
Abstract
Many disturbances are shifting in severity, frequency, and extent due to changing climate and human activities. Altered disturbance regimes can trigger shifts in ecosystem state where recovery to the pre-disturbance ecosystem is uncertain. In the western North American boreal forest, the intensification of wildfire can cause canopy dominance to switch from black spruce (Picea mariana) to deciduous trees such as Alaska paper birch (Betula neoalaskana) and trembling aspen (Populus tremuloides). Understanding the key mechanisms that determine the resilience and stability of these alternative community types is required for the prediction of future forest dynamics. Here, we assess patterns of post-fire tree recovery across a pre-fire gradient of spruce- to deciduous-dominated forests in Interior Alaska and quantify compositional and environmental thresholds that support the resilience of alternative canopy types. We found post-fire organic soil depth of stands on a recovery trajectory to deciduous dominance (7.3 ± 5.5 cm) were similar regardless of pre-fire composition and significantly shallower than spruce (14.9 ± 9.0 cm) or mixed trajectories (10.4 ± 5.9 cm). Deciduous-dominated stands were highly resilient to fire, as 100% remained deciduous-dominated post-fire. Even when deciduous trees only accounted for a small proportion (12%) of the pre-fire stand, deciduous trees often became dominant after wildfire. We conclude that the establishment of deciduous bud banks and seed sources creates a strong hysteresis in stand recovery that reinforces the resilience of deciduous-dominated boreal forests to wildfire. Accounting for the resilience of this alternative stable state to wildfire suggests that shifts from spruce to deciduous dominance caused by shifting wildfire will have long-term effects on future structure and function of boreal forests and vegetation feedbacks to climate change.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Highlights
-
Ecological legacies reinforce resilience of deciduous boreal forests to wildfire.
-
Deciduous legacies create a strong hysteresis in post-fire stand recovery.
-
Fire induced shifts from spruce to deciduous will persist across fire cycles.
Introduction
Patterns of disturbance and recovery shape the structure and function of many ecosystems and play a central role in driving ecosystem response to global change (Seidl and others 2014; McDowell and others 2020). The ability of ecosystems to recover their essential structure and function after a disturbance (that is, ecosystem resilience) is transmitted as legacies of past ecosystems via species traits and materials (Johnstone and others 2016). Many disturbances are exhibiting shifts in their severity, frequency, and extent due to changing climate and human activities (Mollicone and others 2006; Abatzoglou and Williams 2016). These altered disturbance regimes can modify key legacies (for example, seedbanks, seedbeds) and trigger shifts to alternative vegetation states that exhibit hysteresis (Ratajczak and others 2018), where recovery to the pre-disturbance ecosystem is unlikely (Johnstone and others 2010b; Kukavskaya and others 2016; Whitman and others 2019; Coop and others 2020). New ecosystem states may produce novel legacies that will similarly constrain future responses to disturbance, potentially inhibiting recovery to the original state even if external drivers return to prior conditions (Ratajczak and others 2018). Understanding how changes in ecological legacies interact with disturbances is essential for predicting dynamic ecosystem responses to global change (Foster and others 2022b).
Wildfire is the primary large-scale disturbance throughout the northwestern North American boreal forest. Much of this forest is dominated by conifer stands of black spruce (Picea mariana), where stand-replacing fires have historically occurred at approximately 100-year intervals (Johnstone and others 2010a). Forest succession under such short fire cycles is typically driven by early dominance of the post-fire cohort (Gutsell and Johnson 2002), although relay succession of stand composition is possible under longer fire-free intervals (for example, Taylor and Chen 2011). The fire and climate regimes of this region have maintained persistent cycles of black spruce self-replacement for the past ~ 6000 years, indicating a high level of ecosystem resilience to wildfires (Lloyd and others 2006; Higuera and others 2009; Kelly and others 2013; Hoecker and others 2020).
Resilience of black spruce stands depends on legacies of the soil organic layer (SOL) and plant adaptations to disturbance, such as the semi-serotinous cones of black spruce triggered by fire to release stored seed (Johnstone and others 2016). Climate warming and drying in the boreal forest have led to increases in fire severity and frequency (Kelly and others 2013). This can reduce or eliminate the SOL and cause canopy dominance to switch from black spruce to deciduous trees such as Alaska paper birch (Betula neoalaskana) and trembling aspen (Populus tremuloides) (Whitman and others 2019; Johnstone and others 2020; Baltzer and others 2021; Hayes and Buma 2021). Post-fire regeneration patterns of deciduous trees depend on different legacies than black spruce. For successful post-fire regeneration via deciduous seed, mast seeding years must coincide with fires. However, both aspen and birch can regenerate prolifically and rapidly via vegetative reproduction (Greene and Johnson 1999; Frey and others 2003; Chen and others 2009). Once deciduous trees establish via seed following severe wildfire, their asexual regeneration strategies may reinforce the shift in boreal forest composition from the historic black spruce state to deciduous dominance.
As fire regimes continue to intensify in the North American boreal forest (Yue and others 2015; Young and others 2017), changes in forest composition from conifer dominance to alternative deciduous forest types could have important implications for biome C storage, wildlife habitat, forest products, future fire hazard, and feedbacks to earth system dynamics. However, under the rapidly changing environments of the twenty-first century, we often lack sufficient historical analogs to anticipate the resilience of long-lived organisms such as trees to intensifying disturbance regimes. For example, cold soils in high latitude or elevational areas have previously been understood to constrain deciduous-dominated states to a transient stage in successional recovery to conifer dominance (Cleve and others 1991). However, changing climate and fire conditions are altering both drivers and ecosystem feedbacks that may alter the stability of forest types (Johnstone and others 2016; Mekonnen and others 2019). Prediction of future forest dynamics thus depends on understanding the key mechanisms that will determine the resilience and stability of historic or alternative community types (Foster and others 2019; Mekonnen and others 2019). In the North American boreal forest, the potential for shifts from conifer to deciduous broadleaf forests, and their potential to mitigate C losses from fire and future fire risk, depends on whether demographic processes are sufficient to support stable persistence of deciduous stands under future disturbances.
Here, we assess post-fire regeneration across a gradient of spruce- to deciduous-dominated forests in Interior Alaska and quantify compositional and environmental thresholds that support resilience of alternative vegetation types to fire. Specifically, we determine the impact of pre-fire deciduous presence on post-fire recruitment patterns. We hypothesize that pre-fire deciduous presence will result in post-fire deciduous dominance due to reproductive legacies that reinforce the alternate successional trajectory of deciduous-dominated boreal forests. Given the known importance of SOL depth for black spruce self-replacement (for example, Johnstone and others 2010b), we also examine differences in post-fire SOL depth among successional trajectories. Our results not only demonstrate vulnerability of spruce to post-fire replacement by deciduous trees, but also underscore high resilience of mixed and deciduous-dominated boreal forests to wildfire, thereby providing a basis for anticipating the consequences of altered successional trajectories for future forest dynamics.
Methods
Study Area
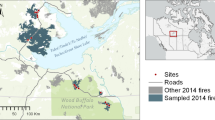
Our study area was the boreal forest ecosystem that lies south of the Brooks Range and north of the Alaskan Range in Interior Alaska, USA (Figure 1). Temperatures across this region are highly continental and seasonally range from − 50 to 38 °C with mean annual temperature of − 3.3 °C and mean annual precipitation of ~ 285 mm, including ~ 40% from snow (Fairbanks International Airport, 1985–2015). This region is underlain by discontinuous permafrost, and soils are undeveloped, primarily (~ 90%) consisting of Inceptisols, Gelisols, Histosols, and Entisols (Osterkamp and Romanovsky 1999). Approximately 57% of the ~ 370,000 km2 Alaskan boreal forest area is dominated by coniferous trees, while 27% is a mix of conifers and deciduous tree species, such as aspen and Alaska paper birch, and 16% is unforested wetland and tundra (Yarie and Billings 2002; Neigh and others 2013).

Sample sites and associated fire perimeters in the boreal forest of Interior Alaska. Forest deciduous fraction in 2000 is also shown for areas with > 20% tree cover based on maps by Massey and others (in review). Fire perimeters are from the Alaska Large Fire database.
Field Methods
We collected and collated data on pre- and post-fire stand composition from 212 sites across Interior Alaska that burned between the years 2004 and 2014 (Figure 1). Sites were established by three independent research projects, but field measurements on pre- and post-fire trees were similar (Walker and others 2017; Johnstone and others 2020). Of the 212 sites examined, we established 88 pre-fire black spruce dominated sites in three separate fire complexes following the 2004 wildfires in Interior Alaska (details in Johnstone and others 2020; Mack and others 2021). In 2010 and 2011, we established an additional 82 sites in the same 2004 fire scars, but sites ranged in pre-fire composition from black spruce dominance to black spruce co-dominance with deciduous species (details in Walker and others 2017). In 2019, we established 42 new sites dispersed between six independent fire scars that were largely deciduous dominated pre-fire. We chose these sites using the Alaska large fire database and forest deciduous fraction maps (Massey and others In Revision). All sites were mature (> 70 years since previous fire) at the time of fire based on historical record of fires in the Alaskan Large Fire Database and confirmed with stand age reconstructions using tree ring analyses. Sampling took place 4–13 years after fire, once the regeneration patterns that set the trajectory of early succession have established (Johnstone and others 2020).
We estimated pre- and post-fire stem density and forest composition by measuring pre-fire (burned) trees and newly established seedlings. We counted all standing or fallen pre-fire trees and measured diameter at breast height (dbh) on all individuals taller than 1.4 m or basal diameter on all trees less than 1.4 m in height that were originally rooted in two parallel 2 m × 30 m belt-transects. Similarly, we estimated post-fire density by counting post-fire seedlings within ten randomly positioned 1 m × 1 m quadrats along the same two transects and measuring the basal diameter of at least one representative species per quadrat. We counted all post-fire stems of spruce, aspen, and birch that were rooted in quadrats, regardless of size. Given the range of time after fire that we sampled (4–13 years) we were unable to differentiate seedlings from vegetative regeneration.
Classification of Successional Trajectories
Using the survey data, we calculated total density (stems ha−1) of each tree species pre- and post-fire. We used published allometric equations to calculate pre-fire (Alexander and Mack 2016) and post-fire (Johnstone and others 2020) species-specific aboveground tree biomass (g m−2). We then calculated the proportion of deciduous stems relative to total stems in both the pre- and post-fire stand (relative deciduous density) and the proportion of deciduous biomass relative to total biomass in both the pre- and post-fire stand (relative deciduous biomass). We classified sites into pre- and post-fire tree species dominance classes based on a deciduous fraction index (DI) calculated from the survey data (DI = relative density plus relative biomass of deciduous tree seedlings divided by two and multiplied by 100). When DI was ≤ 33.33%, sites were classified as black spruce (hereafter Spruce). Sites were classified as mixed black spruce-deciduous (hereafter Mixed) if DI was > 33.33% and < 66.66%, and as deciduous if DI was ≥ 66.66%. We used biomass rather than the more typical basal area because biomass accounts for the different allometric scaling relationships between black spruce and deciduous tree species. Based on pre- and post-fire DI and spruce density, we categorized sites in seven classes representing observed transitions in stand dominance after fire: (1) Spruce to Deciduous, (2) Spruce to Mixed, (3) Spruce to Spruce, (4) Spruce thinning (where stands remained spruce dominated after fire but did not regenerate with sufficient density for self-replacement to occur), (5) Mixed to Mixed, (6) Mixed to Deciduous, and 7) Deciduous to Deciduous.
Statistical Analyses
All analyses were performed using R statistical software version 3.5.2 (R Development Core Team 2018) and model results were plotted using the R package ‘ggplot2’ (Wickham 2016). To assess the impact of pre-fire DI on post-fire DI, we fit a generalized additive model (GAM) in the R package ‘mgcv’ (Wood 2017). GAMs are a nonparametric technique that fit smooth relationships between response and predictor variables (Wood 2017). The smooth for pre-fire DI was fit using thin plate splines and we set the basis dimension to 10 to allow for potentially complex nonlinear relationships. We used a Gaussian distribution and random intercept of fire scar to account for the spatial dependence of sites nested within fire scars. We also used restricted maximum likelihood for automated selection of smoothing parameters (Wood 2011). Model diagnostics were completed using the ‘gratia’ package (Simpson 2022).
To examine differences in post-fire SOL depth among the seven successional trajectories we fit linear mixed effects models (LMMs) using the R package ‘nlme’ (Pinheiro and others 2022). We included a random intercept of fire scar (nine levels) to account for the spatial non-independence of sites within fire scars and a variance structure (VarIdent) to account for changing variance with trajectory that was apparent in the model residuals (Zuur and others 2009). The significance of the fixed effect was assessed using a likelihood ratio test of the full model against the reduced model and verified using AIC (Zuur and others 2009). We confirmed that the statistical assumptions of homogeneity of variance and independence were not violated by visually inspecting residual versus fitted values, the explanatory variable, and each grouping level of the random intercept (Zuur and others 2009). We used Tukey–Kramer post hoc analysis for multiple comparisons in the R package ‘emmeans’ (Lenth and others 2019) with a Bonferroni adjustment to test for differences in marginal means of SOL depths between post-fire trajectories within the same pre-fire class (for example, Spruce to Spruce vs. Spruce to Mixed) and among the same post-fire trajectory with different pre-fire classes (for example, Spruce to Deciduous vs. Deciduous to Deciduous).
Results
Stands with > 12% pre-fire DI were likely to transition to deciduous dominance after fire (that is, post-fire DI was predicted to be > 66%) (Figure 2; pink line). Stands with > 17% pre-fire DI were expected to be exclusively deciduous dominated post-fire based on confidence intervals of fitted relationships (Figure 2; purple line). Out of 157 pre-fire spruce stands, we found 43% transitioned to deciduous dominance and 32% to mixed spruce-deciduous, whereas only 18% remained spruce dominant following fire. The final 7% of pre-fire spruce stands remained spruce dominated after fire but post-fire density was insufficient for self-replacement (that is, post-fire spruce density was lower than pre-fire spruce density; thinning). Notably, 100% of the 35 pre-fire deciduous stands self-replaced, while 93% of the 43 mixed stands transitioned to deciduous dominance, with the remaining 7% of sites continuing as mixed spruce-deciduous post-fire. Our results show that deciduous dominated forests are highly resilient to fire and that even a small component of deciduous stems (~ 12%) in the pre-fire stand can result in a transition of spruce forests to deciduous dominance in the first two decades post-fire.

Post-fire deciduous fraction index as a function of pre-fire deciduous fraction index. Shapes represent different pre-fire species composition (circle = spruce, square = mixed, triangle = deciduous) and colors indicate post-fire succession trajectory (purple = thinning, blue = spruce, green = mixed, and yellow = deciduous). Black line represents generalized additive model fit with shading for the 95% confidence intervals. The pink line represents the threshold in pre-fire DI from which post-fire DI > 66% is likely to emerge based on model fit. The purple line represents the threshold in pre-fire DI from which post-fire DI > 66% will emerge with 95% confidence based on model fit.
Post-fire SOL depth differed between spruce stands that remained spruce versus transitioned to deciduous dominance following wildfire (Figure 3). Specifically, spruce stands that remained spruce had an average post-fire SOL of depth of 14.9 ± 9.0 cm, significantly (p value < 0.05) higher than spruce stands that transitioned to deciduous dominance (8.4 ± 6.4 cm). Spruce stands that transitioned to mixed or thinned had intermediate post-fire SOL depths of 10.7 ± 6.0 cm, and 14.6 ± 8.9 cm, respectively. Post-fire SOL depth of stands on a deciduous recovery trajectory (7.3 ± 5.5 cm) was similar regardless of pre-fire composition. Similarly, post-fire SOL depth of mixed stands (10.4 ± 5.9 cm) did not differ between stands that were pre-fire mixed or pre-fire deciduous (Figure 3).

Post-fire soil organic layer (SOL) depth (cm) grouped according to post-fire trajectory (colors) and pre-fire class (panels). Colors indicate post-fire succession trajectory (purple = thinning, blue = spruce, green = mixed, and yellow = deciduous). Three panels represent pre-fire classes (deciduous, mixed, spruce) based on deciduous index (see classification of successional trajectories section in methods). Different letters above boxplots indicate significant differences (p value < 0.05) among SOL depths of post-fire trajectories within pre-fire dominance classes based on linear mixed effects models and Tukey’s post hoc analyses. We also tested for differences in SOL depth of the same post-fire trajectory among pre-fire classes but did not observe any statistical differences.
Discussion
Our analyses of an extensive dataset on pre-fire to post-fire composition shifts in boreal Alaska demonstrate the presence of deciduous trees in the pre-fire stand strongly impacts post-fire composition, generating strong hysteresis in the successional trajectories and dominant forest states across fire cycles (Figure 4). Stands with pre-fire deciduous fraction as low as 12% transitioned to deciduous dominance within the first 20 years post-fire. This is likely due to the presence of deciduous trees in the pre-fire stand providing a belowground bud bank for asexual regeneration that facilitates rapid and prolific post-fire colonization (Greene and Johnson 1999; Chen and others 2009). However, almost 30% of pure spruce stands (that is, no deciduous trees) still transitioned to deciduous dominance post-fire, highlighting the importance of deciduous seed availability and favorable seedbeds (that is, thin SOL) for the establishment of alternative deciduous trajectories (Johnstone and others 2010b; Brown and others 2015). With continued intensification of wildfire (Young and others 2017), spruce stands are likely to shift to alternative recovery trajectories leading to deciduous dominance, setting in motion the establishment of ecological legacies that will reinforce the resilience of deciduous forest states across multiple fire cycles (Figure 4).

A Fire severity acts as a driver of the initiation of alternative stable states in black spruce forests, through impacts on the depth of the soil organic layer (SOL; underlying dark gray profile). Spruce-dominated stands show high stability and resilience to fire when fire severity is low and a thick SOL persists after fire. When the SOL decreases to an average depth of ~ 10 cm by more severe fire, there is an increased likelihood that seedbeds will support deciduous tree recruitment from seed, creating opportunities for post-fire communities to shift from black spruce (blue) to mixed (green) or even deciduous (yellow) dominance at very high severities (Figure 3). Once deciduous trees are present in the prefire stand, the positive effects of pre-deciduous tree density and biomass on post-fire deciduous recruitment (Figure 2) lead to system hysteresis, where the alternative deciduous-dominated state persists under similar climate and fire regimes. B Step changes in forest composition occur in association with fire events when severe fire, coupled with available seed, allows deciduous trees to colonize. Mixed deciduous-spruce stands may represent an intermediate state that can return to spruce dominance with a long fire-free interval or shift to deciduous dominance with the historical fire return interval. A return from deciduous to spruce dominance is unlikely even if fire severity returns to low levels unless fire frequency also greatly declines. Illustrated by Victor Leshyk, Center for Ecosystem Science and Society, Northern Arizona University.
A key assumption of our findings is that patterns of post-fire regeneration reflect the composition of future boreal forest stands. Stand age reconstructions and longer-term observations of post-fire regeneration support this assumption. Specifically, early successional studies in northwestern boreal forests indicate the relative abundance of tree species recruiting during the initial 3–7 year period of post-disturbance regeneration is maintained through 2–3 decades of succession (Gutsell and Johnson 2002; Johnstone and others 2004; Shenoy and others 2011; Seidl and Turner 2022). This pattern holds even when changes in fire severity cause a shift from spruce to deciduous-dominated recovery (Johnstone and others 2020). Detailed reconstructions of tree ages further show that the trees that recruited immediately after fire dominate the canopy for at least a century of successional development (Johnson and others 1994; Gutsell and Johnson 2002). Relay succession is possible in mixed stands with a sufficiently long fire return interval, due to the shorter lifespan of aspen and birch compared to shade-tolerant conifers (Bergeron 2000; Taylor and Chen 2011) (Figure 4). However, succession in mixed canopy forests rarely leads to pure spruce even after 200 years in Interior Alaska (Fastie and others 2002; Kurkowski and others 2008), while the historic mean fire return interval is 70–120 years (Johnstone and others 2010a; Young and others 2017). Thus, forest composition initially established after fire in Interior Alaska likely persists until the stand burns again. Our results demonstrate that even if deciduous dominance declines to 12% of stand density and biomass (the metrics used in our analyses), this still provides enough of a budbank or seed source to ensure ample regeneration and subsequent dominance of deciduous trees after fire.
Post-fire observations (Johnstone and others 2010b, 2020; Baltzer and others 2021), experimental manipulations of burn depth (Johnstone and Chapin 2006), remote sensing of changes in deciduous fraction over time (Beck and others 2011; Massey and others In Revision), and modeling forecasts (Mekonnen and others 2019; Hansen and others 2020; Foster and others 2022a) provide support for the idea that deeper, more severe burning will lead to increasing abundance of deciduous stands on the landscape. Although all our pre-fire black spruce sites were established following the same year of fire our results align with these previous studies where fire severity drives compositional shifts from black spruce to deciduous dominance. Here, we found that even relatively low pre-fire deciduous presence results in post-fire deciduous dominance, suggesting that once alternative deciduous trajectories are in place, they will likely be resistant to reverting to conifer dominance in subsequent fire cycles. Specifically, the vegetative regeneration strategies of deciduous trees make them highly resilient to disturbance as they can rapidly resprout following fire (Greene and Johnson 1999; Chen and others 2009). Although black spruce is still likely to establish in high abundance following fire in black spruce forests (Johnstone and others 2020; Baltzer and others 2021; Mack and others 2021), if deciduous trees are present their rapid colonization via vegetative regeneration gives them a competitive advantage over the slow growing black spruce, further reinforcing the deciduous trajectory. The combination of (a) increases in fire conditions conducive to deciduous tree recruitment and (b) strong ecological legacies that support subsequent deciduous resilience is likely to cause directional change and hysteresis in forest landscape responses to changing climate and fire. Model simulations that incorporate even a conservative version of this hysteresis predict a substantial conversion of conifer to deciduous-dominated forests in Interior Alaska in the current century (Mann and others 2012; Foster and others 2022b).
Our finding that post-fire SOL was thinner in deciduous trajectories compared to spruce trajectories aligns with previous research on post-fire regeneration dynamics throughout the North American boreal forest (Johnstone and others 2010b, 2020; Baltzer and others 2021; Walker and others 2017). These studies highlight that high fire severity, measured as either total or proportional SOL combustion, can expose mineral soil and facilitate deciduous tree regeneration after wildfire. However, our measurement of post-fire SOL depth is not only driven by burn depth but is also a function of pre-fire SOL depth (Kane and others 2007; Johnstone and others 2020), which is affected by fire frequency and environmental conditions at the site. Deciduous forests are generally located in dry and warm landscape positions (Kurkowski and others 2008; Walker and others 2017) and therefore likely had thinner pre-fire SOL than spruce stands which are generally located at wet and cold landscape positions. In this study, we did not assess burn depth of the SOL and therefore could not reconstruct pre-fire SOL depth and total or proportional combustion. However, we found that post-fire SOL depth of deciduous trajectories did not differ among pre-fire tree compositional dominance classes, suggesting that post-fire SOL representative of seedbed conditions is a more important driver of deciduous recruitment and success than total or proportional combustion.
Historically, North American boreal forests have acted as a net C sink, accumulating C from the atmosphere over numerous fires cycles or centuries (Chapin and others 2006; Bond-Lamberty and others 2007; Kelly and others 2013). With the continued intensification of wildfires, these ecosystems could switch from net C sink to a net C source via deep burning of the SOL (Walker and others 2019). However, deep burning and the associated establishment of deciduous forests also impacts the boreal net ecosystem C balance and can result in fourfold net increase in C storage over the 100-year disturbance cycle (Mack and others 2021). Our results show that once deciduous trees become established, even at low densities in spruce stands, their reproductive strategies frequently enable them to dominate recovery after wildfire, thereby setting the stand on an alternative successional trajectory that is likely to persist across multiple fire cycles in boreal Alaska.
Moreover, although deciduous stands are generally considered resistant to burning (Parisien and others 2011; Girardin and others 2013; Rogers and others 2015), our findings suggest that when they do burn, their asexual regeneration strategies promote continued deciduous dominance. Lower resilience of spruce than deciduous stands to wildfire could leave spruce forests vulnerable to replacement by deciduous forests, especially as wildfires become more frequent and intense with warming over the coming decades (Foster and others 2022a). This switch to deciduous dominance not only alters the pattern of boreal forest C storage (Alexander and Mack 2016; Mack and others 2021), but could cool the atmosphere through higher albedo (Rogers and others 2013) and substantially decrease wildfire activity and risk (Parisien and others 2011; Girardin and Terrier 2015).
Critical next steps for understanding the persistence of deciduous boreal forests include assessing their resilience to other disturbances (for example, insect outbreaks and drought) and determining the drivers of fire occurrence and severity in these forests. As boreal forests store upwards of 30% terrestrial C, understanding these drivers is not only key to predicting the future structure and function of boreal forests but also for quantifying the role of fire in the global C cycle and its feedbacks to climate change.
Data Availability
All data used in this manuscript are archived with the Environmental Data Initiative (EDI) via the Bonanza Creek Long-term Ecological Research Program.http://dx.doi.org/10.6073/pasta/b78161335e3edbd2393378b197735d85.
References
Abatzoglou JT, Williams AP. 2016. Impact of anthropogenic climate change on wildfire across western US forests. Proceedings of the National Academy of Sciences 113:11770–11775.
Alexander HD, Mack MC. 2016. A Canopy Shift in Interior Alaskan Boreal Forests: Consequences for Above- and Belowground Carbon and Nitrogen Pools during Post-fire Succession. Ecosystems 19:98–114.
Baltzer JL, Day NJ, Walker XJ, Greene D, Mack MC, Alexander HD, et al. 2021. Increasing fire and the decline of fire adapted black spruce in the boreal forest. Proc Natl Acad Sci 118:e2024872118.
Beck PSA, Juday GP, Alix C, Barber VA, Winslow SE, Sousa EE, et al. 2011. Changes in forest productivity across Alaska consistent with biome shift. Ecology Letters 14:373–379.
Bergeron Y. 2000. Species and Stand Dynamics in the Mixed Woods of Quebec’s Southern Boreal Forest. Ecology 81:1500–1516.
Bond-Lamberty B, Peckham SD, Ahl DE, Gower ST. 2007. Fire as the dominant driver of central Canadian boreal forest carbon balance. Nature 450:89–92.
Brown CD, Liu J, Yan G, Johnstone JF. 2015. Disentangling legacy effects from environmental filters of postfire assembly of boreal tree assemblages. Ecology 96:3023–3032.
Chapin FSI, Woodwell GM, Randerson JT, Rastetter EB, Lovett GM, Baldocchi DD, et al. 2006. Reconciling carbon-cycle concepts, terminology, and methods. Ecosystems 9:1041–1050.
Chen HY, Vasiliauskas S, Kayahara GJ, Ilisson T. 2009. Wildfire promotes broadleaves and species mixture in boreal forest. Forest Ecology and Management 257:343–350.
Coop JD, Parks SA, Stevens-Rumann CS, Crausbay SD, Higuera PE, Hurteau MD, et al. 2020. Wildfire-driven forest conversion in western North American landscapes. BioScience 70:659–673.
Fastie, C.L., Lloyd, A.H. & Doak, P. (2002). Fire history and postfire forest development in an upland watershed of interior Alaska. Journal of Geophysical Research-Atmospheres, 108.
Foster AC, Armstrong AH, Shuman JK, Shugart HH, Rogers BM, Mack MC, et al. 2019. Importance of tree- and species-level interactions with wildfire, climate, and soils in interior Alaska: Implications for forest change under a warming climate. Ecological Modelling 409:108765.
Foster AC, Shuman JK, Rogers BM, Walker XJ, Mack MC, Bourgeau-Chavez LL, et al. 2022a. Bottom-up drivers of future fire regimes in western boreal North America. Environ. Res. Lett. 17:025006.
Foster AC, Wang JA, Frost GV, Davidson SJ, Hoy E, Turner KW, et al. 2022b. Disturbances in North American boreal forest and Arctic tundra: impacts, interactions, and responses. Environ. Res. Lett. 17:113001.
Frey BR, Lieffers VJ, Landhäusser SM, Comeau PG, Greenway KJ. 2003. An analysis of sucker regeneration of trembling aspen. Can. J. for. Res. 33:1169–1179.
Girardin MP, Ali AA, Carcaillet C, Blarquez O, Hély C, Terrier A, et al. 2013. Vegetation limits the impact of a warm climate on boreal wildfires. New Phytologist 199:1001–1011.
Girardin MP, Terrier A. 2015. Mitigating risks of future wildfires by management of the forest composition: an analysis of the offsetting potential through boreal Canada. Climatic Change 130:587–601.
Greene D, Johnson E. 1999. Modelling recruitment of Populus tremuloides, Pinus banksiana, and Picea mariana following fire in the mixedwood boreal forest. Canadian Journal of Forest Research 29:462–473.
Gutsell SL, Johnson EA. 2002. Accurately ageing trees and examining their height-growth rates: implications for interpreting forest dynamics. Journal of Ecology 90:153–166.
Hansen WD, Fitzsimmons R, Olnes J, Williams AP. 2020. An alternate vegetation type proves resilient and persists for decades following forest conversion in the North American boreal biome. Journal of Ecology 109(1):85.
Hayes K, Buma B. 2021. Effects of short-interval disturbances continue to accumulate, overwhelming variability in local resilience. Ecosphere 12:e03379.
Higuera PE, Brubaker LB, Anderson PM, Hu FS, Brown TA. 2009. Vegetation mediated the impacts of postglacial climate change on fire regimes in the south-central Brooks Range, Alaska. Ecological Monographs 79:201–219.
Hoecker TJ, Higuera PE, Kelly R, Hu FS. 2020. Arctic and boreal paleofire records reveal drivers of fire activity and departures from Holocene variability. Ecology 101:e03096.
Johnson EA, Miyanishi K, Kleb H. 1994. The Hazards of Interpretation of Static Age Structures as Shown by Stand Reconstructions in a Pinus Contorta – Picea Engelmannii Forest. The Journal of Ecology 82:923.
Johnstone J, Chapin F. 2006. Effects of soil burn severity on post-fire tree recruitment in boreal forest. Ecosystems 9:14–31.
Johnstone JF, Allen CD, Franklin JF, Frelich LE, Harvey BJ, Higuera PE, et al. 2016. Changing disturbance regimes, ecological memory, and forest resilience. Front Ecol Environ 14:369–378.
Johnstone JF, Celis G, Chapin FS III, Hollingsworth TN, Jean M, Mack MC. 2020. Factors shaping alternate successional trajectories in burned black spruce forests of Alaska. Ecosphere 11:e03129.
Johnstone JF, Chapin FS, Hollingsworth TN, Mack MC, Romanovsky V, Turetsky M. 2010a. Fire, climate change, and forest resilience in interior AlaskaThis article is one of a selection of papers from The Dynamics of Change in Alaska’s Boreal Forests: Resilience and Vulnerability in Response to Climate Warming. Can. J. for. Res. 40:1302–1312.
Johnstone JF, Chapin Iii F, Foote J, Kemmett S, Price K, Viereck L. 2004. Decadal observations of tree regeneration following fire in boreal forests. Canadian Journal of Forest Research 34:267–273.
Johnstone JF, Hollingsworth TN, Chapin FS, Mack MC. 2010b. Changes in fire regime break the legacy lock on successional trajectories in Alaskan boreal forest. Global Change Biology 16:1281–1295.
Kane ES, Kasischke ES, Valentine DW, Turetsky MR, McGuire AD. 2007. Topographic influences on wildfire consumption of soil organic carbon in interior Alaska: Implications for black carbon accumulation. J. Geophys. Res. 112:G03017.
Kelly R, Chipman ML, Higuera PE, Stefanova I, Brubaker LB, Hu FS. 2013. Recent burning of boreal forests exceeds fire regime limits of the past 10,000 years. Proceedings of the National Academy of Sciences 110:13055–13060.
Kukavskaya EA, Buryak LV, Shvetsov FOREXAMPLE, Conard SG, Kalenskaya OP. 2016. The impact of increasing fire frequency on forest transformations in southern Siberia. Forest Ecology and Management 382:225–235.
Kurkowski TA, Mann DH, Rupp TS, Verbyla DL. 2008. Relative importance of different secondary successional pathways in an Alaskan boreal forest. Canadian Journal of Forest Research 38:1911–1923.
Lenth, R., Singmann, H., Love, J., Buerkner, P. & Herve, M. (2019). emmeans: Estimated Marginal Means, aka Least-Squares Means.
Lloyd AH, Edwards ME, Finney BP, Lynch JA, Barber VA, Bigelow NH. 2006. Holocene development of the Alaskan boreal forest. In: Chapin I, Oswood M, Van Cleve K, Viereck LA, Verbyla DL, Eds. Alaska’s Changing Boreal Forest, . New York: Oxford University Press. pp 62–78.
Mack MC, Walker XJ, Johnstone JF, Alexander HD, Melvin AM, Jean M, et al. 2021. Carbon loss from boreal forest wildfires offset by increased dominance of deciduous trees. Science 372:280–283.
Mann DH, Rupp TS, Olson MA, Duffy PA. 2012. Is Alaska’s boreal forest now crossing a major ecological threshold? Arctic, Antarctic, and Alpine Research 44(3):319–331.
Massey, R., Rogers, B., Berner, L., Cooperdock, S., Mack, M., Walker, X., and others (In Revision). Changes in forest composition and associated biophysical climate feedbacks across boreal North America.
McDowell NG, Allen CD, Anderson-Teixeira K, Aukema BH, Bond-Lamberty B, Chini L, et al. 2020. Pervasive shifts in forest dynamics in a changing world. Science 368:eaaz9463.
Mekonnen ZA, Riley WJ, Randerson JT, Grant RF, Rogers BM. 2019. Expansion of high-latitude deciduous forests driven by interactions between climate warming and fire. Nat Plants 5:952–958.
Mollicone D, Eva HD, Achard F. 2006. Human role in Russian wild fires. Nature 440:436–437.
Neigh CSR, Nelson RF, Ranson KJ, Margolis HA, Montesano PM, Sun G, et al. 2013. Taking stock of circumboreal forest carbon with ground measurements, airborne and spaceborne LiDAR. Remote Sensing of Environment 137:274–287.
Osterkamp TE, Romanovsky VE. 1999. Evidence for warming and thawing of discontinuous permafrost in Alaska. Permafr Periglac Process 10:17–37.
Parisien M-A, Parks SA, Miller C, Krawchuk MA, Heathcott M, Moritz MA. 2011. Contributions of Ignitions, Fuels, and Weather to the Spatial Patterns of Burn Probability of a Boreal Landscape. Ecosystems 14:1141–1155.
Pinheiro, J., Bates, D., & R Core Team. (2022). Nlme: Linear and Nonlinear Mixed Effects Models.
R Development Core Team. (2018). R: A language and environment for statistical computing.
Ratajczak Z, Carpenter SR, Ives AR, Kucharik CJ, Ramiadantsoa T, Stegner MA, et al. 2018. Abrupt Change in Ecological Systems: Inference and Diagnosis. Trends in Ecology & Evolution 33:513–526.
Rogers BM, Randerson JT, Bonan GB. 2013. High-latitude cooling associated with landscape changes from North American boreal forest fires. Biogeosciences 10:699–718.
Rogers BM, Soja AJ, Goulden ML, Randerson JT. 2015. Influence of tree species on continental differences in boreal fires and climate feedbacks. Nature Geoscience 8:228–234.
Seidl R, Schelhaas M-J, Rammer W, Verkerk PJ. 2014. Increasing forest disturbances in Europe and their impact on carbon storage. Nature Climate Change 4:806–810.
Seidl R, Turner MG. 2022. Post-disturbance reorganization of forest ecosystems in a changing world. Proceedings of the National Academy of Sciences 119:e2202190119.
Shenoy A, Johnstone JF, Kasischke ES, Kielland K. 2011. Persistent effects of fire severity on early successional forests in interior Alaska. Forest Ecology and Management 261:381–390.
Simpson, G.L. (2022). Gratia: Graceful ggplot-Based Graphics and Other Functions for GAMs Fitted using mgcv.
Taylor AR, Chen HYH. 2011. Multiple successional pathways of boreal forest stands in central Canada. Ecography 34:208–219.
Van Cleve K, Chapin FS III, Dyrness CT, Viereck LA. 1991. Element cycling in Taiga forests state-factor control. Bioscience 41:78–88.
Walker XJ, Baltzer JL, Cumming SG, Day NJ, Ebert C, Goetz S, et al. 2019. Increasing wildfires threaten historic carbon sink of boreal forest soils. Nature 572:520–523.
Walker XJ, Mack MC, Johnstone JF. 2017. Predicting ecosystem resilience to fire from tree ring analysis in black spruce forests. Ecosystems 20:1137–1150.
Whitman E, Parisien M-A, Thompson DK, Flannigan MD. 2019. Short-interval wildfire and drought overwhelm boreal forest resilience. Sci Rep 9:18796.
Wickham H. 2016. ggplot2: Elegant Graphics for Data Analysis. New York: Springer-Verlag.
Wood SN. 2011. Fast stable restricted maximum likelihood and marginal likelihood estimation of semiparametric generalized linear models. Journal of the Royal Statistical Society: Series B (statistical Methodology) 73:3–36.
Wood SN. 2017. Generalized Additive Models: An Introduction with R, 2nd edn. New York: Chapman and Hall/CRC.
Yarie J, Billings S. 2002. Carbon balance of the taiga forest within Alaska: present and future. Canadian Journal of Forest Research 32:757–767.
Young AM, Higuera PE, Duffy PA, Hu FS. 2017. Climatic thresholds shape northern high-latitude fire regimes and imply vulnerability to future climate change. Ecography 40:606–617.
Yue X, Mickley LJ, Logan JA, Hudman RC, Martin MV, Yantosca RM. 2015. Impact of 2050 climate change on North American wildfire: consequences for ozone air quality. Atmospheric Chemistry and Physics 15:10033–10055.
Zuur, A., Ieno, E.N., Walker, N., Saveliev, A.A. & Smith, G.M. (2009). Mixed effects models and extensions in ecology with R. Springer Science & Business Media.
Acknowledgements
This project was supported by funding from the Department of Defense’s Strategic Environmental Research and Development Program (SERDP) under projects RC-2109 (MCM and JFJ) and RC18-1183 (SJG and MCM), NASA grant NNX15AT71A (MCM), Canada’s NSERC Discovery Grant program (JFJ), the US Joint Fire Sciences Program (05-1-2-06) (JFJ), and by the Bonanza Creek Long-term Ecological Research Program, which is supported by the US NSF (DEB-1636476) and the USDA Forest Service (RJVA-PNW-01-JV-11261952-231) (MCM and JFJ).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflicts of interest.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Walker, X.J., Okano, K., Berner, L.T. et al. Shifts in Ecological Legacies Support Hysteresis of Stand Type Conversions in Boreal Forests. Ecosystems 26, 1796–1805 (2023). https://doi.org/10.1007/s10021-023-00866-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-023-00866-w