
Abstract
Atmospheric deposition can be an important source of phosphorus (P) to watersheds in geologically low-P landscapes like the Precambrian Shield, in south-central Ontario. Recent observations in this region of declining total P (TP) concentrations in streams and lakes, as well as P limitation in vegetation, prompted this investigation into the potential contributing role of atmospheric deposition. Long-term (1980–2014) measurements of bulk TP deposition at four monitoring stations, as well as wet-only deposition at one of the sites (2012–2014) and pollen measurements in 2014, were used to assess the potential contribution of biogenic sources to TP deposition in the region. Ratios of phosphorus to potassium (P:K) in historical bulk deposition and variability in P deposition among the four sites indicated that continuously open, bulk deposition collectors are enriched in ‘local’ biogenic sources of P, particularly in the summer, fall and spring months. Furthermore, in 2014 pollen could account for as much as 30% of bulk TP deposition, and pine trees (Pinus strobus) were the most important source of pollen (>75% of collected pollen). In contrast, winter bulk deposition was the least affected by local, biogenic sources, and likely best reflects net P inputs to this region from long-range (that is, external to the watershed) sources. Catchment input–output budgets for P that use measured annual bulk deposition totals as inputs are generally strongly positive (indicating net retention/accumulation of P within watersheds), whereas annual budgets calculated using extrapolated winter deposition are in approximate balance. Balanced P budgets are more consistent with regional observations of declining surface water TP and P limitation in terrestrial vegetation. Importantly, there was no significant decline in winter TP deposition over the period of record (1980–2014), which suggests that changes in regional deposition are not responsible for declines in surface water TP. Although we recommend extrapolating winter bulk deposition measurements to approximate annual atmospheric TP input to terrestrial ecosystems in non-dusty, oligotrophic landscapes such as this, local sources of P, particularly pollen, are likely important seasonal sources of P nutrition to lakes.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Total phosphorus (TP) concentrations unexpectedly declined over the past several decades at many streams and lakes in the geologically low-P Precambrian Shield landscape of south-central Ontario, despite increases in human development within their catchments (Yan and others 2008; Eimers and others 2009; Palmer and others 2011). Determining the cause of these declines is a high priority given the critical role of P in determining algal production and community composition (Paterson and others 2008) as well as the potential impacts of ‘oligotrophication’ on higher trophic levels (Jeppesen and others 2005). Furthermore, chemical analysis of sugar maple (Acer saccharum) foliage and correlations between soil P concentrations and tree growth in this region suggest that P is transitioning to become the limiting nutrient, as a result of historically high atmospheric nitrogen (N) deposition (Gradowski and Thomas 2006, 2008; Casson and others 2012).
Despite observations of declining surface water TP and suggestions of P limitation in forests, P input–output budgets for Precambrian Shield catchments are generally strongly positive (that is, bulk deposition exceeds stream export; Dillon and Molot 1997; Eimers and others 2009). Positive input–output budgets suggest that P is accumulating within the landscape. The exception is watersheds that have been harvested or undergone substantial wetland disturbance, which show net P loss (Phillips and Watmough 2012; Pinder and others 2014). These observations underscore the need for a reassessment of watershed P budgets and further scrutiny of the accuracy of estimates of atmospheric deposition.
A number of studies have demonstrated the importance of atmospheric deposition as a source of P to watersheds in oligotrophic landscapes where sources of P are otherwise low (for example, Vicars and others 2010), which includes much of the Precambrian Shield region of eastern North America (Dillon and Kirchner 1975). Schindler and others (1976), for example, estimated that atmospheric deposition supplied approximately half of the annual P budget of oligotrophic Rawson Lake in the Experimental Lakes Area of northwestern Ontario, and Cole and others (1990) estimated that bulk deposition in the summer months exceeded fluvial input over the same time period by 50–60 times, at oligotrophic Mirror Lake in New Hampshire. However, the accurate measurement of atmospheric P deposition is notoriously difficult, and numerous studies have reported contamination of deposition collections with local sources of P. ‘Bulk’ deposition collectors, which are continuously open to the atmosphere, are particularly prone to contamination from local dust and P-enriched biological material including pollen, insect parts and bird excrement (Newman 1995; Ahn 1999).
Of primary concern with these potential sources of P is their origin. Sources that originate from within the watershed should be removed from terrestrial or catchment-scale budget calculations, whereas sources that originate from outside of the watershed are considered ‘true inputs’ and should be included in input–output budgets (Ahn and James 2001). Lake budgets on the other hand should include bulk deposition as well as inflows from the terrestrial catchment. Despite their shortcomings, bulk collectors are a simple, cost-effective means of estimating total atmospheric deposition to watersheds. They are the primary source of deposition data for many publicly funded long-term monitoring programs, including the Ontario Ministry of Environment and Climate Change (OMECC) Dorset Environmental Science Centre’s (DESC) monitoring program, which has measured TP inputs and outputs at several Precambrian Shield watersheds since the mid 1970s (Scheider and others 1979).
Pollen is a particularly important source of local contamination in bulk collectors as it contains high concentrations of P and large amounts of tree pollen are released from trees in short, intense bursts every spring and summer (Lee and others 1996b). Deposition of Pinus pollen, for example, may contribute 2.4–6.4 mg P m−2 to annual deposition (Doskey and Ugoagwu 1989; Lee and others 1996b; Lee and Booth 2003; Cho and others 2003). Dust particles also may be enriched in P, although P contamination from dust is likely less important in geologically low P, oligotrophic environments like the Precambrian Shield (Mahowald and others 2008). Fossil fuel and biomass burning are other important sources of atmospheric P deposition in ‘non-dusty’ regions (for example, Tsukuda and others 2006), although these sources are more often considered ‘external’ (rather than internal contamination), particularly in regions that are remote from point sources (Mahowald and others 2008). As fossil fuel emissions and atmospheric deposition of sulfur (S) and nitrogen (N) compounds have declined in Ontario over the past several decades (Kothawala and others 2011), P deposition from these sources may have also declined and contributed to surface water TP trends.
Given the importance of atmospheric P deposition and the considerable uncertainty surrounding its measurement and application, the objectives of this study were to: (1) quantify the contribution of local/internal sources (that is, pollen) to earlier bulk TP deposition measurements and (2) determine whether changes in atmospheric deposition could have contributed to observed declines in surface water TP in this region.
Methods
Study Area
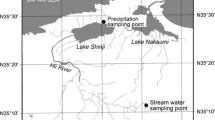
The study region (Figure 1) is located at the southern edge of the Precambrian Shield in the Great Lakes-St. Lawrence ecozone, where forest cover is mixed temperate with Pinus strobus (white pine), Pinus resinosa (red pine) and Tsuga canadensis (eastern hemlock) in the coniferous areas and Betula allegheniensis (yellow birch), Fagus grandifolia (American beech) and sugar maple in the deciduous areas. Surficial geology is characterized by shallow till–rock ridge topography, with bedrock dominated by slowly weathered igneous and metamorphic rock. As such, the contribution of P to lakes via weathering is slow (Kirkwood and Nesbitt 1991; Baker and others 2015). Previous mass budget estimates have shown that atmospheric deposition can represent more than 60% of the total TP input to lakes with relatively small catchment areas relative to lake surface area, like Plastic Lake (PCP; Dillon and Molot 1996). The climate of the region is humid continental with long-term (1980/81–2013/14) mean annual precipitation of 1100 mm year−1 with approximately 35% falling as snow and a mean annual temperature of 5.1 °C. There is virtually no agriculture in this region due to the thin, nutrient poor soils and rocky terrain.

Map of the study area with locations of the four deposition monitoring stations.
The Dorset Environmental Science Centre (DESC) has monitored meteorology and atmospheric bulk deposition in this region since the late 1970s. The Ontario Ministry of Environment and Climate Change (OMECC) first established the DESC calibrated watersheds in order to evaluate the impact of cottage development on nutrient delivery to Precambrian Shield lakes, and several headwater watersheds were selected to encompass a gradient of human disturbance. At the low end of the disturbance spectrum, Plastic Lake (PCP), is surrounded by crown land with no cottage development; Heney Lake (HYP) has a low level of development with only 9 shoreline residences, whereas Harp (HPP) and Paint (PTP) represent more developed lakes with 90 and more than 150 shoreline dwellings, respectively. A meteorological and bulk deposition monitoring station was established at PTP in 1976, located within the backyard of the DESC research station property directly across a paved road from Paint Lake (Figure 1). Similar meteorological and bulk deposition monitoring stations were established within the PCP, HPP and HYP watersheds in the early 1980s (records presented here begin in 1985).
Bulk Deposition Collectors
Field and laboratory methods for measuring bulk deposition at the four sites have been relatively consistent since monitoring began, although the total number of deposition and meteorological monitoring stations has varied from a minimum of three to as many as eight (Dillon and others 1988). Based on early measurements that demonstrated little variability among stations, bulk deposition measurements from at least three stations have been historically pooled to provide a single ‘regional’ estimate for the area (Scheider and others 1979). Since 1985, this has been done by averaging deposition measurements at PCP, HYP and HPP to provide a ‘3-station’ regional estimate, with values from the PTP site used to substitute for one of the other stations during equipment malfunctions or when data were unusable. Deposition monitoring at the Harp Lake watershed ceased in 2013 in response to a landowner request, and therefore records at this site end in 2012/13.
Bulk deposition is collected using continuously open, lined plastic buckets at cleared, open areas at each sampling site. Bulk collectors are located 1.7 m above the ground and are topped with a funnel to limit evaporation. A roll of Nitex screen (10 cm × 10 cm; 500 µm mesh) is placed in the opening of each funnel to limit entry of coarse particles and insects (Scheider and others 1979). Bulk collectors are visited typically weekly, at which time deposition samples are removed and buckets are refitted with clean, food-grade plastic bag liners and Nitex screens. Bulk samples are coarse-filtered (80 µm) and transported to the laboratory in insulated coolers.
The landscape surrounding the deposition collector can influence local sources of P deposition. At Paint Lake (PTP), an open area of approximately 35 m × 35 m surrounds the long-term bulk collector, dominated by Poa pretensis (Kentucky blue grass), which is mowed during summer. This open area is surrounded on three sides by trees dominated by Populus tremuloides (trembling aspen), Betula papyrifera (white birch) and Fraxinus americana (white ash), whereas the OMECC DESC building and a gravel parking lot are located in the other direction (~115 m away). Roads are located in three directions beyond the forest, including a gravel road 60 m away, a secondary paved road for homes and cottages 260 m away, and a two-lane highway (Hwy 117) 440 m away.
The forest surrounding the Plastic Lake site (PCP) is dominated by Pinus strobus, and to a lesser extent Quercus rubra and Acer rubrum (red maple). The open area around the bulk deposition collector is 27, 29, 54, and 70 m in each direction to the tree line and is dominated by Rubus allegheniensis (blackberry), Rubus canadensis, Rubus idaeus var. strigosus (red raspberry), juvenile Acer rubrum, Pteridium aquilinum, Solidago rugosa, Plantago major (plantain), Symphyotrichum lanceolatum var. lanceolatum (panicle aster) and Muhlenbergia frondosa (grass). Shrubs and juvenile trees surrounding the deposition monitoring equipment are cut every few years in order to maintain clearance and limit shading of meteorological sensors, and this was last done in 2010 (C. McConnell OMECC, pers. comm.). The Plastic Lake area has little traffic, and has one, low-use seasonal gravel road, 190 m away from the bulk collector. A two-lane highway (Hwy 35) is located 2.3 km away.
The Heney Lake (HYP) site is located at the edge of a mixed hardwood forest, and Graminoids dominate the open area (20, 25, 26, and 86 m in each direction to the tree line) surrounding the bulk deposition collector. The forest is dominated by Pinus strobus (Eastern white pine), Quercus rubra (Northern red oak), and Populus tremuloides. This site has relatively low traffic and is located at the end of a secondary paved road that leads to a year-round residence. A gravel road continues beyond the meteorological station with little seasonal cottage traffic. The gravel section of the road is 100 m from the bulk deposition collector, the secondary paved section is 1.8 km away, and the two-lane highway (Hwy 117) is 2.5 km away.
Vegetation surrounding the historical HPP monitoring site could not be surveyed in detail like the other monitoring stations. However, like HYP, the forest at Harp Lake is predominantly hardwood, with Acer saccharum (sugar maple), Acer rubrum and Fagus grandifolia (American beech) dominating the uneven-aged canopy (Watmough and Dillon 2003a, b). A paved cottage-access road is located within 200 m of the retired HPP monitoring station.
Wet-Only Deposition Monitoring
Wet-only deposition has been collected at PTP since January 2012 using an ADS 00-120 (N-CON Systems Co. Inc.) automated wet-only precipitation collector, installed 2 m above the ground approximately 30 m away from the bulk collector. Wet-only deposition is only monitored at PTP due to the availability of electrical power at this site. Similar to bulk deposition, wet-only collectors are visited at least weekly, and samples are collected as soon as possible after major precipitation events. All deposition samples are coarse-filtered (80 µm) prior to chemical analysis.
Chemical Analysis of Deposition
Chemical analyses of long-term (1980 onwards) collections of bulk deposition as well as more recent measurements of wet-only deposition (2012 onwards) were conducted at the DESC laboratory. Total P in deposition is measured via auto analyzer using ascorbic acid–molybdate colorimetry after autoclave digestion in sulfuric acid. Sulfate (SO4) and nitrate (NO3–N) concentrations are analyzed by ion chromatography, and potassium (K) samples are acidified and then analyzed using atomic absorption spectroscopy. Annual (hydrologic year: June 1–May 31), seasonal (summer = Jun/Jul/Aug; fall = Sep/Oct/Nov; winter = Dec/Jan/Feb; spring = Mar/Apr/May) and monthly fluxes (mg m−2) are estimated by summing daily fluxes over the specified time period. Daily fluxes are estimated by multiplying the measured chemical concentration on a particular collection date by the amount of precipitation that fell every day since the previous collection date. Concentrations are volume-weighted by dividing the annual or monthly flux by the depth of water deposited over the same time period.
Pollen Deposition in 2014
Durham (1946) pollen collectors were deployed at PTP, PCP and HYP during the 2014 growing season. Each collector consists of two horizontal disks (23 cm in diameter, 15 cm apart), with the lower disk located approximately 1.8 m from the ground (height of bulk collector). A standard 25 mm × 75 mm glass microscope slide covered with a thin film of petroleum jelly to collect pollen was mounted approximately 2 cm above the lower disk using a wooden clothes peg. Five collectors were installed at each station, in a ring around each bulk deposition collector (15 total). From April 24 to August 4 2014 (duration of pollen ‘rain’), microscope slides were collected and replaced with new slides once or twice per week, depending on pollen saturation. Cover slips were placed on each slide for transport to Trent University, and slides were stored at 4 °C prior to enumeration. It should be noted that Durham pollen collectors can have up to 20% undercatch because of the upper disk (Lee and Booth, 2003).
Pollen grains were counted, measured (length, width and depth) and identified to species using a microscope (40–100× magnification) within a randomly selected 1 cm2 area on the collection slide to allow extrapolation of areal deposition (Cho and others 2003). Pollen species were identified following Bassett and others (1978). The average dimensions of a Pinus pollen grain were 46 μm × 69 μm × 49 μm (n = 20), and pollen grain volume was estimated using the formula 4/3πabc, where a = half length, b = half width, and c = half depth of a single pollen grain. Pollen grain weight (1.9 × 10−8 g grain−1) was estimated from the measured mass of 5 mL of air-dried pollen (assuming 50% of the volume was voids; Cho and others 2003) and the calculated volume of an individual pine pollen grain (8.1 × 104 μm3).
To determine whether pollen grains were captured in bulk and wet-only deposition collectors, the amount and type of pollen found in collectors was qualitatively assessed in several grab collections between May 12 and June 24, 2014. In total, pollen grains were counted and all tree pollen was identified to genus level in five wet-only, and eight bulk collections at PTP, six bulk collections at PCP and seven bulk collections at HYP. All collections were first coarse-filtered (80 µm) prior to pollen counting. Immediately before counting, deposition collections were agitated to suspend pollen and then a subsample of known volume was placed in a glass petri dish and pollen grains were identified and counted using a microscope. Pollen counts in subsamples were then extrapolated to the total volume of collected precipitation.
Pollen Macronutrient Analysis
Pollen grains from Pinus strobus and Pinus resinosa cones were harvested by gently shaking cones (still attached to the tree) into paper bags. Pollen was harvested from Pinus trees surrounding PTP, PCP and HYP and composited in order to produce sufficient mass for analysis. Pollen collections (n = 5) were air-dried, weighed (0.2 g per sample for analysis) and block digested in nitric acid for 8 h at 100 °C, and pollen digests were analyzed for P and K by inductively coupled plasma optical emission spectrometry (PerkinElmer ICP-OES). All pollen analyses were conducted at the Environmental Geoscience Research Centre at Trent University. Recovery of P and K was assessed through the analysis of SRM 1515 Apple Leaves and was 90–110%.
Statistical Analysis
Annual, seasonal and monthly bulk deposition (mg m−2) and volume-weighted concentrations (µg L−1) of P and K were calculated between 1980/81 and 2014/15 (PTP), 1985/86 and 2014/15 (PCP and HYP) or 1985/86 and 2012/13 (HPP). Correlations between TP and K have been used to indicate biological sources of P in stream water in this region (Pinder and others 2014), and correlations between TP and K in deposition were explored as a way of assessing the contribution of biological sources of P (that is, plant and animal material) to TP deposition (Mahowald and others 2008). Correlations between P and K concentrations were examined through Pearson’s correlation analysis using raw data from the 30+ year study period at each of the four stations. Differences in chemical concentrations between bulk and wet-only collections of deposition at PTP were evaluated through paired t tests and were considered significant at p < 0.05. Phosphorus and K deposition trends over time were evaluated using the seasonal Mann–Kendall test (Skjelkvale and others 2005; Hamed and Rao 1998) with monthly measurements of P and K deposition. To calculate the contribution of pollen to atmospheric deposition of P and K, the weight of an individual pollen grain was multiplied by the number of pollen grains deposited per unit of area. This weight was then multiplied by the macronutrient concentration of pollen.
Results
Seasonal Patterns and Trends Across Stations
There were no trends in seasonal or annual bulk TP deposition at any of the four long-term monitoring stations (Mann–Kendall test; p > 0.05), and instead, the predominant pattern in the historical record was one of substantial intra-annual, among-year and between-site variability (Figure 2). Seasonal differences in bulk TP deposition were clear at all four stations, with TP inputs consistently highest and most variable in the spring (March–May) and summer (June–August), and lowest and least variable in the winter months (December–February; Figure 2). Although this seasonal pattern was apparent at all four sites, there was substantial variability in seasonal totals across stations (Table 1). Notably, deposition was consistently highest and most variable at Paint Lake in all seasons (see ranges in % CV in Table 1). Differences in seasonal deposition loads among stations were largest in the summer (range 4.0–9.8 mg m−2) and smallest in the winter (range 1.0–1.7 mg m−2). These differences were due to variability in TP concentrations across stations, rather than seasonal precipitation totals, which were very consistent across sites (Table 1).

Seasonal TP deposition loads (mg m−2) at the four monitoring stations. Summer = Jun/Jul/Aug; fall = Sep/Oct/Nov; winter = Dec/Jan/Feb; spring = Mar/Apr/May. Note the different scale of the spring deposition plot.
Phosphorus Versus Potassium Correlations
Total P and K concentrations were significantly positively correlated (p < 0.001) when all raw measurements were considered at each individual station; however, correlations were particularly strong in the summer and spring and were weakest in the winter and fall (Table 2).
Wet-Only Deposition
Concentrations of TP in bulk deposition at PTP were significantly higher than in wet-only deposition (paired t tests; p < 0.05), and differences were largest in the summer (16.5×), fall (5.5×) and spring (2.9×) and smallest in the winter (1.6×; Table 3). Concentrations of K were similarly enriched in bulk deposition collections and were significantly higher than wet-only concentrations in the summer (12×), fall (4×) and spring (1.6×), whereas differences were smallest and statistically insignificant in the winter (1.4×; Table 3). Surprisingly, TP and K concentrations were equally strongly correlated in wet-only deposition (2012–2014) as in bulk deposition at PTP (see Table 2) in the summer (r = 0.68), fall (r = 0.71) and spring (r = 0.99), although unlike the long-term record, there was no correlation between TP and K in winter wet-only deposition (r = −0.009). Strong K:P correlations in wet-only deposition during the growing season suggest that wet-only collectors are vulnerable to contamination from local biogenic sources of P, and high pollen grain counts in grab samples (that is, non-quantitative) of wet-only (and bulk) deposition collections clearly illustrate the vulnerability of wet-only collectors to contamination from pollen (Table 4). For example, 698 pollen grains were counted in just five grab samples of wet-only deposition at PTP, compared with 1794 pollen grains in eight bulk deposition samples at the same site (Table 4).
In contrast to TP and K, concentrations of SO4 and NO3–N, which are commonly associated with the long-range transport of fossil fuel combustion products, were only modestly enriched in bulk compared with wet deposition (10–28% and 3–21%, respectively), and SO4 and NO3–N concentrations were much less variable both within and between seasons (Table 3).
Pollen Deposition
Pollen deposition was quantified at PTP, PCP and HYP during 2014 in order to evaluate the contribution of this biological source to P and K loads at each site. Pollen deposition was greatest at PCP, followed by HYP and PTP, and pine pollen (Pinus strobus and Pinus resinosa) accounted for the majority (>75%) of tree pollen collected at each site (Table 5). Pinus pollen deposition occurred almost entirely in the month of June (97% of total pine pollen collected), whereas pollen from deciduous tree species was collected primarily in May (68–97% of total deciduous pollen collected; temporal data not shown).
The contribution of pine pollen to P and K deposition in 2014 was estimated at each site using measured P and K concentrations in pollen digests and pine pollen deposition rates. Phosphorus and K concentrations in pine pollen were high, and averaged 3.2 ± 0.1 mg g−1and 14.3 ± 0.7 mg g−1, respectively, with a K:P ratio of 4.5:1. When these macronutrient concentrations were applied to pollen deposition rates, pine pollen deposition in the month of June accounted for as much as 30% of annual (2014) bulk TP deposition at PCP, 22% at HYP and 7% at PTP (Table 6). Similar contributions of Pinus pollen to K deposition were estimated for 2014 (Table 6).
The clear enrichment of spring, summer and to a lesser extent fall bulk deposition with biogenic P, which is assumed to originate from within each catchment, suggests that winter-only deposition could be used to approximate the contribution of long-range, external P deposition in this region. Applying the four-station historical winter TP deposition rate (1.4 mg m−2 winter−1; see Table 1) year-round suggests a much lower annual deposition input to this region (5.7 mg m−2 year−1) than previously published estimates (that is, 16–18 mg m−2 year−1; Dillon and Molot 1997; Eimers and others 2009). Recalculation of annual input–output budgets using the lower estimate of deposition further suggests that budgets are much less positive than previous reports and/or are close to being in balance with stream export (Table 7). It should be noted that although we use the calendar definition of winter (December, January, February) in this study, average bulk TP deposition concentrations are very similar in these 3 months (data not shown), suggesting that in this case, the calendar definition of winter matches well with the season of low biotic TP production.
Discussion
Sources of Temporal and Spatial Variability in Deposition
Results of this study have shown that bulk TP deposition is highly spatially and temporally variable, with maximum deposition occurring in the spring, summer and fall months. Strong seasonality in P deposition has been noted in previous studies in temperate regions, with maximum P concentrations and loads typically occurring in the growing season, whereas deposition loads are generally lower in the dormant (that is, winter) months (for example, Brown and others 2011). Higher levels of P deposition in the growing season are generally attributed to greater biological activity in the warm months and associated production of P-rich biological materials including pollen and other plant debris, insect parts and bird droppings (Dammgen and others 2005; Tsukuda and others 2006).
Correlations between TP and K in bulk deposition, which are particularly strong in the spring and summer, support the contribution of biological sources of P deposition in this region. Furthermore, pollen appears to be a particularly important source, as counts in deposition collectors and targeted collections of pollen indicated that 30% of annual bulk TP deposition could be attributed to pollen falling just in the month of June. In temperate forests, pollen is released from plants in large quantities every spring and summer, can be transported long distances and is relatively rich in P (Bassett and others 1978; Doskey and Ugoagwu 1989; Cho and others 2003). Phosphorus and K are usually strongly correlated in biological tissues (Sabota and others 1984), which suggests that K:P ratios can be used to indicate biological contamination in bulk deposition. Ratios of K:P (water-extractable) in pollen from various Pinus species range between 2.8:1 and 3.6:1 (Doskey and Ugoagwu 1992; Lee and others 1996a; Cho and others 2003; Lee and Booth 2003). The K:P ratio measured in pollen digests (acid) in this study (4.5:1) is similar to the previously published water-extractable ratios and was almost identical to the K:P ratio in average (1985–2014) bulk deposition at PCP and HYP (4.4:1 and 4.5:1, respectively) and slightly higher than that at PTP (3.95:1). These results suggest a strong contribution of pollen and other biogenic material to historical deposition collections at all three sites, although the contribution of pollen to bulk deposition likely varies spatially and temporally depending on forest (conifers tend to be larger sources), understory composition (Lee and Booth 2003) and year-to-year differences in weather (Cho and others 2003). Indeed, temporal variability is the norm for pollen deposition studies, and pollen deposition has been observed to account for as little as 5% to as much as 100% of measured annual bulk TP deposition in other studies (Doskey and Ugoagwu 1989).
Large differences in measured bulk TP deposition among the four sites further support the importance of local sources of P deposition, as the contribution of external sources via long-range transport should be relatively consistent among proximal monitoring stations (Tsukuda and others 2006). Total P deposition was particularly high at the PTP site (~25 mg m−2 year−1−1 vs. 15–16 mg m−2 year−1−1 at PCP and HYP), and inter-annual variability was also greatest at this location. Higher TP deposition at PTP is almost entirely due to deposition in August and September, when P deposition at PTP is on average 4 mg m−2 greater than at PCP and HYP (temporal data not shown). Ground vegetation at the PTP station and the extent of human disturbance are also different from the other stations, and the PTP collectors have both the largest area of grass and are located the furthest distance from the tree line. As such, this pulse in P and K deposition in late summer/early fall is likely due to pollen from grasses (and weeds), which typically flower later than pine trees. Decco and others (1998), for example, found that grass pollen was released approximately 4 weeks after the climax release of Pinus pollen at Rochester, Minnesota, which is roughly the same latitude as the study sites.
The K:P ratio is also somewhat different at PTP (3.95:1) compared with the other sites (4.4–4.5:1), which suggests a contribution from vegetation other than Pinus. Interestingly, the mowing practices at PTP have changed over time (K. Somers, retired OMECC scientist, pers. comm.), with less frequent mowing in the early years of record over concern that lawnmower emissions would affect deposition measurements (mainly S and N), whereas in more recent years the lawn has been more consistently maintained. Consequently, in some years grasses may have matured to flowering, whereas in other years the potential contribution of pollen from grass species would have been lower, and this variable maintenance schedule may have contributed to high inter-annual variability at PTP.
The PTP station is also more influenced by roads than the other sites and is within 450 m of a gravel road, a gravel parking lot, a secondary paved road and a paved highway. Depending on region, road dust can contain 0.07% P and 1.87% K (by mass), and therefore inter-annual differences in growing season dryness and dust conditions may contribute to temporal variability at PTP. It should be noted that K:P ratios in road dust are typically much >10:1 (Tamm and Troedsson 1955; Hildemann and others 1991) and are therefore substantially higher than bulk deposition ratios. In addition, the lack of correspondence between TP and aluminum (Al) concentrations in deposition (data not shown) further suggest that dust is not an important source of P enrichment in this region (see Tsukuda and others 2006).
Mass Budget Calculations
In this study, our emphasis was on watershed budgets, and whether historical measurements of bulk deposition were a good indicator of external P inputs to forested landscapes. High intra- and inter-annual variability as well as site-to-site differences suggest that historical bulk deposition measurements in the growing season have been greatly inflated by internal P sources. Although above ground wet-only deposition collectors can help to reduce contamination from insects, bird droppings and local dust (for example, Peters and Reese 1995), high pollen counts in wet-only collections at PTP suggest that they too are sensitive to this local source. Relatively high TP concentrations in wet-only deposition during the growing season compared with the winter months and particularly high wet-only TP levels in the spring further suggest some biological contamination. In contrast, winter-only measurements of bulk deposition are much less variable both between years and among monitoring stations and are more similar to P and K levels in wet-only deposition, suggesting winter deposition may be the best indicator of external or ‘true’ inputs of P deposition in this region. Importantly, P input–output budgets using extrapolated winter deposition are much closer to balance (that is, no net accumulation or export) than estimates using measured bulk deposition in all seasons. For example, previous TP mass budget estimates that used a 3-station annual mean of 16 mg m−2 deposition (Eimers and others 2009) or 18 mg m−2 (Dillon and Molot 1997) indicated substantial net P accumulation (that is, positive budgets) at the majority of watersheds.
High inter-site variability also raises questions over the validity of using a 3-station mean to approximate regional deposition. Although the averaging process is useful for minimizing the effects of high, localized deposition such as that observed at PTP, it obscures some important information on sources of TP deposition. From the perspective of lake budgets, all sources of deposition should be considered, as any source of P could ultimately stimulate aquatic productivity. Based on these results, deposition measurements should be made as close as possible to the target water body, as historical 3-station annual mean deposition (16–19 mg m−2) would clearly underestimate P deposition inputs to Paint Lake (by close to 10 mg m−2 year−1−1), but overestimate inputs to Plastic, Heney and Harp Lakes (that is, contrast with annual totals in Table 1). The ‘error’ associated with using an average, regionally calculated deposition input would be particularly high in lakes with a large surface area-to-drainage area ratio, like Plastic Lake. In the absence of proximal deposition data, the ‘next best’ option would be to use deposition measurements from a site with similar vegetation to capture the potential contribution of pollen. Conversely, terrestrial watershed budgets should exclude all local deposition, and these results suggest that winter-only bulk deposition measurements may be extrapolated to estimate annual P inputs from long-range/external sources, assuming external sources are consistent across all seasons. This assumption is important to confirm and will likely only hold true in areas like the Precambrian Shield, where there are few external/long-range sources of mineral aerosols (for example, fertilized agriculture; desert dust) that could vary seasonally in magnitude (Mahowald and others 2008).
Contribution of Deposition to Observed Shifts in Surface Water and Vegetation P
Long-term monitoring of surface waters in this region has shown that lake and stream TP levels declined since the 1970s, even in lakes that have undergone substantial shoreline development (Eimers and others 2009; Palmer and others 2011). In addition, measurements of foliar chemistry and P addition experiments at forests within the region have suggested that trees (particularly sugar maple) may be shifting from N toward P limitation (Gradowski and Thomas 2006, 2008; Casson and others 2012). Despite these indications of P limitation, previous mass budget estimates for watersheds in this region are generally strongly positive (Dillon and Molot 1997; Eimers and others 2009), suggesting that TP is accumulating within the landscape. These observations were the primary motivation for this study. Our results suggest that trends in TP deposition should only be evaluated using winter-only measurements, as any trends in spring, summer, fall or annual deposition will be strongly influenced by internal biogenic sources of P, which vary year-to-year. Nevertheless, there were no trends in bulk P deposition annually, in any season or at any station, and most importantly, relatively stable winter bulk deposition over time suggests that there has been no change in external atmospheric P loading to this region over the period of record. This is in stark contrast to both S and N deposition, which have declined substantially in this region, although over different time periods (that is, SO4 declined over the entire record; NO3–N declines were primarily post 2000; Kothawala and others 2011; Watmough and others 2016). Negligible shifts in P deposition despite substantial declines in S and N suggest that in contrast to other parts of the world (for example, Japan; Tsukuda and others 2006) that long-range transport of fossil-fuel-derived P is not a substantial source of P deposition in this region.
Overall, these results suggest that year-round, multi-site monitoring of P bulk deposition is warranted to provide atmospheric input data for lake budgets and modeling, whereas winter bulk deposition may be a useful indicator of external P inputs in oligotrophic landscapes like the Precambrian Shield, where the contribution of P from long-range transport of dust is small. Importantly, the lack of decline in winter deposition suggests that external atmospheric loading has not contributed to observed shifts in either forest P limitation or surface water declines in this region.
References
Ahn H. 1999. Statistical modeling of total phosphorus concentrations measured in South Florida rainfall. Ecol Model 116:33–44.
Ahn H, James RT. 2001. Variability, uncertainty, and sensitivity of phosphorus deposition load estimates in South Florida. Water Air Soil Pollut 126:37–51.
Baker SR, Watmough SA, Eimers MC. 2015. Phosphorus forms and response to changes in pH in acid-sensitive soils on the Precambrian Shield. Can J Soil Sci 95:95–108.
Bassett IJ, Crompton CW, Parmelee JA. 1978. An atlas of airborne pollen grains and common fungus spores of Canada. Ottawa, Ontario: Canada Department of Agriculture.
Brown LJ, Taleban V, Gharabaghi B, Weiss L. 2011. Seasonal and spatial distribution patterns of atmospheric phosphorus deposition to Lake Simcoe, ON. J Great Lakes Res 37:15–25.
Casson NJ, Eimers MC, Watmough SA. 2012. An assessment of the nutrient status of sugar maple in Ontario: indications of phosphorus limitation. Environ Monit Assess 184:5917–27.
Cho Y, Kim IS, Kim P, Lee EJ. 2003. Deposition of airborne pine pollen in a temperate pine forest. Grana 42:178–82.
Cole JJ, Caraco NF, Likens GE. 1990. Short-range atmospheric transport: a significant source of phosphorus to an oligotrophic lake. Limnol Oceanogr 35:1230–7.
Dammgen U, Erisman JW, Cape JN, Grunhage L, Fowler D. 2005. Practical considerations for addressing uncertainties in monitoring bulk deposition. Environ Pollut 134:535–48.
Decco ML, Wendland BI, O’Connell EJ. 1998. Volumetric assessment of airborne pollen and spore levels in Rochester, Minnesota, 1992 through 1995. Mayo Clin Proc 73:225–9.
Dillon PJ, Kirchner WB. 1975. The effects of geology and land use on the export of phosphorus from watersheds. Water Res 9:135–48.
Dillon PJ, Lusis M, Reid R, Yap D. 1988. Ten-year trends in sulphate, nitrate and hydrogen deposition in central Ontario. Atmos Environ 22:901–5.
Dillon PJ, Molot LA. 1996. Long-term phosphorus budgets and an examination of a steady-state mass balance model for central Ontario lakes. Water Res 30:2273–80.
Dillon PJ, Molot LA. 1997. Effect of landscape form on export of dissolved organic carbon, iron, and phosphorus from forested stream catchments. Wat Resour Res 33:2591–600.
Doskey PV, Ugoagwu BJ. 1989. Atmospheric deposition of macronutrients by pollen at a semi-remote site in northern Wisconsin. Atmos Environ 23:2761–6.
Doskey PV, Ugoagwu BJ. 1992. Macronutrient chemistry of pollen. Commun Soil Sci Plant Anal 23:15–23.
Durham OC. 1946. The volumetric incidence of atmospheric allergens: IV. A proposed standard method of gravity sampling, counting, and volumetric interpolation of results. J Allergy 17:79–86.
Eimers MC, Watmough SA, Paterson AM, Dillon PJ, Yao W. 2009. Long-term declines in phosphorus export from forested catchments in south-central Ontario. Can J Fish Aquat Sci 66:1682–92.
Gradowski T, Thomas SC. 2006. Phosphorus limitation of sugar maple growth in central Ontario. For Ecol Manag 226:104–9.
Gradowski T, Thomas SC. 2008. Responses of Acer saccharum canopy trees and saplings to P, K and lime additions under high N deposition. Tree Phys 81:173–85.
Hamed KH, Rao AR. 1998. A modified Mann–Kendall trend test for autocorrelated data. J Hydrol 204:182–96.
Hildemann LM, Markowski GR, Cass GR. 1991. Chemical composition of emissions from urban sources of fine organic aerosol. Environ Sci Technol 25:744–59.
Jeppesen E, Søndergaard M, Jensen JP, Havens KE, Anneville O, Carvalho L, Coveney MF, Deneke R, Dokulil MT, Foy BOB, Gerdeaux D. 2005. Lake responses to reduced nutrient loading—an analysis of contemporary long-term data from 35 case studies. Freshwater Biol 50:1747–71.
Kirkwood EE, Nesbitt HW. 1991. Formation and evolution of soils from an acidified watershed: Plastic Lake, Ontario, Canada. Geochim Cosmochim Acta 55:1295–308.
Kothawala DN, Watmough SA, Futter MN, Zhang L, Dillon PJ. 2011. Stream nitrate responds rapidly to decreasing nitrate deposition. Ecosystems 14:274–86.
Lee EJ, Booth T. 2003. Macronutrient input from pollen in two regenerating pine stands in southeast Korea. Ecol Res 18:423–30.
Lee EJ, Kenkel NC, Booth T. 1996a. Pollen deposition in the boreal forest of west-central Canada. Can J Bot 74:1265–72.
Lee EJ, Kenkel NC, Booth T. 1996b. Atmospheric deposition of macronutrients by pollen in the boreal forest. Ecoscience 3:304–9.
Mahowald N, Jickells TD, Baker AR, Artaxo P, Benitez-Nelson CR, Bergametti G, Bond TC, Chen Y, Cohen DD, Herut B, Kubilay N, Losno R, Luo C, Maenhaut W, McGee K, Okin GS, Siefert RL, Tsukuda S. 2008. Global distribution of atmospheric phosphorus sources, concentrations and deposition rates, and anthropogenic impacts. Global Biogeochem Cycles 22:B4026. doi:10.1029/2008GB003240.
Newman EI. 1995. Phosphorus inputs to terrestrial ecosystems. J Ecol 83:713–26.
Palmer ME, Yan ND, Paterson AM, Girard RE. 2011. Water quality changes in south-central Ontario lakes and the role of local factors in regulating lake response to regional stressors. Can J Fish Aquat Sci 68:1038–50.
Paterson AM, Winter JG, Nicholls KH, Clark BJ, Ramcharan CW, Yan ND, Somers KM. 2008. Long-term changes in phytoplankton composition in seven Canadian Shield lakes in response to multiple anthropogenic stressors. Can J Fish Aquat Sci 65:846–61.
Peters NE, Reese RS. 1995. Variations of weekly atmospheric deposition for multiple collectors at a site on the shore of Lake Okeechobee, Florida. Atmos Environ 29:179–87.
Phillips T, Watmough SA. 2012. A nutrient budget for a selection harvest: implications for long-term sustainability. Can J For Res 42:2064–77.
Pinder KC, Eimers MC, Watmough SA. 2014. Impact of wetland disturbance on phosphorus loadings to lakes. Can J Fish Aquat Sci 71:1695–703.
Sabota A, Burovina IV, Pogorelov AG, Solus AA. 1984. Correlation between potassium and phosphorus content and their nonuniform distribution in Acanthamoeba castellanii. Histochemistry 81:201–4.
Scheider WA, Snyder WR, Clark B. 1979. Deposition of nutrients and major ions by precipitation in south-central Ontario. Water Air Soil Pollut 12:171–85.
Schindler DW, Newbury RW, Beaty KG, Campbell P. 1976. Natural water and chemical budgets for a small Precambrian lake basin in central Canada. J Fish Res Board Can 33:2526–43.
Skjelkvale BL, Stoddard JL, Jeffries DS, Tørseth K, Høgasen T, Bowman J, Mannio J, Monteith DT, Mosello R, Rogora M, Rzychon D, Vesely J, Wieting J, Wilander A, Worsztynowicz A. 2005. Regional scale evidence for improvements in surface water chemistry 1990–2001. Environ Pollut 137:165–76.
Tamm C, Troedsson T. 1955. An example of the amounts of plant nutrients supplied to the ground in road dust. Oikos 6:61–70.
Tsukuda S, Sugiyama M, Harita Y, Nishimura K. 2006. Atmospheric phosphorus deposition in Ashiu, Central Japan—source apportionment for the estimation of true input to a terrestrial ecosystem. Biogeochemistry 77:117–38.
Vicars WC, Sickman JO, Ziemann PJ. 2010. Atmospheric phosphorus deposition at a montane site: size distribution, effects of wildfire, and ecological implications. Atmos Environ 44:2813–44.
Watmough SA, Eimers MC, Baker S. 2016. Impediments to recovery from acid deposition. Atmos Environ 146:15–27.
Watmough SA, Dillon PJ. 2003a. Do critical load models adequately protect forests? A case study in south-central Ontario. Can J For Res 33:1544–56.
Watmough SA, Dillon PJ. 2003b. Major element fluxes from a coniferous catchment in central Ontario, 1983–1999. Biogeochemistry 67:369–99.
Yan ND, Somers KM, Girard RE, Paterson AM, Keller W, Ramcharan CW, Rusak JA, Ingram R, Morgan GE, Gunn JM. 2008. Long-term trends in zooplankton of Dorset, Ontario, lakes: the probable interactive effects of changes in pH, total phosphorus, dissolved organic carbon, and predators. Can J Fish Aquat Sci 65:862–77.
Acknowledgements
The authors thank Tim Field, Joe Findeis, Huaxia Yao and other staff at the Dorset Environmental Science Centre for logistical assistance and access to long-term data, and Andrew McDonough and Sarah Elliott for field and laboratory assistance. This research was supported by a Natural Sciences and Engineering Research Council (NSERC) Strategic Grant to MCE (Grant No. STGP 413170).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Eimers, M.C., Hillis, N.P. & Watmough, S.A. Phosphorus Deposition in a Low-Phosphorus Landscape: Sources, Accuracy and Contribution to Declines in Surface Water P. Ecosystems 21, 782–794 (2018). https://doi.org/10.1007/s10021-017-0184-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-017-0184-2