
Abstract
Whereas most studies on overmarking in mammals analysed the rate of overmarking, that those investigate time between exploration of an olfactory stimulus and the response to it remain less common, with inconsistent results. We examined the latency in time between elimination by the sender and sniffing by the receiver, and from sniffing and overmarking, in four captive African equid species to explore differences among species, and among age and sex classes. We investigated these latency time periods in light of three potential hypotheses explaining overmarking behaviour in equids: social bonds, group cohesion, and intrasexual competition. Analysing 1684 events of sniffing and 719 of overmarking among 130 individuals, we found that (i) the time from elimination to overmarking was shorter among female friends and in parent–offspring dyads, proving support to the social bond hypothesis; (ii) intraspecific differences in time periods do not reflect the social organisation of species, thus not supporting the group cohesion hypothesis; (iii) males were more attracted to elimination of conspecifics than females, and female’s eliminations were inspected longer, in line with the sexual competition hypothesis and/or reproductive behaviour. In addition, we found that the younger foals came to sniff eliminations faster than older ones, and in larger groups foals devoted longer time to sniffing the elimination before overmarking. We concluded that examination of the elimination could be driven by motivations other than the decision to overmark. Whereas overmarking serves to express bonds to a familiar individual, the latency of overmarking reflects more reproductive interests.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Scent plays an important role in intraspecific recognition for social animals across taxa: in insects (Dreier et al. 2007), reptiles (Font et al. 2012), birds (Leclaire et al. 2017), and mammals (Ralls 1971). Most mammals have a highly developed olfactory sense which they employ in social communication (Ralls 1971). Volatile chemicals with information on factors including sex and reproductive status are deposited with urine and faeces (Marneweck et al. 2017), and then further intraspecific communication occurs through overmarking of these scents, i.e. applying chemical signals on a deposit of another individual (Ralls 1971; Johnson 1973; Ferkin and Pierce 2007).
Various functions of overmarking have been suggested, resulting in thirteen potential hypotheses (Ralls 1971; Johnson 1973; Ferkin and Pierce 2007; Tučková et al. 2018; Pluháček et al. 2019b). These hypotheses state that overmarking (1) may represent intraspecific competition; (2) be a form of self-advertisement; (3) indicate territorial behaviour; (4) be used to attract mates; (5) provide a means to increase foraging efficiency; (6) be a navigation guide; (7) develop a group cohesion scent; (8) act as a threat behaviour; (9) conceal scent; (10) be part of parent–offspring conflict; (11) act to strengthen social bonds; (12) form a means to identify conspecifics; and (13) simply be an artefact of laboratory studies. Some of these hypotheses are not mutually exclusive (Ferkin and Pierce 2007). It should also be noted that marking behaviour is likely to have more than one function in any one species and different functions in different species (Johnson 1973). The most supported explanation remains intrasexual competition, especially in ungulates (Turner et al. 1981; Penzhorn 1984; Kimura 2000; Marneweck et al. 2019).
When overmarking has been described, most studies focus on the individuals involved, i.e., who is overmarked by whom and the rate of overmarking among them (Brashares and Acrese 1999; Ferkin et al. 2001, 2004; Heymann 1998; Wolf et al. 2002; Wronski et al. 2013). However, some studies also investigated the latency of overmarking once a stimulus was detected (Kappeler 1998; Krueger and Flauger 2011) or latency from marking to sniffing the overmark (Jordan 2007; Jordan et al. 2011; Linklater et al. 2013; Marneweck et al. 2017). Many studies reported sexual differences in overmarking latency or sniffing (Blank et al. 2015; Black-Decima and Santana 2011; Kappeler 1998; Jordan 2007). However, results found across species are inconsistent: whereas in brown brocket deer (Mazama gouazoubira) males overmark with a shorter latency than females (Black-Decima and Santana 2011), the opposite was found in meerkats (Suricata suricatta; Jordan 2007) and there was no difference in white rhinos (Ceratotherium simum; Marneweck et al. 2018). On the other hand, females of the latter species investigated scent marks longer than males. In ring-tailed lemur (Lemur catta), the eliminations (i.e., urine or faeces) of females are overmarked more quickly than those of males; however the sex of the overmarking animal does not affect the latency (Kappeler 1998).
Other studies reported a difference in the time of exploring an elimination based on the type of elimination (Linklater et al. 2013), familiarity between individuals (Black-Decima and Santana Black-Décima and Santana 2011; Cinková and Policht 2015; Jordan et al. 2011), or the oestrus status of the female (Marneweck et al. 2017). In general, it is believed that longer sniffing and shorter latency of overmarking represents higher interest in investigation of the elimination and higher importance of overmarking, respectively (Kappeler 1998; Krueger and Flauger 2011).
Equids represent a good group for studying overmarking for several reasons. First, they differ from many other mammals as they have no dedicated scent glands and therefore mark only with faeces and/or urine (Brown and Macdonald 1985). Second, their overmarking rate is quite frequent (Kimura 2000; Moehlman 1985; Tučková et al. 2018). Third, they are congregatory with variability in their social system (Klingel 1975; Linklater 2013) that is reflected in differences in overmarking (Kimura 2000; Pluháček et al. 2020). Fourth, they represent a unique group of mammals where overmarking is expressed in very young juveniles at an unprecedently high rate (Pluháček et al. 2019b, 2020). Most prior studies on equids favoured the intrasexual competition hypothesis as the main reason for overmarking (Turner et al. 1981; Penzhorn 1984; Kimura 2000). However, recently we provided support for two different hypotheses: social bonds and group cohesion, and found no evidence for intrasexual competition (Tučková et al. 2018; Pluháček et al. 2019a, b). Additionally, we revealed an early expression of male sexual behaviour as a possible explanation of overmarking in male juveniles and subadults (Pluháček et al. 2019b).
In equids, most overmarking is in response to elimination of a conspecific (King and Gurnell 2007). When an individual encounters an elimination they either ignore it or sniff it. After sniffing they may further explore the scent through flehmen, where the odour is drawn through the vomeronasal organ. After exploration by sniffing and/or flehmen the individual may overmark with urine or faeces, or subsequently ignore the stimulus (King and Gurnell 2007; Tučková et al. 2018; Pluháček et al. 2019a, b). Although some studies have investigated the time from exploring (sniffing) a stimulus elimination to overmarking, few have examined time between the elimination being deposited to a conspecific exploring it. Latency of time between deposit of a stimulus and exploration by a conspecific may be related to its attractiveness. As described above, overmarking in equids typically follows the pattern of elimination (stimulus), sniffing (exploration), followed by overmarking. We examined the time between elimination and subsequent steps in this pattern during overmarking events in four captive African equid species to explore differences among species, and among age and sex classes. Specifically, we measured time from elimination by a conspecific to sniffing (TES), and time from sniffing to overmarking (TSO). We also measured overall time from elimination to overmarking (TEO), but as this period was correlated with TES and TSO we did not include it into our analyses. We examined the two periods (TES and TSO) in light of three potential hypotheses explaining overmarking behaviour in equids: (1) social bonds, (2) group cohesion, and (3) intrasexual competition. In line with these hypotheses we predicted that latency should be shorter in (1) friends and mother–offspring pairs, (2) species with less stable group social structure (African wild ass [Equus africanus], Grévy’s zebra [E. grevyi]) than those living in stable groups (mountain zebra [E. zebra] and plains zebra [E. quagga]), and (3) after elimination of adult females and when overmarked by males. In addition, we examined whether any of the time periods were affected by factors such as reproductive or social status, or environmental conditions.
Methods
Animals
We observed four equid species (African wild ass, Grevy’s zebra, plains zebra, and mountain zebra) in five zoos (Brno, Dvůr Králové, Liberec, Ostrava, and Ústí nad Labem) in the Czech Republic. In total, our observations involved 130 individuals including 18 subadults and 43 foals maintained in 15 herds (Table 1). Herd size varied from 3 to 18 individuals and each observed herd included one to seven foals. Foals were defined as dependent juveniles still actively suckling from the mother (Pluháček et al. 2007). For definition of age categories see Tučková et al. (2018). All herds were observed in outdoor enclosures which varied in size from 800 to 2800 m2. Enclosures were covered mostly by gravel with some patches of grass, concrete, sand or soil, with the exception of the enclosure of Grévy’s zebra at Ostrava Zoo and one herd of plains zebra at Dvůr Králové Zoo (#3 in Table 1) which were predominantly covered by grass. Food (grass or hay in feeders) and water was available ad libitum for each herd. Each enclosure was cleaned once a day in the morning before the start of the observation.
Observations were made over 2 years, in 2010 from 23rd June to 25th October and in 2011 from 18th May to 19th September. We performed two observation sessions per day, either from 08:00 to 11:00 or from 14:00 to 17:00; no group was observed twice in the same day. Each group in each year was observed five times a week during any 2 weeks per year. However, these 2 weeks were not consecutive. Thus, we performed ten observation sessions per herd per season. In total, we carried out 600 h of observation (200 sessions over 121 days). In each observation session, we recorded data concerning elimination events (i.e., defecation and urination) within the herd using the ad libitum sampling method (Altmann 1974). This included recording the identity of the eliminator, behaviour of the eliminator prior to the elimination, identity of the individual examining the elimination (i.e., sniffing), and if flehmen and overmarking was observed we recorded details of this individual and the sequence of behaviours. We used the same definition of overmarking as described by other researchers (Turner et al. 1981; Kimura 2000, 2001).
Since overmarking behaviour is highly affected by substrate (Przewalski horses Equus przewalskii: King and Gurnell 2007; all three zebra species: Pluháček et al. 2019b), we included these environmental factors in our analyses. We distinguished whether eliminations occurred near the edge of the enclosure (within 1 m from the fence or wall), whether it occurred under shelter, and the type of surface where it was deposited (concrete, grass, gravel, sand, and soil).
Statistics
All data were analysed using the SAS System, Version 9.4 (SAS Institute, Inc. Cary, NC). We tested factors affecting time from elimination by a conspecific to sniffing (TES) and time from sniffing to overmarking (TSO) using multivariate generalised linear mixed models (GLMM, PROC MIXED, SAS). To approach normal distribution, all dependent variables were log transformed. To avoid the effect of outliers, we excluded values exceeding 180 s (n = 12) in all analyses of TES, and 180 s (n = 11) in all analyses of TSO.
We included the following fixed factors: type of elimination (urination/defecation), sex and age category of the eliminator (adult males/adult females/subadult males/subadult females/male foals/female foals), sex and age category of the animal that examined the elimination, sex of the eliminator, sex of the examining animal, age of the eliminator, age of the examining animal, species, zoo, number of animals within the herd (3–18), number of foals within the herd (1–7), presence of stallion in the herd (yes/no), type of surface (concrete, grass, gravel, sand, or soil), shelter (yes/no), outside temperature, whether the elimination was deposited at the edge of the enclosure (yes /no), whether elimination was on the latrine (yes/no; the latrine was a spot in the enclosure frequently used for elimination although cleaned by keepers every day), pregnancy (yes/no), lactating (yes/no), and oestrus (yes/no) status of the eliminator, pregnancy (yes/no), lactating (yes/no), and oestrus (yes/no) status of the animal examining the elimination, behaviour of the eliminator preceding the elimination (walking/standing/feeding/other), whether the examiner was a parent of the eliminator (yes/no), whether the examiner was offspring of the eliminator (yes/no), and flehmen of the animal examining elimination (yes/no). For the oestrus, lactating, and pregnant variables males and foals were coded as ‘no’. In analyses limited to adult females we also included friendship in the model (friends/middle-friends/non-friends; see Tučková et al. (2018) for details of this distinction). Random factors in the models were identity of the eliminating animal and the year of observation (2010/2011). We tested for intercorrelation of factors and found that sex and age category correlated with sex and age, thus these were not tested in the same model.
Since the explanation of the phenomenon might differ across sex and age categories, we ran GLMMs for each time period (TES, TESFO [time from elimination to sniffing for cases where followed overmarking], TSO) first for all animals, then separately for adult females, adult males, and foals as the exploring individual. In all models we avoided pseudoreplication of the same animal by entering the individual that examined the elimination as a subject in the ‘repeated’ statement. We started with a full model including all fixed effects. We assessed the significance of each fixed effect in the mixed model via F – test paying special attention to hypotheses predictors (age and sex category, species parenthood, friendship). Then we dropped non-significant fixed effects sequentially to identify the model with the lowest AICc value (Tables A1, A2, and A3). Only the factors that were statistically significant are given in the results section. The within-group means were appropriately adjusted for the other effects in the model (LSMEANs statement). Differences between means were tested with t – tests; for multiple comparisons we used the Tukey–Kramer adjustment.
Results
We recorded 4668 eliminations and 1684 cases of reactions to eliminations (i.e., sniffing), with at least one case of reaction from each sex and age category per species (when present; Table 2). In 746 cases, the elimination was overmarked after being sniffed (Table 2).
Correlations among latency time periods
Both TES and TSO were correlated with TEO (TES: Spearman: r = 0.62; P = P < 0.0001; TSO: Spearman: r = 0.61; P < 0.0001). However, TES and TSO were not correlated (Spearman r = 0.08; NS).
Time from elimination to sniffing (TES)
All individuals
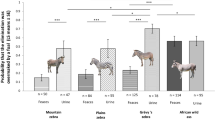
The longest TES we recorded was 32 min and 30 s. Average TES was 28.41±78.01 s (n = 1684 events). TES was affected by age and sex of the animal that examined the elimination (F = 3.92; df = 6, 1510; P = 0.0007; Fig. 1), species (F = 3.84; df = 3, 1510; P = 0.0094; Fig. 2), lactation status of the eliminator (F = 19.83; df = 1, 1510; P < 0.0001), oestrous status of the eliminator (F = 6.72; df = 1, 1510; P = 0.0096), parenthood (F = 5.12; df = 1, 1510; P = 0.0238), and zoo (F = 4.44; df = 4, 1510; P = 0.0014). Elimination of an oestrus or lactating female was sniffed sooner than that of other females, and parents were quicker to sniff their offspring’s eliminations than a non-parent. Grévy’s zebra (t = 3.32; P = 0.0031) and plains zebra (t = 2.59; P = 0.0429) were quicker to sniff an elimination than African wild ass. Male foals sniffed sooner than adult (t = 4.29; P = 0.0004) and subadult females (t = 3.00; P = 0.0429).

Time from elimination (defecation or urination) to sniffing of the elimination (TES) in four African equid species (Equus africanus, E. grevyi. E. quagga, E zebra) at five zoos in the Czech Republic according to the age and the sex of the animal that examined the elimination. The values represent LS means ± SE of means; n refers number of cases involved; significance levels are represented by: *P < 0.05, ***P < 0.001

Time from elimination (defecation or urination) to sniffing of the elimination (TES) in four African equid species (Equus africanus, E. grevyi. E. quagga, E zebra) at five zoos in the Czech Republic according to the species. The values represent LS means ± SE of means; n refers number of cases involved; significance levels are represented by: *P < 0.05, **P < 0.01
Adult females
TES of adult females was affected by parenthood (F = 5.25; df = 1, 612; P = 0.0223): TES was longer when a mother sniffed than another female. We also found an effect of friendship on TES (F = 7.07; df = 2, 220; P = 0.0011) with best friends sniffing faster than non-friends (t = 3.61; P = 0.0011; Fig. 3).

Time from elimination (defecation or urination) to sniffing of the elimination (TES) by females of four African equid species at five zoos in the Czech Republic according to the friendship (Tučková et al. 2018). The values represent LS means ± SE of means; n refers number of cases involved; significance level is represented by: **P < 0.01
Adult males
TES of adult males was affected by species (F = 9.44; df = 2, 228; P = 0.0001), latrine occurrence (F = 7.55; df = 1, 228; P = 0.0065), and by parenthood (F = 4.22; df = 1, 228; P = 0.0410). TES was longer when the male was the father, when the elimination was on the latrine than elsewhere, and in mountain zebra (t = 4.04; P = 0.0002) and Grévy’s zebra (t = 2.43; P = 0.0417) than in plains zebra.
Foals
TES of foals was affected by species (F = 4.78; df = 3, 590; P = 0.0022), oestrus status of the eliminator (F = 9.15; df = 1, 590; P = 0.0026), the age of the foal in days (F = 7.68; df = 3, 590; P = 0.0047, and offspring-parent dyad (F = 5.26; df = 1, 590; P = 0.0221). The TES was shorter when the eliminating animal was in oestrus than when not in oestrus, and when it was a parent of the foal. Younger foals sniffed the elimination faster than older ones (Fig. 4). Grévy’s zebra foals sniffed the elimination faster than African wild ass (t = 3.41; P = 0.0038).

Time from elimination (defecation or urination) to sniffing of the elimination (TES) by foals of four African equid species at five zoos in the Czech Republic according to the age of the foal
Time from elimination to sniffing for situations where overmarking followed only (TESFO)
When we limited the TES only to situations where overmarking followed the sniffing (TESFO) we found that this time was affected by age and sex category of the animal that eliminated (F = 2.81; df = 5, 724; P = 0.0160), parenthood (F = 7.90; df = 1, 724; P = 0.0051), offspring (F = 45.82; df = 1, 724; P < 0.0001), and the zoo (F = 3.05; df = 1, 724; P = 0.0164). TESFO was shorter when a parent inspected the elimination of the offspring and when offspring examined elimination of parent than when other herdmates were involved.
Adult females
When we analysed the TESFO by adult females only, we found that mothers inspected an elimination faster than non-mothers (F = 7.14; df = 1, 261 P = 0.0080). Friendship also reached the level of significance (F = 4.29; df = 2, 78; P = 0.0170) with faster inspection by friends than by non-friends (t = 2.97; P = 0.0109). In addition, oestrus females inspected the elimination of the other female faster than non-oestrus ones (F = 5.47; df = 2, 78; P = 0.0219).
Adult males
When we limited data to eliminations sniffed by adult males only, we found that the TESFO was affected by species (F = 6.75; df = 2, 117; P = 0.0017; longer latency in mountain zebra than in plain zebra: t = 3.37; P = 0.0030), and when at the latrine (F = 5.45; df = 1, 117; P = 0.0213).
Foals
When we limited data to eliminations sniffed by foals only, we found that the TESFO was shorter when the eliminating animal was parent of the foal (F = 42.86; df = 1, 224; P < 0.0001) and when the elimination was not at the edge of the enclosure (F = 5.22; df = 1, 224; P = 0.0233).
Time from sniffing to overmarking (TSO)
All individuals
The longest TSO we recorded was 634 s, with the average lasting 20.74±24.42 s (n = 630 events). When sniffing was accompanied by flehmen, TSO was longer (F = 27.66; df = 1, 514; P < 0.0001; mean with flehmen: 25.81±21.00 s [n = 78 events], mean without flehmen 19.90±24.85 s [n = 552 events]).
Since flehmen strongly affected TSO, we performed separate analyses for the TSO with and without flehmen. When we analysed only those events when the overmarking animal did perform flehmen (n = 78), the TSO was affected by species (F = 2.94; df = 3, 50; P = 0.0418). TSO lasted longer in mountain zebra than in African wild ass (t = 2.95; P = 0.0236).
When events without flehmen (n = 552) were tested, TSO was affected by surface only (F = 3.41; df = 4, 435; P = 0.0093). Zebras and asses overmarked eliminations faster on gravel than on soil (t = 3.11; P = 0.0172).
Adult females
No factor was significant in the final model for adult females.
Adult males
For males, TSO was affected by sex of the eliminating animal (F = 5.45; df = 1, 63; P = 0.0277) with longer TSO after sniffing elimination of females than males.
Foals
TSO increased with increasing herd size for foals (F = 5.17; df = 1, 167; P = 0.0242; Fig. 5).

Time from sniffing an elimination (i.e., faeces or urine) to overmarking it (TSO) by foals of four African equid species according to herd size at five Czech zoos
Discussion
As far as we are aware this is the first observational study that examined the time periods from elimination to overmarking in equids in detail. In addition, we analysed several time periods whereas other studies simply focused on the time from sniffing to overmarking (Marinier et al. 1988; Krueger and Flueger 2011; Hothersall et al. 2010; Linklater et al. 2013; Ghosal et al. 2012; Jezierski et al. 2015). We found that the time from elimination to sniffing (TES) and time from sniffing to overmarking (TSO) were affected by various factors which suggests that these times reflect different needs.
Social bond hypothesis
We found support for the social bond hypothesis of the function of overmarking as TES was shorter among female friends and parent–offspring pairs compared to the rest of the group (Tučková et al. 2018; Pluháček et al. 2019b). The same result was found when only cases of examination that ended by overmarking were included (TESFO). On the other hand, we did not find that friendship and parent–offspring relationship affected TSO; similarly other mammals spent a shorter time inspecting the elimination of familiar individuals than non-familiar (Johnston et al. 1994). Faster TES might indicate either higher attraction of the elimination or may be just associated with closer spatial proximity. Since friendship was defined by proximity in scan samples (Tučková et al. 2018), and the mother closely accompanies the offspring (Tyler 1972), it seems that this explanation is parsimonious. Nevertheless, we found that male foals approached the elimination of other individuals faster than adult females which shows that attractiveness of the elimination played a role as well. In addition, in captive Przewalski’s horses, the decision to examine the elimination and /or to overmark it was not affected by the distance between the two individuals involved (Boyd and Kasman 1986). As it is not possible to disentangle proximity and attractiveness of the elimination of a friend, mother, or offspring in TES we cannot reject the social bond hypothesis.
Group cohesion hypothesis
Zebras reached the elimination to smell it (TES) faster than African wild ass. On the other hand, almost no interspecific differences were found in TSO. These differences seem to reflect phylogenetic differences between asses and zebras (Steiner and Ryder 2011; Steiner et al. 2012) rather than differences in social organisation and thus group stability of the respective species (Joubert 1972; Klingel 1969, 1975, 1977; Ginsberg 1989). Shorter TES of plains zebra could reflect the fact that the individuals form compact groups even in the zoo environment. Since mountain zebras keep longer inter-individual distances than other species (Joubert 1972), the longer latency of time from elimination to sniffing in mountain zebra compared to plains zebra could reflect the longer distance for the male to explore it. Thus, based on our results we did not find any association between latency of overmarking and the group cohesion hypothesis.
Sexual selection hypothesis
Our analyses revealed several differences in latency based either on the sex of the eliminator or the sex of the examining individual, in line with other studies (Black-Decima and Santana 2011; Hothersall et al. 2010; Krueger and Flauger 2011; Marinier et al. 1988). Male foals came to an elimination (TES) faster than subadult and adult females. Males spent more time examining eliminations of females than males (TSO) as reported for other mammals like African elephant (Meyer et al 2008). These results might indicate that males are more attracted to eliminations of other animals and that they devote more time to inspect the elimination than females (Hothersall et al. 2010; Marinier et al. 1988).
These results infer that examination of an elimination is part of sexual competition, especially among males. In our previous study, we found that overmarking by male foals might reflect early expression of sexual behaviour (Pluháček et al. 2019b). We would like to note that whereas the decision to overmark an elimination cannot be associated with intra-male sexual competition, the exploration of the elimination itself by males could be associated with their reproductive behaviour (Kimura 2001; Marinier et al. 1988; Jordan 2007; Jordan et al. 2011). Thus, our data suggest that the latency of overmarking was associated with the sexual competition hypothesis.
Further, we found that the time from sniffing to overmarking by foals increased with increasing group size. This means that in larger groups foals devoted more time to inspecting an elimination before overmarking than when in smaller groups. We suggest that this behaviour might demonstrate that the foal needs to learn recognition of the eliminating animal and this is more difficult in a larger group, thus prolonging the time to overmarking. This also provides indirect evidence that equids need to learn recognition of other individuals from urine and faeces. Since foals might use overmarking of other foals to strengthen social bonds (Pluháček et al. 2020), longer TSO in larger groups could simply reflect the fact that the larger groups included a higher number of foals. Thus, foals may spend more time examining eliminations of other foals to better detect “friends”. However, the number of foals in the group was not associated with TSO. Therefore, it seems that a longer examination time of an elimination before overmarking by foals could be associated more with ontogeny of recognition than with any specific hypothesis explaining overmarking. This suggestion is in line with our other finding that foals reached elimination of another animal faster when they were younger. Younger foals might be more interested and more explorative of other herdmates’ eliminations than older foals who are more familiar with their odours. On the other hand, we cannot exclude that this result is affected by close proximity of the mother in young foals (Tyler 1972).
TSO was highly affected by the performance of flehmen. Although flehmen is often associated with overmarking (Turner et al. 1981; Moehlman 1985) the examination of the elimination via flehmen might be driven by different motivation than the decision to overmark. For example, males might examine most eliminations of females but they do not need to overmark all of them; in many cases flehmen of the elimination was not followed by overmarking (Pluháček et al. in prep.). Thus, even though the latency of overmarking is affected by flehmen, these two behaviours could be partially independent and follow different motivations.
In conclusion, our results provided partial support for two of the three non-exclusive hypotheses explaining overmarking: social bonds, and especially sexual competition. On the other hand, we found no support for the group cohesion hypothesis. In addition, it seems that examination of an elimination could be driven by motivations other than the decision to overmark. Whereas overmarking might serve to express affiliation to a familiar individual, the latency of overmarking, and especially TSO reflecting the examination of elimination, represents interest in the other animal mostly for reproductive reasons. Based on our results, future studies (both observational and experimental) on this topic should distinguish between interpretation of overmarking of an elimination and time examining the elimination.
References
Altmann J (1974) Observational study of behaviour: sampling methods. Behaviour 49:227–267
Black-Décima P, Santana M (2011) Olfactory communication and counter-marking in brown brocket deer Mazama gouazoubira. Acta Theriol 56:179–187
Blank D, Ruckstuhl K, Yang WK (2015) The economics of scent marking with urine and feces in goitered gazelle (Gazella subgutturosa). Mammal Res 60:51–60
Boyd L, Kasman L (1986) The marking behavior of male Przewalski’s horses. In: Duvall D, Muller-Schwarze D, Silverstein RM (eds) Chemical signals in vertebrates, ecology, evolution, and comparative biology, vol IV. Plenum Press, New York, pp 623–626
Brashares JS, Arcese P (1999) Scent marking in a territorial African antelope: II. The economics of marking with faeces. Anim Behav 57:11–17
Brown RE, Macdonald DW (1985) Social odours in mammals, vol 2. Clarendon Press, Oxford
Cinková I, Policht R (2015) Discrimination of familiarity and sex from chemical cues in the dung by wild southern white rhinoceros. Anim Cogn 18:385–392
Dreier S, van Zweden JS, D’Ettorre P (2007) Long-term memory of individual identity in ant queens. Biol Lett 3:459–462
Ferkin MH, Pierce AA (2007) Perspectives on over-marking: is it good to be on top? J Ethol 25:107–116
Ferkin MH, Leonard ST, Bartos K, Schmick MK (2001) Meadow voles and prairie voles differ in the length of time they prefer the top-scent donor of an over-mark. Ethology 107:1099–1114
Ferkin MH, Lee DN, Leonard ST (2004) The reproductive state of female voles affects their scent marking behavior and the responses of male conspecifics to such marks. Ethology 110:257–272
Font E, Barbosa D, Sampedro C, Carazo P (2012) Social behavior, chemical communication, and adult neurogenesis: studies of scent mark function in Podarcis wall lizards. Gen Comp Endocrinol 177:9–17
Ghosal R, Seshagiri PB, Sukumar R (2012) Dung as a potential medium for inter-sexual chemical signaling in Asian elephants (Elephas maximus). Behav Proc 91:15–21
Ginsberg JR (1989) The ecology of female behaviour and male reproductive success in the Grevy’s zebra, Equus grevyi. In: Jewell PA, Maloiy GMO (eds) Symposia of the zoological society of London. Clarendon Press, pp 89–110
Heymann EW (1998) Sex differences in olfactory communication in a primate, the moustached tamarin, Saguinus mystax (Callitrichinae). Behav Ecol Sociobiol 43:37–45
Hothersall B, Harris P, Sortoft L, Nicol CJ (2010) Discrimination between conspecific odour samples in the horse (Equus caballus). Appl Anim Behav Sci 126:37–44
Jezierski T, Jaworski Z, Sobczynska M, Kaminska B, Gorecka-Bruzda A, Walczak M (2015) Excreta-mediated olfactory communication in Konik stallions: a preliminary study. J Vet Behav Clin Appl Res 10:353–364
Johnson RP (1973) Scent marking in mammals. Anim Behav 21:521–535
Johnston RE, Chiang G, Tung C (1994) The information in scent over-marks of golden hamsters. Anim Behav 48:323–330
Jordan NR (2007) Scent-marking investment is determined by sex and breeding status in meerkats. Anim Behav 74:531–540
Jordan NR, Mwanguhya F, Furrer RD, Kyabulima S, Ruedi P, Cant MA (2011) Scent marking in wild banded mongooses: 2. Intrasexual overmarking and competition between males. Anim Behav 81:43–50
Joubert E (1972) The social organisation and associated behaviour in the Hartmann zebra Equus zebra hartmannae. Madoqua Ser I 6:17–56
Kappeler PM (1998) To whom it may concern: the transmission and function of chemical signals in Lemur catta. Behav Ecol Sociobiol 42:411–421
Kimura R (2000) Relationship of the type of social organization to scent-marking and mutual-grromin behaviour in Grevy’s (Equus grevyi) and Grant’s zebra (Equus burchelli bohmi). J Equi Sci 11:91–98
Kimura R (2001) Volatile substances in feces, urine and urine/marked feces of feral horses. Can J Anim Sci 81:411–420
King SRB, Gurnell J (2007) Scent-marking behaviour by stallion: an assessment of function in a reintroduced population of Przewalski horses (Equus ferus przewalskii). J Zool 272:30–36
Klingel H (1969) The social organization and population ecology of the plains zebra (Equus quagga). Zool Afr 4:249–263
Klingel H (1975) A comparison of the social behaviour of the Equidae. In: Geist V, Walther F (eds) The Papers of an International Symposium on The Behaviour of Ungulates and its relation to management. IUCN, pp 124–132
Klingel H (1977) Observations on social organization and behaviour of African and Asiatic wild assess (Equus africanus and E. hemionus). Z Tierpsychol 44:323–331
Krueger K, Flauger B (2011) Olfactory recognition of individual competitors by means of faeces in horse (Equus caballus). Anim Cogn 14:245–257
Leclaire S, Strandh M, Mardon J, Westerdahl H, Bonadonna F (2017) Odour-based discrimination of similarity at the major histocompatibility complex in birds. Proc Roy Soc B. https://doi.org/10.1098/rspb.2016.2466
Linklater WL, Mayer K, Swaisgood RR (2013) Chemical signals of age, sex and identity in black rhinoceros. Anim Behav 85:671–677
Marinier SL, Alexander AJ, Waring GH (1988) Flehmen behaviour in the domestic horse: Discrimination of conspecific odours. Appl Anim Behav Sci 19:227–237
Marneweck C, Jurgens A, Shrader AM (2017) Dung odours signal sex, age, territorial and oestrous state in white rhinos. Proc Roy Soc B. https://doi.org/10.1098/rspb.2016.2376
Marneweck C, Jurgens A, Shrader AM (2018) The role of middens in white rhino olfactory communication. Anim Behav 140:7–18
Marneweck C, Jürgens A, Shrader AM (2019) The first report of urine overmarking of pro-oestrus female dung by a male white rhino. Afr Zool 54:175–179
Meyer JM, Goodwin TE, Schulte BA (2008) Intrasexual chemical communication and social responses of captive female African elephants, Loxodonta africana. Anim Behav 76:163–174
Moehlman PD (1985) The odd-toed ungulates: order Perrisodactyla. In: Brown RE, Macdonald DW (eds) Social odours in mammals, vol 2. Clarendon Press, Oxford, pp 531–549
Penzhorn BL (1984) Observations on mortality of free-ranging Cape Mountain zebras Equus zebra zebra. S Afr J Wild Res 14:89–90
Pluháček J, Bartoš L, Doležalová M, Bartošová-Víchová J (2007) Sex of the foetus determines the time of weaning of the previous offspring of captive plains zebra (Equus burchelli). Appl Anim Behav Sci 105:192–204
Pluháček J, Tučková V, King SRB (2019a) Overmarking behaviour of zebra males: no scent masking, but a group cohesion function across three species. Behav Ecol Sociobiol 73:137
Pluháček J, Tučková V, Šárová R, King SRB (2019b) Test of four hypotheses to explain the function of overmarking in foals of four equid species. Anim Cogn 22:231–241
Pluháček J, Tučková V, Šárová R, King SRB (2020) Effect of social organisation on interspecific differences in overmarking behaviour of foals in African equids. Anim Cogn 23:131–140
Ralls K (1971) Mammalian scent marking. Science 171:443–449
Steiner CC, Ryder OA (2011) Molecular phylogeny and evolution of the Perissodactyla. Zool J Lin Soc 163:1289–1303
Steiner CC, Mitelberg A, Tursi R, Ryder OA (2012) Molecular phylogeny of extant equids and effects of ancestral polymorphism in resolving species-level phylogenies. Mol Phyl Evol 65:573–581
Tučková V, Šárová R, Bartošová J, King SRB, Pluháček J (2018) Overmarking by adult females in four equid species: social bonds and group cohesion. J Zool 306:180–188
Turner JW, Perkins A, Kirkpatrick JF (1981) Elimination marking behavior in feral horses. Can J Zool 59:1561–1566
Tyler SJ (1972) The behaviour and social organisation of the New Forest ponies. Anim Behav Monographs 5:85–196
Wolff JO, Mech SG, Thomas SA (2002) Scent marking in female prairie voles: a test of alternative hypotheses. Ethol 108:483–494
Wronski T, Apio A, Plath M, Ziege M (2013) Sex difference in the communicatory significance of localized defecation sites in Arabian gazelles (Gazella arabica). J Ethol 31:129–140
Acknowledgements
The authors gratefully acknowledge the assistance of the Zoological Garden staff at Dvůr Králové, Brno, Liberec, Ostrava, and Ústí nad Labem, in particular Luděk Čulík, Markéta Čulíková, Aleš Kopecký, Jiří Soumar, Miroslava Kubelková, Miroslava Doležalová, Pavel Moucha, Radek Hlávka, Lubomír Melichar, Jiří Vítek, Petra Padalíková, Pavel Král, Jaroslav Novák, Rostislav Střižík, Eva Zajoncová, and Lenka Málcová. We highly appreciated the comments of an anonymous reviewer and Kazuhiro Goto that helped to improve the manuscript. This work was supported by the Ministry of Agriculture of the Czech Republic (MZE-RO0718) and by grant no. 2011/008 of the Student Grant Agency of the University of South Bohemia.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Compliance for ethical standards
The study was designed according to laws of the European Union and Czech Republic.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Pluháček, J., Tučková, V., Šárová, R. et al. Why wait to mark? Possible reasons behind latency from olfactory exploration to overmarking in four African equid species. Anim Cogn 25, 1443–1452 (2022). https://doi.org/10.1007/s10071-022-01625-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10071-022-01625-5