
Abstract
Objectives
Alveolar bone structures are mostly investigated in small animal models. The majority of these studies examined local influences on the alveolar bone, but only a few examined systemic influencing factors. The hypothalamic-pituitary axis is known to be essential for a vital bone balance. The aim of this study was to analyse the effects that selective hormone treatments have on alveolar bone structure and quality in a sheep model for alveolar bone loss, induced by hypothalamic-pituitary disconnection (HPD).
Methods
Thirty sheep were randomly selected into six groups of five each: control (C), ovariectomy—OVX (O), O + HPD (OH), OH with oestrogen treatment (OHE), OH with thyroxine (T4) treatment (OHT), and OH with a combined treatment of oestrogen and thyroxine (OHTE). After OVX and HPD procedures and an additional 9-month observation/treatment period, structural bone analyses of the mandible were performed by contact radiography, micro-CT, and static histomorphometry.
Results
The HPD procedure caused structural alveolar bone parameters to decrease significantly compared to controls (C). Treatment with oestrogen (OHE) was protective and bone structure was maintained at baseline levels. Thyroxine treatment (OHT) promoted significant bone loss, but the combined treatment (OHTE) improved bone structure and volume parameters even above baseline levels.
Conclusions
Alveolar bone homeostasis significantly underlies systemic regulatory systems. Centrally induced (HPD) bone loss can be prevented by combined peripheral treatment with oestrogen and thyroxine.
Clinical relevance
These results demonstrate the significance of a balanced hormonal regulatory system for steady bone remodelling and maintenance of healthy alveolar bone.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Oral health depends on a good dental status (no decay or tooth erosions), the periodontium and healthy alveolar bone. Local processes like inflammatory reactions in the periodontium can cause significant alveolar bone loss, which can lead to tooth loss contributing to decreased dental health status [1,2,3]. Systemic diseases like osteoporosis cause a significant disruption in the entire skeletal system, with decreased bone quality. This systemic disease not only leads to typical osteoporotic fractures in long and vertebral bones but also causes reduced alveolar bone quality and hence promotes tooth loss [4,5,6].
Some animal models exist regarding alveolar bone loss with local processes like periodontal disease [7], but only a few have been described for alveolar bone loss caused by a systemic disorder. One type of model described alveolar bone loss in ovariectomised rats mimicking postmenopausal bone loss [8,9,10,11], while another model was in sheep with steroid-induced bone loss [12, 13]. Our group recently published a study of surgical hypothalamic-pituitary disconnection (HPD) in a sheep model inducing a low-bone-turnover situation with depressed bone formation and resorption leading to cortical and trabecular bone loss in vertebral and non-vertebral bones [14]. Further analyses demonstrated the systemic influence caused by the HPD and ovariectomy (OVX) procedures, which also affects alveolar bone metabolism, consequently leading to significant alveolar bone loss [15]. Therefore, the effects of OVX and HPD procedures, particularly the latter one, cause a subsequent change in hormone levels. This again leads to a major polysystemic effect on the entire bone metabolism and skeletal system leading to reduced bone quality and biomechanical alterations [16, 17].
The aim of the current study was to analyse whether a surgically induced systemic bone disorder (HPD) can be partially reduced or prevented by hormone treatment (oestrogen, thyroxine, and combined treatments) and hence help with the maintenance of proper alveolar bone structures.
Materials and methods
Animals, biochemical analyses, hypothalamo-pituitary disconnection, and treatment regimens
Thirty adult female Corriedale ewes of normal body weight (58.2 ± 6.2 kg) aged 4–5 years were obtained from a laboratory flock. The sheep were housed together in a paddock and familiarised with the experimental facilities. Five sheep were randomly selected for each of the six groups: no intervention - control entire—C, ovariectomy (OVX)—O, OVX and HPD—OH, OVX + HPD with oestrogen treatment—OHE, OVX + HPD with thyroxine (T4) treatment—OHT, and OVX + HPD with a combined treatment of oestrogen and thyroxine—OHTE.
Under general anaesthesia, bilateral ovariectomy was performed via a paramedian laparotomy in 25 animals (O, OH, OHE, OHT, and OHTE). The HPD animals (n = 20) were all ovariectomised at least 1 week before HPD surgery.
A total of 20 ewes were operated upon with a standardised hypothalamic-pituitary disconnection (HPD), as previously published [18]. In short, the operation used a paramedian, transnasal, and transphenoidal approach to expose the pituitary gland, median eminence, and optic chiasm. The median eminence was entered above the hypophysial portal circulation, then the neural tissue of the tuber cinereum was cleared, leaving the pituitary gland disconnected from the hypothalamus.
An aluminium foil barrier was placed after the entire removal of the local neural tissue between the hypothalamus and the pituitary gland to prevent regenerative tissue growth. This process led to atrophy of the pars nervosa and to hypertrophy of the pars intermedia of the pituitary gland, without causing infarction of the pars distalis, which is essential for the survival of the ewes [18].
Afterwards, one HPD group (OH) did not receive any further treatment. The remaining three groups (OHE, OHT, and OHTE) received hormone replacement therapy (oestrogen, thyroxine, or both) starting 2 weeks after HPD procedure. Oestrogen treatment was administered by silicone implants, 30 × 5 mm each, containing 43.9 mg oestradiol (Compudose 400, Elanco Animal Health, Auckland, New Zealand). The implants were placed subcutaneously in the inguinal region of the abdomen, as previously described [19].
The two groups on thyroxine treatment (OHT and OHTE) received thyroxine (Sigma-Aldrich Chemie, Steinheim, Germany) three times per week in the form of subcutaneous injections of crystalline L-T4 (5 μg/kg), which was dissolved in 0.3 N sodium hydroxide and 50% ethanol before use, and later diluted in a sodium bicarbonate-buffered saline solution for injection as described previously [20]. The group on both treatment regimens (OHTE) received the oestrogen-containing implants and the thrice-weekly thyroxine injections accordingly.
Blood samples were drawn during the experimental period in order to check the thyroxine levels. Thyroxine was checked every other day after injection. As oestrogen substitution via silicone implants is already established [19], measuring of the oestrogen profile was not part of the study. There was no special diet nor were any other treatments given. There were no silages and also no specific soja-free diet fed. All sheep were kept on pasture. Water was given ad libitum.
After 9-months of treatment, the animals were sacrificed by an intravenous overdose injection of sodium pentobarbital (Lethabarb; May and Baker Pty. Ltd., Melbourne, Australia). The entire mandibles were dissected out at death, fixed in 3.7% PBS-buffered formaldehyde for 3 days at 4 °C and subsequently transferred into 70% ethanol for further histomorphometric analyses.
Blood samples (8 ml) were taken from the jugular vein immediately after sacrifice and collected in heparinised tubes. These were centrifuged at 4 °C to obtain plasma, which was stored at −80 °C until it was assayed. Urine samples for the determination of urinary deoxypyridinoline (DPD) cross-lap excretion as a measure of bone resorption were also kept at −80 °C until they were assayed. All serum (sodium, potassium, chloride, calcium, phosphate, alkaline phosphatase (AP), bone alkaline phosphatase (bAP), osteocalcin, and thyroxine) and urine (DPD) parameters were measured with the use of commercial kits.
All procedures and tissue collections were in accordance with the guidelines for the Care and Maintenance of Experimental Animals and with the prior approval of the Monash Medical Centre Animal Ethics Committee and the Animal Experimentation Ethics Committee of Victorian Institute of Animal Science, Victoria, Australia.
Contact radiography and micro-computer tomography
Mandible samples of two regions of interest (ROI)—diastema (ROI 1) and mandibular angle (ROI 2) (Fig. 1)—were analysed by using contact radiography with a Faxitron X-ray cabinet (Faxitron X-ray Corp., Wheeling, IL, USA).

Regions of interest (ROI) are marked by a red line. ROI 1 is the diastema and ROI 2 is the angle of the mandible. Section samples were cut out and were accordingly demonstrated by contact radiography
Diastema (ROI 1) and mandibular angle (ROI 2) sections of all mandibles were scanned (55 kV/145 μA) in a micro-computer tomography (μCT) 40 (Scanco Medical AG, Bruettisellen, Switzerland) for evaluation and 3D visualisation of cortical thickness (Ct.Th) and porosity, as previously described [14, 16]. In addition, ramus sections (ROI 2) were scanned as described above for evaluation and 3D visualisation of bone volume per tissue volume (BV/TV), trabecular thickness (Tb.Th), trabecular number (Tb.N), and trabecular spacing (Tb.Sp).
The generated raw data were manually segmented and assessed using the μCT Evaluation Program V6.0 (Scanco Medical, Bruettisellen, Switzerland). For visualisation, the segmented data were imported and displayed in μCT Ray V3.8 (Scanco Medical, Bruettisellen, Switzerland).
Static histomorphometry
The methods were performed as previously described [16]. After fixation of the bones and transfer into 70% ethanol, they were dehydrated in ascending alcohol concentrations and embedded in methyl methacrylate. The regions of interest (ROIs) were cut into sections of 5 μm in the frontal plane for the diastema and ramus (Fig. 1) with a Microtec rotation microtome (Techno-Med, Munich, Germany). Hereafter, the sections were stained by van Gieson/von Kossa procedure [21].
Static histomorphometry parameters were quantified on three different undecalcified von Kossa-stained sections of 5 μm thickness from the ROI 2, as seen in previous descriptions [22, 23]. Evaluation was carried out according to ASBMR standards [24, 25] of bone volume (BV/TV), trabecular number (Tb.N), trabecular separation (Tb.Sp), trabecular thickness (Tb.Th) by the use of OsteoMeasure histomorphometry system (Osteometrics, Atlanta, GA, USA) and Bioquant (Bioquant Image Analysis, Inc., Nashville, TN, USA) connected to a Zeiss microscope (Carl Zeiss, Jena, Germany).
Statistical analyses
Statistical analysis was conducted using IBM® SPSS® Statistics 14. Analyses’ results are shown as mean ± standard deviation (SD). Assessment for statistical differences between groups was done by one-way ANOVA. The Tukey post hoc honestly significant difference test was used to adjust for multiple comparisons. All statistical tests were two-sided and conducted in an explorative manner at a 5% significance level. The significance levels are marked by * or—p < 0.05 and ** or—p < 0.01.
Results
Animals, ovariectomy, hypothalamo-pituitary disconnection, and treatments
The study setup was initiated by operative procedures and continued with treatment for 9 months. Surgical procedures (OVX and HPD) were successful in all sheep. Treatment implants and injections for the three treatment groups were well tolerated by the animals. None of the animals, including treatment groups, showed signs of infections nor was any hair loss or conspicuous behaviour noted. All ewes grazed on pasture with no further treatments or other special needs. As successful clinical signs of the hypothalamo-pituitary disconnection, polydipsia and polyurea resulting in diabetes insipidus were observed and a moderate gain of body weight of 5.8 ± 0.8 kg overall without significant differences between the groups was noted after 9 months of treatment/observation.
Biochemical profiles
Serum electrolyte levels (sodium, potassium, chloride, calcium, and phosphate) and bone-turnover parameters did not show any significant differences between the six groups at the time of sacrifice. Measurement of serum thyroxine levels throughout the 9-month treatment period demonstrated continuously significant lower levels in the OH and OHE groups than in both control groups (C and O), validating successful HPD surgery. Substitution of thyroxine in the two treatment groups (OHT—9.30 ± 2.46 pmol/l; OHTE—9.69 ± 1.41 pmol/l) was likewise effective since their overall measured thyroxine levels were comparable to levels of control animals (C—8.52 ± 1.59 pmol/l; O—8.28 ± 0.94 pmol/l). Significantly lower values of thyroxine were measured for untreated controls (OH 5.65 ± 1.48 pmol/l; OHE 5.56 ± 0.75 pmol/l) [26]. Oestrogen silicone implants were well tolerated by the animals throughout the entire experiment. The release profile of oestrogen silicone implants is well established and provides oestrogen levels in physiological range for the two substitution groups (OHE and OHTE) [19, 27].
Analysis of cortical and trabecular bone at the diastema and mandibular angle
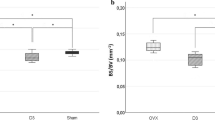
ROI 1 at the diastema was used for analyses of the cortical bone (Fig. 2). Control (C) and OVX (O) groups showed steady levels for the cortical thickness, 2.45 ± 0.21 and 2.32 ± 0.15 mm, respectively. There was a significant decrease in cortical thickness in the OH (1.98 ± 0.20 mm**) and the group treated with thyroxine (OHT) (1.89 ± 0.11 mm**) in comparison to control entire (C). Treatment with oestrogen (OHE) or the combination of oestrogen and thyroxine (OHTE) helped to keep cortical thickness at the baseline levels or even increase it, 2.45 ± 0.17 and 2.59 ± 0.21 mm, respectively. Cortical porosity was measured for the control group (C) at 0.79 ± 0.31% and for OVX group (O) at 1.01 ± 0.29%. It decreased by 12.66% within the oestrogen group (OHE) and by 31.65% within the group of combined oestrogen and thyroxine treatment (OHTE) compared to control entire (C). However, the increase in cortical porosity was more significant, which more than doubled compared to control entire (C), for OH (1.60 ± 0.52%**) and OHT (1.65 ± 0.75%**).

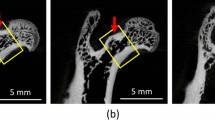
μCT scans of diastema (ROI 1) from control entire (=C) (a), O (b), OH (c), OHE (d), OHT (e), and OHTE (f). An increase in porosity is demonstrated in scans c and e in comparison to control entire (a). After one-way ANOVA for cortical thickness (Ct.Th), porosity (%) and bone mineral density (BMD) significances are visualised in part g with bar graphs (--- significance when p < 0.05 or ___ when p < 0.01 level)
The mandibular angle (ROI 2) was thoroughly analysed on trabecular and cortical structure levels (Fig. 3). Scans of the HPD (OH) (Fig. 3c) and thyroxine treatment group (OHT) (Fig. 3e) showed a decreased trabecular network when compared with the control (Fig. 3a). Bone volume per tissue volume (BV/TV) was 20.2 ± 2.5% in the control group (C). There was a significant reduction in bone volume noted for the HPD group (OH) by 32.17%* and by 25.59%* for the ewes receiving thyroxine treatment (OHT) compared to control entire (C). On the other hand, the ewes on combined treatment (OHTE) had a significant increase of 21.89 and 39.37% in BV/TV (24.6 ± 8.9%*) compared to control entire (C) and OVX (O), respectively. No significant changes to the control ewes (C) were registered for the OVX (O) or for the sheep treated with oestrogen only (OHE). Trabecular thickness was strongly increased by 13.05% in the oestrogen group (OHE) and by 22.02% in the combination treatment group (OHTE) compared to the thyroxine group (OHT) (Fig. 3g). Significance levels for trabecular thickness were missed due to inter-individual differences. However, changes in trabecular thickness were found to have influenced bone volume, most likely stronger than changes in trabecular numbers and separation.

μCT scans of the mandibular angle (ROI 2) for all six groups: control entire (=C) (a), O (b), OH (c), OHE (d), OHT (e), and OHTE (f). Bar graphs in part g show the mean ± standard deviation (SD) for bone volume per tissue volume (BV/TV), trabecular thickness (Tb.Th), trabecular number (Tb.N), trabecular spacing (Tb.Sp). The images of the mandibular angles are at about a 1.2 magnification. A significant decrease in BV/TV for OH and OHT compared to control entire is demonstrated in scans (parts c and e, respectively). Highest BV/TV values are registered for OHTE (part f). One-way ANOVA shows significances (--- significance when p < 0.05 or ___ when p < 0.01 level)
Cortical thickness in the mandibular angle (ROI 2) was analogous to the analyses from the diastema (ROI 1). The lowest levels were documented at 0.78 mm* for the thyroxine group (OHT) and the highest at 1.14 mm for the combination group (OHTE) (23.87% less and 9.70% more compared to control entire (C), respectively).
Static histomorphometry
Kossa-stained histologies from ROI 2 were analysed for all six groups (Fig. 4). Bone volume per tissue volume was the highest in control entire (C) (18.62 ± 3.38%) and OVX samples (O) (19.92 ± 0.27%). A significant decrease in BV/TV of 29.92%* and 29.18%* was observed in HPD sheep (OH) and the thyroxine group (OHT) when compared to control entire (C), respectively. Trabecular thickness increased with oestrogen treatment (OHE) by 6.20% and with the combined thyroxine and oestrogen treatment (OHTE) by 26.81% compared to control ewes (C) (Tb.Th 136.20 ± 10.61 mm). In contrast, trabecular thickness decreased by 13.15% in the HPD group (OH) and by 18.20% in the thyroxine-treated sheep (OHT).

Histological von Kossa-stained sections of the mandibular angle (ROI 2) from control entire—C (a), O (b), OH (c), OHE (d), OHT (e), and OHTE (f). Part g shows bar graphs with the mean ± standard deviation (SD) for bone volume per tissue volume (BV/TV), trabecular thickness (Tb.Th), trabecular number (Tb.N), trabecular spacing (Tb.Sp). BV/TV is significantly decreased in OH when compared with control entire (C) and OVX sheep (O). Tb.Th is significantly increased in OHTE sheep in comparison to OH and OHT groups (--- significance when p < 0.05 level)
Discussion
The present study confirmed the profound negative effect of HPD procedure on alveolar bone mass and structure. The three treatment arms presented interesting results, with oestrogen appearing to be bone protective by maintaining bone volume levels similar to control levels, and thyroxine causing a reduction of bone volume. Combined treatment with oestrogen and thyroxine demonstrated a prevention of bone loss with a tendency towards an increase in bone structure parameters compared to untreated control ewes.
In this current study, we specifically focused on two regions of the mandible (diastema and angle of the mandible) since other studies have placed their focus on these regions when analysing the osseous integration of dental implants [28, 29] and because our previous study analysed these two areas [15].
Alveolar bone is generally influenced by local and systemic factors. Systemic influence of the hypothalamus-pituitary axis on the (alveolar) bone is a complex mechanism regulating bone metabolism, specifically the activity of osteoblasts and osteoclasts for bone turnover [30]. Local parameters can be the mechanical load or inflammatory processes (e.g. periodontal disease). Mechanical loading is an important factor in maintaining good alveolar bone structure. However, here, the local factors were overruled by the HPD procedure leading to a significant decrease in alveolar bone quality in untreated HPD sheep. Furthermore, a systemic therapy was able to protect alveolar bone structure deterioration. Thus, according to our results, systemic influence has a significant effect on alveolar bone metabolism.
The HPD procedure induces a systemic hormonal imbalance, which has been shown to lead to bone loss in the main skeleton as well as in the alveolar bone, most likely due to a low-bone-turnover situation [14,15,16]. The paramount add-on in this current study on alveolar bone was made up off different treatment arms, particularly with oestrogen (OHE) or the combination of oestrogen and thyroxine (OHTE), showing astonishing results in protecting bone structure. This newly added hormonal replacement therapy showed that the surgically HPD-induced hormonal imbalance can be reversed by peripheral drug supplementation.
Oestrogen has been long known to have a stronger impact on inhibiting bone resorption than accelerating bone formation, thus leading to a steady or slightly increased bone mineral density [31, 32]. This positive effect of oestrogen works on a cellular level via the inhibition of osteoclast numbers and function with increased osteoblast recruitment and function [33]. This effect was confirmed by the present results for ewes treated with oestrogen demonstrating similar values for BV/TV and cortical thickness to untreated control ewes. On the contrast, thyroxine supplementation or hyperthyroidism induces a high-bone-turnover situation with increased bone resorption and formation [34, 35]. This supports the current results on why the surgically induced systemic hormonal imbalance in ewes and subsequent thyroxine supplementation do not prevent bone mass loss but rather lead to pronounced bone loss. However, the exact mechanism for thyroid hormones and bone metabolism is not yet fully understood [36]. Recently, an increase in bone turnover was demonstrated to be mediated by thyroxine-specific receptors on osteoclasts [37], whereas others identified TSH receptors on osteoblasts and osteoclasts influencing bone metabolism [35]. However, here, the regulatory system of TSH and thyroid hormones with its feedback mechanism has been disconnected due to the HPD procedure. Therefore, the thyroxine-induced bone loss is most likely explained by a peripheral thyroxine effect leading to a high-bone-turnover situation with the direct stimulation of osteoclast activity, respectively an increase in osteoclastic bone resorption [37].
Interestingly, the treatment arm with the combination of the two hormones (OHTE) showed synergistic effects, as reflected in improved bone structure parameters compared to untreated control ewes. The synergistic effect may be due to oestrogen inhibiting the bone resorptive effect of thyroxine and simultaneously the accelerated thyroxine induction of bone formation leading to a net bone gain [38]. However, this demonstrates the paramount importance of intercellular communication between osteoblasts and osteoclasts—so-called crosstalk—for balanced bone metabolism and consequently bone mass and structure.
The study has some limitations. First, the number of animals is limited. However, this large animal model with initial surgical interventions (OVX and HPD), long-term observation, and treatment demonstrates significant results regarding its impact on bone structure and quality. Second, this follow-up study focuses on mandibular bone only, whereas the systemic impact of the hormonal changes caused by the HPD procedure might be interesting to study also at other anatomic sites, such as the alveolar process of the maxilla. And lastly, the HPD sheep is doubtless a complex model causing systemic changes of different organ systems beyond mimicking low-turnover osteoporosis. In summary, further studies are needed in order to enhance the knowledge of the hormonal regulation of bone metabolism and structure to elucidate these complex interactions also on a cellular base.
Therefore, we can conclude that (i) alveolar bone significantly underlies the systemic regulatory system of the hypothalamus and pituitary in addition to local factors; (ii) the HPD procedure leads to a significant bone loss of the alveolar bone in sheep; (iii) peripheral hormone substitution overrules centrally induced bone loss; (iv) thyroxine alone has a negative effect on bone metabolism, most likely due to a direct activation of osteoclastic bone resorption; and (v) hormonal regulation of bone metabolism in general and the intercellular communication of osteoblasts and osteoclasts in particular is essential for a balanced bone turnover, respectively bone mass and structure.
References
Rawlinson SC (2009) Ovariectomy vs. hypofunction: their effects on rat mandibular bone. J Dent Res 88:615–620. doi:10.1177/0022034509340132
Sanfilippo F, Bianchi AE (2003) Osteoporosis: the effect on maxillary bone resorption and therapeutic possibilities by means of implant prostheses—a literature review and clinical considerations. Int J Periodontics Restorative Dent 23:447–457
Tatakis DN, Kumar PS (2005) Etiology and pathogenesis of periodontal diseases. Dent Clin North Am 49:491-516, v. doi:10.1016/j.cden.2005.03.001
Kribbs PJ, Chesnut CH 3rd, Ott SM, Kilcoyne RF (1989) Relationships between mandibular and skeletal bone in an osteoporotic population. J Prosthet Dent 62:703–707
Krall EA, Garcia RI, Dawson-Hughes B (1996) Increased risk of tooth loss is related to bone loss at the whole body, hip, and spine. Calcif Tissue Int 59:433–437
Jonasson G, Jonasson L, Kiliaridis S (2006) Changes in the radiographic characteristics of the mandibular alveolar process in dentate women with varying bone mineral density: a 5-year prospective study. Bone 38:714–721. doi:10.1016/j.bone.2005.10.008
Oz HS, Puleo DA (2011) Animal models for periodontal disease. J Biomed Biotechnol 2011:754857. doi:10.1155/2011/754857
Yamashiro T, Takano-Yamamoto T (2001) Influences of ovariectomy on experimental tooth movement in the rat. J Dent Res 80:1858–1861
Tanaka M, Ejiri S, Toyooka E, Kohno S, Ozawa H (2002) Effects of ovariectomy on trabecular structures of rat alveolar bone. J Periodontal Res 37:161–165
Tanaka M, Toyooka E, Kohno S, Ozawa H, Ejiri S (2003) Long-term changes in trabecular structure of aged rat alveolar bone after ovariectomy. Oral Surg Oral Med Oral Pathol Oral Radiol Endod 95:495–502. doi:10.1067/moe.2003.135
Rawlinson SC, Boyde A, Davis GR, Howell PG, Hughes FJ, Kingsmill VJ (2009) Ovariectomy vs. hypofunction: their effects on rat mandibular bone. J Dent Res 88:615–620. doi:10.1177/0022034509340132
Dvorak G, Gruber R, Huber CD, Goldhahn J, Zanoni G, Salaberger D, Watzek G, Haas R (2008) Trabecular bone structures in the edentulous diastema of osteoporotic sheep. J Dent Res 87:866–870
Dvorak G, Reich KM, Tangl S, Goldhahn J, Haas R, Gruber R (2011) Cortical porosity of the mandible in an osteoporotic sheep model. Clin Oral Implants Res 22:500–505. doi:10.1111/j.1600-0501.2010.02031.x
Beil FT, Oheim R, Barvencik F, Hissnauer TN, Pestka JM, Ignatius A, Rueger JM, Schinke T, Clarke IJ, Amling M, Pogoda P (2012) Low turnover osteoporosis in sheep induced by hypothalamic-pituitary disconnection. J Orthop Res 30:1254–1262. doi:10.1002/jor.22066
Oheim R, Beil FT, Krause M, Bindl R, Ignatius A, Pogoda P (2013) Mandibular bone loss in ewe induced by hypothalamic-pituitary disconnection. Clin Oral Implants Res 25:1239–1244. doi:10.1111/clr.12259
Oheim R, Beil FT, Kohne T, Wehner T, Barvencik F, Ignatius A, Amling M, Clarke IJ, Pogoda P (2013) Sheep model for osteoporosis: sustainability and biomechanical relevance of low turnover osteoporosis induced by hypothalamic-pituitary disconnection. J Orthop Res 31:1067–1074. doi:10.1002/jor.22327
Oheim R, Schinke T, Amling M, Pogoda P (2016) Can we induce osteoporosis in animals comparable to the human situation? Injury 47(Suppl 1):S3–S9. doi:10.1016/S0020-1383(16)30002-X
Clarke IJ, Cummins JT, de Kretser DM (1983) Pituitary gland function after disconnection from direct hypothalamic influences in the sheep. Neuroendocrinology 36:376–384
Prickett TC, Barrell GK, Wellby M, Yandle TG, Richards AM, Espiner EA (2008) Effect of sex steroids on plasma C-type natriuretic peptide forms: stimulation by oestradiol in lambs and adult sheep. J Endocrinol 199:481–487. doi:10.1677/JOE-08-0267
Billings HJ, Viguie C, Karsch FJ, Goodman RL, Connors JM, Anderson GM (2002) Temporal requirements of thyroid hormones for seasonal changes in LH secretion. Endocrinology 143:2618–2625
Amling M, Priemel M, Holzmann T, Chapin K, Rueger JM, Baron R, Demay MB (1999) Rescue of the skeletal phenotype of vitamin D receptor-ablated mice in the setting of normal mineral ion homeostasis: formal histomorphometric and biomechanical analyses. Endocrinology 140:4982–4987
Oheim R, Amling M, Ignatius A, Pogoda P (2012) Large animal model for osteoporosis in humans: the ewe. Eur Cell Mater 24:372–385
Pogoda P, Egermann M, Schnell JC, Priemel M, Schilling AF, Alini M, Schinke T, Rueger JM, Schneider E, Clarke I, Amling M (2006) Leptin inhibits bone formation not only in rodents, but also in sheep. J Bone Miner Res 21:1591–1599. doi:10.1359/jbmr.060709
Parfitt AM, Drezner MK, Glorieux FH, Kanis JA, Malluche H, Meunier PJ, Ott SM, Recker RR (1987) Bone histomorphometry: standardization of nomenclature, symbols, and units. Report of the ASBMR Histomorphometry Nomenclature Committee J Bone Miner Res 2:595–610. doi:10.1002/jbmr.5650020617
Dempster DW, Compston JE, Drezner MK, Glorieux FH, Kanis JA, Malluche H, Meunier PJ, Ott SM, Recker RR, Parfitt AM (2013) Standardized nomenclature, symbols, and units for bone histomorphometry: a 2012 update of the report of the ASBMR Histomorphometry Nomenclature Committee. J Bone Miner Res 28:2–17. doi:10.1002/jbmr.1805
Oheim R, Simon MJ, Steiner M, Vettorazzi E, Barvencik F, Ignatius A, Amling M, Clarke IJ, Pogoda P, Beil FT (2017) Sheep model for osteoporosis: the effects of peripheral hormone therapy on centrally induced systemic bone loss in an osteoporotic sheep model. Injury. doi:10.1016/j.injury.2017.02.009
Turner AS, Mallinckrodt CH, Alvis MR, Bryant HU (1995) Dose-response effects of estradiol implants on bone mineral density in ovariectomized ewes. Bone 17:421S–427S
Duncan WJ, Lee MH, Bae TS, Lee SJ, Gay J, Loch C (2015) Anodisation increases integration of unloaded titanium implants in sheep mandible. Biomed Res Int 2015:857969. doi:10.1155/2015/857969
Trisi P, Berardini M, Falco A, Podaliri Vulpiani M, Perfetti G (2014) Insufficient irrigation induces peri-implant bone resorption: an in vivo histologic analysis in sheep. Clin Oral Implants Res 25:696–701. doi:10.1111/clr.12127
Driessler F, Baldock PA (2010) Hypothalamic regulation of bone. J Mol Endocrinol 45:175–181. doi:10.1677/JME-10-0015
Raisz LG (1999) Physiology and pathophysiology of bone remodeling. Clin Chem 45:1353–1358
Pacifici R (1998) Cytokines, estrogen, and postmenopausal osteoporosis—the second decade. Endocrinology 139:2659–2661. doi:10.1210/endo.139.6.6087
Khosla S, Oursler MJ, Monroe DG (2012) Estrogen and the skeleton. Trends Endocrinol Metab 23:576–581. doi:10.1016/j.tem.2012.03.008
De Rosa G, Testa A, Giacomini D, Carrozza C, Astazi P, Caradonna P (1997) Prospective study of bone loss in pre- and post-menopausal women on L-thyroxine therapy for non-toxic goitre. Clin Endocrinol 47:529–535
Abe E, Marians RC, Yu W, Wu XB, Ando T, Li Y, Iqbal J, Eldeiry L, Rajendren G, Blair HC, Davies TF, Zaidi M (2003) TSH is a negative regulator of skeletal remodeling. Cell 115:151–162
Nicholls JJ, Brassill MJ, Williams GR, Bassett JH (2012) The skeletal consequences of thyrotoxicosis. J Endocrinol 213:209–221. doi:10.1530/JOE-12-0059
Monfoulet LE, Rabier B, Dacquin R, Anginot A, Photsavang J, Jurdic P, Vico L, Malaval L, Chassande O (2011) Thyroid hormone receptor beta mediates thyroid hormone effects on bone remodeling and bone mass. J Bone Miner Res 26:2036–2044. doi:10.1002/jbmr.432
Heino TJ, Hentunen TA, Vaananen HK (2002) Osteocytes inhibit osteoclastic bone resorption through transforming growth factor-beta: enhancement by estrogen. J Cell Biochem 85:185–197
Acknowledgements
The authors appreciate the tremendous help and technical assistance of Mr. Bruce Doughton and Ms. Lynda Morrish and would like to thank them, as well as Ms. Marion Dietzmann, for her help in histological processing. This work was supported by the German Research Foundation (DFG) within the framework of the DFG Research Group 793 to PP and MA. FTB, MJKS, and RO are fellows of the German Research Foundation (DFG AM 103/13-1 and DFG AM 103/14-1).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Funding
This work was supported by the German Research Foundation (DFG) within the framework of the DFG Research Group 793 to PP and MA. FTB, MJKS, and RO are fellows of the German Research Foundation (DFG AM 103/13-1 and DFG AM 103/14-1).
Ethical approval
All applicable national and institutional guidelines for the care and use of animals were followed. All procedures and tissue collections were in accordance with the guidelines for the Care and Maintenance of Experimental Animals and with the prior approval of the Monash Medical Centre Animal Ethics Committee and the Animal Experimentation Ethics Committee of Victorian Institute of Animal Science, Victoria, Australia.
Rights and permissions
About this article

Cite this article
Simon, M.J.K., Beil, F.T., Pogoda, P. et al. Is centrally induced alveolar bone loss in a large animal model preventable by peripheral hormone substitution?. Clin Oral Invest 22, 495–503 (2018). https://doi.org/10.1007/s00784-017-2138-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00784-017-2138-7