
Abstract
Nicotine has a profound influence on the carotenoid metabolism in halophilic Archaea of the class Halobacteria. In a study of Halobacterium salinarum, Haloarcula marismortui and Halorubrum sodomense, using different analytical techniques to monitor the production of different carotenoids as a function of the presence of nicotine, we showed that the formation of α-bacterioruberin was inhibited in all. In Hbt. salinarum, addition of nicotine led to a significant change in the color of the culture due to the accumulation of lycopene, in addition to the formation of bisanhydrobacterioruberin which does not differ in color from α-bacterioruberin. Very little or no lycopene was formed in Har. marismortui and in Hrr. sodomense; instead bisanhydrobacterioruberin was the only major carotenoid found in nicotine-amended cultures. The findings are discussed in the framework of the recently elucidated biochemical pathway for the formation of the different carotenoid pigments encountered in the Halobacteria.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Archaea of the class Halobacteria are generally colored pink–red due to C50 carotenoids, especially bacterioruberin and its mono-, bis- and trisanhydro derivatives (Straub 1987). Their structures were elucidated in the early 1970s (Kelly et al. 1970; Kushwaha et al. 1975). Small amounts of other carotenoid compounds are often present, including lycopene, the precursor for the biosynthesis of C50 bacterioruberins and of C40 carotenoids, notably β-carotene that yields the retinal moiety of the light-driven proton pump bacteriorhodopsin and other retinal proteins. No consistent correlation was found between the amount of C50 and C40 carotenoids, suggesting that the biosynthetic pathways are independent (Kushwaha et al. 1974). Minor C50-carotenoids have also been detected, including bacterioruberin derivatives with an epoxy group or a peroxide end group (Rønnekleiv et al. 1995). Haloferax volcanii contained bacterioruberin (82%), monoanhydrobacterioruberin (7%), bisanhydrobacterioruberin (3%), and minor compounds including 2-isopentenyl-3,4-dehydrorhodopin (C45) (1%) and lycopene (C40) (0.3%) (Rønnekleiv and Liaaen-Jensen 1995). Haloarcula japonica had 68.1% bacterioruberin, 22.5% monoanhydrobacterioruberin, 9.3% bisanhydrobacterioruberin, < 0.1% isopentenyldehydrorhodopin, and traces of lycopene and phytoene (Yatsunami et al. 2014). No less than 30 carotenoids, including 7 geometric isomers for bacterioruberin, monoanhydrobacterioruberin and bisanhydroruberin, were identified in Haloterrigena turkmenica (Squillaci et al. 2017). Occasionally high contents of ketocarotenoids were reported in members of the group: “Haloferax alexandrinus” was claimed to accumulate canthaxanthin in amounts up to 34% of the total carotenoids (Asker and Ohta 1999, 2002). 3-Hydroxy-echinenone (11%) and trans-astaxanthin (24%) were found in Hbt. salinarum by Calo et al. (1995), who detected these pigments also in Har. hispanica and in Hfx. mediterranei. However, the analytical techniques used were not described in detail, and the findings need further confirmation.
Carotenoids have different functions in the Halobacteria. Growth of a colorless mutant of Hbt. salinarum was inhibited by strong light (Dundas and Larsen 1962, 1963). A colorless mutant also showed much lower resistance to UV, hydrogen peroxide, and DNA strand-breaks induced by ionizing radiation (Shahmohammadi et al. 1998). Antioxidant activity of bacterioruberins of Har. japonica was much higher than that of an equivalent amount of β-carotene (Yatsunami et al. 2014). Antioxidant activity was also reported for the pigments of Htg. turkmenica (Squillaci et al. 2017) and other members of the class (Hou et al. 2017). Bacterioruberins may also have a structural function: they reinforce the cell membrane (Lazrak et al. 1988) and they can be associated with retinal proteins (Yoshimura and Kouyama 2008).
A number of applications are being considered for the pigments of the Halobacteria (Rodrigo-Baños et al. 2015), including use as coloring agents, antioxidants and anti-cancer compounds, with Halobacterium, Haloferax, and Halorubrum spp. as proposed production strains (de la Vega et al. 2016; Sikkandar et al. 2013). Optimization of the cultivation of Hfx. mediterranei for pigment production has been reported (Chen et al. 2015; Fang et al. 2010).
Analytical methods used for the study of Halobacteria carotenoids included thin-layer chromatography in the early studies (Kelly et al. 1970; Kushwaha et al. 1975) and HPLC in more recent years (Oren and Rodríguez-Valera 2001; Squillaci et al. 2017). Resonance Raman spectroscopy was introduced in such studies about 10 years ago (Marshall et al. 2007; Fendrihan et al. 2009; Jehlička and Oren 2013; Jehlička et al. 2013, 2014a, b; Camacho-Córdova et al. 2014). However, the ability of Raman spectroscopy to discriminate between structurally slightly differing bacterioruberins is limited (Jehlička et al. 2014a, b; Harris et al. 2015).
To elucidate the carotenoid biosynthesis pathway in the Halobacteria, a cell-free system was developed for Hbt. salinarum in the 1970s; it converted isopentenyl diphosphate to trans-phytoene to lycopene via trans-phytofluene, ζ-carotene, and neurosporene (Kushwaha et al. 1976). Analysis of the intermediates accumulating in mutants of Har. japonica enabled elucidation of the complete pathway from lycopene to bacterioruberin (Yang et al. 2015) (Fig. 1).

The biosynthesis of bacterioruberins and other carotenoid pigments in halophilic Archaea, as based on the studies on Har. japonica by Yang et al. (2015), indicating the inhibitory effect of nicotine on different stages in the process leading to the accumulation of mainly bisanhydrobacterioruberin in Hrr. sodomense and Har. marismortui, with lycopene as a major product in Hbt. salinarum
Another approach that has contributed to the study of the biosynthesis of carotenoids of Halobacteria is the use of nicotine as a specific inhibitor. The effect of nicotine was earlier explored in studies on Mycobacterium, Flavobacterium and purple nonsulfur bacteria, where nicotine led to accumulation of lycopene and inhibition of cyclic carotenoids formation (Howes and Batra 1970; McDermott et al. 1973a, b). Nicotine at concentrations between 1 and 9 mM inhibited formation of bacterioruberin and monoanhydrobacterioruberin as well as of β-carotene in Hbt. salinarum and in Halococcus sp. and led to accumulation of lycopene and to a lesser extent of bisanhydrobacterioruberin (Kushwaha and Kates 1976). Labelling studies with [14C]mevalonate showed that 3 mM nicotine inhibited incorporation of mevalonate into bacterioruberin and monoanhydrobacterioruberin in Hbt. salinarum, while labeling of lycopene and bisanhydrobacterioruberin increased. Following transfer to nicotine-free medium, the labeling of bacterioruberin and monoanhydrobacterioruberin increased at the expense of lycopene and bisanhydrobacterioruberin. It was concluded that nicotine inhibits the hydration steps that convert bisanhydrobacterioruberin into monoanhydrobacterioruberin and finally into bacterioruberin, and that lycopene is the precursor for the C50 carotenoids (Kushwaha and Kates 1979a, b), supporting the earlier hypothesis that bacterioruberin is formed by addition of two isoprene units to a C40 skeleton with isopropylidene end groups, followed by introduction of four hydroxyl groups (Kelly et al. 1970). Hydroxyl groups at C1 and C1′ are introduced first by an enzyme system insensitive to nicotine, the remaining hydroxyls being later introduced by a nicotine-sensitive enzyme system.
As no further studies about the effects of nicotine on the carotenoid content of halophilic Archaea have been reported since the 1970, we examined its effect on Hbt. salinarum and on other members of the class Halobacteria in which such inhibitor studies were not earlier performed.
Materials and methods
Organisms
Halobacterium salinarum strain R1 (DSM 671) was grown in medium containing (g L−1): NaCl, 250; MgCl2·6H2O, 5.0; KCl, 5.0; NH4Cl, 5.0 and yeast extract, 10.0, pH 7.0. The medium for Halorubrum sodomense (ATCC 33755T) contained: NaCl, 125; MgCl2·6H2O, 160, K2SO4, 5.0; CaCl2·2H2O, 0.1; yeast extract, 1.0, casamino acids, 1.0, and soluble starch, 2.0; pH 7.0. Haloarcula marismortui ATCC 43049T was grown in medium containing NaCl, 206; MgSO4·7H2O, 36; KCl, 0.37; CaCl2·2H2O, 0.5; MnCl2, 0.013, and yeast extract, 5.0. Cultures were grown in 100 ml portions in 250 ml Erlenmeyer flasks or in 500 ml portions in 1 l flasks with shaking (120 rpm) at 35 °C in the light (45 μmol quanta m−2 s−1) in a New Brunswick Innova incubator. Nicotine (Sigma) was added at concentrations of 1 or 5 mM as indicated. At the end of the exponential growth phase cultures were harvested by centrifugation (8000×g for 20 min at 4 °C).
Cellular pigment absorption spectra
Carotenoids were extracted from cell pellets with methanol–acetone (1:1 by volume), the extracts were cleared by centrifugation, and the absorption spectra were measured against the solvent in a Cary 300 Bio UV–Visible spectrophotometer.
Analysis of carotenoids by HPLC
Pigments were extracted by grinding cell pellets with mortar and pestle in acetone: dichloromethane (1:1 by volume). Cell debris was discarded by centrifugation and the extracts were dried under a stream of N2. The carotenoids were dissolved in acetone and saponified by adding an equal volume of 60% KOH: ethanol (1:10) and incubating overnight at 4 °C. For extraction of carotenoids, an equal volume of diethyl ether and 10% volume of 12% NaCl were added to the saponification solution. The carotenoids were extracted following vortexing and separation of the hydrophobic fraction. The solvent was dried under a stream of N2 and the residue was dissolved in 100% acetone. Pigments were analyzed by HPLC as previously described, using a Waters 996 photodiode array detector (Ronen et al. 1999; Neuman et al. 2014). Carotenoids were identified by their characteristic absorption spectra and distinctive retention times. Quantitation was performed by integrating the peak areas of the HPLC results using Millennium chromatography software (Waters).
Raman spectroscopy analysis
Micro-Raman analyses of cell material dried by lyophilization were performed on a multichannel Renishaw In ViaReflex spectrometer coupled with a Peltier-cooled CCD detector. Excitation was provided by a 514.5 nm Ar laser (power ~ 10 mW). To improve signal-to-noise ratios, Raman signals from 30 to 60 scans were accumulated, each of 20-s exposure. Spectra were recorded at a resolution of 2 cm−1 between 800 and 2000 cm−1. Ten spectra were recorded at different points in the sample. Benzonitrile, silicon and diamond were used for spectral wavenumber calibration. Raman spectra were exported into the Galactic *.SPC format. Spectra were compared using GRAMS AI (V. 8.0, Thermo Electron Corp., Waltham, MA, USA). Raman spectra were not subjected to any data manipulation or processing techniques.
Results and discussion
The effect of nicotine on the pigmentation of different species of halophilic Archaea
When grown in the presence of nicotine (1 or 5 mM), cultures of Hbt. salinarum R1 were yellow–orange, as compared to the bright red color of cells grown without the inhibitor. At the higher nicotine concentration, growth was reduced. Microscopic examination did not show any changes in cell morphology between cells grown in the absence and in the presence of nicotine. Growth in the presence of nicotine did not lead to any visible color change of Hrr. sodomense and Har. marismortui cultures; however, growth rates were reduced, in the case of Har. marismortui already noticeable when added at a concentration of 1 mM.
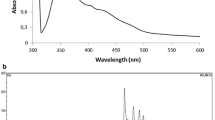
The absorption spectra of cell extracts in methanol–acetone 1:1 are shown in Fig. 2. When grown without nicotine, all three species show the characteristic absorption spectrum of α-bacterioruberin with a major absorption peak at 496 nm, a second maximum at 530 nm and a shoulder around 470 nm. When Hbt. salinarum was grown in the presence of nicotine, the 530 nm peak nearly disappeared, the 496 nm peak shifted to ~ 500 nm, and a prominent maximum appeared at 472–474 nm. The pigment spectrum of Har. marismortui grown in the presence of 1 mM nicotine was no different from that of the control without inhibitor. When grown with 5 mM nicotine the relative size of the 530 nm peak was somewhat reduced and the major 496 nm peak shifted to ~ 500 nm. No prominent differences were observed in the absorption spectra of extracts of Hrr. sodomense grown without and with 1 or 5 mM nicotine.

Absorption spectra of methanol–acetone extracts of cells of Hbt. salinarum, Har. marismortui and Hrr. sodomense grown with and without nicotine (1 and 5 mM)
HPLC analysis (Figs. 3, 4) showed prominent differences between the types of carotenoids produced in the presence and in the absence of nicotine in all three species, also in cases where the absorption spectra as shown in Fig. 2 were not significantly different. In all three organisms, addition of nicotine led to the disappearance of the major peak of α-bacterioruberin. Lycopene appeared as a major product only in Hbt. salinarum, confirming the findings of Kushwaha and Kates (1976, 1979a, b). However, only little lycopene was found in pigment extracts of Har. marismortui incubated with the higher concentration of nicotine, and none was detected in Hrr. sodomense at any nicotine concentration. The pigment accumulating instead of α-bacterioruberin could be identified on the basis of its elution time and its absorption spectrum as bisanhydrobacterioruberin (Fig. 4, Table 1). We, thus, confirm that nicotine inhibits the hydration steps that convert bisanhydrobacterioruberin into monoanhydrobacterioruberin and finally into bacterioruberin, and that lycopene is the precursor for the C50 carotenoids, as reported in studies with a strain of Hbt. salinarum (Kushwaha and Kates 1979a, b). However, the ratio in which lycopene and bisanhydrobacterioruberin are formed differs greatly in different members of the Halobacteria. This explains the lack or very small extent of color change in cultures of Hrr. sodomense and Har. marismortui when grown in the presence of nicotine (Fig. 2). The absorption spectrum of bisanhydrobacterioruberin differs very little from that of α-bacterioruberin, but the absorption peaks of lycopene are blue-shifted by more than 20 nm (Fig. 4).

HPLC (chromatogram of pigment extracts of Hbt. salinarum, Har. marismortui and Hrr. sodomense grown with and without nicotine. The absorbance is given in relative units, based on the maximum absorbance–wavelength of each fraction. The fractions can tentatively be identified as (1) apocarotenoid (likely retinal); (2) an unidentified bacterioruberin derivative; (3) α-bacterioruberin; (4) monoanhydrobacterioruberin; (5) bisanhydrobacterioruberin; (6) lycopene

Spectra of HPLC peaks indicated in Fig. 3 as recorded during elutions in acetonitrile–H2O/ethylacetate gradients. Tentative assignments of the peaks: (1) apocarotenoid (likely retinal); (2) a bacterioruberin; (3) α-bacterioruberin; (4) monoanhydrobacterioruberin; (5) bisanhydrobacterioruberin; (6) lycopene; (7): unidentified
Another inhibitor known to interfere with the production of normal carotenoids and cause accumulation of probable biosynthetic intermediates is diphenylamine (McDermott et al. 1973b). It was earlier reported that diphenylamine at concentrations as low as 30 μM inhibits growth of Hbt. salinarum (Kushwaha and Kates 1979b). We found that 50 μM diphenylamine also completely inhibited growth of Har. marismortui and Hrr. sodomense. This inhibitor is, thus, not suitable for studies on carotenoid biosynthesis in the Halobacteria.
The results of the resonance Raman spectroscopy assays are summarized in Fig. 5 and in Table 2. Bacterioruberin and derivatives have characteristic Raman bands at 1505/1508 cm−1 [ν(C=C)], 1151/1154 cm−1 [(ν(C–C)], and 1000/1003 cm−1 [δ(C=CH)] (Marshall et al. 2007; Fendrihan et al. 2009; Jehlička and Oren 2013; Jehlička et al. 2013; Camacho-Córdova et al. 2014). The wavenumber positions of both ν1 and ν2 bands depend on the length of the conjugated chain (Merlin 1985; Withnall et al. 2003). In the case of all microorganisms grown in the absence of nicotine, Raman spectra show the dominant bacterioruberin with possible traces of its derivatives. The ν1 band obtained for the nicotine-treated sample of Hbt. salinarum is slightly shifted to 1512 cm−1. This can be attributed to the presence of a longer polyene chain carotenoid such as lycopene with eleven C=C conjugated bonds (de Oliveira et al. 2010). However, the less intense, complex broad band contains also a component with roughly the same wavelength as observed in the untreated sample (1506 cm−1), confirming the possible presence of a bacterioruberin derivative. No significant differences were observed in the carotenoid Raman spectra of Har. marismortui and Hrr. sodomense as a result of the presence of nicotine, consistent with low amounts of lycopene, if at all, detected in these samples using HPLC; any other possible qualitative changes in the types of carotenoids present were not reflected in the Raman spectra recorded. Raman spectroscopy is not sufficiently sensitive to differentiate, e.g., between α-bacterioruberin and bisanhydrobacterioruberin, compounds that do not differ in the length of the conjugated chain. Overall, the ability of Raman spectroscopy to discriminate between structurally slightly differing carotenoid pigments is limited (Jehlicka et al. 2014a, b; Harris et al. 2015).

Raman spectra of lyophilized cells of Hbt. salinarum grown without (a) and in the presence of 5 mM nicotine (b). The spectra contain the characteristic bands of carotenoids: the ν1 band located at around 1506 cm−1 (C=C in-phase stretching vibrations), the ν2 band at 1152 cm−1 (C–C stretching vibrations) and the feature of medium intensity at 1000/1003 cm−1 (ν3) (attributed to the in-plane rocking modes of the CH3 groups attached coupled with C–C bonds to the polyene chain)
The scheme presented in Fig. 1, based on the biosynthetic pathway of carotenoid pigments in the Halobacteria as elucidated for Har. japonica (Yang et al. 2015), explains the observations presented. The enzymatic step that is strongly affected by nicotine in all three organisms examined is the conversion of bisanhydrobacterioruberin into monoanhydrobacterioruberin, the direct precursor of α-bacterioruberin. Therefore, addition of nicotine leads to the accumulation of bisanhydrobacterioruberin. A second reaction that is sensitive to nicotine to a different extent in the organisms tested is the elongation of lycopene (C40) to a C45 intermediate (dihydroisopentyldehydrorhodopin). Thus, lycopene accumulates in Hbt. salinarum, as observed already in the 1970s (Kushwaha and Kates 1976, 1979a, b). But this effect was absent or nearly absent in the other two species included in our study.
References
Asker D, Ohta Y (1999) Production of canthaxanthin by extremely halophilic bacteria. J Biosci Bioengin 88:617–621
Asker D, Ohta Y (2002) Production of canthaxanthin by Haloferax alexandrinus under non-aseptic conditions and a simple, rapid method for its extraction. Appl Microbiol Biotechnol 58:743–750
Calo P, de Miguel T, Sieiro C, Velazquez JB, Villa TG (1995) Ketocarotenoids in halobacteria: 3-hydroxy-echinenone and trans-astaxanthin. J Appl Bacteriol 79:282–285
Camacho-Córdova DI, Camacho-Ruíz RM, Córdova-López JA, Cervantes-Martínez J (2014) Estimation of bacterioruberin by Raman spectroscopy during the growth of halophilic archaeon Haloarcula marismortui. Appl Opt 53:7470–7475
Chen CW, S-h Hsu, Lin M-T, Y-h Hsu (2015) Mass production of C50 carotenoids by Haloferax mediterranei in using extruded rice bran and starch under optimal conductivity of brine medium. Bioprocess Biosyst Env 38:2361–2367
de la Vega M, Sayago A, Ariza J, Barneto AG, León R (2016) Characterization of a bacterioruberin-producing haloarchaea isolated from the marshlands of the Odiel River in the southwest of Spain. Biotechnol Progr 32:592–600
de Oliveira VE, Castro HV, Edwards HGM, de Oliveira LFC (2010) Carotenes and carotenoids in natural biological samples: a Raman spectroscopic analysis. J Raman Spectrosc 41:642–650
Dundas ID, Larsen H (1962) The physiological role of the carotenoid pigments of Halobacterium salinarium. Arch Mikrobiol 44:233–239
Dundas ID, Larsen H (1963) A study on the killing by light of photosensitized cells of Halobacterium salinarium. Arch Mikrobiol 46:19–28
Fang C-J, Ku K-L, Lee M-H, Su N-W (2010) Influence of nutritive factors on C50 carotenoids production by Haloferax mediterranei ATCC 33500 with two-stage cultivation. Biores Technol 101:6487–6493
Fendrihan S, Musso M, Stan-Lotter H (2009) Raman spectroscopy as a potential method for the detection of extremely halophilic archaea embedded in halite in terrestrial and possibly extraterrestrial samples. J Raman Spectrosc 40:1996–2003
Harris LV, McHugh M, Hutchinson IB, Ingley R, Malherbe C, Parnell J, Marshall AO, Edwards HGM (2015) Avoiding misidentification of bands in planetary Raman spectra. J Raman Spectrosc 46:863–872
Hou J, Gao X, Lü Z-Z, Li Y, Zhou Y, Cui H-L (2017) In vitro antioxidant, antihemolytic and anticancer activity of the carotenoids from halophilic archaea. Curr Microbiol. https://doi.org/10.1007/s00284-017-1374-z
Howes CD, Batra PP (1970) Accumulation of lycopene and inhibition of cyclic carotenoids in Mycobacterium in the presence of nicotine. Biochim Biophys Acta 222:174–179
Jehlička J, Oren A (2013) Raman spectroscopy in halophile research. Front Microbiol 10:380
Jehlička J, Edwards HGM, Oren A (2013) Bacterioruberin and salinixanthin carotenoids of extremely halophilic Archaea and Bacteria: a Raman spectroscopic study. Spectrosc Acta A 106:99–103
Jehlička J, Edwards HGM, Oren A (2014a) Raman spectroscopic of microbial pigments. Appl Environ Microbiol 80:3286–3295
Jehlička J, Edwards HGM, Osterrothová K, Novotná J, Nedbalová L, Kopecký J, Němec I, Oren A (2014b) Potential and limits of Raman spectroscopy for carotenoid detection in microorganisms: implications for astrobiology. Philos Trans R Soc A 372:20140199
Kelly M, Norgård S, Liaaen-Jensen S (1970) XXXI. C50 carotenoids of Halobacterium salinarium, especially bacterioruberin. Acta Chem Scand 24:2169–2182
Kushwaha SC, Kates M (1976) Effect of nicotine on biosynthesis of C50 carotenoids in Halobacterium cutirubrum. Can J Biochem 54:824–829
Kushwaha SC, Kates M (1979a) Studies on the biosynthesis of C50 carotenoids in Halobacterium cutirubrum. Can J Microbiol 25:1292–1297
Kushwaha SC, Kates M (1979b) Effect of nicotine on carotenogenesis in extremely halophilic bacteria. Phytochemistry 18:2061–2062
Kushwaha SC, Gochnauer MB, Kushner DJ, Kates M (1974) Pigments and isoprenoid compounds in extremely and moderately halophilic bacteria. Can J Microbiol 20:241–245
Kushwaha SC, Kramer JKG, Kates M (1975) Isolation and characterization of C50-carotenoid pigments and other polar isoprenoids from Halobacterium cutirubrum. Biochim Biophys Acta 398:303–314
Kushwaha SC, Kates M, Porter JW (1976) Enzymatic synthesis of C40 carotenes by cell-free preparation from Halobacterium cutirubrum. Can J Biochem 54:816–823
Lazrak T, Wolff G, Albrechts A-M, Nakatani Y, Ourisson G, Kates M (1988) Bacterioruberins reinforce reconstituted Halobacterium lipid membranes. Biochim Biophys Acta 939:160–162
Marshall CP, Leuko S, Coyle CM, Walter MR, Burns BP, Neilan BA (2007) Carotenoid analysis of halophilic archaea by resonance Raman spectroscopy. Astrobiology 7:631–643
McDermott JCB, Ben-Aziz A, Singh RK, Britton G, Goodwin TW (1973a) Recent studies of carotenoid biosynthesis in bacteria. Pure Appl Chem 35:29–46
McDermott JCB, Britton G, Goodwin TW (1973b) Effect of inhibitors of zeaxanthin synthesis in a Flavobacterium. J Gen Microbiol 77:161–171
Merlin JC (1985) Resonance Raman spectroscopy of carotenoids and carotenoid-containing systems. Pure Appl Chem 57:785–792
Neuman H, Galpaz N, Cunningham FX Jr, Zamir D, Hirschberg J (2014) The tomato mutation nxd1 reveals a gene necessary for neoxanthin biosynthesis and demonstrates that violaxanthin is a sufficient precursor for abscisic acid biosynthesis. Plant J 78:80–93
Oren A, Rodríguez-Valera F (2001) The contribution of halophilic Bacteria to the red coloration of saltern crystallizer ponds. FEMS Microbiol Ecol 36:123–130
Rodrigo-Baños M, Garbayo I, Vílchez C, Bonete MJ, Martínez-Espinosa RM (2015) Carotenoids from haloarchaea and their potential in biotechnology. Mar Drugs 13:5508–5532
Ronen G, Cohen M, Zamir D, Hirschberg J (1999) Regulation of carotenoid biosynthesis during tomato fruit development: expression of the gene for lycopene epsilon-cyclase is down-regulated during ripening and is elevated in the mutant Delta. Plant J 17:341–351
Rønnekleiv M, Liaaen-Jensen S (1995) Bacterial carotenoids 53, C50-carotenoids 23; carotenoids of Haloferax volcanii versus other halophilic bacteria. Biochem Syst Ecol 23:627–634
Rønnekleiv M, Lenes M, Norgård S, Liaaen-Jensen S (1995) Three dodecaene C50-carotenoids from halophilic bacteria. Phytochemistry 39:631–634
Shahmohammadi HR, Asgarani E, Terato H, Saito T, Ohyama Y, Gekko K, Yamamoto O, Ide H (1998) Protective roles of bacterioruberin and intracellular KCl in the resistance of Halobacterium salinarium against DNA-damaging agents. J Radiat Res 39:251–262
Sikkandar S, Murugan K, Al-Sohaibani S, Rayappan F, Nair A, Tilton F (2013) Halophilic bacteria—a potent source of carotenoids with antioxidant and anticancer potentials. J Pure Appl Microbiol 7:2825–2830
Squillaci G, Parrella R, Carbone V, Minasi P, La Cara F, Morana F (2017) Carotenoids from the extreme halophilic Haloterrigena turkmenica: identification and antioxidant activity. Extremophiles 21:933–945
Straub O (1987) In: Pfander H, Gerspacher M, Rychener M, Schwabe R (eds) Key to carotenoids, 2nd edn. Birkhäuser Verlag, Basel, pp 11–218
Withnall R, Chowdhry BZ, Silver J, Edwards HGM, de Oliveira LFC (2003) Raman spectra of carotenoids in natural products. Spectrochim Acta A 59:2207–2212
Yang Y, Yatsunami R, Miyoko N, Fukui T, Takaichi S, Nakamura S (2015) Complete biosynthetic pathway of the C50 carotenoid bacterioruberin from lycopene in the extremely halophilic archaeon Haloarcula japonica. J Bacteriol 197:1614–1623
Yatsunami R, Ando A, Yang Y, Takaichi S, Kohno M, Matsumara Y, Ideka H, Fukui T, Nakasone K, Fujita N, Sekine M, Takashina T, Nakamura S (2014) Identification of carotenoids from the extremely halophilic archaeon Haloarcula japonica. Front Microbiol 5:100
Yoshimura K, Kouyama T (2008) Structural role of bacterioruberin in the trimeric structure of archaerhodopsin-2. J Mol Biol 375:1267–1281
Acknowledgements
We thank Lily Mana for technical assistance. AO was supported by Grant no. 2221/15 from the Israel Science Foundation. This study was further supported by the Erasmus+ inter-institutional agreement between the Charles University, Prague, and the Hebrew University of Jerusalem. JJ was funded by the Czech Science Foundation Project 17-04270S. Work in the laboratory of JH is funded by Israel Science Foundation Grant ISF 850/13.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by G. Antranikian.
Rights and permissions
About this article

Cite this article
Oren, A., Hirschberg, J., Mann, V. et al. Effects of nicotine on the biosynthesis of carotenoids in halophilic Archaea (class Halobacteria): an HPLC and Raman spectroscopy study. Extremophiles 22, 359–366 (2018). https://doi.org/10.1007/s00792-018-0995-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00792-018-0995-x