
Abstract
In the early 1990’s, reserves adjacent to Kruger National Park (KNP) removed their fences to create a continuous landscape within the Kruger to Canyons Biosphere Reserve. Understanding how these interconnected multi-management systems responded to changes in environmental factors and management regimes can help to maintain natural large-scale landscape heterogeneity and ecological resilience. Our objective was to analyze remote sensing-derived vegetation metric changes between the different management types pre- and post-fence removal. The study area included fourteen reserves and the central section of KNP. We calculated the residuals between TIMESAT-derived metrics (from AVHRR NDVI time series) and rainfall to analyze changes in vegetation from 1985 to 2006. We then compared vegetation-rainfall residuals between different management types pre- and post-fence removal using mean–variance plots, nonmetric multidimensional scaling plots, and permutational multivariate analysis of variance to statistically identify and analyze changes. All management types experienced increased greenness. Reserves that removed their fences had greater changes in vegetation post-fence removal compared to reserves that remained fenced and KNP. Our findings suggest managers may need to address landscape changes by implementing management regimes such as reducing artificial surface water to counterbalance increased grazing pressure as a result of increased animal mobility across artificially created resource gradients. Habitat connectivity within and between protected area networks can be achieved by removing fences across adjacent conservation areas thus potentially increasing ecological resilience, which is vital to effective long-term conservation.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Highlights
-
Greenness increased in all protected areas in the Kruger to Canyons Biosphere Reserve.
-
Reserves that restored connectivity via fence removal had the greatest change in greenness.
-
Managers may need to counterbalance increased grazing pressure post-fence removal.
Introduction
Advancing large-scale ecological connectivity between core habitat areas is a major focus within the conservation community (CBD 2011; CBD 2020; Hilty and others 2020; Secretariat of the Convention on Biological Diversity 2020) and is a key component for rewilding ecosystems to regain their natural ecological processes (Soulé and Reed 1998; Carver and others 2021). Habitat connectivity conserves metapopulations by promoting organism dispersal, genetic variation, seasonal migration, and response to climate change through migration and refugia (Boone and Hobbs 2004; Hayward and Kerley 2009; Lindsey and others 2009; Smit and others 2020). Although global conservation area connectivity has increased slightly over the past decade (Saura and others 2019), international conservation efforts have not met the 2020 goal set by the Convention of Biological Diversity (Secretariat of the Convention on Biological Diversity 2020; Carrasco and others 2021). In many regions, conservation has shifted from individual protected areas to large-scale ecological networks for conservation (Fitzsimons and Wescott 2008; Santini and others 2016; Guzmán Wolfhard and Raedig 2019; Hilty and others 2020). Protected area networks can improve ecological connectivity by removing fences between adjacent conservation areas (Hayward and Kerley 2009; Lindsey and others 2009; Durant and others 2015), which has become an increasingly common practice in southern Africa (Newmark 2008; Peel and Smit 2020; Smit and others 2020). Conservation areas experiencing different management regimes will need to be connected to meet future conservation goals, including but not limited to Other Effective Area-based Conservation Measures (OECMs) (for example, Territories and Areas Conserved by Indigenous Peoples and Local Communities, Biodiversity Partnership Areas), lands managed under biodiversity stewardship arrangements, ecological corridors, private nature reserves, contractual parks, and national parks (Reid 2001; Ramutsindela 2003; Peel and others 2005; Kreuter and others 2010; Dudley and others 2018; Mitchell and others 2018; IUCN-WDPA 2019; Hilty and others 2020). Protected area network designs will directly influence their management (Margules and Pressey 2000), making successful systematic conservation planning vital to conserving these landscapes.
Although multiple types of conservation areas are integrated across protected area networks, understanding how different management regimes influence the landscape is important to provide effective conservation at large spatiotemporal scales. Historically, managers have used the equilibrium and non-equilibrium approaches to prevent degradation to land or vegetation, with these approaches having different underlying philosophies. In systems where managers subscribe to the equilibrium approach, the underlying premise is that plant biomass is controlled by herbivores, where grazing pressure, in combination with fire, maintains a relative constant amount of forage (Ellis and Swift 1988; Westoby and others 1989). In these equilibrium systems, managers place an emphasis on controlling herbivore numbers to maintain safe stocking rates below the ecological carrying capacity and retain a functional ecosystem during dry years, thereby reducing drought mortality (Ellis and Swift 1988; Vetter 2005). In contrast, a non-equilibrium management approach allows plant biomass to be controlled by fluctuating biotic and abiotic factors, such as rainfall, fire, herbivory, and predation, and assumes a system has discrete states of alternative persistent vegetation communities with transitions occurring between states (Ellis and Swift 1988; Westoby and others 1989; Stringham and others 2003). Managers subscribing to the non-equilibrium approach will allow herbivore populations to fluctuate with rainfall, with animal numbers increasing during wet years due to increased fecundity and decreasing due to high mortality during dry years (Ellis and Swift 1988; Vetter 2005). Mobility is key for non-equilibrium and large-scale systems, as it allows herbivores to move across the landscape in response to resource availability (Vetter 2005; Staver and others 2019; Smit and others 2020). When possible, rangeland management, especially in larger protected areas, has shifted toward the non-equilibrium approach to allow greater spatiotemporal flexibility, which in turn fosters landscape heterogeneity and ecological resilience (du Toit and others 2003; Cumming 2004; Vetter 2005).
Promoting landscape heterogeneity benefits biodiversity and ecological processes within conservation areas. Heterogeneous landscapes containing different vegetation types provide multiple niches that can be exploited by various organisms, providing the means to maintain high levels of biodiversity and ecological resilience (Lindenmayer and Fischer 2006; Fuhlendorf and others 2017). Herbivory in savannas influences ecosystem heterogeneity by modifying vegetation biomass (van Coller and Siebert 2015) and the structure and composition of woody and herbaceous vegetation (Scholes and Archer 1997; Levick and Rogers 2008; Hempson and others 2015). In order to maintain habitat diversity, surface water needs to be spaced far enough apart across the landscape to control overgrazing and homogenous grazing regimes (Smit and others 2007; Child and others 2013), which may be a factor that increases bush encroachment (Sinclair and Fryxell 1985), erosion risk, and starvation-induced mortality (Walker and others 1987) and may ultimately trigger an unfavorable alternative stable state (Scheffer and others 2001; Vetter 2005). Changes in ecosystem resilience can be interpreted using system states, which are considered resistant and resilient domains of ecological processes (for more information, see Supplementary Information 1).
In the early 1990’s, reserves adjacent to Kruger National Park (KNP) began to remove their fences to create a continuous landscape known as the “Greater Kruger Ecosystem” (Peel and others 2005). The Kruger to Canyons Biosphere Reserve (K2C) protected areas are managed across a spectrum of the equilibrium and non-equilibrium approaches, with fenced reserves aiming to contain herbivore numbers at or below ecological carrying capacity, reserves open to KNP embracing animal mobility over larger scales, and KNP acting as a larger natural ecosystem with the goal of maintaining natural biodiversity and heterogeneity (Peel and others 1998; du Toit and others 2003). Although the reserves adjacent to KNP aim to incorporate the national park’s management goals, they operate at smaller spatial extents and encounter various management challenges due to their size and varied management regimes (Peel and others 1998, 2005). Our goal was to determine whether fence removal between the reserves and KNP influenced the vegetation within the K2C protected area network. To answer this question, we analyzed how vegetation changed between 1985 and 2006 across three broad management types: “fenced” reserves remained individually fenced parcels throughout the whole study period, “recently unfenced” reserves removed the fences between them and the larger KNP ecosystem around 1993, and “KNP” as the central portion of Kruger National Park, which is open to the much larger KNP ecosystem. Our objective was to analyze changes in remote sensing-derived vegetation metrics of greenness and heterogeneity in greenness between the different management types. We predicted the recently unfenced reserves would show marked changes from pre- to post-fence removal, while fenced reserves and KNP would experience no or more limited change over the study period.
Methods
Study Area
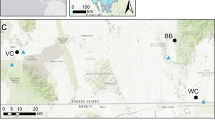
The study area was located within the Kruger to Canyons Biosphere Reserve (K2C) in the eastern Lowveld of South Africa between 30°35′ E and 30°40′ E and 24°00′ S and 25°00′ S (Peel and others 2007) (Figure 1). This included fourteen reserves and the central section of Kruger National Park (KNP), defined as the area between the Sabie and Olifants rivers. These managed protected areas fell within the savanna biome (Peel and others 2007) and cover an area of about 6,100 km2. The average annual rainfall varied over the study area, ranging from about 400 mm to 650 mm with the rainfall increasing from north to south (Peel and others 2007). Rainfall occurred primarily between the months of October and April (Venter and others 2003). The geology within the reserves was predominantly granitic rock, with some sections of amphibolitic and gabbro rock. The western side of KNP contained the same types of geology as the adjacent private and provincial reserves (granitic, amphibolitic, and gabbro), while the eastern side of KNP contained high-nutrient geology such as basaltic, rhyolitic, sandstone, and shale (Keyser 1997; Venter and others 2003). As such, we restricted our KNP study site to the western part with similar geology to the adjacent protected areas. Of the fourteen reserves, seven removed their fences within the period 1985 to 2006 therefore becoming open and connected to the Greater Kruger Ecosystem, while seven reserves remained fenced (Figure 1). Although both KNP and the other protected areas are conservation areas, the objectives and hence management approaches often differ, with the private protected areas often more intensely managed (for example, higher density of artificial water provision; clearing of woody vegetation in order to increase tourism viewing and photographic opportunities; and smaller management fires) with a strong focus on high-end tourism, while KNP focuses primarily on biodiversity conservation (Child and others 2013).

The area analyzed within the Kruger to Canyons Biosphere Reserve, including fenced reserves, recently unfenced reserves, and the central section of Kruger National Park containing nutrient poor geology.
Data
We utilized a historic 1-km2 normalized difference vegetation index (NDVI) dataset covering South Africa between 1985 and 2006, which incorporates National Oceanic and Atmospheric Administration (NOAA) Advanced Very High Resolution Radiometer (AVHRR) imagery for the years 1985–1998 and Satellite Pour L'Observation de la Terre VEGETATION (SPOT VGT) for the years 1998–2006 (Swinnen and Veroustraete 2008). Although integrated sensors can make it difficult to know whether detected changes result from ecological shifts or switching sensors, this was required in order to obtain a time series that spanned pre- and post-fence removal. Data from 1994 were unavailable due to failure of the NOAA-13 satellite, leading to a year-long gap within the dataset. Swinnen and Veroustraete (2008) provide details on the processing and integration of the NDVI dataset.
We used TIMESAT (Jönsson and Eklundh 2002, 2004) to fit a smooth continuous curve for the AVHRR NDVI time series dataset. We applied the adaptive Savitsky-Golay filter with the season cutoff value set to one and used the second spike method (STL replace) with one envelope iteration and a spatial window size of three. A higher threshold percentage is required when defining the end of the growing season in TIMESAT (Olsen and others 2015), which led us to define the end of the growing season as 45% of the seasonal amplitude as measured from the right minima, and the beginning of the growing season as 10% of the seasonal amplitude as measured from the left minima. We explored trends in two TIMESAT variables that measure vegetation productivity: maximum NDVI value (Max NDVI) and seasonal amplitude (Jönsson and Eklundh 2002, 2004). Although the inability to distinguish greenness as herbaceous or woody vegetation restricted our interpretation abilities, Max NDVI and seasonal amplitude provided robust metrics that maintained spatiotemporal consistency, which allowed comparison across many reserves and over a long period of time.
Since savanna systems are heavily responsive to rainfall (Scholes and Archer 1997) and we were interested in vegetation changes from management regimes instead of rainfall differences over time, we calculated the residuals between rainfall and the vegetation metrics for each year as a measure of vegetation changes resulting from non-rainfall causes. We used a historic 1-km2 interpolated rainfall dataset covering South Africa, depicting average total rainfall for a 12-month period beginning in July and ending in June of the next year (Malherbe and others 2016; Agricultural Research Council Institute for Soil, Climate, and Water). We resampled the 1-km2 rainfall data to align with the NDVI dataset and extracted values for rainfall, Max NDVI, and seasonal amplitude for each 1 km2 pixel (ESRI 2014). We then used linear models to calculate the vegetation-rainfall residuals between the log-transformed rainfall and each vegetation metrics (Max NDVI and seasonal amplitude) and summarized by management type and time period (see Supplementary Information 2 for statistical assumption confirmations).
We compared vegetation pre- and post-fence removal by splitting the time series data into distinct time periods. Since most reserves in the study area started removing fences in 1993 (Supplementary Information 3), the TIMESAT metrics extracted between 1985 and 1993 corresponded to pre-fence removal, and the metrics extracted between 1995 and 2006 corresponded to post-fence removal. Previous studies found it took time for animals to adjust to newly connected reserves (Druce and others 2008), and that 5 years was roughly the amount of time it took for the vegetation to fully respond to the removal of a fence (Hiscocks 1999; de Boer and others 2015), leading us to split the data into three time periods: (1) “Early” defined as the metrics extracted between 1985 and 1993 (pre-fence removal), (2) “Transition” defined as the metrics extracted between 1995 and 1998 (up to 5 years after fence removal), and (3) “Late” defined as the metrics extracted between 1999 and 2006 (more than 5 years after fence removal). The metrics were also summarized by protected area management type: (1) “fenced” reserves retained their fences through the entire 1985–2006 time period, (2) “recently unfenced” reserves removed their fences around 1993, and (3) “KNP” represented the central section of Kruger National Park falling between the Sabie and Olifants rivers. In order to keep the comparison between KNP and the reserves consistent, we only analyzed the vegetation that fell on sections of granitic, amphibolitic, and gabbro rock within the central sections of KNP, as geology is an important driver of vegetation patterns (Keyser 1997; Venter and others 2003). Further, we acknowledge the unavoidable spatial autocorrelation of recently unfenced reserves being adjacent to KNP and fenced reserves being farther from KNP, which is an artifact of using restored connectivity across a protected area network as a natural experiment to better understand the influence of large-scale fence removal. We calculated the mean and variance of the vegetation-rainfall residuals for each management type per year as a proxy for vegetation greenness and greenness heterogeneity.
Assessing Changes in Vegetation
We performed a mean–variance analysis in order to identify and measure changes over time and across different management types. Washington-Allen and others (2008) demonstrated that mean–variance plots can be used in semi-arid systems to analyze the ecological resilience of a landscape by splitting the landscape trajectory into four quadrants. Increases along the x-axis depict increased greenness (as a measure of vegetation biomass and vigor), and increases along the y-axis represent increased heterogeneity of greenness. This type of analysis allowed us to measure changes in two continuous variables (mean and variance), as well as to characterize landscape resilience within the four quadrants (Figure 2). Because we calculated the mean and variance of the vegetation-rainfall residuals for Max NDVI and seasonal amplitude, we assumed movement toward different quadrats within the mean–variance plots indicated changes not influenced by rainfall.

Hypothetical statistical quadrants of the interannual mean–variance dynamics of a savanna landscape’s vegetation-rainfall residuals (revised from Washington-Allen and others 2008).
We used k-means clustering to further analyze the locations of clusters within the mean–variance plots. We used the NbClust package (Charrad and others 2014) to determine the optimal number of clusters within each mean–variance plot, and this value was used to perform the k-means clustering analysis in RStudio (RStudio Team 2016; R Core Team 2020).
We performed a permutational multivariate analysis of variance (PERMANOVA) to detect whether the Euclidean distances significantly differed between clusters in the mean–variance plots. We used the vegan package (Oksanen and others 2019) to determine that Max NDVI-rainfall residuals and seasonal amplitude-rainfall residuals met the assumption of equal dispersion between clusters (Supplementary Information 2). We then used the smacof (de Leeuw and Mair 2009) and tidyverse (Wickham and others 2019) packages to visualize how the clusters differed across coordinate space using nonmetric multidimensional scaling (NMDS) plots and tested for significant differences between these clusters using the vegan (Oksanen and others 2019) and RVAideMemoire (Hervé 2020) packages.
We explored vegetation changes by assessing how the remote sensing metrics changed across the mean–variance defined quadrats. Once Max NDVI-rainfall residuals and seasonal amplitude-rainfall residuals met statistical assumptions (Supplementary Information 2), we performed a Tukey Honest Significant Differences (HSD) post hoc test to analyze comparisons between all management type levels and time period levels.
Results
Changes in Vegetation
The mean–variance plots of vegetation greenness and heterogeneity of greenness and the k-means cluster analysis identified four and five distinct clusters within fenced reserves for Max NDVI-rainfall residuals and seasonal amplitude-rainfall residuals, respectively, five distinct clusters within recently unfenced reserves for both Max NDVI-rainfall residuals and seasonal amplitude-rainfall residuals, and five and three distinct clusters within KNP for Max NDVI-rainfall residuals and seasonal amplitude-rainfall residuals, respectively (Figure 3). Across all management types, a general pattern of temporal change along the mean axis, with Early time period on the left and Late time period on the right, indicated increased greenness after accounting for rainfall from 1985 to 2006 throughout the whole landscape (Figure 3). An exception to this general pattern was in 1998, a severe drought year, which had low mean and variance for both vegetation metrics for fenced and recently unfenced reserves. Since 1998 was an exception to the pattern, we believe the system experienced a “reversible change” to increased greenness. With two or more distinct clusters identified visually for each management type, we proceeded with statistical analysis to test for significant differences between the clusters.

Mean–variance plots of the Max NDVI-rainfall residuals (left: a, b, c) and seasonal amplitude-rainfall residuals (right: d, e, f) for fenced reserves (a, d), recently unfenced reserves (b, e), and KNP (c, f).
Overall, the cluster centroids significantly differed between time periods for the Max NDVI-rainfall residuals (F = 16.41, P < 0.001) and seasonal amplitude-rainfall residuals (F = 14.07, P < 0.001) (Table 1). The centroids also significantly differed between the management types for the Max NDVI-rainfall residuals (F = 3.34, P = 0.041) (Table 1). See Supplementary Information 4 for full PERMANOVA statistics on the main level effects for Max NDVI and seasonal amplitude.
While Figure 3 visually illustrates a shift to higher greenness from the start to the end of the study period for all management types, pairwise comparisons confirmed a statistically significant shift in the centroids within the NMDS plots for all reserve types including fenced (Max NDVI: F = 8.68, P = 0.015; seasonal amplitude: F = 8.10, P = 0.014), recently unfenced (Max NDVI: F = 11.63, P = 0.009; seasonal amplitude: F = 12.90, P = 0.003), and KNP (Max NDVI: F = 5.89, P = 0.032; seasonal amplitude: F = 6.05, P = 0.035) (Table 2). Although the pairwise comparisons demonstrated that the centroids differed between the early and late periods across all management types for both Max NDVI-rainfall residuals and seasonal amplitude-rainfall residuals (Table 2), the amount of overlap between the early and late polygons in the NMDS plots suggest this difference may have been more pronounced in recently unfenced reserves compared to the fenced reserves and KNP (Figure 4). This indicates that although the whole protected area network seemed to experience a large-scale change during the study period, KNP and fenced reserves may have shifted less than the recently unfenced reserves.

Nonmetric Multidimensional Scaling (NMDS) plots comparing Max NDVI-rainfall residuals (left: a, b, c) and seasonal amplitude-rainfall residuals (right: d, e, f) between time periods within fenced reserves (a, d), recently unfenced reserves (b, e), and KNP (c, f).
Vegetation-rainfall residuals in recently unfenced reserves were similar to fenced reserves at the start of the study and became more similar to KNP during the transition and late time periods (Figure 5). The Tukey HSD analysis revealed that Max NDVI-rainfall residuals and seasonal amplitude-rainfall residuals were significantly higher at the end of the study compared to the start of the study (P < 0.001 in both cases) (Table 3). KNP also had significantly higher Max NDVI-rainfall residuals compared to fenced reserves (P = 0.04) (Table 3). In addition to the significant differences for pairwise comparisons for the main effects, there were also several significant differences for pairwise comparisons within the Management Type:Time Period interaction term, although the interaction term itself was not significant (Table 1). We cautiously interpreted these pairwise comparisons as follows. Within recently unfenced reserves, both Max NDVI-rainfall residuals and seasonal amplitude-rainfall residuals increased during the study period (P = 0.03 in both cases) (Table 3, Figure 5). Recently unfenced reserves also had significantly higher Max NDVI-rainfall residuals and seasonal amplitude-rainfall residuals at the end of the study compared to fenced reserves at the start of the study (P = 0.02 in both cases) (Table 3). Additionally, KNP also had significantly higher Max NDVI-rainfall residuals and seasonal amplitude-rainfall residuals at the end of the study than fenced reserves (Max NDVI: P < 0.001; seasonal amplitude: P = 0.01) and recently unfenced reserves (Max NDVI: P < 0.001; seasonal amplitude: P = 0.02) at the start of the study (Table 3).

Mean Max NDVI-rainfall residuals (a) and mean seasonal amplitude-rainfall residuals (b) for Early, Transition, and Late time periods, by management type.
Discussion
Understanding how vegetation responds to changes in management (for example, the removal of fencing or artificial provision of water) can help guide landscape-scale management to maintain or enhance ecological resilience and spatial conservation planning. In a first of its kind study of large-scale fence removal within a protected area network, we found relatively small changes in medium resolution-derived greenness and heterogeneity within an overall trend of increasing greenness. Our results suggest that recently unfenced reserves experienced greater changes in vegetation from the start to the end of the study compared to the fenced reserves and KNP and those vegetation changes may be associated with their connectivity to the larger KNP landscape post-fence removal. Our findings suggest all management types within the K2C protected area network experienced a “reversible change” with a gradual increase in greenness, although the system did not cross a threshold into a new state since the fenced, recently unfenced, and KNP systems all reversed during the drought in 1998. We postulate recently unfenced reserves experienced at least three changes that may have interacted in unknown ways: (1) the gradual increase in greenness the whole system encountered, (2) a sudden increase in animal mobility after the reserves became connected to KNP, and (3) a gradual shift in management strategies to account for increased connectedness with the surrounding landscape.
Vegetation greenness gradually increased across the entire K2C protected area network during the study period regardless of management type or rainfall levels, representing a large-scale change. While inconsistent with our prediction, this finding is supported by other studies that showed global increases in greenness (Nemani and others 2003; Zhu and others 2016) as well as increased greenness regardless of rainfall in African landscapes (Fensholt and others 2012; Gibbes and others 2014). Although we cannot be sure what process drove this pattern in our study system, a possible ecological explanation is increased woody encroachment resulting from heightened CO2 levels, which is a documented phenomenon across the globe (Stevens and others 2016). Woody encroachment may influence ecosystems by impacting species richness (Blaum and others 2007; Sirami and others 2009; Ratajczak and others 2012), decreasing grazing species in turn shifting predator guilds, and changing fire intervals and intensity (Smit and Prins 2015), which may require management intervention to maintain biodiversity and resilience.
Increased animal mobility within the recently unfenced reserves may have caused an influx of water-dependent species due to the higher waterpoint densities in the recently unfenced reserves compared to KNP (Child and others 2013). For example, de Boer and others (2015) showed a 16-fold increase in elephant density in a recently unfenced reserve and attributed this to a combination of population growth and movement from KNP. Artificial waterpoints are known to have a strong influence on herbivore distribution and can result in unnaturally high pressure on the surrounding vegetation (Owen-Smith 1996; Chamaillé-Jammes and others 2007; Smit and others 2007). Graz and others (2012) modeled the joint management actions of fence removal with waterpoint closure and predicted that removing fences while retaining high levels of waterpoints would spread grazing activity across the landscape (thereby expanding grazing impact), whereas removing fences while closing some waterpoints resulted in the two management changes counterbalancing one-another, with some areas experiencing an increase in herbivory (analogous to recently unfenced reserves in our study) while other areas experienced a decrease in herbivory (analogous to KNP) (Graz and others 2012). Our empirical findings align with the Graz and others (2012) models, suggesting that reserve managers may want to consider reducing the number or density of artificial waterpoints simultaneously with fence removal to prevent sudden animal influx and reduction in vegetation heterogeneity (Cook and others 2017). Constructive engagement and dialog between private reserves and KNP about the strong gradient in artificial waterpoints, changes in animal mobility, and conservation objectives may be needed to continue the mutual benefits of collaborative conservation.
Along with a gradual increase in greenness and sudden increase in animal mobility, recently unfenced reserves may alter management regimes to account for increased connectedness with the surrounding landscape. Increased mobility may have shifted management strategies from focusing on maintaining animal numbers under the carrying capacity (that is, equilibrium system) to embrace the natural and spatial heterogeneity of the savanna landscape (that is, non-equilibrium system) (Peel and others 1998). Increased mobility may also motivate bush clearing to increase viewing opportunities of highly desired species. Given diverse management goals among reserves, vegetation changes stemming from fence removal may have positive and negative effects that may be enhanced or exacerbated by management actions. In the future, reserves planning to remove fences may want to consider adjusting other aspects of the system such as limiting wide-scale artificial water provision to offset a sudden increase in animal mobility (Graz and others 2012; Cook and others 2017), as well as coordinating management and monitoring (for example, fire regimes) across neighboring conservation areas to enable the larger landscape to be managed cooperatively between multiple managers (Lindsey and others 2009) and maintain heterogeneity across the entire connected landscape (van Wilgen and others 2022).
Connecting conservation areas managed under different objectives directly aligns with Goal A in the post-2020 Global Biodiversity Framework to increase the “area, connectivity, and integrity of natural ecosystems” (CBD 2020), making the K2C protected area network a vital landscape for learning how increased protected area connectivity may alter ecosystems. Along with its innovative large-scale fence removal, the K2C region could be considered an important case study, providing valuable lessons for incorporating flexibility while managing its complex socio-ecological system (du Toit and others 2003; Holness and Biggs 2011; Roux and Foxcroft 2011; van Wilgen and Biggs 2011; Biggs and others 2015). In order to maintain heterogeneity and ecological resilience within protected savannas effectively, managers need to incorporate the complexity of spatiotemporal uncertainties and fluctuations (Scheffer and others 2001; Rogers 2003; Folke and others 2004; Fuhlendorf and others 2017), which include biotic and abiotic changes that may be gradual or sudden (Ratajczak and others 2018). Future management practices will need to cooperatively conserve the world’s remaining ecosystems which are experiencing multiple threats, often have diverse management goals, and are managed at various spatial scales. Taking this large-scale ecosystem recovery approach is the best chance to globally rewilding ecological networks by integrating core habitat connectivity, the dynamic changing nature of ecosystems, and co-existence through local community conservation (Carver and others 2021).
Conclusion
Our study examined vegetation change, by proxy of moderate resolution remote sensing metrics, between 1985 and 2006 in a South African protected area network to determine if landscape change differed between management types, and whether fence removal across adjacent protected areas influenced the vegetation. This is the first study we are aware of to analyze landscape changes across a protected area network before and after large-scale fence removal, which can provide insight for global protected area networks rewilding ecosystems to improve ecological resilience, restore ecological processes, and adapt to climate change (Carver and others 2021). Although the area and number of parks have grown extensively over the past century in southern Africa, the average park size decreased while fencing increased and protected areas became fragmented “ecological islands” (Cumming 2004). Most reserve boundaries were created without considering large-scale natural environmental processes such as seasonal movements and migration patterns (Cumming 2004), causing serious fragmentation problems within fenced ecosystems (Boone and Hobbs 2004; Lindsey and others 2012). Although connecting different reserves can at least partly resolve some of these issues (Lindsey and others 2009), new challenges arise when attempting to manage various conservation objectives through multiple land owners and stakeholders at a large scale. Our findings suggest managers may need to address landscape changes by counterbalancing management regimes such as adapting artificial surface water provision when increasing animal mobility through fence removal. It is crucial to better understand how these interconnected systems respond to gradual and sudden changes in environmental factors and management regimes to maintain natural landscape heterogeneity and enhance ecological resilience.
Data Availability
Data and code are publicly available on GitHub: https://github.com/ellielinden/K2C_VegetationChange.
References
Biggs R, Rhode C, Archibald S, Kunene LM, Mutanga SS, Nkuna N, Ocholla PO, Phadima LJ. 2015. Strategies for managing complex social-ecological systems in the face of uncertainty: examples from South Africa and beyond. Ecol Soc 20:52.
Blaum N, Rossmanith E, Popp A, Jeltsch F. 2007. Shrub encroachment affects mammalian carnivore abundance and species richness in semiarid rangelands. Acta Oecol 31:86–92.
Boone RB, Hobbs NT. 2004. Lines around fragments: effects of fencing on large herbivores. Afr J Range Forage Sci 21:147–158.
Carrasco L, Papeş M, Sheldon KS, Giam X. 2021. Global progress in incorporating climate adaptation into land protection for biodiversity since Aichi targets. Glob Chang Biol:1788–801.
Carver S, Convery I, Hawkins S, Beyers R, Eagle A, Kun Z, Van Maanen E, Cao Y, Fisher M, Edwards SR, Nelson C, Gann GD, Shurter S, Aguilar K, Andrade A, Ripple B, Davis J, Sinclair A, Bekoff M, Noss R, Foreman D, Pettersson H, Root-Bernstein M, Svenning J-C, Taylor P, Sophie W-J, Featherstone, Alan Watson Fløjgaard C, Stanley-Price M, Navarro LM, Aykroyd T, Parfitt A, Soulé M. 2021. Guiding principles for rewilding. Conserv Biol:1–31.
Chamaillé-Jammes S, Fritz H, Murindagomo F. 2007. Climate-driven fluctuations in surface-water availability and the buffering role of artificial pumping in an African savanna: potential implication for herbivore dynamics. Austral Ecol 32:740–748.
Charrad M, Ghazzali N, Boiteau V, Niknafs A. 2014. NbClust: an R package for determining the relevant number of clusters in a data set. J Stat Softw 61:1–36. http://www.jstatsoft.org/v61/i06/.
Child MF, Peel MJS, Smit IPJ, Sutherland WJ. 2013. Quantifying the effects of diverse private protected area management systems on ecosystem properties in a savannah biome, South Africa. Oryx 47:29–40. http://search.ebscohost.com/login.aspx?direct=true&db=8gh&AN=84657751&site=ehost-live.
Convention on Biological Diversity (CBD). 2011. Report of the tenth meeting of the conference of the parties to the convention on biological diversity. Montreal, Canada www.cbd.int/doc/meetings/cop/cop-10/ official/cop-10–27-en.pdf.
Convention on Biological Diversity (CBD). 2020. Zero draft of the post-2020 global biodiversity framework.
Cook RM, Witkowski ETF, Helm CV, Henley MD, Parrini F. 2017. Recent exposure to African elephants after a century of exclusion: rapid accumulation of marula tree impact and mortality, and poor regeneration. For Ecol Manag 401:107–116. https://doi.org/10.1016/j.foreco.2017.07.006.
Cumming DHM. 2004. Performance of parks in a century of change. In: Child B, Ed. Parks in transition: biodiversity, rural development and the bottom line, . London: Earthscan. pp 105–124.
de Leeuw J, Mair P. 2009. Multidimensional scaling using majorization: SMACOF in R. J Stat Softw 31:1–30. http://www.jstatsoft.org/v31/i03/.
de Boer WF, van Oort JWA, Grover M, Peel MJS. 2015. Elephant-mediated habitat modifications and changes in herbivore species assemblages in Sabi Sand, South Africa. Eur J Wildl Res 61:491–503.
Druce HC, Pretorius K, Slotow R. 2008. The response of an elephant population to conservation area expansion: phinda private game reserve, South Africa. Biol Conserv 141:3127–3138.
du Toit JT, Rogers KH, Biggs HC. 2003. The kruger experience: ecology and management of savanna heterogeneity. New York: Island Press.
Dudley N, Jonas H, Nelson F, Parrish J, Pyhälä A, Stolton S, Watson JEM. 2018. The essential role of other effective area-based conservation measures in achieving big bold conservation targets. Glob Ecol Conserv 15:1–7.
Durant SM, Becker MS, Creel S, Bashir S, Dickman AJ, Beudels-Jamar RC, Lichtenfeld L, Hilborn R, Wall J, Wittemyer G, Badamjav L, Blake S, Boitani L, Breitenmoser C, Broekhuis F, Christianson D, Cozzi G, Davenport TRB, Deutsch J, Devillers P, Dollar L, Dolrenry S, Douglas-Hamilton I, Dröge E, FitzHerbert E, Foley C, Hazzah L, Hopcraft JGC, Ikanda D, Jacobson A, Joubert D, Kelly MJ, Milanzi J, Mitchell N, M’Soka J, Msuha M, Mweetwa T, Nyahongo J, Rosenblatt E, Schuette P, Sillero-Zubiri C, Sinclair ARE, Price MRS, Zimmermann A, Pettorelli N. 2015. Developing fencing policies for dryland ecosystems. J Appl Ecol 52:544–51. http://doi.wiley.com/https://doi.org/10.1111/1365-2664.12415
Ellis JE, Swift DM. 1988. Stability of African pastoral ecosystems: alternate paradigms and implications for development. J Range Manag 41:450.
ESRI. 2014. ArcMap (version 10.6) software.
Fensholt R, Langanke T, Rasmussen K, Reenberg A, Prince SD, Tucker CJ, Scholes RJ, Le QB, Bondeau AlberteEastman J, Eastman JR, Epstein H, Gaughan AE, Hellden U, Mbow C, Olsson L, Paruelo JM, Schweitzer C, Seaquist JW, Wessels KJ. 2012. Greenness in semi-arid areas across the globe 1981–2007—an earth observing satellite based analysis of trends and drivers. Remote Sens Environ 121:144–158. https://doi.org/10.1016/j.rse.2012.01.017.
Fitzsimons JA, Wescott G. 2008. The role of multi-tenure reserve networks in improving reserve design and connectivity. Landsc Urban Plan 85:163–173.
Folke C, Carpenter S, Walker B, Scheffer M, Elmqvist T, Gunderson L, Holling CS. 2004. Regime shifts, resilience, and biodiversity in ecosystem management. Annu Rev Ecol Evol Syst 35:557–581.
Fuhlendorf SD, Fynn RWS, McGranahan DA, Twidwell D. 2017. Heterogeneity as the basis for rangeland management. In: Briske DD, Ed. Rangeland systems: processes, management and challenges. Springer. pp 169–96.
Gibbes C, Southworth J, Waylen P, Child B. 2014. Climate variability as a dominant driver of post-disturbance savanna dynamics. Appl Geogr 53:389–401.
Graz FP, Westbrooke ME, Florentine SK. 2012. Modelling the effects of water-point closure and fencing removal: a GIS approach. J Environ Manag 104:186–194. https://doi.org/10.1016/j.jenvman.2012.03.014.
Guzmán Wolfhard LV, Raedig C. 2019. Connectivity conservation management: linking private protected areas. In: Nehren U, Schlϋter S, Raedig C, Sattler D, Hissa H, Eds. Strategies and tools for a sustainable rural Dio de Janeiro. Switzerland (AG): Springer. pp 155–71.
Hayward MW, Kerley GIH. 2009. Fencing for conservation: restriction of evolutionary potential or a riposte to threatening processes? Biol Conserv 142:1–13.
Hempson GP, Archibald S, Bond WJ, Ellis RP, Grant CC, Kruger FJ, Kruger LM, Moxley C, Owen-Smith N, Peel MJS, Smit IPJ, Vickers KJ. 2015. Ecology of grazing lawns in Africa. Biol Rev 90:979–94. https://doi.org/10.1111/brv.12145.
Hervé M. 2020. RVAideMemoire: testing and plotting procedures for biostatistics. https://cran.r-project.org/package=RVAideMemoire.
Hilty J, Worboys G, Keeley A, Woodley S, Lausche B, Locke H, Carr M, Pulsford I, Pittock J, White W, Theobald D, Levine J, Reuling M, Watson J, Ament R, Tabor G. 2020. Guidance for conserving connectivity through ecological networks and corridors. Gland, Switzerland: IUCN. https://portals.iucn.org/library/node/49061.
Hiscocks K. 1999. The impact of an increasing elephant population on the woody vegetation in southern Sabi Sand Wildtuin, South Africa. Koedoe 42:47–55.
Holness SD, Biggs HC. 2011. Systematic conservation planning and adaptive management. Koedoe 53:1–9.
IUCN-WDPA. 2019. Recognising and reporting other effective area-based conservation measures. Switzerland.
Jönsson P, Eklundh L. 2002. Seasonality extraction by function-fitting to time-series of satellite sensor data. IEEE Trans Geosci Remote Sens 40:1824–1832.
Jönsson P, Eklundh L. 2004. TIMESAT—a program for analyzing time-series of satellite sensor data. Comput Geosci 30:833–845.
Keyser N. 1997. Geological map of the republic of South Africa and the kingdoms of Lesotho and Swaziland.
Kreuter U, Peel MJS, Warner E. 2010. Wildlife conservation and community-based natural resource management in Southern Africa’s private nature reserves. Soc Nat Resour An Int J 23:507–24. http://www.informaworld.com/https://doi.org/10.1080/08941920903204299.
Levick SR, Rogers KH. 2008. Patch and species specific responses of savanna woody vegetation to browser exclusion. Biol Conserv 141:489–498.
Lindenmayer DB, Fischer J. 2006. Habitat fragmentation and landscape change: an ecological and conservation synthesis. Washington: Island Press.
Lindsey PA, Romañach SS, Davies-Mostert HT. 2009. The importance of conservancies for enhancing the value of game ranch land for large mammal conservation in southern Africa. J Zool 277:99–105.
Lindsey PA, Masterson CL, Beck AL, Romañach SS. 2012. Ecological, Social and financial issues related to fencing as a conservation tool in Africa. In: Somers MJ, Hayward MW, Eds. Fencing for conservation: restriction of evolutionary potential or a riposte to threatening processes? pp 215–34.
Malherbe J, Dieppois B, Maluleke P, Van Staden M, Pillay DL. 2016. South African droughts and decadal variability. Nat Hazards 80:657–681.
Margules CR, Pressey RL. 2000. A framework for systematic conservation planning. Nature 405:243–53. www.nature.com
Mitchell BA, Fitzsimons JA, Stevens CMD, Wright DR. 2018. PPA or OECM? Differentiating between privately protected areas and other effected area-based conservation measures on private land. Parks 24:49–60.
Nemani RR, Keeling CD, Hashimoto H, Jolly WM, Piper SC, Tucker CJ, Myneni RB, Running SW. 2003. Climate-driven increases in global terrestrial net primary production from 1982 to 1999. Science 300(80):1560–1563.
Newmark WD. 2008. Isolation of African protected areas. Front Ecol Environ 6:321–328.
Oksanen J, Blanchet, F. Guillaume Friendly M, Kindt R, Legendre P, McGlinn D, Minchin PR, O’Hara RB, Simpson GL, Solymos P, Stevens MHH, Szoecs E, Wagner H. 2019. vegan: community ecology package. https://cran.r-project.org/package=vegan.
Olsen JL, Miehe S, Ceccato P, Fensholt R. 2015. Does EO NDVI seasonal metrics capture variations in species composition and biomass due to grazing in semi-arid grassland savannas? Biogeosciences 12:4407–4419.
Owen-Smith N. 1996. Ecological guidelines for waterpoints in extensive protected areas. S Afr J Wildl Res 26:107–112.
Peel MJS, Smit IP. 2020. Drought amnesia: lessons from protected areas in the eastern Lowveld of South. African. J Range Forage Sci 37:81–92.
Peel MJS, Biggs HC, Zacharias PJK. 1998. The evolving use of stocking rate indices currently based on animal number and type in semi-arid heterogeneous landscapes and complex land-use systems. African J Range Forage Sci 15:117–127.
Peel MJS, Kruger JM, Zacharias PJK. 2005. Environmental and management determinants of vegetation state on protected areas in the eastern Lowveld of South Africa. Afr J Ecol 43:352–361.
Peel MJS, Kruger JM, Macfadyen S. 2007. Woody vegetation of a mosaic of protected areas adjacent to the Kruger National Park, South Africa. J Veg Sci 18:807–814.
Ramutsindela M. 2003. Land reform in South Africa’s national parks: a catalyst for the human-nature nexus. Land Use Policy 20:41–49.
Ratajczak Z, Nippert JB, Collins SL. 2012. Woody encroachment decreases diversity across North American grasslands and savannas. Ecology 93:697–703.
Ratajczak Z, Carpenter SR, Ives AR, Kucharik CJ, Ramiadantsoa T, Stegner MA, Williams JW, Zhang J, Turner MG. 2018. Abrupt change in ecological systems: inference and diagnosis. Trends Ecol Evol 33:513–526. https://doi.org/10.1016/j.tree.2018.04.013.
Reid H. 2001. Contractual national parks and the Makuleke community. Hum Ecol 29:135–155.
Rogers KH. 2003. Adopting a heterogeneity paradigm: implications for management of protected savannas. In: du Toit JT, Rogers KH, Biggs HC, Eds. The kruger experience: ecology and management of savanna heterogeneity, . New York: Island Press. pp 41–58.
Roux DJ, Foxcroft LC. 2011. The development and application of strategic adaptive management within South African National Parks. Koedoe 53:1–5.
RStudio Team. 2016. RStudio: integrated development for R.
Santini L, Saura S, Rondinini C. 2016. Connectivity of the global network of protected areas. Divers Distrib 22:199–211.
Saura S, Bertzky B, Bastin L, Battistella L, Mandrici A, Dubois G. 2019. Global trends in protected area connectivity from 2010 to 2018. Biol Conserv 238:108183. https://doi.org/10.1016/j.biocon.2019.07.028.
Scheffer M, Carpenter SR, Foley JA, Folke C, Walker B. 2001. Catastrophic shifts in ecosystems. Nature 413:591–6. http://www.nature.com/doifinder/https://doi.org/10.1038/35098000.
Scholes RJ, Archer S. 1997. Tree-grass interactions in savannas. Annu Rev Ecol Syst 28:517–544.
Secretariat of the Convention on Biological Diversity. 2020. Global biodiversity outlook 5. Montreal
Sinclair ARE, Fryxell JM. 1985. The Sahel of Africa: ecology of a disaster. Can J Zool 63:987–994.
Sirami C, Seymour C, Midgley G, Barnard P. 2009. The impact of shrub encroachment on savanna bird diversity from local to regional scale. Divers Distrib 15:948–957.
Smit IPJ, Prins HHT. 2015. Predicting the effects of woody encroachment on mammal communities, grazing biomass and fire frequency in African savannas. PLoS One 10:1–16. https://doi.org/10.1371/journal.pone.0137857.
Smit IPJ, Grant CC, Devereux BJ. 2007. Do artificial waterholes influence the way herbivores use the landscape? Herbivore distribution patterns around rivers and artificial surface water sources in a large African savanna park. Biol Conserv 136:85–99.
Smit IPJ, Peel MJS, Ferreira SM, Greaver C, Pienaar DJ. 2020. Megaherbivore response to droughts under different management regimes: lessons from a large African savanna. African J Range Forage Sci 37:65–80.
Soulé M, Reed N. 1998. Rewilding and biodiversity: complementary goals for continental conservation. Wild Earth 8:19–28.
Staver AC, Wigley-Coetsee C, Botha J. 2019. Grazer movements exacerbate grass declines during drought in an African savanna. J Ecol 107:1482–1491.
Stevens N, Erasmus BFN, Archibald S, Bond WJ. 2016. Woody encroachment over 70 years in South African savannas: overgrazing, global change or extinction aftershock? Philos Trans R Soc B Biol Sci 371:1–9.
Stringham TK, Krueger WC, Shaver PL. 2003. State and transition modeling: an ecological process approach. J Range Manag 56:106–113.
Swinnen E, Veroustraete F. 2008. Extending the SPOT-VEGETATION NDVI time series (1998–2006) back in time with NOAA-AVHRR data (1985–1998) for Southern Africa. IEEE Trans Geosci Remote Sens 46:558–572.
Team RC. 2020. R: a language and environment for statistical computing. https://www.r-project.org/.
van Coller H, Siebert F. 2015. Herbaceous biomass—species diversity relationships in nutrient hotspots of a semi-arid African riparian ecosystem. Afr J Range Forage Sci 32:213–223.
van Wilgen BW, Biggs HC. 2011. A critical assessment of adaptive ecosystem management in a large savanna protected area in South Africa. Biol Conserv 144:1179–1187. https://doi.org/10.1016/j.biocon.2010.05.006.
Venter FJ, Scholes RJ, Eckhardt HC. 2003. The abiotic template and its associated vegetation pattern. In: du Toit JT, Rogers KH, Biggs HC, Eds. The kruger experience: ecology and management of savanna heterogeneity, . New York: Island Press. pp 83–129.
Vetter S. 2005. Rangelands at equilibrium and non-equilibrium: recent developments in the debate. J Arid Environ 62:321–341.
Washington-Allen RA, Ramsey RD, West NE, Norton BE. 2008. Quantification of the ecological resilience of drylands using digital remote sensing. Ecol Soc 13.
Westoby M, Walker B, Noy-Meir I. 1989. Opportunistic management for rangelands not at equilibrium. J Range Manag 42:266–274.
Wickham H, Averick M, Bryan J, Chang W, McGowan L, François R, Grolemund G, Hayes A, Henry L, Hester J, Kuhn M, Pedersen T, Miller E, Bache S, Müller K, Ooms J, Robinson D, Seidel D, Spinu V, Takahashi K, Vaughan D, Wilke C, Woo K, Yutani H. 2019. Welcome to the tidyverse. J Open Source Softw 4:1686.
Zhu Z, Piao S, Myneni RB, Huang M, Zeng Z, Canadell JG, Ciais P, Sitch S, Friedlingstein P, Arneth A, Cao C, Cheng L, Kato E, Koven C, Li Y, Lian X, Liu Y, Liu R, Mao J, Pan Y, Peng S, Peuelas J, Poulter B, Pugh TAM, Stocker BD, Viovy N, Wang X, Wang Y, Xiao Z, Yang H, Zaehle S, Zeng N. 2016. Greening of the earth and its drivers. Nat Clim Chang 6:791–795.
Acknowledgements
We would like to thank Else Swinnen and Johan Malherbe for providing the historic AVHRR NDVI and historic rainfall datasets used in this analysis. We would also like to thank all reserve managers and SANParks staff for taking the time to discuss the complexities of the K2C system. We thank Mike Gill, David Eldridge, and two anonymous reviewers for their reviews of earlier drafts, which significantly improved this manuscript. The University of Connecticut Department of Natural Resources and the Environment provided the funding to travel to South Africa and gain invaluable insights on the K2C system.
Funding
No funding was received for conducting this study.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Author contributions: EL, CR, IS, MP, IO designed the study; EL performed the analysis; EL and CR wrote the manuscript, which was revised by IS and MP.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Linden, E., Rittenhouse, C.D., Peel, M.J.S. et al. Vegetation Changes Following Large-scale Fence Removal Across a Protected Area Network Within the Kruger to Canyons Biosphere Reserve, South Africa. Ecosystems 26, 768–783 (2023). https://doi.org/10.1007/s10021-022-00792-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-022-00792-3