
Abstract
Large-scale phylogenetic studies of animal cognition have revealed robust links between absolute brain volume and species differences in executive function. However, past comparative samples have been composed largely of primates, which are characterized by evolutionarily derived neural scaling rules. Therefore, it is currently unknown whether positive associations between brain volume and executive function reflect a broad-scale evolutionary phenomenon, or alternatively, a unique consequence of primate brain evolution. Domestic dogs provide a powerful opportunity for investigating this question due to their close genetic relatedness, but vast intraspecific variation. Using citizen science data on more than 7000 purebred dogs from 74 breeds, and controlling for genetic relatedness between breeds, we identify strong relationships between estimated absolute brain weight and breed differences in cognition. Specifically, larger-brained breeds performed significantly better on measures of short-term memory and self-control. However, the relationships between estimated brain weight and other cognitive measures varied widely, supporting domain-specific accounts of cognitive evolution. Our results suggest that evolutionary increases in brain size are positively associated with taxonomic differences in executive function, even in the absence of primate-like neuroanatomy. These findings also suggest that variation between dog breeds may present a powerful model for investigating correlated changes in neuroanatomy and cognition among closely related taxa.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Background
Comparative studies suggest that variance in brain size is linked to species differences in some aspects of cognition (Benson-Amram et al. 2016; Deaner et al. 2007; Garamszegi and Eens 2004; Kotrschal et al. 2013, 2015; MacLean et al. 2014; Madden 2001; Overington et al. 2009; Reader and Laland 2002; Shultz and Dunbar 2010; Sol et al. 2005, 2008). Studies of relative brain size have revealed interspecific links with problem solving in carnivores (Benson-Amram et al. 2016) and innovativeness in birds (Overington et al. 2009), while intraspecific examinations have linked relative brain size to numerical learning, spatial learning, and reversal learning in guppies (Buechel et al. 2017; Kotrschal et al. 2013, 2015). Phylogenetic studies of absolute brain size have demonstrated a strong relationship between absolute brain volume and components of executive function (a suite of cognitive abilities involved in behavioral control, including working memory and inhibition) (Deaner et al. 2007; MacLean et al. 2014; Shultz and Dunbar 2010). However, much of the work linking absolute brain size to executive function has been conducted with primates, and recent advances in comparative neuroanatomy reveal that primates are characterized by derived neural scaling properties, which contrast with those of other orders (Herculano-Houzel 2012, 2017; Herculano-Houzel et al. 2007). Specifically, in most mammals, as brain volume increases, average neuron size tends to increase, and neurons become less densely populated in the brain (Herculano-Houzel 2014, 2017; Herculano-Houzel et al. 2006). In contrast, as primate brains increase in volume, both neuron size and density remain constant, resulting in isometric (i.e., 1:1) scaling between these variables (Herculano-Houzel et al. 2007). As a result, for a given primate and non-primate mammal of equivalent brain size, the primate’s brain is expected to contain more neurons and to have greater neuron density. Similarly, in primates, a twofold change in brain volume is expected to lead to a twofold increase in the number of neurons in the larger brain, whereas a twofold change in volume leads to the addition of many fewer neurons in non-primate mammals. This phenomenon has been proposed to account for the uniqueness of the human brain [but see (Mortensen et al. 2014) for evidence that long-finned pilot whales have more neocortical neurons that humans, but lower neuron density], the ‘primate advantage’ of increases in brain volume, and previously observed associations between brain volume and species differences in executive function (Deaner et al. 2007; Herculano-Houzel 2012; MacLean et al. 2014). Consequently, it remains unknown whether the relationship between brain volume and executive function reflects a broad-scale evolutionary phenomenon, or alternatively, a unique consequence of primate brain evolution.
Therefore, an important test of the association between brain volume and executive function requires analysis with a large sample of taxa that do not adhere to primate-like neural scaling rules. Domesticated dogs, with their extraordinary degree of intraspecific morphological variation [including variation in brain size (Kruska 1988; Wosinski et al. 1996)], offer a unique opportunity for such a study. New evidence shows that dogs adhere to the same cortical scaling rules as other non-primate mammals, and that domestication appears to have had no effect on allometric brain scaling in dogs (or other domesticated carnivores examined) (Jardim-Messeder et al. 2017). In addition, the previous phylogenetic studies of links between brain size and cognition have been critiqued for ignoring heterogeneity resulting from variation in evolutionary trajectories or different selection pressures on neuroanatomy across taxa (Logan et al. 2018). As the relative sizes of different brain structures are likely to be less variable within, than across species (Finlay and Darlington 1995; Gould et al. 2013; Healy and Krebs 1992; Lucas et al. 2004) dogs allow us to assess the cognitive correlates of changes in absolute brain size while better controlling for changes in the relative sizes of specific brain regions, or other factors that vary considerably across diverse taxonomic groups (Barton and Harvey 2000; Barton and Venditti 2014; Gould et al. 2013). Although breed-level variation in neuroanatomy remains largely unexplored, preliminary data suggest that the relative sizes of neural structures do not vary significantly across dog breeds with large degrees of variation in brain volume and skull shape (Roberts et al. 2010; Thames et al. 2009).
Here, we report the results of analyses investigating the association between estimated brain weight and breed differences in dog cognition, measured in a sample of more than 7000 individuals from 74 breeds, on a battery of cognitive tests designed to probe diverse aspects of cognition [broadly defined as the processes through which animals acquire, store, and act on information from the environment (Shettleworth 2009)].
Methods
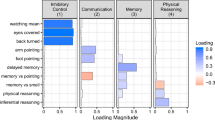
Cognitive data were collected through Dognition.com, a citizen science (Bonney et al. 2009; Cooper 2016) website that provides dog owners with instructions for completing cognitive experiments with pet dogs in their homes. Analyses of these data replicate findings from similar tests conducted in laboratory settings (Stewart et al. 2015), and psychometric analyses of citizen science data yield underlying factors consistent with those obtained using conventional approaches (MacLean et al. 2017). We included data from all ten of Dognition’s primary cognitive tasks in our analyses, measuring a wide range of cognitive skills including components of executive function, inferential and causal reasoning, and communicative processes (Table 1, Online Resource 1). We analyzed data from 7397 purebred adult dogs representing 74 breeds (Online Resource 1). Breed-average body weights were compiled from the Canine Behavioral Assessment and Research Questionnaire (C-BARQ) (Hsu and Serpell 2003; McGreevy et al. 2013), and breed-average brain weights were estimated from a scaling function described by Bronson (1979) and validated using C-BARQ body weights for a sample of 24 breeds with known brain weights (Online Resource 1). To control for genetic relatedness between breeds, the associations between estimated brain weight and cognitive measures were tested using Efficient Mixed Modeling for Association studies (EMMA) (Kang et al. 2008; Zhou and Stephens 2012). Genetic relatedness among breeds was incorporated using a breed-average identity-by-state (IBS) matrix (Boyko et al. 2010), using molecular data from Hayward and colleagues (2016). For analyses including individual-level data across breeds, breed-level IBS matrices were multiplied by an individual-level incidence matrix to generate an individual-level IBS matrix. To avoid the assumption that members of the same breed were clonal, pairwise within-breed IBS values were set to the average IBS value between members of that breed. Tests were conducted using the ‘EMMREML’ package (Akdemir and Godfrey 2015) in the R environment (v.3.3.1) (R Core Team 2016). Associations were considered significant at an alpha level of 0.05. Data are available as electronic supplementary material (Online Resource 2).

Scores on a measure of short-term memory (delayed memory) as a function of estimated absolute brain weight across dog breeds. The dashed lines show the regression slopes from linear mixed models controlling for genetic relatedness between breeds a across all trials, b across short delays of 60 and 90 s, and c across long delays of 120 and 150 s. Each breed included in the analyses had at least 20 individuals complete this task, and is represented by one diamond
Results
Absolute brain size predicts breed differences in executive function
Based on the previous studies linking absolute brain size to executive function, we hypothesized that larger-brained breeds would score higher on cognitive measures related to executive function. To test this hypothesis, we fit mixed models predicting breed-average scores on two tasks indexing components of working memory (short-term memory) and of inhibitory control (self-control) as a function of estimated absolute brain weight. These models revealed significant associations between estimated brain weight and performance on both tasks (Table 1; Figs. 1, 2).

Difference scores between the watching and not watching conditions in the cunning task (a) and scores on measures of self-control (b cunning: watching condition; c cunning: not watching condition) as a function of estimated brain weight across dog breeds. The dashed lines show the regression slopes from linear mixed models controlling for genetic relatedness between breeds. Each breed included in the analyses had at least 20 individuals complete this task, and is represented by one diamond
In the delayed memory task, larger-brained breeds correctly remembered the location of hidden food at significantly higher levels than smaller-brained breeds when examining performance after all delay lengths combined [β = 0.0021, χ2(1) = 14.18, p < 0.001; Fig. 1a], as well as after short (60 s and 90 s) and long (120 s and 150 s) delays [short delays: β = 0.0017, χ2(1) = 7.91, p = 0.005, Fig. 1b; long delays: β = 0.0023, χ2(1) = 9.70, p = 0.001, Fig. 1c]. The slope of the regression relating estimated brain weight to performance was greater at long delays than short delays, suggesting that the gap in performance between breeds of varying brain sizes generally widens as a function of increased cognitive demands. The effect was weakest at 60 s (β = 0.0014, χ2(1) = 2.84, p = 0.09), but became significant as delay length increased [90 s: β = 0.0020, χ2(1) = 6.99, p = 0.01; 120 s: β = 0.0027, χ2(1) = 5.78, p = 0.02; 150 s: β = 0.0020, χ2(1) = 5.18, p = 0.02].
In the cunning task, dogs were prohibited from eating a visible food reward, and whether the experimenter watched the dog varied across conditions. Although designed as a measure of sensitivity to human perception, all conditions in this task pose demands on self-control, as dogs were required to inhibit a desire to consume the visible food [see Müller et al. (2016) for a similar measure assessing self-control in dogs]. In this task, larger-brained breeds were significantly slower to pilfer prohibited food placed directly in front of them than smaller-brained breeds in a condition where a human’s eyes were closed or back was turned (not watching), compared to a condition where a human actively watched the subject [watching; β = − 0.10, χ2(1) = 13.15, p < 0.001; Fig. 2]. Although greater self-control is likely required to inhibit the desire to steal food when not being actively monitored, variation in perspective-taking ability may also contribute to this effect, or smaller-brained breeds may simply be more likely to restrain their actions while being monitored. Thus, we conducted separate analyses of the watching and not watching conditions for this task. In both conditions, larger-brained breeds exhibited greater self-control by waiting significantly longer to eat the forbidden food [(cunning) watching: β = 0.33, χ2(1) = 9.98, p = 0.002; (cunning) not watching: β = 0.43, χ2(1) = 17.94, p < 0.001]. In addition, the slope of the regression relating estimated brain weight to time to pilfer food was greater in the not watching condition than in the watching condition, supporting the idea that greater self-control is required to resist temptation when not being actively monitored. Therefore, with increased task difficulty in both the cunning and delayed memory tasks, the slopes of the relationships between cognitive performance and brain size generally become more pronounced.
Associations between brain size and cognition vary across cognitive domains
A fundamental question in the cognitive sciences is whether skills for solving diverse problems are subserved by a common set of cognitive processes that are flexibly applied across contexts (domain generality), or alternatively, by specialized processes that are differentially applied in specific contexts (domain specificity). Due to the practical challenges of conducting cognitive experiments with large samples of species, most previous studies investigating links between brain volume and cognition used a small number of cognitive tasks, precluding assessment of the specificity of associations between brain size and measures of cognition. To test the predictions of the domain-general and domain-specific hypotheses, we investigated whether estimated brain weight broadly predicts breed differences across diverse cognitive measures, or alternatively, if these associations are limited to executive function.
Consistent with domain-specific hypotheses (Amici et al. 2012; MacLean et al. 2017), the relationship between estimated brain weight and breed differences in cognition was highly variable across the ten cognitive measures (Table 1; Fig. 3). For six of the ten tasks, there was no association between estimated brain weight and breed-average performance [(contagious) yawning: β = − 0.00053, χ2(1) = 1.19, p = 0.27; eye contact: β = − 0.018, χ2(1) = 0.12, p = 0.73; foot pointing: β = − 0.00013, χ2(1) = 0.15, p = 0.70; memory vs. smell: β = − 0.00023, χ2(1) = 0.15, p = 0.69; inferential reasoning: β = 0.00011, χ2(1) = 0.03, p = 0.87; physical reasoning (visual causality): β = 0.00042, χ2(1) = 0.30, p = 0.59]. In addition to significant associations between estimated brain weight and components of executive function, larger-brained breeds were also significantly more likely to search for food in a location indicated by an arm-pointing gesture [arm pointing: β = 0.00068, χ2(1) = 4.18, p = 0.041], and were significantly more likely to rely on this pointing gesture when it was pitted against their own memory of where food had been hidden [memory vs. pointing: β = − 0.0013, χ2(1) = 3.99, p = 0.046].

Scores on all cognitive tasks except delayed memory and cunning (see Figs. 1, 2) as a function of estimated brain weight across dog breeds. The dashed lines show the regression slopes from linear mixed models controlling for genetic relatedness between breeds. Each breed included in the analyses had at least 20 individuals complete a given task, and is represented by one diamond
To explore whether behavioral data were stable across trials within tasks significantly associated with estimated brain weight, and whether any variation across time differed as a function of estimated brain weight, we fit mixed models predicting task performance from trial number alone, as well as from a trial number by estimated brain weight interaction term. To account for repeated measures, all models included a random intercept for breed. Trial number alone was a significant predictor of task performance in cunning due to dogs pilfering forbidden food significantly faster over the course of the trials in the watching condition but not in the not watching condition, as well as in memory vs. pointing, such that dogs became less likely to follow the pointing gesture over time (Online Resource 1). However, in no cases were there any significant trial number by estimated brain weight interactions (Online Resource 1). The lack of significant interactions between trial number and brain weight demonstrates that, while performance varied across trials in some measures, it did not vary differentially across breeds as a function of brain weight. Therefore, the significant associations identified between estimated brain weight and cognitive performance in our main analyses are unlikely to be due to differential learning between small and large-brained breeds over the course of each task.
Individual-level analyses
To examine how within-breed variation in cognition may influence our results, we fit mixed models predicting scores on each task as a function of breed-average estimated brain weight on an individual level (Online Resource 1). Results from these individual-level analyses largely mirrored those from the main breed-level analyses: Estimated brain weight was again a significant predictor of performance in each measure which primarily indexed components of executive function [delayed memory: β = 0.0015, χ2(1) = 11.06, p < 0.001; (cunning) watching: β = 0.27, χ2(1) = 10.25, p = 0.001; (cunning) not watching: β = 0.33, χ2(1) = 15.34, p < 0.001], as well as difference scores between conditions in the cunning task [β = − 0.080, χ2(1) = 13.40, p < 0.001] and performance in arm pointing [β = 0.00068, χ2(1) = 6.51, p = 0.01]. However, unlike results from the breed-level analyses, the association between estimated brain weight and performance in memory vs. pointing fell slightly above the significance threshold [β = − 0.0011, χ2(1) = 3.64, p = 0.056], while a significant association emerged between estimated brain weight and performance in physical reasoning [β = 0.00092, χ2(1) = 4.00, p = 0.045].
Body size
As breed-average brain weights were estimated from breed-average body weights in our main analyses, we expected to find many similar associations between cognition and body size. Mixed models predicting task scores as a function of C-BARQ-reported breed-average body weight generally revealed the same pattern of results as those using breed-average estimated brain weight as the predictor; body weight was a significant predictor of performance in delayed memory, cunning, and the watching and not watching conditions analyzed separately, but associations between body weight and arm pointing (p = 0.07) and memory vs. pointing (p = 0.06) fell slightly above the significance threshold (Online Resource 1). In the majority of cases, linear models using estimated brain weight as the predictor had a lower AIC and explained a larger proportion of the variance in cognitive measures as compared to models using body weight as the predictor, but differences in AIC and adjusted R2 were generally small (Online Resource 1). Even in studies using more direct measures of brain size rather than estimation, the challenge of disentangling associations with absolute brain size and body size is evident as the two variables are very highly correlated (MacLean et al. 2014).
To explore whether the same associations between cognition and brain size hold when estimating breed-average brain weight independently from body weight, we compiled breed-average skull measurements from Boyko et al. (2010) for a subset of breeds (n = 27) for whom data were available. To assess which skull measures best predicted brain weight, we first fit a multiple regression model predicting the known breed-average brain weights from Bronson (1979) using breeds for which both skull measurements and brain weights were available (n = 12) as a function of cranial depth, maximum cranial width, and least cranial width. As results from this model showed that only cranial depth was a significant predictor of brain weight controlling for the other predictors, we next fit a simple linear regression model predicting brain weight from cranial depth alone. Breed-average cranial depth explained a greater proportion of the variance in brain weight and had a lower model AIC [β = 0.01, F(1, 10) = 119.4, p < 0.001, adjusted R2 = 0.92, AIC = 82.13] than did C-BARQ-reported breed-average body weight in this same subset [β = 1.03, F(1, 10) = 62.83, p < 0.001, adjusted R2 = 0.85, AIC = 89.03], suggesting that cranial depth was a better predictor of brain weight than was body weight in this sample. We then used cranial depth to predict breed-average brain weight across the 27 breeds for which cognitive data and skull measurements were available. Finally, we fit mixed models (controlling for genetic relatedness between breeds) predicting scores on each cognitive task as a function of these new brain weight estimates. Breed-average brain weight estimated from cranial depth significantly predicted performance in each measure which primarily indexed components of executive function [delayed memory: β = 0.0014, χ2(1) = 4.91, p = 0.03; (cunning) watching: β = 0.43, χ2(1) = 4.51, p = 0.03; (cunning) not watching: β = 0.49, χ2(1) = 6.72, p = 0.01], but did not significantly predict performance in arm pointing [β = − 0.0002, χ2(1) = 0.09, p = 0.76], memory vs. pointing [β = 0.0002, χ2(1) = 0.02, p = 0.89], or difference scores between conditions in the cunning task [β = − 0.06, χ2(1) = 0.69, p = 0.41]. Therefore, estimates of breed-average brain weight derived independently of breed-average body weight were significantly associated with the measures of short-term memory and self-control, but not other cognitive measures.
The findings above reveal a range of associations between estimated brain weight and breed differences in cognition. However, it remains possible that additional factors, which covary with body weight and brain weight, may partially account for these associations. To address this possibility, we conducted additional analyses with potentially confounding variables.
Perceptual factors
Previous studies suggest that skull shape is associated with visual perception in dogs. Specifically, brachycephalic dogs [defined by a high cephalic index (CI), see Online Resource 1] are characterized by more forward-facing eyes and greater ocular overlap (Gácsi et al. 2009; Helton and Helton 2010). Gácsi et al. (2009) found that brachycephalic breeds were significantly better at following human pointing cues as compared to dolichocephalic breeds (low CI), and suggested that this result may be partially attributable to breed differences in visual perception. Our sample contained 50 breeds for which CI measurements were reported in (McGreevy et al. 2013) or (Boyko et al. 2010), and in these breeds, CI was significantly negatively correlated with estimated brain weight (r = − 0.43, p = 0.002). Thus, explanations invoking breed differences in vision yield predictions opposite to what we observed with measures of short-term memory, self-control, and gesture following. Indeed, in breeds for which both cognitive and CI data were available, CI was significantly negatively correlated with performance in measures of short-term memory and self-control [delayed memory: r = − 0.47, p = 0.02; (cunning) watching: r = − 0.41, p = 0.02; (cunning) not watching: r = − 0.52, p = 0.002], and was not significantly correlated with the measures of gesture following (arm pointing: r = − 0.19, p = 0.24; memory vs. pointing: r = − 0.04, p = 0.85). In addition, because we do not find consistent relationships between estimated brain weight and cognitive performance across tasks incorporating highly similar experimental set-ups and identical stimulus presentation distances, it is highly unlikely that breed differences in vision play a large role in the relationships we observed.
Training history
A second important consideration relates to possible effects of training history on these cognitive measures. Using questionnaire data submitted to Dognition.com, we found that breeds with larger brains were more likely to have attended at least one obedience class (r = 0.71, p < 0.001), and were more likely to have been extensively trained (r = 0.75, p = 0.01). Although this information was not available for the majority of our subjects (precluding inclusion as a covariate in the main breed-level analyses), we fit mixed models predicting scores on each task as a function of breed-average-estimated brain weight and owner-reported extent of training history on an individual level including dogs for which both measures were available (Table 2). These analyses revealed that, controlling for training history, estimated brain weight was a significant predictor of performance in each measure which primarily indexed components of executive function [delayed memory: β = 0.0015, χ2(1) = 4.22, p = 0.04; (cunning) watching: β = 0.35, χ2(1) = 12.95, p < 0.001; (cunning) not watching: β = 0.38, χ2(1) = 17.93, p < 0.001], but was not a significant predictor of performance in any other measures [yawning: β = − 0.00043, χ2(1) = 0.21, p = 0.64; eye contact: β = − 0.068, χ2(1) = 1.17, p = 0.28; arm pointing: β = 0.00056, χ2(1) = 1.78, p = 0.18; foot pointing: β = − 0.00020, χ2(1) = 0.17, p = 0.68; cunning: β = − 0.028, χ2(1) = 0.25, p = 0.62; memory vs. pointing: β = − 0.00095, χ2(1) = 0.91, p = 0.34; memory vs. smell: β = − 0.00021, χ2(1) = 0.078, p = 0.78; inferential reasoning: β = 0.00082, χ2(1) = 1.17, p = 0.28; physical reasoning: β = 0.0011, χ2(1) = 2.42, p = 0.12]. Therefore, as in the breed-level analyses, estimated brain weight was a significant predictor of performance on measures of short-term memory and self-control, even after controlling for training history on an individual level. However, unlike results from the main breed-level analyses (but mirroring the pattern of results obtained using cranial depth rather than body weight to estimate brain weight in a subset of our sample), estimated brain weight was not a significant predictor of difference scores between conditions in the cunning task or of reliance on gesture following in arm pointing or memory vs. pointing after controlling for training history. We also found that, controlling for estimated brain weight, training history was a significant predictor of performance in the watching [β = 8.67, χ2(1) = 31.08, p < 0.001] and not watching [β = 8.55, χ2(1) = 30.02, p < 0.001] conditions of the cunning task, as well as in eye contact [β = 3.60, χ2(1) = 15.87, p < 0.001], arm pointing [β = 0.024, χ2(1) = 8.18, p = 0.004], memory vs. pointing [β = − 0.041, χ2(1) = 5.94, p = 0.015], and inferential reasoning [β = 0.029, χ2(1) = 3.96, p = 0.047], but was not a significant predictor of performance in the six remaining measures [yawning: β = − 0.018, χ2(1) = 0.95, p = 0.33; foot pointing: β = 0.0097, χ2(1) = 1.11, p = 0.29; cunning: β = 0.35, χ2(1) = 0.16, p = 0.69; memory vs. smell: β = − 0.027, χ2(1) = 3.55, p = 0.059; delayed memory: β = 0.026, χ2(1) = 3.46, p = 0.063; physical reasoning: β = 0.18, χ2(1) = 1.71, p = 0.19]. It is important to note that, in measures of self-control (watching and not watching conditions of cunning), both estimated brain weight and training history made independent significant contributions to task performance, the latter of which is unsurprising given that owners verbally forbid subjects from taking visible food (a commonly trained command) as a part of the task.
Functional breed group classification
A third consideration is that modern dog breeds have been selected for a variety of functional roles, with some roles (e.g., hunting and herding) requiring extensive cooperation with humans (Coppinger and Schneider 1995). Previous studies suggest that dogs bred for cooperative roles may be more likely to follow human gestures (Udell et al. 2014; Wobber et al. 2009). Thus, to control for potential effects of breed group, we fit models including American Kennel Club (AKC) breed group designation and estimated brain weight as predictors of performance. For the memory vs. pointing task, but none of the other measures significantly associated with estimated brain weight in the main analyses, breed group was a significant predictor of performance controlling for estimated brain weight [χ2(6) = 1.35, p = 0.03] but estimated brain weight was not a significant predictor of performance controlling for breed group [χ2(1) = 0.04, p = 0.15]. AKC-classified “sporting” and “working” breeds were the most likely to follow an arm-pointing gesture in memory vs. pointing (proportion of choices based on memory—sporting: mean = 0.60, SEM = 0.02; working: mean = 0.63, SEM = 0.02), while breeds in the “terrier” and “herding” groups were the least likely (proportion of choices based on memory—terrier: mean = 0.70, SEM = 0.01; herding: mean = 0.69, SEM = 0.01). While increased tendencies to follow an arm-pointing gesture over memory in the memory vs. pointing task may be better explained by differences in AKC breed group than by differences in brain size, we find only mixed support for the idea that selection for cooperative roles is primarily responsible for this result, as breeds belonging to the herding group were among the least likely to follow gestures in this task.
Discussion
Our findings support the hypothesis that increases in absolute brain size are associated with variation in executive function, even in the absence of primate-like neuroanatomy. This result raises new questions about the cognitive consequences of evolutionary changes in brain size, and suggests that, even within a species, brain size is associated with some aspects of cognition. By investigating intraspecific variance in a species characterized by high levels of morphological diversity, this study circumvents some challenges inherent to the previous interspecific comparisons. Specifically, across species, differences in absolute brain volume can be confounded with changes in the relative proportions of specific brain regions. This phenomenon can arise both due to specialization of specific brain structures [mosaic brain evolution (Barton and Harvey 2000; Barton and Venditti 2014; Gould et al. 2013)] resulting from heterogeneity in selection pressures on neuroanatomical variation across taxa (Logan et al. 2018), or due to conserved developmental processes involving the timing of neurogenesis, which yield disproportionate enlargement of late developing structures (e.g., the neocortex) in larger-brained taxa (Finlay and Darlington 1995; Kaas 2000). Regarding the latter possibility, there are currently few data on the development of brain structures in dogs, but preliminary data suggest that breed differences in brain volume are less subject to these conserved developmental constraints which induce allometry (deviation from 1:1 scaling) between total brain volume and specific regions of the brain. Specifically, in contrast to comparative mammalian samples, imaging studies have shown that there are no significant differences in the proportional sizes of the cerebellum, forebrain, or brainstem across dog breeds with widely varying total brain volumes (Thames et al. 2009), or in absolute size of the olfactory lobe across breeds with widely varying skull shapes (Roberts et al. 2010). This finding is consistent with the notion that allometric relationships which apply across species often do not hold within species (Armstrong 1990), leading researchers to suggest that the extraordinary morphological variation of dogs makes this species an ideal candidate for investigating intraspecific brain scaling and the cognitive implications thereof (Jardim-Messeder et al. 2017).
Importantly, even with a lack of isometric (i.e. 1:1) scaling between brain volume and the number of neurons in the brain, larger dog brains are still expected to contain more neurons than smaller dog brains. In the only comparison of dog neuron counts to date, Jardim-Messeder et al. found that the cortex of a golden retriever with a body weight of 32 kg contained 627 million neurons, while the cortex of an unknown breed with a body weight of 7.45 kg contained 429 million neurons (Jardim-Messeder et al. 2017). Thus, if the number of neurons in the brain is expected to predict taxonomic differences in executive function (Herculano-Houzel 2017), our findings are consistent with this hypothesis. However, because increases in volume are expected to result in the addition of many fewer neurons in dog brains than primate brains, we would theoretically expect differences in the slope of the relationship between brain volume and measures of executive function between these taxonomic groups given adequate control of extraneous variables.
A second major finding from this study is that the relationship between estimated brain weight and breed differences in cognition varied widely across cognitive tasks. For example, we found only limited evidence for an association between estimated brain weight and reasoning about causal properties of the world, and no evidence for an association between estimated brain weight and inferential reasoning, use of a novel communicative gesture (foot pointing), or measures of eye contact with a human. After controlling for training history at an individual level, we also found no evidence for an association between estimated brain weight and reliance on following communicative pointing cues to locate food. This result supports domain-specific accounts of cognition, and is consistent with previous comparative studies which have revealed links between brain volume and executive function, but no association between brain volume and other domains of cognition (MacLean et al. 2013). As suggested by domain specificity, absolute brain size is, therefore, unlikely to be an informative predictor of all cognitive abilities. However, absolute brain size may be particularly important for executive function, because these processes exert high-level ‘supervisory’ control over a variety of more specialized cognitive functions. In addition, these results suggest that the significant associations between estimated brain weight and measures of executive function are unlikely to be artifacts of breed differences in motivation or perception, in which case we would have expected similar associations for all the tasks involving a search for food, or requiring attention to visual stimuli in the frontal field.
One limitation of this study stems from the lack of individual-level brain weight data from the dogs in our sample. While the scaling function used to estimate breed-average brain weights described by Bronson (1979) explained an overwhelming percentage of the variance in brain weight (93%) in Bronson’s sample (26 breeds ranging from 3.6 kg to 55.0 kg in body weight), and a comparable percentage (91%) when using C-BARQ breed-average body weight as a predictor, we cannot rule out the possibility that brain-to-body size scaling in breeds absent from Bronson’s sample deviate from the scaling relationship used in our estimates. However, while estimates of brain weight based on body weight are subject to prediction error, the effects of this error are minimized in comparisons of taxa characterized by large differences in body size. Given the over 17-fold variation in body weight across breeds in our sample (range 3.35–59.75 kg), and confirmation of a strong link between body and brain weight in dogs (Bronson 1979), it is highly unlikely that prediction error substantially influenced our results. As expected given the strong association between body and brain weight, the observed associations between cognition and estimated absolute brain size were generally the same as those that exist between cognition and body weight in our sample. However, we found the same links between absolute brain size and executive function in a subset of our sample using brain weight estimated from skull measurements, rather than body weight. Relatedly, the exact intraspecific neural scaling rules of dog brains remain unknown, but it is of note that the two individuals examined in Jardim-Messeder et al. (2017) fit the interspecific neural scaling rules that apply across non-primate mammals. Future work can address these limitations by examining how differences in cognition relate to variation in brain size and neuron count on an individual level both within and across dog breeds. Similarly, without independent measures of brain and body weight, we were unable to explore possible associations between relative brain size and performance on the cognitive tasks.
In conclusion, our findings suggest that evolutionary relationships between executive function and absolute brain size do not require primate-like neural scaling rules, that these relationships may be independent of changes in the relative sizes of specific areas of the brain, and that associations between brain size and cognition vary across cognitive domains. These findings raise new questions about how evolutionary shifts in brain size influence cognition across taxa, and suggest that dog breeds provide a powerful and highly tractable model for assessing the cognitive consequences of neuroanatomical variation among close genetic relatives.
References
Akdemir D, Godfrey OU (2015) EMMREML: fitting mixed models with known covariance structures. https://cran.r-project.org/web/packages/EMMREML/EMMREML.pdf
Amici F, Barney B, Johnson VE, Call J, Aureli F (2012) A modular mind? A test using individual data from seven primate species. PLoS One. https://doi.org/10.1371/journal.pone.0051918
Armstrong E (1990) Brains, bodies and metabolism. Brain Behav Evol 36:166–176. https://doi.org/10.1159/000115305
Barton RA, Harvey PH (2000) Mosaic evolution of brain structures in mammals. Nature 405(1997):1055–1058
Barton RA, Venditti C (2014) Rapid evolution of the cerebellum in humans and other great apes. Curr Biol 24(20):2440–2444. https://doi.org/10.1016/j.cub.2014.08.056
Benson-Amram S, Dantzer B, Stricker G, Swanson EM, Holekamp KE (2016) Brain size predicts problem-solving ability in mammalian carnivores. Proc Natl Acad Sci 113(9):2532–2537. https://doi.org/10.1073/pnas.1505913113
Bonney R, Cooper CB, Dickinson J, Kelling S, Phillips T, Rosenberg KV, Shirk J (2009) Citizen science: a developing tool for expanding science knowledge and scientific literacy. Bioscience 59(11):977–984. https://doi.org/10.1525/bio.2009.59.11.9
Boyko AR, Quignon P, Li L, Schoenebeck JJ, Degenhardt JD, Lohmueller KE, Ostrander EA (2010) A simple genetic architecture underlies morphological variation in dogs. PLoS Biol 8(8):49–50. https://doi.org/10.1371/journal.pbio.1000451
Bronson RT (1979) Brain weight-body weight scaling in breeds of dogs and cats. Brain Behav Evol 16:227–236
Buechel SD, Boussard A, Kotrschal A, van der Bijl W, Kolm N (2017) Brain size affects performance in a reversal-learning test. Proc R Soc Lond Ser B Biol Sci. https://doi.org/10.1098/rspb.2017.2031
Cooper C (2016) Citizen science: How ordinary people are changing the face of discovery. The Overlook Press, New York
Coppinger R, Schneider R (1995) Evolution of working dogs. In: Serpell J (ed) The domestic dog: its evolution, behaviour and interactions with people. Cambridge University Press, pp 21–47. https://www.gwern.net/docs/genetics/heritable/1995-serpellthedomesticdog.pdf
Deaner RO, Isler K, Burkart J, Van Schaik C (2007) Overall brain size, and not encephalization quotient, best predicts cognitive ability across non-human primates. Brain Behav Evol 70(2):115–124. https://doi.org/10.1159/000102973
Finlay B, Darlington R (1995) Linked regularities in the development and evolution of mammalian brains. Science 268(5217):1578–1584. https://doi.org/10.1126/science.7777856
Gácsi M, McGreevy PD, Kara E, Miklósi Á (2009) Effects of selection for cooperation and attention in dogs. Behav Brain Funct BBF 5:31. https://doi.org/10.1186/1744-9081-5-31
Garamszegi LZ, Eens M (2004) The evolution of hippocampus volume and brain size in relation to food hoarding in birds. Ecol Lett 7(12):1216–1224. https://doi.org/10.1111/j.1461-0248.2004.00685.x
Gould KL, Gilbertson KE, Seyfer AL, Brantner RM, Hrvol AJ, Kamil AC, Nelson JC (2013) Differences in relative hippocampus volume and number of hippocampus neurons among five corvid species. Brain Behav Evol 81(1):56–70. https://doi.org/10.1146/annurev-immunol-032713-120240.Microglia
Hayward JJ, Castelhano MG, Oliveira KC, Corey E, Balkman C, Baxter TL, Boyko AR (2016) Complex disease and phenotype mapping in the domestic dog. Nat Commun 7(10460):1–11. https://doi.org/10.1038/ncomms10460
Healy SD, Krebs JR (1992) Foord storing and the hippocampus in corvids: amount and volume are correlated. Proc R Soc Lond 248:241–245. https://doi.org/10.1098/rspb.1992.0068
Helton WS, Helton ND (2010) Physical size matters in the domestic dog’s (Canis lupus familiaris) ability to use human pointing cues. Behav Process 85(1):77–79. https://doi.org/10.1016/j.beproc.2010.05.008
Herculano-Houzel S (2012) Neuronal scaling rules for primate brains: the primate advantage. In: Hofman MA, Falk D (eds) Progress in brain research, vol 195, 1st edn. Elsevier B.V., pp. 325–340. https://doi.org/10.1016/B978-0-444-53860-4.00015-5
Herculano-Houzel S (2014) The glia/neuron ratio: how it varies uniformly across brain structures and species and what that means for brain physiology and evolution. Glia 62(9):1377–1391. https://doi.org/10.1002/glia.22683
Herculano-Houzel S (2017) Numbers of neurons as biological correlates of cognitive capability. Curr Opin Behav Sci 16:1–7. https://doi.org/10.1016/j.cobeha.2017.02.004
Herculano-Houzel S, Mota B, Lent R (2006) Cellular scaling rules for rodent brains. Proc Natl Acad Sci 103(32):12138–12143. https://doi.org/10.1016/B0-12-370878-8/00345-1
Herculano-Houzel S, Collins CE, Wong P, Kaas JH (2007) Cellular scaling rules for primate brains. Proc Natl Acad Sci USA 104(9):3562–3567. https://doi.org/10.1073/pnas.0611396104
Hsu Y, Serpell JA (2003) Development and validation of a questionnaire for measuring behavior and temperament traits in pet dogs. J Am Vet Med Assoc 223(9):1293–1300
Jardim-Messeder D, Lambert K, Noctor S, Pestana FM, de Castro Leal ME, Bertelsen MF, Herculano-Houzel S (2017) Dogs have the most neurons, though not the largest brain: trade-off between body mass and number of neurons in the cerebral cortex of large carnivoran species. Front Neuroanat 11:1–18. https://doi.org/10.3389/fnana.2017.00118
Kaas JH (2000) Why is brain size so important: design problems and solutions as neocortex gets bigger or smaller. Brain Mind 1(1):7–23. https://doi.org/10.1023/A:1010028405318
Kang HM, Zaitlen NA, Wade CM, Kirby A, Heckerman D, Daly MJ, Eskin E (2008) Efficient control of population structure in model organism association mapping. Genetics 178:1709–1723. https://doi.org/10.1534/genetics.107.080101
Kotrschal A, Rogell B, Bundsen A, Svensson B, Zajitschek S, Brannstrom I, Kolm N (2013) Artificial selection on relative brain size in the guppy reveals costs and benefits of evolving a larger brain. Curr Biol 23:168–171
Kotrschal A, Corral-Lopez A, Amcoff M, Kolm N (2015) A larger brain confers a benefit in a spatial mate search learning task in male guppies. Behav Ecol 26(2):527–532. https://doi.org/10.1093/beheco/aru227
Kruska D (1988) Effects of domestication on brain structure and behavior in mammals. Hum Evol 3(6):473–485. https://doi.org/10.1007/BF02436333
Logan CJ, Avin S, Boogert N, Buskell A, Cross FR, Currie A, Montgomery SH (2018) Beyond brain size: uncovering the neural correlates of behavioral and cognitive specialization. Comp Cogn Behav Rev 13:55–89. https://doi.org/10.3819/CCBR.2018.130008
Lucas JR, Brodin A, de Kort SR, Clayton NS (2004) Does hippocampal size correlate with the degree of caching specialization? Proc R Soc B Biol Sci 271(1556):2423–2429. https://doi.org/10.1098/rspb.2004.2912
MacLean EL, Sandel AA, Bray J, Oldenkamp RE, Reddy RB, Hare B (2013) Group size predicts social but not nonsocial cognition in lemurs. PLoS One 8(6):1–9. https://doi.org/10.1371/journal.pone.0066359
MacLean EL, Hare B, Nunn CL, Addessi E, Amici F, Anderson RC, Zhao Y (2014) The evolution of self-control. Proc Natl Acad Sci 111(20):E2140–E2148. https://doi.org/10.1073/pnas.1323533111
MacLean EL, Herrmann E, Suchindran S, Hare B (2017) Individual differences in cooperative communicative skills are more similar between dogs and humans than chimpanzees. Anim Behav 126:41–51. https://doi.org/10.1016/j.anbehav.2017.01.005
Madden J (2001) Sex, bowers and brains. Proc Biol Sci 268(1469):833–838. https://doi.org/10.1098/rspb.2000.1425
McGreevy PD, Georgevsky D, Carrasco J, Valenzuela M, Duffy DL, Serpell JA (2013) Dog behavior co-varies with height, bodyweight and skull shape. PLoS One. https://doi.org/10.1371/journal.pone.0080529
Mortensen HS, Pakkenberg B, Dam M, Dietz R, Sonne C, Mikkelsen B, Eriksen N (2014) Quantitative relationships in delphinid neocortex. Front Neuroanat 8:1–10. https://doi.org/10.3389/fnana.2014.00132
Müller CA, Riemer S, Virányi Z, Huber L, Range F (2016) Inhibitory control, but not prolonged object-related experience appears to affect physical problem-solving performance of pet dogs. PLoS One 11(2):1–19. https://doi.org/10.1371/journal.pone.0147753
Overington SE, Morand-Ferron J, Boogert NJ, Lefebvre L (2009) Technical innovations drive the relationship between innovativeness and residual brain size in birds. Anim Behav 78(4):1001–1010. https://doi.org/10.1016/j.anbehav.2009.06.033
R Core Team (2016) A Language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://doi.org/citeulike-article-id:2400517. Accessed 21 June 2016
Reader SM, Laland KN (2002) Social intelligence, innovation, and enhanced brain size in primates. Proc Natl Acad Sci 99(7):4436–4441. https://doi.org/10.1073/pnas.062041299
Roberts T, McGreevy P, Valenzuela M (2010) Human induced rotation and reorganization of the brain of domestic dogs. PLoS One. https://doi.org/10.1371/journal.pone.0011946
Shettleworth SJ (2009) Cognition, evolution, and behavior, 2nd edn. Oxford University Press, Oxford
Shultz S, Dunbar RIM (2010) Species differences in executive function correlate with hippocampus volume and neocortex ratio across nonhuman primates. J Comp Psychol 124(3):252–260. https://doi.org/10.1037/a0018894
Sol D, Duncan RP, Blackburn TM, Cassey P, Lefebvre L (2005) Big brains, enhanced cognition, and response of birds to novel environments. Proc Natl Acad Sci USA 102(15):5460–5465. https://doi.org/10.1073/pnas.0408145102
Sol D, Bacher S, Reader SM, Lefebvre L (2008) Brain size predicts the success of mammal species introduced into novel environments. Am Nat 172(Suppl):S63–S71. https://doi.org/10.1086/588304
Stewart L, MacLean EL, Ivy D, Woods V, Cohen E, Rodriguez K, Hare B (2015) Citizen science as a new tool in dog cognition research. PLoS One 10(9):1–16. https://doi.org/10.1371/journal.pone.0135176
Thames RA, Robertson ID, Flegel T, Henke D, O’Brien DP, Coates JR, Olby NJ (2009) Development of a morphometric magnetic resonance paramater suitable for distinguishing between normal dogs and dogs with cerebellar atrophy. Canine Brain Morphometry 51(3):246–253. https://doi.org/10.1111/j.1740-8261.2009.01655.x
Udell MAR, Ewald M, Dorey NR, Wynne CDL (2014) Exploring breed differences in dogs (Canis familiaris): does exaggeration or inhibition of predatory response predict performance on human-guided tasks? Anim Behav 89:99–105. https://doi.org/10.1016/j.anbehav.2013.12.012
Wobber V, Hare B, Koler-Matznick J, Wrangham R, Tomasello M (2009) Breed differences in domestic dogs’ (Canis familiaris) comprehension of human communicative signals. Interact Stud. https://doi.org/10.1075/is.10.2.06wob
Wosinski M, Schleicher A, Zilles K (1996) Quantitative analysis of gyrification of cerebral cortex in dogs. Neurology 4:441–468
Zhou X, Stephens M (2012) Genome-wide efficient mixed model analysis for association studies. Nat Genet 44(7):821–824. https://doi.org/10.1038/ng.2310
Acknowledgements
We thank Laurie Santos, Richard Wrangham, David Ivy, Eliot Cohen, Kip Frey, and all other members of the Dognition.com team for their help in the creation of Dognition.com; Adam Boyko, Martin Schmidt, and James Serpell for sharing data used in this project; Stacey Tecot and David Raichlen for valuable feedback on previous drafts; and especially all of the dog owners who participated in Dognition.com experiments as citizen scientists.
Funding
DJH was supported by an Emil W. Haury Fellowship from the School of Anthropology at the University of Arizona, and a Graduate Access Fellowship from the Graduate College at the University of Arizona. ÁM was supported by the Hungarian Academy of Sciences (MTA-ELTE Comparative Ethology Research Group, MTA 01 031).
Author information
Authors and Affiliations
Contributions
BH, JC, JK, ÁM, and ELM conceived and designed the experiments. DJH and ELM analyzed the data. DJH, BH, JC, JK, ÁM, and ELM wrote the paper.
Corresponding author
Ethics declarations
Conflict of interest
BH is a founder of Dognition.com and a member of its Scientific Advisory Board. JC, JK, and ÁM are also members of the Dognition.com Scientific Advisory Board.
Data accessibility
Data are available as electronic supplementary material.
Ethical standards
All animals included in this study were pet dogs tested by citizen scientists in their own homes. The use of third-party data from Dognition.com was approved by Duke University IACUC protocol A138-11-06 and data were collected in accordance with relevant guidelines and regulations.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Horschler, D.J., Hare, B., Call, J. et al. Absolute brain size predicts dog breed differences in executive function. Anim Cogn 22, 187–198 (2019). https://doi.org/10.1007/s10071-018-01234-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10071-018-01234-1