
Abstract
The study aimed to identify two lactic acid bacteria from human breast milk and evaluate for their probiotic potential. Lactobacillus fermentum strains showed various degree of antibiotic susceptibility profile to clinical commercial antibiotics. The MA-7 and MA-8 strains have never lost their vitality at pH 2 and 3. Lactobacillus fermentum MA-8 exhibited more resistance to 0.3 and 1% bile concentrations than MA-7 strain. Both of the L. fermentum strains can tolerate the simulated gastric and intestinal juices. The strains showed high auto-aggregation percentages varied from 95 to 98%. Both isolates also exhibited gamma hemolytic activity. The cholesterol lowering activity of MA-7 and MA-8 strains was varied from 34.84 to 91.15%. The supernatants showed higher anti-cholesterol activity compared to pellets. The results demonstrated that both L. fermentum strains maybe good probiotic candidates and may have potential as health biotherapeutic with cholesterol-lowering effects.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Human milk contains various bioactive compounds involved in various microorganisms, oligosaccharides and other substances, which play important roles in host-microbial interactions for infant health (Gomez-Gallego et al., 2016). The bacterial species belong to Staphylococcus, Streptococcus, Lactobacillus, Micrococcus, Enterococcus and Bifidobacterium are generally found in healthy human breast milk (Pérez-Cano et al., 2010). Lactobacilli are important group for the healthy human intestinal microbiota and also control and maintain the microbial balance (Reuter, 2001). Lactobacillus fermentum, L. gasseri, L. rhamnosus, isolated from human breast milk, belong to the group of Lactobacilli, can be regarded as potential probiotic bacteria (Cabrera-Rubio et al., 2016; Jost et al., 2015).
Probiotics are defined by Food and Agriculture Organization (FAO) and World Health Organization (WHO) as “Live microorganisms which when administered in adequate amounts confer a health benefit on the host” (FAO/WHO, 2002) and play a role in improving gut barrier, competing with pathogenic microbiota for their adhesion and improvement of probiotic microorganism colonization (Rao et al., 2016). Bacteria isolated from human milk useful for health are considered attractive probiotic microorganisms (Martín et al., 2004).
Consumption of cholesterol rich food can lead to increased blood cholesterol levels and develop hypercholesterolaemia (Castorena-Alba et al., 2018). It has also been reported that hypercholesterolemia is a health problem associated with the dysbiosis of the intestinal microbiota (Lye et al., 2017). Probiotic microorganisms have an important role in lowering the level of serum cholesterol, which is one of the most common diseases of the modern age and consequently results in cardiovascular diseases, a major cause of death worldwide (WHO, 2018a). By 2030, WHO estimates that cardiovascular diseases will remain to be the most important cause of death and affect approximately 23.6 million people worldwide (WHO, 2018b). Functional foods containing probiotic microorganisms reduce the risk of cardiovasicular disease, and therefore, the strains of probiotics and also characterization of the bio-active properties are important (Shahidi, 2012).
The present study was aimed to carry out identification and characterization of L. fermentum originated from human breast milk and evaluated for some desirable traits in probiotic microorganisms including antibiotic sensitivity, resistance to gastrointestinal system conditions; transit tolerance, hemolytic and auto-aggregation activities. The cholesterol- lowering effects of L. fermentum isolated from different sources (koumiss, traditional dairy products, commercial yoghurt, Tulum cheese, food) have been investigated in the literature (Bao et al., 2010; Castorena-Alba et al., 2018; Pan et al., 2011; Raghavan et al., 2011; Tulumoglu et al., 2014). However, a very limited number of studies have been reported on cholesterol lowering potential of L. fermentum strains originated from human breast milk. For this reason, the cholesterol-lowering effects of the tested strains were investigated in order to determine the health promoting biotherapeutic effects.
Material methods
Characterization of isolates by API 50 CHL system
The gram-positive and catalase-negative isolates were tested for carbohydrate fermentation using API 50 CHL (bioMérieux, Marcy-l’Etoile, France) consisting of 50 biochemical tests. The inoculums adjusted to a density of Mc Farland 2 were incubated in each strip at 37 °C until 48 h. The biochemical profile of the strains was determined according to the color changes that occur as a result of fermentation of the carbohydrate sources. The identification was made by evaluating the results as positive and negative with software APIweb™.
Molecular identification
The selected strains according to the biochemical test described above were sent to Iontek (Istanbul-Turkey) to identify by 16 S rRNA PCR analysis. DNA fragments encoding 16S rRNA were amplifiedby using the universal forward (5′-AGAGTTTGATCMTGGCTCAG-3′) and reverse (3′-TACGGYTACCTTGTTACGACTT-5′) primers under following conditions: initial denaturation at 95 °C for 15 min for 30 cycles, each consisting of denaturation at 95 °C for 30 s, annealing at 56 °C for 45 s, extension at 72 °C for 40 s and final extension step at 72 °C for 10 min.
Determination of antibiotic sensitivity of strains
Antibiotic sensitivity of the strains was tested according to the method defined by Sharma et al. (2016). The susceptibility of the strains against eleven clinical commercial antibiotics (Oxoid, ThermoFisher Scientific, Wesel, Germany) (Erythromycin (15 µg), Amikacin (10 µg), Gentamycin (10 µg), Kanamycin (30 µg), Amoxicillin (30 µg), Ampicillin (10 µg), Penicillin G (10 µg), Cloxacillin (5 µg), Chloramphenicol (10 µg), Nalidixic Acid (5 µg) and Ofloxacin (5 µg)) which are frequently used in the treatment of human microorganism diseases was determined by disc diffusion method. The suspension densities of active cultures were adjusted to McFarland 0.5. The isolates (100 μL) were spread onto MRS agar (Merck, Darmstadt, Germany). Each antibiotic disc was placed on the inoculated MRS agar in triplicate and then allowed to incubate for 24 h at 37 °C. At the end of the incubation period, the inhibition zone diameters around the antibiotic discs were measured with Vernier callipers and the results were evaluated according to CLSI (Clinical and Laboratory Standards Institute) (Sharma et al., 2016).
Resistance to gastrointestinal system conditions
Acid tolerance
The tolerance to the low pH conditions was evaluated against pH 2 and 3 in MRS broth at 37 °C. Spectrophotometric (Beckman Coulter DU 730, California, USA) measurements were made at 600 nm for 0, 1 and 3 h of incubation twice for each sample. 0.1 mL inoculum was then spread onto MRS agar and incubated at 37 °C for 24 h under anaerobic conditions. The experiment was performed with three replicates. Colony counts were performed at the end of the incubation period and these values were calculated in Log10 (CFU/mL) (Zoral, 2013).
Bile tolerance
The bile tolerance of the strains was determined the method described by Bao et al. (2010) with some slight modifications. The inoculated MRS broth, containing 0.3% and 1% of bile (Oxoid, Hampshire, UK) was incubated for 4 h at 37 °C. Spectrophotometric measurements were made at 600 nm at 0 and 4 h of incubation twice for each sample. In addition, MRS agar media was inoculated from the test and control groups at these times and incubated for 24 h at 37 °C under anaerobic conditions twice for each sample. Colony counts were performed at the end of the incubation period and the values were calculated in Log10 (CFU/mL) (Tokatlı, 2013).
Simulated gastric and pancreatic juice (transit) tolerance
The simulated gastric solutions (3 g/L pepsin (Sigma-Aldrich, Saint Louis, USA) at pH 2 and 3 were prepared to determine the pepsin tolerance of the strains. The cell densities of isolates were adjusted to McFarland 0.5 standard. The isolates were inoculated (1%) into gastric solutions and incubated for 3 h at 37 °C incubation twice for each sample. MRS agar media were inoculated at the intervals of 0 and 3 h of incubation and left to incubate for 24 h under anaerobic condition. The gastric juice tolerance was determined by counting the viable cell. The survival rate was calculated in Log10 (CFU/mL) (Yürümez, 2011).
The simulated small intestine solution (1 g/L pancreatin (Sigma-Aldrich, Saint Louis, USA) and 0.03 g/L bile salt (Oxoid, Hampshire, UK) was prepared to determine the pancreatic tolerance of the strains. The cultures (McFarland 0.5 standard turbidity, 1%) were inoculated into pancreatin solution twice for each sample and incubated for 3 h at 37 °C. At 0 and 3 h of incubated groups were inoculated onto MRS agar medium and left to incubate for 24 h under anaerobic condition. At the end of the incubation period, the colonies were counted on the plate and these values were calculated in the same manner as for the determination of the gastric juice tolerance (Yürümez, 2011).
Hemolytic activity
For hemolytic activity, the strains were inoculated Colombia sheep blood agar (OR-BAK, Ankara, Turkey) and incubated at 37 °C for 24 h under anaerobic condition. The greening zone formation around the colonies was evaluated as α-hemolytic, the clear zone formation as beta hemolytic and no zone as gamma hemolytic (Eryılmaz, 2011).
Auto-aggregation activity
The cell suspension were adjusted to 0.6 ± 0.02 at 600 nm and incubated at 37 °C for 4 h without any movement in three replicates. At the end of incubation period, 0.1 mL of sample was suspended in 3.9 mL of PBS buffer. The OD values of the samples were read at 600 nm (Tomás et al., 2005). In addition, the auto-aggregation abilities of cultures were monitored under the light microscope (Leica, Wetzlar, Germany) after gram staining. The percentage of auto-aggregation was calculated using following formula:
OD1: Pre-incubation optical density, OD2: Optical density after incubation
Cholesterol assimilation
The cholesterol assimilation was determined using the “o-phthalaldehyde” method described by Hosono (1999) with some modifications. Bacterial supernatants and pellets were evaluated separately to determine the cholesterol lowering activity of the strains. The cell suspensions were adjusted to 0.6 at 600 nm. Each sample of the assimilation experiment and spectrophotometric measurements were recorded twice. MRS broth containing cholesterol (Sigma-Aldrich, Saint Louis, USA) (100 µg/mL) and bile (0.3 and 1%) (Oxoid, Hampshire, UK) were inoculated with the strains (2%) and incubated for 24 h at 37 °C. After the incubation, the supernatants were separated from the pellets. The pellets were suspended in distilled water equal volume to the previous original media. Then, 3 mL of 99% ethanol was added into 0.5 mL of supernatant or pellet and vortex-mixed. 2 mL of 50% KOH (Sigma-Aldrich, Saint Louis, USA) was added to the samples and heated in a water bath (60 °C) for 10 min and then cooled at room temperature. 5 mL of hexane was added to the samples and the mixture was stirred for 20 s. Distilled water (3 mL) was added to each sample and gently mixed. The samples were kept at room temperature for 15 min. 2.5 mL of the hexane layer was taken and completely evaporated in a water bath at 60 °C. 4 mL of o-phthalaldehyde reagent (Sigma-Aldrich, Saint Louis, USA) was added to each tube. After 10 min, 2 mL of pure sulfuric acid (Merck, Darmstadt, Germany) was added onto the samples. The samples were mixed and then incubated at room temperature for 10 min. Absorbance was read using a spectrophotometer at 550 nm. Cholesterol assimilation was calculated using the equation:
A: Cholesterol assimilation (%), B: The amount of cholesterol in the nutrient medium inoculated (µg), C: The amount of cholesterol in the non-inoculated medium (µg)
Statistical analysis
The data obtained for antibiotic susceptibility, pH and bile tolerance, simulated gastric and pancreatic juices resistance were analyzed by using the Mann–Whitney U-test. The cholesterol assimilation data was subjected to statistical analysis for checking the significance correlation using Origin Graphing & Analyzing 2017. The statistical significance was accepted at p < 0.005 and r = 1.
Results and Discussions
Identification of Lactic Acid Bacteria
The strains were identified by API 50 CHL System as L. fermentum MA-7 (93.7%) and L. fermentum MA-8 (99.6%). The strains were also identified molecularly. According to the 16S rRNA sequences, the results confirmed that the isolated two bacteria were L. fermentum.
Antibiotic sensitivity
L. fermentum MA strains were tested for their sensitivity against antibiotics which are commonly used on various pathogenic microorganisms. The strains were sensitive to Amoxicillin, Ampicillin, Erythromycin and Penicillin G. The strains exhibited moderate sensitivity to Chloramphenicol. The two strains showed resistance against Amikacin, Gentamicin, Kanamycin, Nalidixic Acid, Ofloxacin and Cloxacillin (Table 1). The means for Amoxicillin inhibition zone values were 31.463 mm and 25.433 mm for L. fermentum MA-7 and MA-8 strains. The average of MA-7 is larger than the MA-8 strain. There are mean differences between the observational data of antibiotic susceptibility of MA-7 and MA-8 strains for Amoxicillin antibiotic. However, the statistical analysis indicated that there was no statistically significant difference in the significance level of 0.05 (p > 0.05). The probable cause is that both bacteria tested are same bacteria (L. fermentum) but different strains (MA7 and MA-8 strains).
The most important factor in the safety of probiotics is sensitivity to antibiotics. A potential risk of probiotic application is the spread of antibiotic resistance genes to pathogenic bacteria because of horizontal transfer of the antibiotic resistance gene (Courvalin, 2006). However, it has been reported that probiotic microorganisms contain an antibiotic resistance gene in the genomic structure is not a safety concern in itself, as long as the genes are not mobilized and transferred horizontally to other bacteria (Darsanaki et al., 2013). As a result, the resistance of L. fermentum MA strains may supply the viability of these strains during treatment with these clinical antibiotics and therefore strains may be used for probiotic and therapeutic purposes.
Resistance to gastrointestinal system conditions
Tolerance to low pH conditions
The results of low pH conditions (pH 2 and 3) on the MA strains are presented in Table 2 as spectrophotometrically and cell viability. A probiotic strain is primarily expected to be tolerant to the low pH conditions of stomach ranges from 1.5 to 4.5 during fasting and after meal, and food digestion is carried out for 3 h (Jacobsen et al., 1999; Lebeer et al., 2008). The strains have never lost their viability at pH 2 and 3. The spectrophotometric and cell viability mean values of MA-7 and MA-8 strains are different for control group at 3rd h in the Table 2. The averages of MA-7 strains (0.730 and 8.43) were different from MA-8 strains (0.276 and 10.27) for experiments but there was no statistically significant difference between two strains in the significance level of 0.05 (p > 0.05). Khalil et al. (2018) found the survival ability of L. fermentum DUR 18 originated from fermented food at pH 3.0 conditions as 6.37 Log10 CFU/mL with 86.20% survival rate. Lactobacillus fermentum CH58 isolated from Brazilian food products reported by Ramos et al. (2013), was able to growth at pH2.0 after 3 h. Our results indicated that L. fermentum MA strains are able to survive under low pH conditions; therefore, they are likely to be able to viable through the gastrointestinal system.
Tolerance to bile
Strains that pass the acidity barrier of the stomach, in the next step, they expose to bile fluid. For this reason, tolerance to bile is another criteria to identified them as probiotic (Ammor and Mayo, 2007; Khater et al., 2010). The survival in different bile salt concentrations was varied depend on the strain. The spectrophotometric mean values of the bile tolerance for L. fermentum MA-7 and MA-8 strains were 0.106 and 0.055 at 0.3% bile concentration at 4th h in the Table 3. The observational averages are different but there were no statistically significant differences in the significance level of 0.05 in bile tolerance mean values between the MA strains, neither at spectrophotometric, nor viable cell value averages. Also, there was no difference statistically between spectrophotometric value means at 1% bile concentration at 0 h. The survival rate of two MA strains was approximately or over 100% for both bile concentrations throughout the incubation period (4 h) (Table 3). These results are similar to the results obtained by Ramos et al. (2013), found that L. fermentum CH58 viable cell numbers increased from 4.46 to 6.07 during the condition of 0.3% bovine bile salts for 3 h. Khalil et al. (2018) reported that the strain L. fermentum DUR 18 exhibited ability to survive at 0.3% bile salt for 3 h. Lactobacillus fermentum UCO-979C, isolated from a human gastric biopsy, showed tolerance [4.2 × 106 CFU/mL (6.62 log 10)] to 2.0% bile salts through 2 h (Garcíaet al., 2017). The resistance to bile salts in the gastrointestinal tract is an essential property to evaluate the ability of the strains as probiotic (Ren et al., 2014). The ability to survive of the L. fermentum MA strains at different bile conditions make them suitable for use as probiotic.
Tolerance to gastric and pancreatic juice
The effects of gastric and pancreatic juice on the viability of L. fermentum MA strains are presented in the Table 4. The tolerance of the MA strains to artificial gastric juice increased from pH 2.0 to pH 3.0. The survival rates of L. fermentum MA-8 for both acidic conditions (pH 2.0 and 3.0) were higher than MA-7 strain in gastric solution. Although its higher resistance to pancreatic juice, there is a bit difference between the observational data of simulated gastric and pancreatic juices resistance in the Table 4. The statistical analysis also confirmed that there was no significant difference between of the tested two L. fermentum MA strains (p > 0.05). The possible cause of is that both bacteria tested are the same bacteria but different strains.
About 2.5 L of gastric juice at a pH of about 2.0 and 0.7 L of pancreatic juice at approximately pH 8.0 are secreted into stomach and small intestine of human each day (Charteris et al., 1998). These secretions, low pH of the stomach and the antimicrobial activity of the pepsin are important barriers to microorganisms entering the intestinal tract and colonizing (Huang and Adams, 2004). The Joint FAO/WHO Working Group Report suggested that microorganisms should have minimum characteristics that can predict the probiotic potential include resistance to passage through the stomach in the presence of acid and pepsin, and the viability in the small intestine in the presence of pancreatin and bile salts (FAO/WHO, 2002). In a study conducted by Bao et al. (2010), the viability of L. fermentum F6 originated from traditional dairy products in simulated gastric juice at pH 2.5 was 86.7% after 3 h. The high tolerance of L. fermentum MA-7 and 8 strains to gastric and pancreatic juices indicated that the MA strains could be resistant to the effects of pepsin during gastric transit and pancreatin during the small intestinal transit.
Auto-aggregation activity
The auto-aggregation ability of the bacterial strains used as probiotics is an important feature to adhere to epithelial cells (Collado et al., 2008). Lactobacillus fermentum MA-7 and MA-8 strains showed percentage of auto-aggregation 95% and 98%. These results are higher than that demonstrated by Ramos et al. (2013) who reported that L. fermentum CH58 had 55.61% auto-aggregation or that reported by Collado et al. (2008), who demonstrated auto-aggregation after 4 and 24 h for L. fermentum ME3 strain as 29.7% and 70.2%. The MA-7 and MA-8 strains with their high auto-aggregation ability can be a physico-chemical barrier by allowing the formation of biofilms in the gastrointestinal tract (Castagliuolo et al., 2005) and can be good barrier against colonization of pathogenic bacteria.
Hemolytic activity
Hemolytic activity of lactic acid bacteria is an important in vitro selection criterion and one of the safety requirements to evaluate as probiotic strain. Non-hemolytic (γ-hemolytic) activity is considered as safety precondition to select a probiotic strain (FAO/WHO, 2002). L. fermentum MA-7 and MA-8 did not exhibit hemolytic activity. Similar to our results, Owusu-Kwarteng et al. (2015) observed 16 L. fermentum strains as α- and γ-hemolytic. García et al. (2017) indicated L. fermentum UCO-979C as α-hemolytic. The MA strains, showed no red blood cell lysis activity, can be considered safe for hemolytic activity.
Cholesterol assimilation
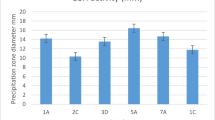
The cholesterol assimilating activity at different bile concentrations (0.3% and 1%) of L. fermentum MA-7 and MA-8 strains was determined for pellets and supernatants. The MA strains were able to assimilate cholesterol ranging from 34.84 to 91.15% (Fig. 1). According to the obtained data, it was observed that L. fermentum MA-7 and MA-8 strains had significant cholesterol assimilation activities even at the increased bile rate. In addition, statistical data were correlated with the percentage of cholesterol assimilation data (p < 0.05). The pellet and supernatant of L. fermentum MA-8 showed higher cholesterol lowering activity for both bile concentrations than MA-7 strain. The supernatant of MA-8 strain had higher anti-cholesterol activity than its pellet at both bile conditions. The similar results were also observed for cholesterol lowering activity of L. fermentum MA-7. The supernatant of MA-7 had higher cholesterol assimilating activity when compared to its pellet in the presence of two bile concentrations. Hypercholesterolemia is an important health problem and can cause death in humans. The properties of anti-cholesterol of some probiotic lactic acid bacteria have been reported in the literature. Castorena-Alba et al. (2018) reported the anti-cholesterol activity of L. fermentum food isolates strains as 57.65% at 4% concentration of oxgall and cholesterol (100 μg/mL). Tulumoglu et al. (2014) reported cholesterol assimilation ability of seven L. fermentum strains isolated from Tulum cheese ranging from 12.1 to 45.3%. In our previous study, the various Lactobacillus gasseri strains originated from human breast milk showed cholesterol assimilation ranged from 23.67 to 88.94% (Gunyakti and Asan-Ozusaglam, 2018).

Cholesterol lowering activity of L. fermentum MA strains. The cholesterol assimilation activity of the strains was studied in two replicates according to the o-phthalaldehyde method. (A) Spectrophotometric results of cholesterol amount in media for MA-7 strain; mean ± SD. (B) Spectrophotometric results ofcholesterol amount in media for MA-8 strain characterize mean ± SD. (C) Percentage of cholesterol assimilation values of MA-7 and MA-8 strains
The likely reason is that Lactobacillus strains can lower cholesterol by decreasing the solubility of cholesterol and thereby reducing cholesterol uptake (Oh et al., 2018). It has been reported that the cholesterol degradation of Bacillus subtilis SFF34, a non-probiotic strain, originated from fermented seafood, was 31% and 50% of the cholesterol in the medium after 36 and 60 h incubation (Kim et al., 2002). The results revealed that L. fermentum MA strains exhibit high cholesterol lowering activities. This suggests that both strains are important in natural bio-therapy for reducing hypercholesterolemia and related cardiovascular diseases.
In conclusion, L. fermentum MA-7 and MA-8 strains are safe due to gamma hemolytic activity, and resistant to low pH, bile and gastric and intestinal conditions in the gastrointestinal tract. Both L. fermentum have high-level colon epithelial adhesion activity, indicating that they can stay in the gastrointestinal tract for a long time. Currently, the development of cholesterol-lowering bio-therapeutics is an important area. L. fermentum MA-7 and MA-8 can achieve elimination of cholesterol in the gastrointestinal tract with good cholesterol-lowering effects. L. fermentum MA-7 and MA-8 can be suggested as probiotic candidates and bio-agents after further studies.
References
Ammor MS, Mayo B. Selection criteria for lactic acid bacteria to be used as functional starter cultures in dry sausage production: An update. Meat Sci. 76: 138–146 (2007)
Bao Y, Zhang Y, Zhang Y, Liu Y, Wang S, Dong X, Wang Y, Zhang H. Screening of potential probiotic properties of Lactobacillus fermentum isolated from traditional dairy products. Food Control. 21: 695–701 (2010)
Cabrera-Rubio R, Mira-Pascual L, Mira A, Collado MC. Impact of mode of delivery on the milk microbiota composition of healthy women. J. Dev. Orig. Health Dis. 7: 54–60 (2016)
Castagliuolo I, Galeazzi F, Ferrari S, Elli M, Brun P, Cavaggioni A, Tormen D, Sturniolo GC, Morelli L, Palu` G. Beneficial effect of auto-aggregating Lactobacillus crispatus on experimentally induced colitis in mice. FEMS Immunol. Med. Microbiol. 43: 197–204 (2005)
Castorena-Alba MM, Vázquez-Rodríguez JA, López-Cabanillas Lomelí M, González-Martínez BE. Cholesterol assimilation, acid and bile survival of probiotic bacteria isolated from food and reference strains. Cyta J. Food. 16: 36–41 (2018)
Charteris WP, Kelly P, Morelli L, Collins JK. Development and application of an in vitro methodology to determine the transit tolerance of potentially probiotic Lactobacillus and Bifidobacterium species in the upper human gastrointestinal tract. J. Appl. Microbiol. 84: 759–768 (1998)
Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing: Twenty- Second Informational Supplement. CLSI Document M100-S22, Clinical Laboratory Standard Institute, Wayne, PA. ISBN 1-56238-785-5, (Print); ISBN 1-56238-786-3(2012) (Electronic)
Collado MC, Meriluoto J, Salminen S. Adhesion and aggregation properties of probiotic and pathogen strains. Eur. Food Res. Technol. 226: 1065–1073 (2008)
Courvalin P. Antibiotic resistance: The pros and cons of probiotics. Dig. Liver Dis. 38: 261–265 (2006)
Darsanaki RK, Aliabadi MA, Chakoosari MMD. Antibiotic resistance of lactic acid bacteria. Sci. J. Microbiol. 2: 201–206 (2013)
Eryılmaz TF. Identification of potential probiotic properties of some lactic acid bacterial genera which is isolated from vaginal secretion. PhD thesis, University of Ankara, Ankara, Turkey (2011)
FAO/WHO. Guidelines for the evaluation of probiotics in food. World Health Organization and Food and Agriculture Organization of the United Nations, London Ontario, Canada (2002)
García A, Navarro K, Sanhueza E, Pineda S, Pastene E, Quezada M, Henríquez K, Karlyshev A, Villena J, González C. Characterization of Lactobacillus fermentum UCO-979C, a probiotic strain with a potent anti-Helicobacter pylori activity. Electron. J. Biotechnol. 25: 75–83 (2017)
Gomez-Gallego C, Garcia-Mantrana I, Salminen S, Collado MC. The human milk microbiome and factors influencing its composition and activity. Semin. Fetal Neonatal Med. 21: 400-405 (2016)
Gunyakti A, Asan-Ozusaglam M. Investigation of the potential use of Lactobacillus gasseri originated from human breast milk as food additive. LWT Food Sci. Technol. 93: 613–619 (2018)
Hosono UA. Bile tolerance, taurocholate deconjugation, and binding of cholesterol by Lactobacillus gasseri strains. J. Dairy Sci. 82: 243–248 (1999)
Huang Y, Adams MC. In vitro assessment of the upper gastrointestinal tolerance of potential probiotic dairy propionibacteria. Int. J. Food Microbiol. 91: 253–260 (2004)
Jacobsen CN, Nielsen VR, Hayford AE, Møller P L, Michaelsen KF, Paerregaard A, Sandström B, Tvede M, Jakobsen M. Screening of probiotic activities of forty seven strains of Lactobacillus spp. by in vitro techniques and evaluation of colonization ability of five selected strains in human. Appl. Environ. Microbiol. 65: 4949–4956 (1999)
Jost T, Lacroix C, Braegger C, Chassard C. Impact of human milk bacteria and oligosaccharides on neonatal gut microbiota establishment and gut health. Nutr. Rev. 73: 426–437 (2015)
Khalil ES, Abd Manap MY, Mustafa S, Alhelli AM, Shokryazdan P. Probiotic Properties of exopolysaccharide-producing Lactobacillus strains isolated from Tempoyak. Molecules. 23: 398 (2018)
Khater KAA, Ali MA, Ahmed EAM. Effect of encapsulation on some probiotic criteria. J. Am. Sci. 6: 836–845 (2010)
Kim KP, Rhee CH, Park HD. Degradation of cholesterol by Bacillus subtilis SFF34 isolated from Korean traditional fermented flatfish. Lett. Appl. Microbiol. 35: 468–472 (2002)
Lebeer S, Vanderleyden J, De Keersmaecker SCJ. Genes and molecules of lactobacilli supporting probiotic action. Microbiol. Mol. Biol. Rev. 72: 728–764 (2008)
Lye HS, Kato T, Low WY, Taylor TD, Prakash T, Lew LC, Liong MT. Lactobacillus fermentum FTDC 8312 combats hypercholesterolemia via alteration of gut microbiota. J. Biotechnol. 262: 75–83 (2017)
Martín R, Langa S, Reviriego C, Jimenez E, Marin LM, Olivares M, Boza J, Jimenez J, Fernandez L, Xaus J, Rodriguez JM. The commensal microflora of human milk: New perspectives for food bacteriotherapy and probiotics. Trends Food Sci. Technol. 15: 121–127 (2004)
Oh NS, Joung JY, Lee JY, Kim Y. Probiotic and anti-inflammatory potential of Lactobacillus rhamnosus 4B15 and Lactobacillus gasseri 4M13 isolated from infant feces. PLoS One. 13: e0192021 (2018)
Owusu-Kwarteng J, Tano-Debrah K, Akabanda F, Jespersen L. Technological properties and probiotic potential of Lactobacillus fermentum strains isolated from West African fermented millet dough. BMC Microbiol. 15: 261 (2015)
Pan DD, Zeng XQ, Yan YT. Characterization of Lactobacillus fermentum SM-7 isolated from koumiss, a potential probiotic bacterium with cholesterol-lowering effects. J. Sci. Food Agric. 91: 512–518 (2011)
Pérez-Cano FJ, Dong H, Yaqoob P. In vitro immunomodulatory activity of Lactobacillus fermentum CECT5716 and Lactobacillus salivarius CECT5713: Two probiotic strains isolated from human breast milk. Immunobiology. 215: 996–1004 (2010)
Rao SC, Athalye-Jape GK, Deshpande GC, Simmer KN, Patole SK. Probiotic supplementation and late-onset sepsis in preterm infants: a meta-analysis. Pediatrics. 137: 1–16 (2016)
Raghavan CM, Nanda A, Yuvaraj R, Murugan AS, Raja RB. Assimilation of cholesterol by Lactobacillus species as probiotics. World Appl. Sci. J. 14: 552–560 (2011)
Ramos CL, Thorsen L, Schwan RF, Jespersen L. Strain-specific probiotics properties of Lactobacillus fermentum, Lactobacillus plantarum and Lactobacillus brevis isolates from Brazilian food products. Food Microbiol. 36: 22–29 (2013)
Ren D, Li C, Qin Y, Yin R, Du S, Ye F, Liu C, Liu H, Wang M, Li Y. In vitro evaluation of the probiotic and functional potential of Lactobacillus strains isolated from fermented food and human intestine. Anaerobe. 30: 1–10 (2014)
Reuter G. The Lactobacillus and Bifidobacterium microbiota of the human intestine: composition and succession. Curr. Issues Intest. Microbiol. 2: 43–53 (2001)
Shahidi F. Nutraceuticals, functional foods and dietary supplements in health and disease. J. Food Drug Anal. 20: 226–230 (2012)
Sharma P, Tomar SK, Sangwan V, Goswami P, Singh R. Antibiotic resistance of Lactobacillus sp. isolated from commercial probiotic preparations. J. Food Saf. 36: 38–51 (2016)
Tokatlı M. Identification of lactic acid bacteria isolated from pickles in Ankara Çubuk region, determination of their technological and functional properties and their potential for using as a starter culture, PhD thesis, University of Ankara, Ankara, Turkey (2013)
Tomás J, Wiese B, Nader-Macías ME. Effects of culture conditions on the growth and auto-aggregation ability of vaginal Lactobacillus johnsonii CRL 1294. J. Appl. Microbiol. 99: 1383–1391 (2005)
Tulumoğlu Ş, Kaya Hİ, Şimşek Ö. Probiotic characteristics of Lactobacillus fermentum strains isolated from Tulum cheese. Anaerobe. 30: 120–125 (2014)
WHO. Cardiovascular diseases (CVDs). Available from http://www.who.int/news-room/fact sheets/detail/cardiovascular-diseases-(cvds). Accessed Dec. 05, 2018a
WHO. Cardiovascular diseases. Available from http://www.wpro.who.int/mediacentre/factsheets/cardiovascular_disease/en/. Accessed Dec. 06, 2018b
Yürümez E. Probiotics properties of some lactic acid bacteria isolated from faeces samples. MSc thesis, University of Ankara, Ankara, Turkey (2011)
Zoral S. İnsan kaynaklı Lactobacillus spp suşlarının probiyotik özelliklerinin belirlenmesi. MSc thesis, University of Ahi Evran, Kırşehir, Turkey (2013)
Acknowledgements
This work was financially supported by The Scientific Research Projects Coordinator of Aksaray University-Turkey (Project number 2017/031).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors of the article declare no conflict of interest.
Rights and permissions
About this article

Cite this article
Asan-Ozusaglam, M., Gunyakti, A. Lactobacillus fermentum strains from human breast milk with probiotic properties and cholesterol-lowering effects. Food Sci Biotechnol 28, 501–509 (2019). https://doi.org/10.1007/s10068-018-0494-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10068-018-0494-y