardized Precipitation–Evapotranspiration index) uses the monthly difference (D) between precipitation (P) and potential evapotranspiration (PET) (Vicente-Serrano et al. 2010):
SPEI data were calculated for the May–August period, since May droughts were typical for the study area (Fig. 1). All climatic data were averaged for a cell size 0.5° × 0.5° (~33 × 56 km).
Field studies
Field studies were conducted in the summers of 2013 and 2014. The main goal of on-ground studies was tree sampling for dendrochronology analysis. Ancillary collected data included a description of soil and ground cover and forest inventory measurements (including forest health estimation). The latter were also used as ground truth for Landsat digital image analysis.
Test sites (TS) were preselected based on Landsat and QuickBird data analyses. QuickBird’s on-ground resolution (0.6 m) enabled differentiation of live and dead trees by expert visual photointerpretation. It was found that stands with signs of decline and mortality occupied the 500–1050 m a.s.l. elevation belt. Forest-tundra and alpine tundra occupied the upper elevations. TS (N = 9) were established along the elevation gradient within a range of 500–950 m, where the majority of forest decline and mortality were observed. TS were selected within typical declining and dead stands (N = 7) and stands without noticeable damage (N = 2; Fig. s2). Thus, TS were likely to be representative of the entire study area.
For dendrochronological analysis, dominant trees were randomly sampled within ±10 m elevation range from the TS centerpoint coordinate. The sampling area for each TS was about 0.5 ha. Samples were taken by increment bore or by chainsaw (disks above root tension zone). On each TS at least 10 samples were taken to ensure reliable statistics. So the total sample set is 114 (94 living plus 20 dead)—24 removed = 90 plus 18 regeneration = 118. The latter was defined as trees with height about ¼ of the upper canopy. Twenty-four living trees were removed from analysis because they were invalid for cross-dating.
Forest inventory data were obtained on circular (radius = 9.8 m) test plots within each TS. The data included a description of forest type and species composition, tree heights and diameter at breast height (dbh), canopy closure, trees and regeneration vigor, regeneration abundance and composition, signs of insect attacks (insect and larvae galleries within the bark and xylem) and fungi impacts, and signs of wildfire impact (i.e., charcoal and fire scars on the boles). Ground cover, soil type, depth to bedrock and relief features (i.e., azimuth, slope steepness, elevation) were described. For the Landsat digital analysis, trees were divided into living, declining and dead groups. The category “dying trees” included trees with crown density decrease, needles discoloration (yellow, green–yellow or gray) and progressive radial increment decrease. The category “dead trees” included dead and severely damaged trees (a partial loss of small branches and bark).
Satellite scenes analysis
Landsat scenes, essentially cloud-free, were used for mapping stand mortality dynamics within the study area. Scenes (with pixel size 15–30 m) were acquired for the summers of 1989, 2000, 2003, 2006–2010, 2012 with a total of 23 images (obtained from glovis.usgs.gov). Landsat data analysis was based on ground truth data from established TS, QuickBird scenes (n = 3; pixel size was 0.6 m) and expert knowledge. Maximum likelihood classification techniques with thresholding were applied using ERDAS Imagine software (geospatial.intergraph.com).
First, a mask of fir stands before mortality was generated based on the Landsat scenes of 1989. For this purpose, 10 sample areas of dark needle stands (with about ~7600 pixels per sample area) were selected within the Landsat scene. The generated mask was used for tracking temporal fir mortality within dark needle stands only.
Second, dead stands (i.e., consist of >75% of dead trees) were mapped. The other ten sample areas were used for dead and declining stands (with about 105 pixels per sample area), and three sample areas were taken for healthy stands (with about 12840 pixels per sampled area). Two statistics were calculated: First, the total dead stand’s area for a given year and second, dead stands area increase (area increment) for a given period.
Third, classification accuracy was estimated based on test points with 728 points for dead stands and 285 test points for healthy stands. All points were selected based on the QuickBird scenes interpretation. Overall classification accuracy was good (87%, Kappa-statistic was 0.71; Table s1).
GRACE gravimetric data was applied for detection of soil water anomalies. We used monthly EWTA (Equivalent of Water Thickness Anomalies) for the analysis. EWTA were measured with accuracy 10–30 mm month−1 (Long et al. 2014). GRACE spatial resolution was one by one degree (~66 × 112 km at latitude 54°). These data have been available since 2003 (www.grace.jpl.nasa.gov). The satellite data were analyzed using ERDAS Imagine software (www.geospatial.intergraph.com) and ESRI ArcGIS software (www.esri.com). StatSoft Statistica (StatSoft Inc. 2013) was used for statistical analysis.
Geospatial analysis
Topographic analysis was based on the Deutsche Zentrum für Luft- und Raumfahrt (DLR) SRTM-XSAR DEM with absolute and relative horizontal accuracy of ±20 and ±15 m, respectively, and with absolute and relative vertical accuracy of ±16 and ±6 m, respectively (http://eoweb.dlr.de:8080/index.html). The analyzed elevation range interval was 50 m. Aspect, slope steepness and curvature (i.e., convex/concave slope parameters) data were calculated from the DEM using ArcGIS tools. The aspect data were quantized to eight directions (i.e., by 45° corresponding to north, northeast, east).
The distribution of topography features with a given altitude, azimuth and slope steepness was uneven within the study area and thus could lead to biased analysis. To avoid this, the data were normalized by the following procedure. The analyzed forested area (about 52,000 ha) was stratified into intervals of azimuth (45°), slope steepness (1°), curvature (0.5) and elevation (100 m) and related to the total study area (which was about 108,400 ha and included both, forested and non-forested areas) with similar parameters:
where the c(i) subscript represents the ith category of relief feature c, AF c(i) is the area of the given on-ground class within the ith category of the relief feature c, and AT c(i) is the area of the ith category of relief feature c over the total territory. The “normalized” results are presented in relative units below.
Dendrochronology analysis
The surface of each tree disk or core sample was sanded and treated with a contrast enhancing powder. The tree ring widths (TRW) were measured with 0.01 mm precision using a linear table instrument (LINTAB-III). The TSAP and COFECHA computer programs were used in tree ring analysis (Holmes 1983; Rinn 1996). Dates of tree mortality were determined based on the master-chronology method (Fritts 1991). A master chronology was constructed based on 70 living trees. This master chronology was used for cross-dating dead trees. The mean coefficient of correlation between individual tree ring series and master chronology was 0.44. The mean sensitivity of individual series included into master chronology was satisfactory (0.221). The final dataset was divided into “surviving” (n = 70) and “dead” (n = 20) groups. For both groups, standard and residual chronologies were constructed. Standard chronologies were indexed using ARSTAN software [i.e., double detrending to remove long-term trends (Cook and Holmes 1986)]. The resulting chronologies were a unitless index of radial tree growth. Statistical analysis was carried out using Microsoft Excel and StatSoft software (StatSoft Inc. 2013).
Results
Study site inventory data
Within the study area, mean fir tree height was about 15.0 m, mean dbh 19.0 cm, and mean age 110 years. Crown closure of stands was about 70–80%. The rooting depth of fir trees was about 20–60 cm. Ground cover was mesophytic dominated by sedges. The soil type was mountain sandy–clay rocky podzolized underlain by stony clay at depths of 5–50 cm. There were no outward signs of fires within the test sites, i.e., there were no charcoal or fire scars visible on the boles.
Satellite-derived spatial and temporal patterns of fir stand mortality
The total study area was about 108,400 ha, including 52,600 ha of fir stands (with 6500 ha of dead stands). The Landsat analysis showed that the area of dead stands increased since the year 2000 and reached the maximum value in the year 2010 [i.e., 6500 ha, or 12% of total stand area (Fig. 2a)]. Fir mortality (expressed as % of dead trees) started on convex shaped ridge tops and steep south-facing slopes (Figs. 2, 3). Initially the area covered by dead stands was greatest at higher elevations and shifted with time to lower elevations (about 720 m) and to gentle (5°–7°) slopes (Figs. 2a, 3a). Stand mortality was uneven along the elevational gradient with maximum mortality within the 500–700 m elevation belt, decreasing with an increase in elevation (Fig. 2b). The mortality started on steep southern slopes, increasing with an increase in slope steepness (with saturation at about 20°). Stands with at least 50% mortality were located within the slope range of 18°–25° (Fig. 3a, b). The dead stands occupied steeper slopes almost within all elevation ranges with the exception of 900–1100 m (Fig. 3c). Mortality was also sensitive to terrain curvature, increasing within convex relief features (Fig. 3d). With respect to azimuth, the maximum area of dead stands was located on southwest slopes, whereas total stands area distribution was uniform (Fig. 2c, d).

Dead stand area dynamics with respect to elevation (a, b) and azimuth (c, d). a Elevational distribution of dead stands in 2003, 2007 and 2010. Maximum values are shown by vertical lines. Note: the sum of the area under each curve corresponded to the dead stand area in a given year. b Elevational distributions of dead stands and all stands. c Azimuth distribution of dead stands in the years 2003, 2007 and 2010. d The final azimuth distribution of dead and all stands (year 2010). Data presented in relative units (“normalized” against total study area based on the Eq. 2)

Distribution of stand area with respect to slope steepness and terrain curvature. a Dead stand area since beginning of mortality (in 2003), and area increments since 2003 (2007), and since 2007 (2010). b Distribution of the dead and total stand area. Medians on (a, b) are indicated by vertical dashed lines. c Median of slope steepness per elevation class for dead stand and total stand area. d Stand mortality with respect to study area curvature (concave are negative, convex are positive)
Dendrochronology data
The tree ring width (TRW) trajectories of “survivors” and “decliners” tree cohorts diverged after drought in 1990 (Fig. 4). The “decliners” cohort eventually died at the beginning of the twenty-first century, whereas the “survivors” cohort showed a growth release. A similar event (i.e., a growth depression with a following growth increase) happened at the beginning of the twentieth century (Fig. 4).

Tree ring width (TRW) chronology of “survivors” and “decliners” tree cohorts and regeneration. Confidence interval (p < 0.05) shown by gray background. Black bars: the percentage of sampled dead trees that died in the given year. Arrows indicate droughts. Note: scale for regeneration given with scale coefficient 0.5 for better reading
Tree ring width and ecological variables
Correlation analysis showed that trees in both “survivors” and “decliners” cohorts became sensitive to climate variables (i.e., a correlation becomes significant) since the beginning of the 1980s (Fig. 4; Table s2). The TRW of the “survivors” cohort showed positive correlations with drought decrease (note that an SPEI increase means drought decrease by definition), whereas negative correlations were obtained with VPD and number of May–July days with frosts (Fig. 5). Similarly, the tree ring width of the “decliners” cohort was positively correlated with SPEI and negatively correlated with a VPD, whereas no correlations were observed with frost days (Table s2). Since 1990, a significant change in the SPEI annual pattern was observed: an increase in drought during May and July (Fig. 1f). For both cohorts correlation with summer temperatures and precipitation were insignificant. The climatogram (Fig. 1e) showed that temperature increases were mainly observed during winter months. The time intervals (May–June, May–August) were selected based on the criteria of maximum correlation between TRW and climate variables. TRW correlated with soil water anomalies in June (Fig. 5b). Notably, the highest correlations were found between TRW and ecological variables of the previous year (Fig. 5a, b, c; Table s2). Similarly, autocorrelation (i.e., correlation of current year TRW with previous ones) reached maximal values with the previous year TRW (Fig. s4). Thus, fir is sensitive to anomalies in atmospheric humidity and soil water content in both current and previous years.

Relationship between tree ring width (TRW) of all trees and climate variables of the previous (a, b, c) and the current (b, d) year. a SPEI for previous year May–August (1981–2006). b Soil water anomalies (EWTA) for June (dots, current year) and September (triangles, previous year); period: 2003–2011. c Mean VPD for previous year May–June (1985–1999). d Sum of frost days for current year May–June (1990–2011)
Discussion
Fir mortality and relief features
The area of dead stands gradually increased after the 1990, 1998 and 2008 drought episodes (Figs. s3, 4), and the increase was uneven with respect to elevation, exposure, slope steepness and convexity (Figs. 2, 3). There was a general trend of decreasing tree mortality with an increase in elevation within the study area. Maximum mortality was observed within the 500–700 m elevation belt (Fig. 2b). That decrease in mortality could be attributed to the elevational gradient of precipitation and humidity, as well as a potential evapotranspiration decrease at higher elevations. Meanwhile, locally within that elevation belt, mortality started on hilltops with convex shape and hills with steep south-facing slopes. Over time, mortality shifted downward to gentle slopes. Dead stands were found on steeper slopes, and the mortality increased as steepness increased. The majority of stand mortality was located on these steeper slopes with approximately 20% higher solar radiation in comparison with the plains (i.e., an 18°–25° range). The slope aspect and steepness effect on tree mortality were also described in other papers (e.g., Stephenson 1990; Lausch et al. 2013). However, in those studies there was a lack of quantitative analysis between relief features and mortality.
Thus, stand mortality was located mainly on terrain features with maximal risk of water stress. In addition, drought impact on trees was amplified by shallow well-drained rocky soils, especially on steep convex slopes, where bedrock was at about 10–25 cm depth.
Fir mortality and ecological variables
The forests within our study area location (i.e., within the transition between “dark needle” taiga on the northeast and forest-steppe on southwest) were found to be especially sensitive to anomalies in atmospheric humidity and soil water content. Tree ring width positively correlated with drought decrease and soil water content, and negatively with vapor pressure deficit (VPD). The highest correlation was found using climatic conditions of the previous year (i.e., drought index, VPD soil water content; Table s2). A positive correlation with soil water content in September of the previous year (Fig. 5b) indicated the importance of soil water captured in previous year autumn for tree growth in the current year. That should be of special importance for the water-stress risk areas (steep slopes of southward exposure; Fig. 2) because snowmelt water mainly runs off over the still frozen root zone. Autocorrelation also showed a high impact (r = 0.8) of the previous year into the current year (Fig. s4).
The impact of the previous year growth conditions on tree ring formation is known although commonly the main input is attributed to current year variables. For example, according to Kagawa et al. (2006), xylem formation of Larix gmelinii was approximately 43% dependent on the previous year’s photoassimilates. These observations agree with our data, but differ in the effect, i.e., higher effect of the previous year’s input in comparison with the current one. That effect may be attributed to the drought-induced trees becoming susceptible to biotic impacts. Thus, bark beetles and fungi impacts on the drought-weakened trees may lead to a decrease in the current year growth increment. Negative impacts of late spring frosts on the TRW (Fig. 5d) might be attributed to the known fir sensitivity to frosts that kill “flushed” apical shoots.
“Survivors” and “decliners” cohorts response to climate variables was similar before the droughts at the end of the twentieth century. After these droughts, “decliners” showed a negative trend of TRW growth and ultimately died (Fig. 4). For both groups, the period of greatest sensitivity of TRW to climate variables was observed since the end of the 1980s (Fig. s4). This was at the beginning of noticeable climate changes in general (Stocker et al. 2013) and drought increase within the study site in particular (Fig. 1f).
The regeneration survived drought and showed a growth release after upper canopy mortality (Fig. 4). A similar phenomenon has also been described in other works. For example, spruce (Picea ajansis) and fir (Abies nephrolepis) mortality in Russian Far East, as well as Siberian pine mortality in southern Siberia, was observed for mainly mature stands, whereas the regeneration survived (Man’ko et al. 1998; Kharuk et al. 2013b). The possible causes were a decrease in water stress due to canopy shading and lower regeneration leaf area index (LAI) in comparison with mature trees (Rautiainen et al. 2012).
The reason for fir drought-sensitivity (as well as that of Siberian pine and spruce) is a high LAI, a major determinant of water balance. For example, mixed fir, Siberian pine and spruce stands had LAI up to 7–8, whereas LAI of drought-tolerant Scotch pine stands is about 3–4 (Utkin 1975, re-calculated data). Due to its high LAI, fir does not tolerate low relative humidity, especially in spring. This agrees with high TRW correlation with vapor pressure deficit in May–June, when probability of drought is high (Fig. 5). SPEI data also showed a May drought increase during recent decades (Fig. 1f). Spring–early summer droughts (accompanied by high solar radiation) occurred when the rooting zone is still frozen and leads to needle desiccation and chlorosis (typically on the sun-facing part of the crown). In years of extreme atmospheric droughts in spring fir stands partly turned red on vast territories (upwards of hundreds of thousands of ha). Sites with thick moss and lichen ground cover experienced more damage due to decrease in soil insolation and, consequently, delay of root zone thawing.
The dendrochronology record indicated similar drought-induced fir mortality at the beginning of the twentieth century, when fir experienced lower annual TRW growth followed by a large growth increase (Fig. 4). The latter was similar to the recorded fir regeneration growth after the droughts at the end of the twentieth century (Fig. 4). The growth depression at the beginning of the twentieth century coincided with air temperature and precipitation anomalies (Fig. 1) revealed in the historical records of severe drought in 1912–1914 (Kirillov and Shherbakov 1961). The surviving regeneration formed stands, which experienced drought impacts at the end of the twentieth century. Currently regeneration is forming fir stands with life expectancies until the next drought. However, according to climate scenarios (Stocker et al. 2013) aridity increase in combination with catastrophic droughts is expected to be more frequent in the future, which may lead to elimination of Abies sibirica from part of its current range and its substitution by more tolerant species (e.g., Larix sibirica, Pinus sylvestris). A similar process is expected for Siberian pine (Kharuk et al. accepted) and Picea abies on the western part of the “dark needle conifer” range, for example, in Belarus forests (Kharuk et al. 2015a).
Fir mortality and biotic impacts
It is known that drought-weakened trees are the targets for insect and fungal attacks (e.g., Fettig et al. 2013). Fir mortality, as mentioned above, was located mainly on terrain with maximal water-stress risk. Declining and dead trees also exhibited signs of bark-beetle (Monochamus urussovi Fisch) attacks and larvae galleries within the bark and xylem, as well as fungal (Heterobasidion annosum) impacts. Thus, it appears likely that drought-stressed trees became susceptible to biotic impacts in subsequent years. Also, post-drought TRW decreases and mortality area increases (Figs. 3a, 4) indicate possible post-drought biotic impacts. Similarly, drought-induced spruce (Picea obovata) mortality in Belarus was accompanied by a bark-beetle outbreak and root fungi impacts (Sazonov et al. 2013). Thus, fungi and insect attacks seem to be covarying factors explaining fir mortality in Siberia, whereas increasing drought points toward water stress are the primary cause of decline.
The latter agrees with the current concept of multiple mechanisms of drought-induced mortality (i.e., hydraulic failure, xylem embolism, insect, fungi and bacterial attacks) (McDowell 2011; Choat et al. 2012; Anderegg et al. 2013; Fettig et al. 2013).
In general, “dark needle conifer” (Abies sibirica, Pinus sibirica and Picea obovata) mortality in Siberia was mainly observed within the margins of the DNC range, i.e., within the DNC-hardwoods and DNC-steppe ecotones. Within the interior regions of the DNC range, mortality was located on the relief features with maximum water stress where “diffuse-type” mortality (i.e., scattered individual trees or tree clusters) occurred. Under future climate scenarios (Hijioka et al. 2014), fir (and Siberian pine) is at risk to be eliminated from portions of their present area and to be substituted by drought-tolerant species (e.g., Pinus sylvestris, Larix sibirica).
The fir mortality in Siberia considered here is part of broader phenomenon of “dark needle conifer” decline and mortality in the European and Asian parts of Russia (Man’ko et al. 1998; Chuprov 2008; Zamolodchikov 2012; Sazonov et al. 2013). Notably, all reported areas of DNC mortality in Russia coincided with observed drought episodes or drought index increase. Along with fir, Siberian pine mortality was documented within the southern part of its range (Kharuk et al. 2013b, 2015b). Scenarios of climate changes project further increases in frequency and severity of droughts in some forested areas (e.g., Sterl et al. 2008; Anderegg et al. 2013; Stocker et al. 2013). This may lead to increases in DNC stress and mortality in synergy with projected increases in climate-related impacts from insects (Lloyd and Bunn 2007). On the other hand, climate warming opens opportunities for DNC migration northward into the larch-dominated regions and into alpine tundra (Kharuk et al. 2006, 2010).
Conclusion
Recent fir decline and mortality in southern Siberian Mountains are caused primary by water-stress increase and periodic drought episodes. Drought-weakened trees become more sensitive to bark beetles and fungi attacks; thus, the synergy of water-stress and biotic impact leads to stand mortality. We found that previous year ecological variables (SPEI, VPD, soil water anomalies) have a significant impact on the current tree ring growth. At geographical scale, fir mortality occurred within the transition zone between fir-dominant stands and forest-steppe. In a future climate with increased aridity and drought frequency fir (and Siberian pine) as a highly drought-sensitive species may be partly eliminated from its current range and substituted by drought-tolerant species (such as Pinus sylvestris and Larix sibirica).
References
Aitken SN, Yeaman S, Holliday JA, Wang T, Curtis-McLane S (2008) Adaptation, migration or extirpation: climate change outcomes for tree populations. Evol Appl 1(1):95–111. doi:10.1111/j.1752-4571.2007.00013.x
Allen CD, Breshears DD, McDowell NG (2015) On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 6(8):129. doi:10.1890/ES15-00203.1
Anderegg LDL, Anderegg WRL, Berry JA (2013) Tree physiology review: not all droughts are created equal: translating meteorological drought into woody plant mortality. Tree Physiol 33(7):701–712. doi:10.1093/treephys/tpt044
Arhipova NG (2013) On the problem of spruce (Picea abies (L.) Karst.) decline in Latvia. In: The problem of spruce stands decline. Reports of International seminar. Colorpoint Publishing House, Minsk [In Russian]
Breda N, Huc R, Granier A, Dreyer E (2006) Temperate forest trees and stands under severe drought: a review of ecophysiological responses, adaptation processes and long-term consequences. Ann For Sci 63:625–644. doi:10.1051/forest:2006042
Choat B, Jansen S, Brodribb TJ, Cochard H, Delzon S, Bhaskar R, Bucci SJ, Field TS, Gleason SM, Hacke UG, Jacobsen AL, Lens F, Maherali H, Martínez-Vilalta J, Mayr S, Mencuccini M, Mitchell PJ, Nardini A, Pittermann J, Pratt RB, Sperry JS, Westoby M, Wright IJ, Zanne AE (2012) Global convergence in the vulnerability of forests to drought. Nature 491:752–755. doi:10.1038/nature11688
Chuprov NP (2008) About problem of spruce decay in European North of Russia. Rus J For 1:24–26 [In Russian]
Cook ER, Holmes RL (1986) Users manual for program Arstan. In: Holmes RL, Adams RK, Fritts HC (eds) Tree-ring chronologies of western North America: California, eastern Oregon and northern Great Basin. Laboratory of Tree-Ring Research, University of Arizona, Tucson, pp 50–65
Fettig CJ, Reid ML, Bentz BJ, Sevanto S, Spittlehouse DL, Wang T (2013) Changing climates, changing forests: a western North American perspective. J For 111(3):214–228. doi:10.5849/jof.12-085
Fritts HC (1991) Reconstruction large-scale climatic patterns from tree-ring data: a diagnostic analysis. University of Arizona Press, Tucson
Guarın A, Taylor AH (2005) Drought triggered tree mortality in mixed conifer forests in Yosemite National Park, California, USA. For Ecol Manag 218:229–244. doi:10.1016/j.foreco.2005.07.014
Hijioka Y, Lin E, Pereira JJ, Corlett RT, Cui X, Insarov GE, Lasco RD, Lindgren E, Surjan A (2014) Asia. In: Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part B: Regional Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change [Barros, V.R., C.B. Field, D.J. Dokken, M.D. Mastrandrea, K.J. Mach, T.E. Bilir, M. Chatterjee, K.L. Ebi, Y.O. Estrada, R.C. Genova, B. Girma, E.S. Kissel, A.N. Levy, S. MacCracken, P.R. Mastrandrea, and L.L.White (eds.)]. Cambridge University Press, Cambridge, New York, NY, pp 1327–1370
Holmes RL (1983) Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull 44:69–75
Kagawa A, Sugimoto A, Maximov TC (2006) Seasonal course of translocation, storage and remobilization of 13C pulse-labeled photoassimilate in naturally growing Larix gmelinii saplings. N Phytol 171:793–804. doi:10.1111/j.1469-8137.2006.01780.x
Kharuk VI, Ranson KJ, Im ST, Dvinskaya ML (2006) Forest-tundra larch forests and climatic trends. Rus J Ecol 37(5):291–298. doi:10.1134/S1067413606050018
Kharuk VI, Im ST, Dvinskaya ML, Ranson KJ (2010) Climate-induced mountain treeline evolution in southern Siberia. Scand J For Res 25(5):446–454. doi:10.1080/02827581.2010.509329
Kharuk VI, Im ST, Oskorbin PA, Petrov IA, Ranson KJ (2013a) Siberian pine decline and mortality in southern Siberian mountains. For Ecol Manag 310:312–320. doi:10.1016/j.foreco.2013.08.042
Kharuk VI, Ranson KJ, Oskorbin PA, Im ST, Dvinskaya ML (2013b) Climate induced birch mortality in trans-Baikal lake region, Siberia. For Ecol Manag 289:385–392. doi:10.1016/j.foreco.2012.10.024
Kharuk VI, Im ST, Dvinskaya ML, Golukov AS, Ranson KJ (2015a) Climate-induced mortality of spruce stands in Belarus. Environ Res Lett. doi:10.1088/1748-9326/10/12/125006
Kharuk VI, Ranson KJ, Im ST, Petrov IA (2015b) Climate-induced larch growth response within central Siberian permafrost zone. Environ Res Lett. doi:10.1088/1748-9326/10/12/125009
Kirillov MV, Shherbakov YuA (eds) (1961) Krasnoyarsk region. Krasnoyarsk publishing house, Krasnoyarsk [In Russian]
Krylov GV (1961) Forests of West Siberia. AN SSSR, Moscow [In Russian]
Lausch A, Heurich M, Fahse L (2013) Spatio-temporal infestation patterns of Ips typographus (L.) in the Bavarian Forest National Park, Germany. Ecol Indic 31:73–81. doi:10.1016/j.ecolind.2012.07.026
Lloyd AH, Bunn AG (2007) Responses of the circumpolar boreal forest to 20th century climate variability. Environ Res Lett 2(4): 045013. doi: 10.1088/1748-9326/2/4/045013. http://iopscience.iop.org/1748-9326/2/4/045013. Accessed 22 Apr 2014
Long D, Longuevergne L, Scanlon BR (2014) Uncertainty in evapotranspiration from land surface modeling, remote sensing, and GRACE satellites. Water Resour Res 50(2):1131–1151
Man’ko YI, Gladkova GA, Butovets GN, Kamibayashi N (1998) An experience of monitoring fir-spruce forest decline in the central Sikhote–Alin. Rus J Forestry 1:3–16 [In Russian]
Martínez-Vilalta J, Lloret F, Breshears DD (2012) Drought-induced forest decline: causes, scope and implications. Biol Lett 8(5):689–691. doi:10.1098/rsbl.2011.1059
McDowell NG (2011) Mechanisms linking drought, hydraulics, carbon metabolism, and vegetation mortality. Plant Physiol 155:1051–1059. doi:10.1104/pp.110
Pavlov IN, Ruhullaeva OV, Barabanova OA, Ageev AA (2008) Estimation of root pathogens impact on forest resources of Siberian federal district. Boreal Zone Conifers 3(4):262–268 [In Russian]
Rautiainen M, Heiskanen J, Korhonen L (2012) Seasonal changes in canopy leaf area index and MODIS vegetation products for a boreal forest site in central Finland. Boreal Environ Res 17:72–84
Rinn F (1996) TSAP V 3.6 Reference manual: computer program for tree-ring analysis and presentation. Frank Rinn, Heidelberg
Russian Federal Agency of Forestry (2013) Annual report about state and exploitation of forests in Russian Federation in 2012. http://www.rosleshoz.gov.ru/docs/other/79. Accessed 28 July 2015 [in Russian]
Sazonov AA, Kuhta VN, Blinzov AI, Zvyaginzev VB, Ermohin MV (2013) The problem of large-scale spruce mortality in Belarus, and the ways of its solving. For Wildl 7:10–15 [In Russian]
Kharuk VI, Im ST, Petrov IA. “Dark needle conifer” decline in the Baikal Lake region. Contemp probl ecol (accepted)
StatSoft Inc. (2013) Electronic statistics textbook. http://www.statsoft.com/textbook. Accessed 29 May 2015
Stephenson NL (1990) Climatic control of vegetation distribution: the role of water balance. Am Nat 135:649–670. doi:10.1086/285067
Sterl A, Severijns C, Dijkstra H, Hazeleger W, van Oldenborgh GJ, van den Broeke M, Burgers G, van den Hurk B, van Leeuwen PJ, van Velthoven P (2008) When can we expect extremely high surface temperatures? Geophys Res Lett 35(14):L14703. doi: 10.1029/2008GL034071. http://onlinelibrary.wiley.com/doi/10.1029/2008GL034071/pdf. Accessed 22 Apr 2012
Stocker TF, Qin D, Plattner G-K, Tignor M, Allen SK, Boschung J, Nauels A, Xia Y, Bex V, Midgley PM (eds) (2013) Climate change 2013: the physical science basis. Contribution of working group I to the fifth assessment report of the intergovernmental panel on climate change. Cambridge University Press, Cambridge and New York. http://www.climatechange2013.org. http://www.ipcc.ch. Accessed 22 Apr 2014
Utkin AI (1975) Biological productivity of forests: study methods and results. For Silvic 1:9–189 [In Russian]
Vasilyauskas V (2013) Main causes of spruce decline in Lithuania. In: The problem of spruce stands decline. Reports of International seminar. Colorpoint Publishing House, Minsk. p 6–21. [In Russian]
Vicente-Serrano SM, Beguería S, López-Moreno I (2010) A multiscalar drought index sensitive to global warming. The standardized precipitation evapotranspiration index. J Clim 23:1696–1718. doi:10.1175/2009JCLI2909.1
Worrall JJ, Marchetti SB, Egeland L, Mask RA, Eager T, Howell B (2010) Effects and etiology of sudden aspen decline in southwestern Colorado USA. For Ecol Manag 260(5):638–648. doi:10.1016/j.foreco.2010.05.020
Yousefpour R, Hanewinkel M, Le Moguédec G (2010) Evaluating the suitability of management strategies of pure norway spruce forests in the black forest area of southwest Germany for adaptation to or mitigation of climate change. Environ Manag 45(2):387. doi:10.1007/s00267-009-9409-2
Zamolodchikov DG (2012) An estimate of climate related changes in tree species diversity based on the results of forest fund inventory. Biol Bull Rev 2(2):154–163. doi:10.1134/S2079086412020119
Acknowledgements
Authors thank anonymous reviewers for valuable comments. This research was supported by Russian Science Fund (RNF) Grant (No. 14-24-00112). K. J. Ranson’s contribution was supported in part by the NASA’s Terrestrial Ecology Program.
Funding
This study was supported by Russian Science Fund (RNF) (Grant No. 14-24-00112). K. J. Ranson’s contribution was supported in part by the NASA’s Terrestrial Ecology Program.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Editor: Christopher Reyer.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Kharuk, V.I., Im, S.T., Petrov, I.A. et al. Fir decline and mortality in the southern Siberian Mountains. Reg Environ Change 17, 803–812 (2017). https://doi.org/10.1007/s10113-016-1073-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10113-016-1073-5