
Abstract
In this paper we review the fragmentary fossil pollen record of Hamamelidaceae which extends back to the middle Eocene until the Early Pleistocene in the Iberian Peninsula. Records of fossil palynomorphs related to Hamamelidaceae are very scarce in the European Cenozoic, in part due to difficulty in confident identification at a generic or even familial rank. The present review contributes to improving the accuracy of identification of Cenozoic Hamamelidaceae in standard palynological studies, which should help trace its poorly understood fossil and evolutionary history. The review of all published Iberian records indicates a considerable generic diversity. At least four genera can be confidently identified. The examination of new material from the Oligocene of north-west Spain has led to the formal description of three new fossil pollen genera that are related to extant genera currently restricted geographically to Asia: Disanthuspollenites, Embolantherapollenites and Parrotiapollenites. Based on SEM analysis, a new combination of the fossil species Tricolpopollenites indeterminatus, traditionally used to designate fossil pollen related to Hamamelidaceae in Europe, is proposed. Additionally, we discuss the occurrence of pollen forms that can be tentatively attributed to Corylopsis and Fothergilla.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
According to the most recent classifications, Hamamelidaceae comprises about 100 species distributed in 27 genera and three subfamilies: Exbucklandioideae, Rhodoleioideae and Hamamelidoideae (Endress 1989b, 1993). The Altingioideae, which contain Liquidambar, were traditionally included within Hamamelidaceae, but they are now regarded as an independent family (Magallón 2007; APG III 2009). Members of Hamamelidaceae are widely distributed across the world, including tropical Africa, South Africa, Madagascar, the Middle East region, Asia, New Guinea, north-east Australia, eastern North America and Central America (Takhtajan 2009). However, most species are represented by scattered and relict occurrences (Endress 1993). At present day, Europe is devoid of members of Hamamelidaceae and the main diversity centre is Asia.
The oldest fossil records of Hamamelidaceae are floral remains from the Late Cretaceous (late Santonian) of Sweden and Georgia, USA (Crepet et al. 1992; Magallón-Puebla et al. 1996; Magallón et al. 2001; Friis et al. 2011). These mesofossils are attributed to the extinct genera Allonia and Androdecidua. Morphological features characteristic of the present-day genera first occur in the Maastrichtian of Germany, where Knobloch and Mai (1986) found seeds in clusters similar to extant Rhodoleia. However, it is not until the Eocene that present-day genera become generalized in the fossil record (Manchester 1994; Radtke et al. 2005; Benedict et al. 2008).
Reliable records of dispersed Hamamelidaceae pollen are scarce. It has proved difficult to confidently ascribe dispersed Cretaceous palynomorphs (not preserved in anatomical connection with floral remains) to Hamamelidaceae as they show features that are also similar to extant Platanaceae, a family widely distributed in the Northern Hemisphere during the Late Cretaceous (Crepet et al. 1992; Denk and Tekleva 2006). Fossil palynomorphs unequivocally related to Hamamelidaceae are not reported until the Paleogene. In the European Cenozoic the first records of individual genera are from the Eocene, although further evidence would be needed for a more confident assignment of their botanical affinity at a generic level (Muller 1981). Thus, various authors report fossil species such as Corylopsispollenites microreticulatus, Retitricolpites henisensis and Tricolpites compactus (Gruas-Cavagnetto 1978; Châteauneuf 1980; Kedves 1998; Worobiec and Gedl 2018), possibly related to extant Fothergilla and Corylopsis. In other cases, identification is made at family level only, and records are not well documented with images or descriptions of the specimens, which renders them less valuable. In the Neogene, Hamamelidaceae pollen become more common. In Eastern Europe, the species Tricolporopollenites indeterminatus and Tricolporopollenites staresedloensis, both of uncertain generic affinity within Hamamelidaceae, have been recorded throughout the Neogene (Stuchlik et al. 2014). During the Quaternary, the extensive glaciations that took place in mid-latitude regions led to the extinction of Hamamelidaceae in Europe (Zhang and Lu 1995).
The scarcity of fossil pollen records of individual hamamelidaceous genera is partly due to the difficulty in identifying palynomorphs at a generic level using standard light microscopy (LM) counting techniques. Scanning electron microscopy (SEM) analysis on Hamamelidaceae pollen is very useful in obtaining a higher taxonomic resolution, as has been shown by Grímsson et al. (2015a). However, despite SEM analysis being widely used nowadays, it is not always a time/cost effective technique to be used routinely. While it is true that many genera within Hamamelidaceae produce pollen with similar morphological features which hamper reliable identification at a generic level, some genera are recognizable without the need for SEM examination. Therefore, unlike foliar and seed macroremains, which are virtually impossible to separate at a generic rank (Tiffney 1986; Mai 1995), some hamamelidaceous pollen do allow precise identification. This fact confers them significant, although mostly untapped, potential to provide insights on the poorly understood fossil history and biodiversity of the family during the Cenozoic.
This paper aims to contribute to improving the accuracy in the identification of Cenozoic Hamamelidaceae fossil pollen by describing in detail six pollen types related to this family. Three pollen types are new genera that occur consistently in the Iberian Peninsula. We discuss their botanical affinity and confirmed stratigraphic ranges. Additionally, we review the records of Hamamelidaceae reported in the literature which, although not always documented with micrographs, suggest a higher past biodiversity.
Palaeogeographical and geological context
The distribution and composition of Iberian vegetation throughout the Cenozoic was mainly controlled by the climate and the palaeogeographic and tectonic processes that gave rise to the major sedimentary basins. An overview of the evolution of Cenozoic vegetation in the Iberian Peninsula is summarized in Barrón et al. (2010).
Throughout the Late Cretaceous–early Eocene, the area that at present day forms the Iberian Peninsula comprised two emerged landmasses located in its western half (Hesperian Massif) and in north-east Spain (Ebro and Catalan Massifs) (López-Martínez 1989). The latter islands became connected with each other and with the Central European emerged land over the course of the Eocene, remaining as a single geographic unit until the present day (Meulenkamp and Sissingh 2003).
Broadly, the Cenozoic geological evolution of the Iberian Peninsula was primarily driven by the convergence of the Iberian and European plates. During the Cenozoic the Iberian plate motion had two main components: a lateral movement towards the east and an anticlockwise rotation which led to strong compressional deformation during most of the Paleogene and, to a lesser extent, in part of the Miocene. This plate movement resulted in the formation of the major mountain ranges (Pyrenees, Cantabrian, Iberian and Betic) (Sanz de Galdeano 1996; Alonso-Zarza et al. 2002).
Deposition during the Cenozoic was extensive in both the inner part of the peninsula and in coastal regions. Cenozoic basins cover a large area, spreading across the territory, and comprising a broad variety of basins in terms of size and origin (Civis 2004). Five major Cenozoic basins were formed: the Ebro, Duero, Guadalquivir, Lower Tagus and Tagus. The Ebro Basin originated in the late Palaeocene–Eocene as a foreland depression associated with the uplift of the Pyrenees and the Iberian Range (Fig. 1a). It was connected to the Atlantic during the early–middle Eocene; however, it lost its marine connection in the late Eocene following a significant regression. From the late Eocene to the late Miocene, the Ebro Basin was endorheic and fluvial/alluvial sedimentation dominated in its margins, while a lacustrine system developed in its central part (Anadón et al. 1985; Pardo et al. 2004). In the late Miocene, various rivers started to act as drainage for the lacustrine system, connecting it to the Mediterranean. The Duero and Tagus Basins (Fig. 1b, c) are two large intraplate depressions separated by a central horst (the Central System mountain range) (Alonso-Zarza et al. 2002). Both basins occupy most of the northern and southern Iberian plateaus in central Spain. During the Neogene, they became endorheic basins dominated by lacustrine depositional settings that led to carbonate, siliciclastic and evaporite deposition. The Lower Tagus Basin in Portugal (Fig. 1d) was an endorheic basin in the Paleogene, becoming connected to the Atlantic in the early Miocene (Pais 2004). The Neogene is characterized by marine deposits in the distal part of the basin, while in more proximal parts a mixture of brackish and continental deposition prevailed. The Pliocene is eminently fluvial. The Guadalquivir Basin is the youngest of the major Cenozoic basins (Fig. 1e). This foreland depression, linked to the uplift of the Betic Range, formed in the middle–late Miocene and comprises primary marine deposits. Originally, it was part of the North Betic Strait which, together with the South Rif Strait in Morocco, was the main seaway connecting the Atlantic and Mediterranean realms. In the mid–late Tortonian, the connection with the Mediterranean ceased and the Guadalquivir Basin became linked only to the Atlantic (Civis 2004).

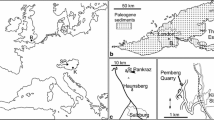
Major Cenozoic Iberian basins and geographic location of the sites that have yielded hamamelidaceous palynomorphs. a Ebro Basin, b Duero Basin, c Tagus Basin, d Lower Tagus Basin, e Guadalquivir Basin. 1 Sarral, Ebro Basin: late Eocene (Cavagnetto and Anadón 1996); 2 S. Coloma de Queralt, Ebro Basin: middle Eocene–Oligocene (Cavagnetto and Anadón 1996); 3 As Pontes Basin: early Oligocene–early Miocene (Casas-Gallego 2018); 4 Izarra: early Miocene (Barrón et al. 2006a); 5 Rubielos de Mora: early Miocene (Barrón et al. 2006b; Jiménez-Moreno et al. 2007); 6 Tagus Basin: early Miocene (Pais 1986); 7 Arcas del Villar: middle Miocene (Gaudant et al. 2015); 8 Duero Basin: middle Miocene (Rivas Carballo 1991; Rivas Carballo et al. 1994; Valle et al. 1995); 9 La Cerdanya Basin: late Miocene (Barrón et al. 2016); 10 Venta del Moro: late Miocene (Van Campo 1989; Casas-Gallego et al. 2015); 11 San Onofre: Pliocene (Bessais and Cravatte 1988); 12 Barcelona area: Pliocene (Suc and Cravatte 1982); 13 Baza Basin: Pleistocene (Altolaguirre et al. 2020)
In addition to the above major basins, multiple smaller depressions are present, such as the Neogene basins in the eastern Iberian range (Fig. 1, locations 5 and 10), various Neogene intramontane basins in the Pyrenees (Fig. 1, location 9), and the Galician interior basins (Fig. 1, location 3) associated with strike-slip faults. Because the Cenozoic Iberian basins are mostly continental, their age determination often relies on micromammals. In the absence of marine fossils, a more solid knowledge of the terrestrial microfloras is believed to have the potential for complementing mammals in age determination and intra and inter-basin correlations.
Material and methods
In order to assemble the current knowledge of the Hamamelidaceae fossil record in the Iberian Peninsula, we have reviewed all published palaeobotanical works that report the occurrence of the family. To achieve this, an extensive review of palaeobotanical records in the region produced by Postigo-Mijarra et al. (2009) was a mandatory starting point. The area that concentrates more records of Hamamelidaceae pollen is the eastern part of the peninsula, close to the Mediterranean coast [Fig. 1, e.g. Suc (1980), Jiménez Moreno (2005), Bessais and Cravatte (1988)]. In addition to the records reported in the regional literature, the palynological assemblages from the early Miocene of Rubielos de Mora, middle Miocene of Arcas del Villar and late Miocene of Venta del Moro and La Cerdanya, all housed within the collections of the Spanish Geological Survey in Madrid, have been re-examined for this study, paying special attention to the occurrence of Hamamelidaceae.
Moreover, the palynoflora of the Oligocene–early Miocene As Pontes Basin, which is exceptionally well preserved in lignite and clays rich in organic matter, has also been re-examined (Casas-Gallego 2018). The taxonomic descriptions provided in this work derive from specimens recovered from this basin. Since the 1960s, multiple palynological investigations in As Pontes have resulted in the identification of more than 300 palynomorph species (Médus and Nonn 1963; Médus 1965; Cavagnetto 2002; Casas-Gallego 2018). The re-examination of the material has led to the discovery of new pollen forms displaying hamamelidaceous affinity that have not previously been documented.
The palynological slides were studied using LM and SEM. For the SEM analysis, we used the single grain technique, initially proposed by Zetter (1989) and subsequently used successfully and improved in various studies (Grímsson et al. 2011, 2015b; Denk et al. 2012; Kmenta and Zetter 2013). A small portion of the organic residue obtained after the palynological processing was transferred onto a glass slide and mixed with glycerine. Using a × 20 microscope objective, specific palynomorphs of interest for this work were targeted. The relatively high density of the glycerine facilitates the collection of selected palynomorphs using the tip of a dissecting needle. Picked palynomorphs were transferred to a metal SEM stub and thoroughly rinsed with absolute ethanol to dissolve the glycerine. At that stage, the samples were air-dried and carbon and gold-coated to enable examination using SEM. The morphologies of the fossil specimens were compared with those of extant pollen of Hamamelidaceae. Various authors have described the pollen of specific extant genera (Fritz and Allesch 1999; Zhang 2001), although the most comprehensive review of Hamamelidaceae pollen is that of Bogle and Philbrick (1980), which documents the morphology of 26 of the 27 currently accepted genera from good quality SEM images.
The intrafamilial classification of Hamamelidaceae used in this study is based on Endress (1989b). The suprafamilial classification is that proposed by the Angiosperm Phylogeny Group (APG III 2009). The terminology used to describe the palynomorphs is based on Punt et al. (2007). The slides with the holotypes are lodged in the collections of the Geominero Museum of the Geological Survey of Spain (IGME) in Madrid.
Pollen record of Hamamelidaceae in the Iberian Peninsula
To date, 14 hamamelidaceous genera have been reported from fossil pollen in the Iberian Peninsula (Fig. 5). However, among them only six have been comprehensively described and/or documented with images: Disanthuspollenites, Embolantherapollenites, Parrotiapollenites, Tricolpopollenites indeterminatus, cf. Corylopsispollenites and cf. Fothergilla. These have all been reviewed in the section ‘Systematic paleontology’.
There is no evidence of Hamamelidaceae pollen in the Palaeocene of the Iberian Peninsula, although this might be due to the limited study of these deposits in the region, partly because they crop out sparsely. The absence of Hamamelidaceae pollen in the Iberian Palaeocene is in line with the microfloral record for the Northern Hemisphere, which shows that pre-Eocene specimens potentially related to the family lack features that can undisputedly confirm such relationship (Muller 1981; Zetter et al. 2011). The first pollen records of the family in the Iberian Peninsula are from the middle Eocene (Bartonian) of the Ebro Basin. In particular, Cavagnetto and Anadón (1996) reported the occurrence of Fothergilla, Corylopsis and Loropetalum from the Bartonian of Igualada (eastern Ebro Basin). The first occurrences of hamamelidaceous pollen related to extant genera in Europe have a similar (Bartonian and Priabonian) age (Gruas-Cavagnetto 1978; Châteauneuf 1980; Worobiec and Gedl 2018). Despite the fact that the Eocene specimens reported by Cavagnetto and Anadón (1996) as supposedly related to Fothergilla, Corylopsis and Loropetalum have not been figured and need further confirmation, they are valuable because they clearly indicate that pollen produced by Hamamelidaceae was diverse by the middle Eocene, and also because they represent the oldest Iberian palaeobotanical remains ascribed to the family.
It seems that the generic diversity increased significantly during the Oligocene (Fig. 5). The presence of specimens tentatively related to Fothergilla and Corylopsis have been confirmed in the Rupelian and Chattian, respectively (Figs. 3k–n, 4k–n). In the Oligocene we also found the oldest confident occurrences of the ancestors of extant Disanthus, Parrotia and Embolanthera (Figs. 2 and 3). Some of the Oligocene records are controversial, such as Eustigma, Hamamelis, Parrotiopsis and Sinowilsonia. Palynomorphs assigned to these genera have been reported from Oligocene deposits in north-west and north-east Spain (Cavagnetto and Anadón 1996; Cavagnetto 2002), but no images or illustrations were provided to support such identifications. Because palynological studies often report dozens to hundreds of taxa based on the examination of thousands of palynomorphs, palynologists cannot possibly document every taxon with images. It is therefore understandable that for well-known, formally described species it is not necessary to photograph these in every study. However, when dealing with specimens that have not been documented previously, a detailed description becomes imperative until there is a general consensus as to the botanical affinity of the palynomorph. It is questionable whether Paleogene fossil pollen can be assigned to certain extant genera such as Eustigma, Hamamelis, Parrotiopsis and Sinowilsonia without the use of SEM. Grímsson et al. (2015a) showed that the identification of various hamamelidaceous genera such as Fortunearia and Distylium must rely on SEM analysis, as the features observable under LM are too similar to closely related genera to allow for differentiation (Bogle and Philbrick 1980). In some cases, even the SEM study might not be enough to infer the botanical affiliation of the fossil palynomorph, as in the case of Tricolpopollenites indeterminatus (Fig. 4a–j).

a–k Specimens of Embolantherapollenites calvicolpatus sp. nov. from the Chattian of the As Pontes Basin. a SEM image showing the structure of the reticulum and colpus margin, b–c and g equatorial view at high, medium, and low focus, respectively, d–f holotype in polar view at high, medium, and low focus, respectively, h–k polar view of two different specimens at high and low focus. l–o Specimens of Disanthuspollenites operculatus sp. nov, l holotype in polar view, Chattian of As Pontes Basin, m polar view, Burdigalian of Rubielos de Mora, n–o polar view, Messinian of Venta del Moro. The scale bar represents 20 µm, unless otherwise indicated

a–j Specimens of Parrotiapollenites asper sp. nov. a–b holotype at high and low focus, Chattian of As Pontes Basin, c–d specimen in polar view from the Chattian of As Pontes at high and low focus, e–f SEM images showing in detail the microbacula-bearing reticulum and microgranulate colpi, g–h two different specimens from the Burdigalian of Rubielos de Mora, i–j two specimens from the Chattian of As Pontes. k–n Specimens tentatively attributed to Corylopsispollenites E.Worobiec from the Chattian of As Pontes. k–l Polar view, m–n tangential, nearly equatorial, view. The scale bar represents 20 µm, unless otherwise indicated

a–j specimens of Tricolpopollenites indeterminatus comb. nov. from the Chattian and Rupelian of As Pontes Basin. a–d Three specimens in polar view, e–f SEM images showing in detail the reticulate exine, g–h proposed neotype in equatorial view at high and low focus, respectively, i–j specimen in equatorial view at high and low focus. k–n Two specimens tentatively attributed to Fothergilla from the Chattian of As Pontes. Note the distinctive reduction in size of lumina in the central part of the polar areas. The scale bar represents 20 µm, unless otherwise indicated
The confirmed record of Embolantherapollenites is so far restricted to the Oligocene, and those of T. indeterminatus, cf. Corylopsispollenites sp. and cf. Fothergilla sp. extend until the early Aquitanian. Despite the last confirmed occurrences of the latter taxa, high generic diversity continued until the middle Miocene. The first occurrences of Distylium, Exbucklandia and Rhodoleia are reported from lower and middle Miocene deposits (Fig. 5) (Jiménez-Moreno et al. 2007, 2010). Again, the records of Distylium and Exbucklandia must be taken cautiously, as more detailed work is needed to conclusively link fossil pollen with those extant genera but attest to the co-occurrence of various hamamelidaceous forms in the early and middle Miocene. Pollen produced by living Rhodoleia is distinctive, as it possesses a very fine perforate exine (Bogle and Philbrick 1980), and could therefore be one of the hamamelidaceous genera that can be identified without the need for SEM, although its presence in the Miocene of the Iberian Peninsula requires documentation.

Known stratigraphic range of hamamelidaceous palynomorphs in the Iberian Peninsula. The solid line represents confirmed records described in this study. Reported records that need further confirmation (not sufficiently documented with images or descriptions) are represented with a dotted line
Among the confirmed records, palynomorphs related to extant Disanthus and Parrotia are the best represented in the Iberian Cenozoic. The occurrence of Disanthuspollenites throughout the Miocene has been shown through various studies (Jiménez Moreno 2005; Casas-Gallego et al. 2015; Gaudant et al. 2015). In the Pliocene, the only confirmed occurrence is that of Parrotiapollenites, for which there is micro- and macrofossil evidence (Roiron 1983; Bessais and Cravatte 1988; Leroy 1997; Jiménez-Moreno et al. 2013). Parrotia persisted in the Iberian Peninsula until the Early Pleistocene (Julia Bruguès and Suc 1980; González-Sampériz et al. 2010; Postigo-Mijarra et al. 2010; Altolaguirre et al. 2019). Parrotia also seems to have become extinct in the Early Pleistocene (Calabrian) in other southern European regions (Italy and Greece) (Magri et al. 2017).
Climatic and ecological significance of fossil Hamamelidaceae
Palaeoclimatic and palaeoecological inferences based on fossil pollen mostly rely on three assumptions associated with the concept of a nearest living relative (NLR) (Utescher et al. 2014):
-
1.
It is assumed that for some fossil taxa it is possible to assign a NLR based on the degree of morphological similarity between the fossil and the extant taxa. Thus, it is accepted that similar fossil and extant morphologies reflect systematic closeness.
-
2.
The principle of physiological actualism or uniformitarianism assumes that similar morphologies also reflect similar genotypes and, ultimately, a comparable set of ecological and climatic tolerances (Tiffney 2008).
-
3.
The current geographic distribution of plants reflects with enough accuracy the climatic ranges that suit them best.
The above assumptions pose some difficulties when trying to interpret the palaeoclimatic significance of Hamamelidaceae. Firstly, as shown in the systematic palaeontology section, the NLR of some fossil hamamelidaceous taxa cannot be assigned confidently, such as Tricolpopollenites indeterminatus and the palynomorphs tentatively ascribed to Corylopsis and Fothergilla. Secondly, some present-day genera within Hamamelidaceae are monotypic or have only two species, displaying a relict geographic distribution, like Disanthus, Embolanthera, Fothergilla and Parrotia (Endress 1993). These genera are likely to have comprised a higher number of species and a wider geographic distribution throughout the Cenozoic. Therefore, any palaeoclimatic inference derived from fossil Hamamelidaceae must be taken cautiously.
Current members of Hamamelidaceae have a wide geographic distribution, living in areas within a broad variety of climates and ecosystems, ranging from tropical rainforests to mid-altitude mountains in regions with temperate climate. This pattern is the result of a worldwide spread during the Cretaceous and the Paleogene (Zhang and Lu 1995).
Extant Disanthus includes a single species of deciduous shrubs which occurs only in eastern China and Japan. Studies on the ecological requirements of Disanthus are lacking, but its foliar strategy and current habitat indicate a preference for deciduous and mixed evergreen-deciduous broadleaved forests (Zhang et al. 2003). The relatively broad stratigraphic range of Disanthuspollenites (from Oligocene to upper Miocene) suggests a considerable ability to adapt to different climatic and ecological regimes. Probably the plant that produced Disanthuspollenites formed part of primary evergreen forests characteristic of north-west Iberia during the Oligocene (Casas-Gallego 2018), and became able to thrive in warm temperate forests dominated by mesophyllous plants during most of the Neogene. The fossil record also suggests that the parent plant of Disanthuspollenites persisted during the drier periods of the late Miocene, possibly associated with azonal local wetter conditions linked to riparian habitats. Furthermore, the changes in abundance of Disanthuspollenites in the Messinian succession of Venta del Moro (eastern Spain) indicates that it was not able to tolerate seasonally flooded soils. In the latter locality, a shallow freshwater marsh developed during the Messinian. It has been shown that an increase in the water level of the wetland resulted in a drastic replacement of Disanthus by Acer (Casas-Gallego et al. 2015).
At present, Embolanthera comprises two species: E. glabrescens, which is known only from north Vietnam, and E. spicata, which is endemic of the Philippines (Endress 1993). Both are evergreen trees that live in humid subtropical (Cwa in the Köppen-Geiger climatic classification) and tropical rainforest (Af) climates, respectively. This geographic distribution indicates that Embolanthera species are among the most thermophilous members of Hamamelidaceae. This thermic affinity accounts for its occurrence in the evergreen broadleaved forests that developed in north-western Iberia during the Rupelian and early Chattian (Casas-Gallego and Barrón 2020).
Parrotia is a deciduous tree currently represented by two species. P. persica occurs only in the Caspian Hyrcanian mixed forests of northern Iran and Azerbaijan, where it shares habitat with other relict species such as Gleditsia caspica, Zelkova carpinifolia and Pterocarya fraxinifolia, at the lower altitudes of montane and lowland forests (Endress 1993; Akhani et al. 2010). The Caspian forest has been envisaged by Suc et al. (1995) as a potential modern analogue of the vegetation that dominated large areas of the Mediterranean contour during the late Miocene and Pliocene. In fact, despite Parrotia fossil leaves being hard to differentiate from closely similar genera, like Fothergilla (Mai 1995), the rich reported macrofossil record of Parrotia suggests that it was widespread in Europe during the Miocene and Pliocene (Leroy and Roiron 1996; Zidianakis et al. 2007), when it attained its maximum geographic extent and abundance. The second species is P. subaequalis, a deciduous tree endemic to eastern China that has just a few populations in subtropical montane forests, where it co-occurs with occasional evergreen broadleaved taxa such as Cyclobalanopsis glauca (Li and Zhang 2015).
In the Cenozoic of the Iberian Peninsula, Parrotiapollenites is the best represented hamamelidaceous fossil pollen, with a stratigraphic range from the lower Oligocene to the lower Pliocene. Likewise, the European macrofossil record of Parrotia indicates that it occurs consistently from the Oligocene attaining a peak in abundance in the Mio-Pliocene (Mai 1995). This pattern suggests a preference for warm temperate climatic regimes characteristic of the Neogene and Pliocene. It is possible that the parent plant that produced the specimens of Parrotiapollenites from the Oligocene of northern Spain had more similar climatic requirements to extant P. subaequalis.
Corylopsis currently comprises at least 29 species of mainly deciduous shrubs or small trees in south-eastern China, India, Japan and Korea, where they usually grow in montane forests under warm temperate conditions. A few species are semi-evergreen and thrive under subtropical climates at lower altitudes (Zhang et al. 2003). Based on its current distribution and the vegetation of which it forms part, the climatic preference of Corylopsis seems to be similar to that of Disanthus or slightly better adapted to warmer conditions. Corylopsis is primary entomophilous, and therefore, it produces little pollen. Even so, the specimens of Corylopsispollenites are encountered in very low numbers suggesting that the role of this plant in the vegetation was limited.
The current species of Fothergilla are deciduous shrubs native to south-eastern North America, where they grow in woodlands of low-altitude mountains and lowland swamp settings, tolerating a broad range of temperature (Ranney and Lynch 2007). It is likely that Paleogene and Neogene Fothergilla inhabited a similar range of environments, as a mosaic of vegetation types functionally similar to those of south-eastern USA today, including swamps, are known to have been widespread in Europe throughout most of the Cenozoic (Mai 1995; Collinson and Hooker 2003).
Conclusions
The revision of the reported occurrences of fossil Hamamelidaceae pollen in the Iberian Peninsula shows a significant generic diversity of the family from the middle Eocene until the Early Pleistocene. Four genera can be confidently identified, including the first evidence of Embolanthera in Europe, and two more are described as potentially related to extant Corylopsis and Fothergilla. The occurrence of eight additional pollen forms/genera has been reported, although their botanical affiliation has not been sufficiently substantiated.
Newly examined deposits from various Oligocene, early, middle and late Miocene Iberian localities have allowed three new fossil hamamelidaceous genera to be formally described: Disanthuspollenites, Parrotiapollenites and Embolantherapollenites, as well as to propose a new combination and emendation of Tricolpopollenites indeterminatus, a species of uncertain affinity within Hamamelidaceae. These genera possess unique morphological features which normally enable their recognition without the need for SEM analysis. Given the difficulty in generic differentiation of macrofossil remains, this study highlights the potential from fossil pollen to obtain knowledge on the past biodiversity of Hamamelidaceae. The new data provided contribute to the understanding of the evolution and palaeobiogeography of Hamamelidaceae and are expected to help in the identification of the family in standard palynological studies.
Systematic palaeontology
Order Saxifragales
Family Hamamelidaceae
Subfamily Exbucklandioideae
Disanthuspollenites M.Casas-Gallego & E.Barrón, gen. nov.
Type species: Disanthuspollenites operculatus (Fig. 2l–o).
Etymology: From extant genus Disanthus.
Diagnosis: Medium sized (20–30 µm) tricolpate pollen with sub-spheroidal shape. The colpi have sharply pointed ends and enclose opercula that are clearly visible in polar view as islands of isolated exine. The exine is reticulate, with small lumina of more or less uniform size over the entire surface.
Notes: Disanthuspollenites is here proposed to designate fossil pollen related to extant Disanthus. The consistent development of operculum in the colpus area is a unique feature among hamamelidaceous pollen (Bogle and Philbrick 1980), which is the main basis for the botanical assignment and allows for straightforward identification in routine palynological studies. The species Psilatricolporites operculatus, related to extant genus Alchornea (Van der Hammen and Wymstra 1964), as well as the extant genera Potentilla and Teucrium, among others, also possesses tricolpate operculate pollen (Reille 1992), but the combination of prolate spheroidal shape, three long apertures with pointed ends and reticulate exine, is exclusive to Disanthuspollenites.
Disanthuspollenites operculatus M.Casas-Gallego & E.Barrón, sp. nov.—HOLOTYPE: Fig. 2l. Slide Aα-1. Equatorial diameter: 24 µm. Type locality: As Pontes Basin (Galicia, Spain), late Oligocene (Chattian), 43.47ºN, 7.90ºW.
Figures: This study, Fig. 2l–o. 1989 – Distylium; Van Campo, p. 22, pl. 5, images 1–8. 2005 – Alchornea; Jiménez-Moreno, p. 121, pl. 1, images 25–26. 2005 – Disanthus; Jiménez-Moreno, p. 124, pl. 4, images 9–10. 2005 – Corylopsis; Jiménez-Moreno, p. 124, pl. 4, images 11–12. 2015 – Disanthus; Casas-Gallego et al., p. 89, pl. 1, images 4–6. 2015 – Disanthus type; Gaudant et al., p. 109, Fig. 3i.
Known species stratigraphy: Upper Oligocene (Chattian) to upper Miocene (Messinian).
Etymology: From Latin operculum: cover, lid – with reference to the development of operculate colpi.
Diagnosis: Tricolpate pollen with rounded, nearly circular, outline in polar view. The polar axis is slightly longer than the equatorial, resulting in a P/E (polar/equatorial axis) ratio of 1.0–1.1. The colpi are operculate, 12–16 µm long, not reaching the polar area. The polar area index is 0.35–0.40. The exine is reticulate, with small lumina of more or less uniform size over the entire surface. The exine in the operculum area possesses the same reticulation pattern as in the rest of the surface. Lumina of the reticulum are up to 1 µm long. The murus of the reticulum is about 1 µm high.
Dimensions: The polar and equatorial axes range in length between 21 and 30 µm (20 specimens measured).
Notes: In the Iberian Peninsula, fossil pollen virtually identical to the specimens from the Oligocene of As Pontes Basin described here have been identified in the early Miocene (Burdigalian) of Rubielos de Mora (Jiménez Moreno 2005; Jiménez-Moreno et al. 2007), middle Miocene of central Spain (Gaudant et al. 2015) and late Miocene (Messinian) of Venta del Moro, in eastern Spain (Van Campo 1989; Casas-Gallego et al. 2015). Van Campo (1989) assigned these tricolpate, operculate pollen forms to Distylium. However, Distylium is one of the few hamamelidaceous genera that produce mainly pantoporate (occasionally also tetracolpate) pollen (Bogle and Philbrick 1980; Endress 1993). Based on the uniqueness that the presence of operculum represents among extant Hamamelidaceae, we consider that the specimens from Venta del Moro (Fig. 2n–o; Van Campo, 1989, pl. 5, images 1–10; Casas-Gallego et al., 2015, pl. 1, images 4–6) are more accurately placed within Disanthuspollenites, suggesting a relationship with extant Disanthus. Using the same criteria, the tricolpate, reticulate and operculate specimens reported by Jiménez Moreno (2005) as Alchornea and Corylopsis should be allocated in Disanthuspollenites. Extant Alchornea produces operculate, psilate pollen (Gosling et al. 2013). Alchornea is nowadays a pantropical genus with a well-known stratigraphic distribution in low-latitude regions ranging from the Eocene up to the present day (Germeraad et al. 1968; Salard-Cheboldaeff 1990). In Europe, palynomorphs assigned to Psilatricolporites operculatus (related to Alchornea) have been reported mainly from the Eocene (Châteauneuf 1980; Schuler 1990; Cavagnetto and Anadón 1996), when tropical-like climatic conditions predominated. However, it is unlikely that climatic conditions in Europe during the Neogene were suitable for the occurrence of this thermophilous genus. Also, the presence of operculum in the specimens identified by Jiménez Moreno (2005) as Corylopsis is the basis for including them in the synonymy list of Disanthuspollenites operculatus.
The specimens of D. operculatus from the late Oligocene (Chattian) of As Pontes represent the oldest record of this pollen form. Because of the relatively long stratigraphic range covered by the occurrences reported in Spain (Fig. 5), it is possible that various species evolved. However, except for slight differences in the height of the murus and the width of the operculum, there are no significant morphological differences between the specimens of different ages and localities (Fig. 2l–o).
Subfamily Hamamelidoideae
Corylopsispollenites E.Worobiec in Stuchlik et al. (2014)
cf. Corylopsispollenites sp. (Fig. 3k–n).
Type species: Corylopsispollenites microreticulatus. Middle Miocene of central Poland.
Description: Tricolpate pollen with a prolate/subprolate shape and circular outline in polar view. The colpi are straight, sharply ended and possess smooth, clean margins. The colpi are long, resulting in a reduced polar area (index 0.15–0.2). The exine is 1–2 µm thick, microreticulate, with lumens less than 1 µm long uniformly covering the entire surface.
Dimensions: Polar axis about 25 µm. Equatorial axis 21–22 µm.
Notes: The genus Corylopsispollenites was erected to accommodate fossil palynomorphs related to extant Corylopsis. The original diagnosis indicates the occurrence of weakly visible pores, which have neither been observed in our specimens nor in extant Corylopsis pollen (Bogle and Philbrick 1980). Apart from the latter difference, which may be due to the interpretation of the colpi shape by Stuchlik et al. (2014), the specimens from the Chattian of the As Pontes Basin (Fig. 3k–n) share the general morphological features outlined in the original diagnosis of the genus. However, the taxonomic assignment is questionable, as we consider that, in the absence of SEM images, the original diagnosis, as well as extant pollen of Corylopsis, does not provide sufficient morphological features to differentiate it from other Hamamelidaceae genera such us Hamamelis, Loropetalum and Ostrearia. Therefore, SEM analysis is needed to conclusively assign fossil pollen to Corylopsis.
The presence of Corylopsis in the European Cenozoic has been demonstrated with abundant seed occurrences (Manchester 1999). In the Iberian Peninsula, however, the reported records derive from palynomorphs that have not been sufficiently described or documented with images. Fossil pollen identified generally as Corylopsis have been reported from the middle to late Eocene (Bartonian and Priabonian) of the Ebro Basin (Cavagnetto and Anadón 1996) and the early Miocene (Burdigalian) of the Rubielos de Mora Basin, in eastern Spain (Jiménez Moreno 2005). The specimens presented in the latter study as Corylopsis [Jiménez Moreno (2005), page 124, pl. 4, images 11–12] are operculate and have been included in the list of synonyms of Disanthuspollenites operculatus in the present work. Jiménez-Moreno and Suc (2007) also indicate the occurrence of pollen identified questionably as Corylopsis in middle Miocene deposits of eastern Spain.
Embolantherapollenites M.Casas-Gallego & E.Barrón, gen. nov.
Type species: Embolantherapollenites calvicolpatus (Fig. 2a–k).
Etymology: From extant genus Embolanthera.
Diagnosis: Medium size (18–27 µm) tricolpate pollen with sub-spheroidal shape and colpi with sharply pointed ends. The exine ornamentation consists of a coarse, deep and homobrochate reticulum resting upon short baculae. The outline of the lumina is angular. The colpi margins are very distinctive, displaying a smooth, non-ornamented surface, a feature clearly visible in LM on both polar and equatorial views.
Notes: The development of non-ornamented colpi is a distinctive feature that can be observed in polar and equatorial views and with both LM and SEM (Fig. 2a–k). Among extant Hamamelidaceae, this kind of colpi occurs only in Embolanthera and Maingaya (Bogle and Philbrick 1980). Embolanthera and Maingaya have been shown to be phylogenetically close and are currently included in the subtribe Loropetalinae (Endress 1989b). The only morphological differences between the pollen of these two genera are the larger size of the non-ornamented area in the colpi membrane and the development of shorter baculae supporting the reticulum in Embolanthera. The latter features strongly suggest that our specimens are related to Embolanthera, providing solid evidence for the presence of this genus in Europe.
Embolantherapollenites calvicolpatus M.Casas-Gallego & E.Barrón, sp. nov. (Fig. 2a-k).—HOLOTYPE: Fig. 2d–f. Slide DC-1. Equatorial diameter: 25 µm. Type locality: As Pontes Basin (Galicia, Spain), 43.47ºN, 7.90ºW.
Known species stratigraphy: Rupelian and Chattian of the As Pontes Basin (Galicia, Spain).
Etymology: From Latin calvus, meaning bald, in reference to the lack of ornament in the colpi margins.
Diagnosis: Tricolpate pollen with rounded, nearly circular outline in polar view. The polar axis is as long or slightly longer than the equatorial axis, resulting in a P/E ratio of about 1.0–1.1. Colpi are around 12–15 µm long, not meeting at the poles. The polar area index is 0.25–0.3. The reticulum is about 2 µm deep, rather homobrochate, supported by a single row of short and scarce bacula. The lumina of the reticulum are up to 3 µm long and have a more or less uniform size over the entire surface. The outline of the lumina is an angular.
Dimensions: The equatorial axis is 18–26 µm long, while the polar axis is 20–26 µm long (12 specimens measured).
Notes: Palynomorphs ascribed generally to Embolanthera have been reported from the Pliocene of north-east Spain (Suc and Cravatte 1982) and from a few localities on the Spanish Mediterranean coast with a general middle Miocene to Pliocene age (Jiménez-Moreno et al. 2010). Unfortunately, these records have never been documented with pictures or descriptions, which renders them questionable. The Neogene floristic dynamic in the Iberian Peninsula is characterized by the gradual extinction of taxa that are nowadays typical of tropical and subtropical regions, such as Embolanthera (Postigo-Mijarra et al. 2009). Given that by the late Miocene most thermophilous taxa that were once present in Iberia had become extinct regionally and a major part of the subtropical-like habitats had disappeared, the occurrence of Embolanthera in the late Miocene–Pliocene would be a significant finding that requires further evidence.
Fothergilla Murray in Linnaeus 1774.
cf. Fothergilla sp. (Fig. 4k–n).
Description: Tricolpate pollen with subprolate shape. The colpi margins are even, smooth and slightly thickened. The colpi have pointed ends, with no microgranules preserved. The exine is about 1.5 µm thick, reticulate, with heterobrochate reticulum. The lumina of the reticulum are up to 2 µm long in the mesocolpium, significantly diminish in size towards the polar area. The muri is about 1 µm deep.
Dimensions: Equatorial diameter 35–45 µm (3 specimens measured).
Notes: The considerable reduction in size of the lumina in the polar area is a feature that occurs only in Fothergilla and Eustigma among extant Hamamelidaceae (Bogle and Philbrick 1980). However, the fact that Eustigma produces pollen with considerably wider murus allows this genus to be discounted as a likely living relative. Our specimens are very similar to extant Fothergilla but are here assigned tentatively due to the lack of SEM images and the low number of specimens available for study, which are considered insufficient to erect a new fossil genus. Palynomorphs attributed to Fothergilla have been reported from the middle Eocene to early Oligocene (Bartonian to Rupelian) of north-east Spain (Cavagnetto and Anadón 1996) and from the middle Miocene of eastern Spain (Jiménez-Moreno and Suc 2007; Jiménez-Moreno et al. 2010); however, none of these occurrences is sufficiently documented.
Parrotiapollenites M.Casas-Gallego & E.Barrón, gen. nov.
Type species: Parrotiapollenites asper (Fig. 3a–j).
Etymology: From extant genus Parrotia.
Diagnosis: Tricolpate pollen with sub-spheroidal shape. The colpi possess bluntly rounded ends and irregular, ragged margins covered with numerous microgranules. The exine is reticulate, with heterobrochate reticulum. The reticulation pattern is more or less uniform over the entire surface. The muri of the reticulum bear numerous microbacula/verrucae visible under SEM.
Notes: Parrotiapollenites can be differentiated from other genera within Hamamelidaceae by having colpi with characteristic rounded ends and irregular margins densely covered by microgranules. Occasionally, isolated patches of exine also occur in the colpi area (Fig. 3f, g), but they do not form a well-developed operculum. The structure of the reticulum, supporting multiple microbacula, and the nature of the colpi margins, as revealed by the observation with SEM (Fig. 3e–f), denote a clear relationship with extant Parrotia (Bogle and Philbrick 1980).
Parrotiapollenites asper M.Casas-Gallego & E.Barrón, sp. nov.—HOLOTYPE: Fig. 3a–b. Slide F-4. Equatorial diameter: 31 µm. Type locality: As Pontes Basin (Galicia, Spain), 43.47ºN, 7.90ºW.
Figures: This study, Fig. 3a–j. 1980 – Parrotia cf. persica; Suc, pl. 4, images 10–11. 1980 – Parrotia cf. persica; Julia Bruguès & Suc, pl. 1, images 11–12. 1996 – cf. Parrotia; Barrón, p. 733, pl. 9, image 9. 2005 – Parrotia cf. persica; Jiménez-Moreno, p. 124, images 28–31 and p. 133, Fig. 23, images 1–4. 2016 – Tricolporopollenites indeterminatus; Barrón et al., p. 105, pl. 1, image s.
Known species stratigraphy: Chattian to Lower Pleistocene.
Etymology: From Latin asper, meaning rough, irregular, uneven – in reference to the irregular, ragged nature of the colpi margins.
Diagnosis: Tricolpate pollen with rounded outline and slightly sunken apertures in polar view. Colpi range in length from 8 to 18 µm, resulting in a polar area index of 0.25–0.35. Colpi edges irregular, bearing numerous microgranules, usually visible in LM. The outer layer of the exine is clearly thicker than the inner layer. Low heterobrochate reticulum with lumina rounded in shape and ranging in size between 0.5 and 1.8 µm. Muri about 1 µm high, bearing numerous microbacula-verrucae.
Dimensions: The equatorial diameter is 25–33 µm long (15 specimens measured).
Notes: Although scarce, pollen assigned to Parrotia are the most commonly identified among hamamelidaceous genera in the Iberian Peninsula. Parrotia pollen have been reported from lower Miocene to Lower Pleistocene deposits (Suc 1980; Suc and Cravatte 1982; Bessais and Cravatte 1988; Leroy 1997; Jiménez-Moreno et al. 2007; Barrón et al. 2016; Altolaguirre et al. 2020). The palynomorphs of Rupelian and Chattian age from the As Pontes Basin represent the oldest palaeobotanical record of Parrotia in the Iberian Peninsula (Fig. 3a–j). Parrotia is believed to have become extinct in Iberia during the Early Pleistocene, around 1.2 Ma (Postigo-Mijarra et al. 2010).
Parrotiapollenites asper differs from the closely similar species Tricolporopollenites indeterminatus primarily by having a more spherical shape, which results in an increased tendency to become preserved in polar view compared to T. indeterminatus. Also, T. indeterminatus lacks microgranules in the colpi area, or they occur in much fewer numbers. The colpi in T. indeterminatus can also develop rounded ends, but not as broadly rounded as in Parrotiapollenites.
Parrotiapollenites asper does not differ substantially from other records in the Iberian Peninsula, such as that of Parrotia cf. persica figured in Suc (1980) and Jiménez-Moreno et al. (2005). Therefore, although the possibility for the occurrence of more than one species in the Iberian Cenozoic cannot be discounted, the lack of distinctive morphological features precludes the confirmation of intrageneric diversification.
In the Iberian Peninsula, macrofloral remains assigned to Parrotia have been reported from Chattian deposits in north-eastern Spain, where a single leaf was identified (Moreno-Domínguez et al. 2015), the early Miocene (Burdigalian) of the Ribesalbes Basin in eastern Spain, and the late Miocene (Tortonian) of the La Cerdanya Basin in the eastern Pyrenees, where leaves of ‘Parrotia’ pristina were recovered (Barrón and Postigo-Mijarra 2011; Barrón et al. 2014). However, as indicated by the latter authors, the leaves of Parrotia are virtually indistinguishable from those of Fothergilla, Corylopsis and Hamamelis. Therefore, palynomorphs are the most reliable source of palaeobotanical information for this genus.
Subfamily uncertain.
Tricolpopollenites Pflug & Thomson in Thomson & Pflug, 1953.
Tricolpopollenites indeterminatus Romanowicz, comb. nov. and emend.—HOLOTYPE: Pollenites indeterminatus (Romanowicz 1961; page 355, plate 21, image 275). NEOTYPE (designated here): Fig. 4g–h. Slide C-B. Neotype locality: As Pontes Basin (Galicia, Spain), 43.47ºN, 7.90ºW.
Figures: This study, Fig. 4a–j. 1961 – Pollenites indeterminatus n. f.; Romanowicz, p. 355, pl. 21, image 275. 1994 – Tricolporopollenites indeterminatus (Romanowicz) Ziembińska-Tworzydło; Ziembińska-Tworzydło, pl. 16, images 19–20. 2007 – cf. Parrotia; Ivanov, p. 376, Fig. 6, images 6–7. 2009 – Tricolporopollenites indeterminatus (Romanowicz) Ziembińska-Tworzydło; Worobiec, p. 33, pl. 11, image 13. 2014 – Tricolporopollenites indeterminatus; Stuchlik et al., pl. 17, image 4.
Original diagnosis sensu Romanowicz (1961): The specimen found in the sediments from Kruszyna 10 borehole have a size of 38 µm, an oval outline, three continuous furrows (colpi) not interrupted by pores, and a surface covered with a dense mesh (reticulate).
Holotype: Pollenites indeterminatus (Romanowicz 1961; page 355, plate 21, image 275).
Emended diagnosis: Tricolpate pollen with rounded, nearly circular outline in polar view and elliptical outline in equatorial view. Pollen prolate or subprolate (P/E ratio = 1.24–1.53). The colpi are straight, with a length of 15–20 µm, not meeting at the poles. The colpi ends are slightly rounded and the colpi margins are well defined (not irregular), usually smooth, although occasionally they may bear a few scattered microgranules. The exine is reticulate, about 1.5 µm thick. The lumina of the reticulum form different geometric shapes with rounded corners which are up to 2 µm long. The murus is smooth, not supporting any kind of additional ornament, slightly higher at the nodes of the reticulum. The murus is about 1 µm high and it is supported by extremely short bacula.
Dimensions: The polar axis is 28–41 µm. The equatorial axis is 25–35 µm (25 specimens measured).
Known species stratigraphy: Oligocene (Rupelian to Chattian).
Notes: The original description of this species from the ‘middle’ Oligocene of Poland did not explicitly designate a holotype (Romanowicz 1961) despite such designation became mandatory in 1958. However, according to the current version of the International Code of Nomenclature for algae, fungi and plants (Turland et al. 2018), the specimen shown in image 275 of plate 21 in Romanowicz (1961) can be considered to be the holotype as per section b of article 9.1 of the code: ‘a holotype of a name of a species or infraspecific taxon is the one specimen or illustration either (a) indicated by the author(s) as the nomenclatural type or (b) used by the author(s) when no type was indicated’. The poor original diagnosis, which does not allow for a clear differentiation from other similar species, combined with the contradicting description provided in its subsequent transference to T. indeterminatus (Ziembińska-Tworzydło 1974), which has been widely followed in later palynological studies (Stuchlik et al. 2014), introduced considerable confusion regarding the taxonomic status of this species. This may have resulted in various authors using this species name to designate fossil palynomorphs with different botanical affinities.
The new combination and emendation proposed here is based on well-preserved specimens from the Oligocene (Rupelian and Chattian) of the As Pontes Basin. It is intended to throw some light on the historically poor definition of the species. Because the current location of the material used by Romanowicz (1961) is unknown and the original diagnosis is supported only by a single micrograph that cannot be critically identified or used for purposes of precise application of the species name, we propose the specimen in figures g–h of Fig. 4 as the neotype to serve as an interpretative type of the ambiguous original specimen (Turland et al. 2018). Here we follow the proposal for neotypification of fossil-taxa names introduced by Gravendyck et al. (2020) which is particularly useful in palaeopalynology, given the large number of types of fossil names lost or damaged. Although the original diagnosis is very succinct, Romanowicz (1961) described a tricolpate (not tricolporate) pollen. Therefore, in the absence of any emendation, the new combination carried out by Ziembińska-Tworzydło (1974) transferring the species to the genus Tricolporopollenites should not be effective. The specimens from the Oligocene of Poland attributed by Ziembińska-Tworzydło (1974) to T. indeterminatus are clearly colporate (page 397, plate 24, images 9a and 9b) and were described as possessing well-developed pores. While it is not impossible that tricolporate pollen evolved within Hamamelidaceae, to date the development of tricolporate pollen has not been demonstrated in any extant or fossil genera of the family, which is essentially tricolpate; rarely polycolpate/polyporate in Distylium, Matudea and Sycopsis (Bogle and Philbrick 1980; Endress 1989a; 1993).
Tricolpopollenites indeterminatus has traditionally been one of the few palynomorph species used to designate fossil pollen related to Hamamelidaceae in the European Cenozoic. It has been frequently reported in Eastern Europe (Poland, Czech Republic, Slovakia), in deposits ranging in age from Oligocene to late Miocene (Grabowska 1965; Ziembińska-Tworzydło 1974; Ziembińska-Tworzydło et al. 1994; Worobiec 2009, 2014; Kováčová et al. 2011). In the Iberian Peninsula, it has been encountered in the As Pontes Basin. Traditionally, a link between T. indeterminatus and extant Parrotia or Distylium has been assumed (Kováčová et al. 2011; Stuchlik et al. 2014; Worobiec 2014). The SEM analysis of T. indeterminatus provided here for the first time (Fig. 4e–f) confirms a relationship with Hamamelidaceae, as indicated by the colpi and reticulum shape and structure but suggests that a link with Parrotia and Distylium as living relatives is unlikely. The reticulum of T. indeterminatus does not bear the microbacula characteristic of Parrotia and Distylium (Bogle and Philbrick 1980). Also, T. indeterminatus differs from Parrotiapollenites asper by having a more prolate shape, which makes it more prone to being preserved in equatorial view (Fig. 4g–j), longer colpi with ends only slightly rounded (not as rounded as in Parrotia) and, more importantly, more or less regular (not ragged) margins with only few or no microgranules. The reticulum of T. indeterminatus also possesses larger lumina. Tricolpopollenites indeterminatus displays some similarities with extant Parrotiopsis, although these are not conclusive for a confident botanical affinity determination, the possibility also exists that it may represent an extinct genus within Hamamelidaceae.
References
Akhani H, Djamali M, Ghorbanalizadeh A, Ramezani E (2010) Plant biodiversity of Hyrcanian relict forests, N Iran: An overview of the flora, vegetation, palaeoecology and conservation. Pakistan J Bot 42:231–258
Alonso-Zarza AM, Armenteros I, Braga JC, Muñoz A, Pujalte V, Ramos E, Aguirre J, Alonso-Gavilán G, Arenas C, Baceta JI, Carballeira J, Calvo JP, Corrochano A, Fornós JJ, González A, Luzón A, Martín JM, Pardo G, Payros A, Pérez A, Pomar L, Rodriguez JM, Villena J (2002) Tertiary. In: Gibbons W, Moreno T (eds) The Geology of Spain. Geological Society of London, London, pp 293–334
Altolaguirre Y, Postigo-Mijarra JM, Barrón E, Carrión JS, Leroy SAG, Bruch AA (2019) An environmental scenario for the earliest hominins in the Iberian Peninsula: Early Pleistocene palaeovegetation and palaeoclimate. Rev Palaeobot Palynol 260:51–64. https://doi.org/10.1016/j.revpalbo.2018.10.008
Altolaguirre Y, Bruch AA, Gibert L (2020) A long Early Pleistocene pollen record from Baza Basin (SE Spain): Major contributions to the palaeoclimate and palaeovegetation of Southern Europe. Quatern Sci Rev 231:106199. https://doi.org/10.1016/j.quascirev.2020.106199
Anadón P, Cabrera L, Guimerà J, Santanach P (1985) Paleogene strike-slip deformation and sedimentation along the southeastern margin of the Ebro Basin. Soc Econ Paleontol Mineral Spec Publ 37:303–318. https://doi.org/10.2110/pec.85.37.0283
Barrón E, Postigo-Mijarra JM (2011) Early Miocene fluvial-lacustrine and swamp vegetation of La Rinconada mine (Ribesalbes-Alorca basin, Eastern Spain). Rev Palaeobot Palynol 165:11–26. https://doi.org/10.1016/j.revpalbo.2011.02.001
Barrón E, Hernández JM, López-Horgue MA, Alcalde-Olivares C (2006a) Palaeoecology, biostratigraphy and palaeoclimatology of the lacustrine fossiliferous beds of the Izarra Formation (Lower Miocene, Basque-Cantabrian Basin, Álava province, Spain) based on palynological analysis. Rev Esp Micropaleontol 38:321–338
Barrón E, Lassaletta L, Alcalde-Olivares C (2006b) Changes in the Early Miocene palynoflora and vegetation in the east of the Rubielos de Mora Basin (SE Iberian Ranges, Spain). Neues Jahrb Geol Palaeontol Abh 242:171–204. https://doi.org/10.1127/njgpa/242/2006/171
Barrón E, Rivas Carballo R, Postigo Mijarra JM, Alcalde Olivares C, Vieira M, Castro L, Pais J, Valle Hernández M (2010) The Cenozoic vegetation of the Iberian Peninsula: A synthesis. Rev Palaeobot Palynol 162:382–402. https://doi.org/10.1016/j.revpalbo.2009.11.007
Barrón E, Postigo Mijarra JM, Diéguez C (2014) The late Miocene macroflora of the La Cerdanya Basin (Eastern Pyrenees, Spain): towards a synthesis. Palaeontographica Abt B 291:85–129
Barrón E, Postigo-Mijarra JM, Casas-Gallego M (2016) Late Miocene vegetation and climate of the La Cerdanya Basin (eastern Pyrenees, Spain). Rev Palaeobot Palynol 235:99–119. https://doi.org/10.1016/j.revpalbo.2016.08.007
Benedict JC, Pigg KB, DeVore ML (2008) Hamawilsonia boglei gen. et sp. nov. (Hamamelidaceae) from the Late Paleocene Almont flora of central north Dakota. Int J Pl Sci 169:687–700. https://doi.org/10.1086/533607
Bessais E, Cravatte J (1988) Les écosystèmes végétaux pliocènes de Catalogne méridionale. Variations latitudinales dans le domaine nord-ouest méditerranéen. Geobios 21:49–63. https://doi.org/10.1016/S0016-6995(88)80031-7
Bogle AL, Philbrick CT (1980) A generic atlas of Hamamelidaceous pollens. Contrib Gray Herb 210:29–103
Casas-Gallego, M 2018. Estudio palinológico del Oligoceno-Mioceno Inferior de la Cuenca de As Pontes (Galicia, España). Serie Tesis Doctorales, 31. Instituto Geológico y Minero de España, Madrid
Casas-Gallego M, Barrón E (2020) New pollen genera and species from the Oligocene of northern Spain and a systematic, biostratigraphic and biogeographic re-evaluation of coeval taxa. J Syst Palaeontol 18:1961–1994. https://doi.org/10.1080/14772019.2020.1863488
Casas-Gallego M, Lassaletta L, Barrón E, Bruch AA, Montoya P (2015) Vegetation and climate in the eastern Iberian Peninsula during the pre-evaporitic Messinian (late Miocene). Palynological data from the Upper Turolian of Venta del Moro (Spain). Rev Palaeobot Palynol 215:85–99. https://doi.org/10.1016/j.revpalbo.2015.01.001
Cavagnetto C (2002) La palynoflore du Bassin d’As Pontes en Galice dans le Nord Ouest de l’Espagne à la limite Rupélien-Chattien (Oligocène). Palaeontographica Abt B 263:161–204
Cavagnetto C, Anadón P (1996) Preliminary palynological data on floristic and climatic changes during the Middle Eocene-Early Oligocene of the eastern Ebro Basin, northeast Spain. Rev Palaeobot Palynol 92:281–305. https://doi.org/10.1016/0034-6667(95)00096-8
Châteauneuf JJ (1980) Palynostratigraphie et paléoclimatologie de l’Eocene Supérieur et de l’Oligocene du Bassin de Paris. Mem BRGM 116:1–357
Civis J (2004) Cuencas cenozoicas. In: Vera JA (ed) Geología de España. SGE-IGME, Madrid
Collinson ME, Hooker JJ (2003) Paleogene vegetation of Eurasia: framework for mammalian faunas. Deinsea 10:41–83
Crepet WL, Nixon KC, Friis EM, Freudenstein JV (1992) Oldest flowers of hamamelidaceous affinity, from the Upper Cretaceous of New Jersey, North America. Proc Natl Acad Sci USA 89:8986–8989. https://doi.org/10.1073/pnas.89.19.8986
Denk T, Tekleva MV (2006) Comparative pollen morphology and ultrastructure of Platanus: Implications for phylogeny and evaluation of the fossil record. Grana 45:195–221. https://doi.org/10.1080/00173130600873901
Denk T, Grímsson F, Zetter R (2012) Fagaceae from the early Oligocene of Central Europe: Persisting new world and emerging old biogeographic links. Rev Palaeobot Palynol 169:7–20. https://doi.org/10.1016/j.revpalbo.2011.09.010
Endress PK (1989b) A suprageneric taxonomic classification of the Hamamelidaceae. Taxon 38:371–376. https://doi.org/10.2307/1222267
Endress PK (1989a) Aspects of evolutionary differentiation of the Hamamelidaceae and the Lower Hamamelididae. Pl Syst Evol 162:193–211. https://doi.org/10.1007/978-3-7091-3972-1_10
Endress PK (1993) Hamamelidaceae. In: Kubitzki K (ed) The families and genera of vascular plants, vol. 2. Springer, Berlin, Heidelberg, New York, pp 322–331
Friis EM, Crane PR, Pedersen KR (2011) Early flowers and angiosperm evolution. Cambridge University Press, Cambridge
Fritz A, Allesch K (1999) Pollenmorphologische Bilddokumentation ausgewählter Sippen der ehemals Kätzchenblütigen. Carinthia II 189:471–490
Gaudant J, Barrón E, Anadón P, Reichenbacher B, Peñalver E (2015) Palaeoenvironmental analysis of the Miocene Arcas del Villar gypsum sequence (Spain), based on palynomorphs and cyprinodontiform fishes. Neues Jahrb Geol Palaeontol Abh 277:105–124. https://doi.org/10.1127/njgpa/2015/0503
Germeraad JH, Hopping CA, Muller J (1968) Palynology of Tertiary sediments from tropical areas. Rev Palaeobot Palynol 6:189–348. https://doi.org/10.1016/0034-6667(68)90051-1
González-Sampériz P, Leroy SAG, Carrión JS, Fernández S, García-Antón M, Gil-García MJ, Uzquiano P, Valero-Garcés B, Figueiral I (2010) Steppes, savannahs, forests and phytodiversity reservoirs during Pleistocene in the Iberian Peninsula. Rev Palaeobot Palynol 162:427–457. https://doi.org/10.1016/j.revpalbo.2010.03.009
Gosling WD, Miller CS, Livingstone DA (2013) Atlas of the tropical West African pollen flora. Rev Palaeobot Palynol 199:1–135. https://doi.org/10.1016/j.revpalbo.2013.01.003
Grabowska I (1965) The Middle Oligocene age of the Torun clays, based on the spore and pollen analysis. Kwart Geol 9:815–836
Gravendyck J, Bachelier JB, Kürschner WM, Herendeen PS (2020) A proposal to solve a paradox when neotypifying names of fossil-taxa. Taxon 669:628. https://doi.org/10.1002/tax.12249
Grímsson F, Zetter R, Baal C (2011) Combined LM and SEM study of the Middle Miocene (Sarmatian) palynoflora from the Lavanttal Basin, Austria: Part I. Bryophyta, Lycopodiophyta, Pteridophyta, Ginkgophyta, and Gnetophyta. Grana 50:102–128. https://doi.org/10.1080/00173134.2011.585804
Grímsson F, Meller B, Bouchal JM, Zetter R (2015a) Combined LM and SEM study of the middle Miocene (Sarmatian) palynoflora from the Lavanttal Basin, Austria: part III. Magnoliophyta 1 – Magnoliales to Fabales. Grana 54:85–128. https://doi.org/10.1080/00173134.2015.1007081
Grímsson F, Zetter R, Grimm GW, Pedersen GK, Pedersen AK, Denk T (2015b) Fagaceae pollen from the early Cenozoic of West Greenland: revisiting Engler’s and Chaney’s Arcto-Tertiary hypotheses. Pl Syst Evol 301:809–832. https://doi.org/10.1007/s00606-014-1118-5
Gruas-Cavagnetto C (1978) Étude palynologique de l’Éocène du Bassin Anglo-Parisien. Mem Soc Geol Fr 131:1–64
Iii APG (2009) An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Bot J Linn Soc 161:105–121. https://doi.org/10.1111/j.1095-8339.2009.00996.x
Jiménez Moreno G (2005) Utilización del análisis polínico para la reconstrucción de la vegetación, clima y estimación de paleoaltitudes a lo largo del arco alpino europeo durante el Mioceno (21–8 Ma). Departamento de Estratigrafía y Paleontología, Laboratoire Paléoenvironnements et Paléobiosphères, University of Granada, Granada
Jiménez-Moreno G, Suc J-P (2007) Middle Miocene latitudinal climatic gradient in Western Europe: evidence from pollen records. Palaeogeogr Palaeoclimatol Palaeoecol 253:208–225. https://doi.org/10.1016/j.palaeo.2007.03.040
Jiménez-Moreno G, Rodríguez-Tovar FJ, Pardo-Igúzquiza E, Fauquette S, Suc JP, Müller P (2005) High-resolution palynological analysis in late early-middle Miocene core from Pannonian Basin, Hungary: climatic changes, astronomical forcing and eustatic fluctuations in the Central Paratethys. Palaeogeogr Palaeoclimatol Palaeoecol 216:73–97. https://doi.org/10.1016/j.palaeo.2004.10.007
Jiménez-Moreno G, Fauquette S, Suc JP, Abdul Aziz H (2007) Early Miocene repetitive vegetation and climatic changes in the lacustrine deposits of the Rubielos de Mora Basin (Teruel, NE Spain). Palaeogeogr Palaeoclimatol Palaeoecol 250:101–113. https://doi.org/10.1016/j.palaeo.2007.03.004
Jiménez-Moreno G, Fauquette S, Suc JP (2010) Miocene to Pliocene vegetation reconstruction and climate estimates in the Iberian Peninsula from pollen data. Rev Palaeobot Palynol 162:403–415. https://doi.org/10.1016/j.revpalbo.2009.08.001
Jiménez-Moreno G, Burjachs F, Exposito I, Oms O, Carrancho A, Villalaín JJ, Agustí J, Campeny G, Gómez de Soler B, Made J (2013) Late Pliocene vegetation and orbital-scale climate changes from the western Mediterranean area. Global Planet Change 108:15–28. https://doi.org/10.1016/j.gloplacha.2013.05.012
Julia Bruguès R, Suc J-P (1980) Analyse pollinique des dépôts lacustresdu Pléistocène inférieur de Banyoles (Bañolas, site de la Bòbila Ordis - Espagne): un élément nouveau dans la reconstitution de l’histoire paléoclimatique des régions méditerranéennes d’Europe occidentale. Geobios 13:5–19
Kedves M (1998) Études palynologiques des couches du Tertiaire inférieur de la région parisienne. IX Pl Cell Biol Dev 9:28–42
Kmenta M, Zetter R (2013) Combined LM and SEM study of the upper Oligocene/lower Miocene palynoflora from Altmittweida (Saxony): providing new insights into Cenozoic vegetation evolution of Central Europe. Rev Palaeobot Palynol 195:1–18. https://doi.org/10.1016/j.revpalbo.2013.03.007
Knobloch E, Mai DH (1986) Monographie der Früchte und Samen in der Kreide von Mitteleuropa. Rozpr Ústredního Ústavu Geol, Praha 47:1–219
Kováčová M, Doláková N, Kováč M (2011) Miocene vegetation pattern and climate change in the northwestern Central Paratethys domain (Czech and Slovak Republic). Geol Carpath 62:251–266. https://doi.org/10.2478/v10096-011-0020-0
Leroy SAG (1997) Climatic and non-climatic lake-level changes inferred from a Plio-Pleistocene lacustrine complex of Catalonia (Spain): palynology of the Tres Pins sequences. J Paleolimnol 17:347–367. https://doi.org/10.1023/A:1007929213952
Leroy SAG, Roiron P (1996) Latest Pliocene pollen and leaf floras from Bernasso palaeolake (Escandorgue Massif, Hérault, France). Rev Palaeobot Palynol 94:295–328. https://doi.org/10.1016/S0034-6667(96)00016-4
Li W, Zhang G-F (2015) Population structure and spatial pattern of the endemic and endangered subtropical tree Parrotia subaequalis (Hamamelidaceae). Flora 112:10–18. https://doi.org/10.1016/j.flora.2015.02.002
López-Martínez N (1989) Tendencias en Paleobiogeografía. El futuro de la biogeografía del pasado. In: Aguirre E (ed) Paleontología. Nuevas tendencias, CSIC, Madrid, pp 271–296
Magallón S (2007) From Fossils to Molecules: Phylogeny and the Core Eudicot Floral Groundplan in Hamamelidoideae (Hamamelidaceae, Saxifragales). Syst Bot 32:317–347. https://doi.org/10.1600/036364407781179617
Magallón S, Herendeen PS, Crane PR (2001) Androdecidua endressii gen. et sp. nov., from the Late Cretaceous of Georgia (United States): further floral diversity in Hamamelidoideae (Hamamelidaceae). Int J Pl Sci 162:963–983. https://doi.org/10.1086/320770
Magallón-Puebla S, Herendeen PS, Endress PK (1996) Allonia decandra: floral remains of the tribe Hamamelideae (Hamamelidaceae) from Campanian strata of southeastern USA. Pl Syst Evol 202:177–198. https://doi.org/10.1007/BF00983381
Magri D, Di Rita F, Aranbarri J, Fletcher W, González-Sampériz P (2017) Quaternary disappearance of tree taxa from Southern Europe: Timing and trends. Quat Sci Rev 163:23–55. https://doi.org/10.1016/j.quascirev.2017.02.014
Mai DH (1995) Tertiäre vegetationsgeschichte Europas. G. Fischer. Jena, Stuttgart, New York
Manchester SR (1994) Fruits and seeds of the middle Eocene Nut Beds flora, Clarno Formation, Oregon. Palaeontogr Amer 58:1–205
Manchester SR (1999) Biogeographical relationships of North American Tertiary floras. Ann Missouri Bot Gard 86:472–522. https://doi.org/10.2307/2666183
Médus J (1965) L’évolution biostratigrafique d’une lagune Néogene de Galice (Espagne). Pollen Spores 7:381–393
Médus J, Nonn H (1963) Premieres résultats d’analyses polliniques à Puentes de García Rodríguez (province de Galice, Espagne) et conclusions géomorphologiques qui en découlent. CR Acad Sci Ser II A Sci Terre Planetes 256:1570–1572
Meulenkamp JE, Sissingh W (2003) Tertiary palaeogeography and tectonostratigraphic evolution of the Northern and Southern Peri-Tethys platforms and the intermediate domains of the African-Eurasian convergent plate boundary zone. Palaeogeogr Palaeoclimatol Palaeoecol 196:209–228. https://doi.org/10.1016/S0031-0182(03)00319-5
Moreno-Domínguez R, Diez JB, Jacques FMB, Ferrer J (2015) First macroflora data from La Val (Late Oligocene/Early Miocene), Estadilla (Huesca, Spain). Hist Biol 27:469–489. https://doi.org/10.1080/08912963.2014.941830
Muller J (1981) Fossil pollen record of extant angiosperms. Bot Rev 47:1–142
Pais J (1986) Évolution de la végétation et du climat pendant le Miocène au Portugal. Cienc Terra 8:179–191
Pais J (2004) The Neogene of the Lower Tagus Basin (Portugal). Rev Esp Paleontol 19:229–242
Pardo G, Arenas C, González A, Luzón A, Muñoz A, Pérez A, Pérez-Ribarés FJ, Vázquez-Urbez M, Villena J (2004) La cuenca del Ebro. In: Vera JA (ed) Geología de España. SGE-IGME, Madrid, pp 533–543
Postigo-Mijarra JM, Barrón E, Gómez Manzaneque F, Morla C (2009) Floristic changes in the Iberian Peninsula and Balearic Islands (south-west Europe) during the Cenozoic. J Biogeogr 36:2025–2043. https://doi.org/10.1111/j.1365-2699.2009.02142.x
Postigo-Mijarra JM, Morla C, Barrón E, Morales Molino C, García S (2010) Patterns of extinction and persistence of Arctotertiary flora in Iberia during the Quaternary. Rev Palaeobot Palynol 162:416–426. https://doi.org/10.1016/j.revpalbo.2010.02.015
Punt W, Hoen PP, Blackmore S, Nilsson S, Le Thomas A (2007) Glossary of pollen and spore terminology. Rev Palaeobot Palynol 143:1–81. https://doi.org/10.1016/j.revpalbo.2006.06.008
Radtke MC, Pigg KB, Wehr WC (2005) Fossil Corylopsis and Fothergilla leaves (Hamamelidaceae) from the Lower Eocene flora of Republic Washington, U.SlA, and their evolutionary and biogeographic significance. Int J Pl Sci 166:347–356. https://doi.org/10.1086/427483
Ranney TG, Lynch NP (2007) Clarifying taxonomy and nomenclature of Fothergilla (Hamamelidaceae) cultivars and hybrids. HortScience 42:470–473. https://doi.org/10.21273/HORTSCI.42.3.470
Reille M (1992) Pollen et Spores d'Europe et d'Afrique du nord. Laboratoire de Botanique Historique et Palynologie, Marseille
Rivas Carballo MR (1991) The development of vegetation and climate during the Miocene in the south-eastern sector of the Duero Basin (Spain). Rev Palaeobot Palynol 67:341–351. https://doi.org/10.1016/0034-6667(91)90049-9
Rivas Carballo MR, Alonso-Gavilán G, Valle MF, Civis J (1994) Miocene palynology of the central sector of the Duero basin (Spain) in relation to palaeogeography and palaeoenvironment. Rev Palaeobot Palynol 82:251–264. https://doi.org/10.1016/0034-6667(94)90079-5
Roiron P (1983) Nouvelle étude de la macroflore plio-pleitocène de Crespià (Catalogne, Espagne). Geobios 16:687–715. https://doi.org/10.1016/S0016-6995(83)80087-4
Romanowicz I (1961) Spore and pollen analysis of Tertiary sediments from the vicinity of Bolesławiec and Zebrzydowa. Biul Inst Geol 158:325–409
Salard-Cheboldaeff M (1990) Intertropical African palynostratigraphy from the Cretaceous to Late Quaternary times. J Afr Earth Sci 11:1–24. https://doi.org/10.1016/0899-5362(90)90072-M
Sanz de Galdeano C (1996) Tertiary tectonic framework of the Iberian Peninsula. In: Friend PF, Dabrio CJ (eds) Tertiary bains of Spain, The stratigraphic record of crustal kinematics. Cambridge University Press, Cambridge, pp 9–14
Schuler M (1990) Palynologie et biostratigraphie de l’Eocène et de l’Oligocène inférieur dans le fossés rhénan, rhodanien et de Hesse. Doc BRGM 190:1–503
Stuchlik L, Ziembińska-Tworzydło M, Kohlman-Adamska A, Grabowska I, Słodkowska B, Worobiec E, Durska E (2014) Atlas of pollen and spores of the Polish Neogene. W. Szafer Institute of Botany, Polish Academy of Sciences, Kraków
Suc J-P (1980) Contribution à la connaissance du Pliocene et du Pleistocene Inférieur des regions mediterranéennes d'Europe occidentale par l'analyse palynologique des dépots du Languedoc-Roussillon (sud de la France) et de la Catalogne (nord-est de l'Espagne). Université des Sciences et Techniques du Languedoc, Languedoc
Suc J-P, Cravatte J (1982) Étude palynologique du Pliocène de Catalogne (Nord-Est de l’Espagne). Paleobiol Cont 13:1–31
Suc J-P, Diniz F, Leroy S, Poumot C, Bertini A, Dupont L, Clet M, Bessais E, Zheng Z, Fauquette S, Ferrier J (1995) Zanclean (Brunssumian) to early Piacenzian (early-middle Reuverian) climate from 4° to 54° north latitude (West Africa, West Europe and West Mediterranean areas). Mededel Geol Dienst 52:43–56
Takhtajan A (2009) Flowering plants. Springer, Berlin
Tiffney BH (1986) Fruit and seed dispersal and the evolution of the Hamamelidae. Ann Missouri Bot Gard 73:394–416. https://doi.org/10.2307/2399119
Tiffney BH (2008) Phylogeography, fossils, and Northern Hemisphere biogeography: the role of physiological uniformitarianism. Ann Missouri Bot Gard 95:135–143. https://doi.org/10.3417/2006199
Turland NJ, Wiersema JH, Barrie FR, Greuter W, Hawksworth DL, Herendeen PS, Knapp S, Kusber W-H, Li D-Z, Marhold K, May TW, McNeill J, Monro AM, Prado J, Price MJ, Smith GF (2018) International Code of Nomenclature for algae, fungi, and plants (Shenzhen Code) adopted by the Nineteenth International Botanical Congress Shenzhen, China, July 2017. Regnum Vegetabile 159. Koeltz Botanical Books, Glashutten
Utescher T, Bruch AA, Erdei B, François L, Ivanov D, Jacques FMB, Kern AK, Liu Y-SC, Mosbrugger V, Spicer RA (2014) The Coexistence Approach – Theoretical background and practical considerations of using plant fossils for climate quantification. Palaeogeogr Palaeoclimatol Palaeoecol 410:58–73. https://doi.org/10.1016/j.palaeo.2014.05.031
Valle MF, Alonso Gavilán G, Rivas Carballo MR (1995) Analyse palynologique préliminaire du Miocène dans le NE de la dépression du Duero (aire de Belorado, Burgos, España). Geobios 28:407–412. https://doi.org/10.1016/S0016-6995(95)80017-4
Van Campo E (1989) Flore pollinique du Miocene Superieur de Venta del Moro (Espagne). Acta Palynol 1:9–32
Van der Hammen T, Wymstra TA (1964) A palynological study of the Tertiary and Upper Cretaceous of British Guiana. Leidse Geol Meded 30:183–241
Worobiec E (2009) Middle Miocene palynoflora of the Legnica lignite deposit complex, Lower Silesia, Poland. Acta Palaeobot 49:5–133
Worobiec E (2014) The palynology of late Miocene sinkhole deposits from Upper Silesia, Poland. Rev Palaeobot Palynol 211:66–77. https://doi.org/10.1016/j.revpalbo.2014.09.001
Worobiec E, Gedl P (2018) Upper Eocene palynoflora from Łukowa (SE Poland) and its palaeoenvironmental context. Palaeogeogr Palaeoclimatol Palaeoecol 492:134–146. https://doi.org/10.1016/j.palaeo.2017.12.019
Zetter R (1989) Methodik und Bedeutung einer routinemäßig kombinierten lichtmikroskopischen und rasterelektonenmikroskopischen Untersuchung fossiler Mikrofloren. Courier Forschungsinst Senckenberg 109:41–50
Zetter R, Farabee MJ, Pigg KB, Manchester SR, DeVore ML, Nowak MD (2011) Palynoflora of the late Paleocene silicified shale at Almont, North Dakota, USA. Palynology 35:179–211. https://doi.org/10.1080/01916122.2010.501164
Zhang ZY (2001) Pollen morphology and variation of the genus Fortunearia (Hamamelidaceae) endemic to China. Isr J Pl Sci 49:61–66. https://doi.org/10.1560/TG08-LJ1V-J79P-JF67
Zhang ZY, Lu AM (1995) Hamamelidaceae: Geographic distribution, fossil history and origin. Acta Phytotaxon Sin 33:313–339
Zhang ZY, Zhang H, Endress PK (2003) Hamamelidaceae. In: XXX authors missing! Flora of China. Vol. 9, Pittosporaceae through Connaraceae. Science Press, Beijing, and Missouri Botanical Garden, St Louis, pp. 18–42
Zidianakis G, Mohr BAR, Fassoulas C (2007) A late Miocene leaf assemblage from Vrysses, western Crete, Greece, and its paleoenvironmental and paleoclimatic interpretation. Geodiversitas 29:351–377
Ziembińska-Tworzydło M (1974) Palynological characteristics of the Neogene of western Poland. Acta Palaeontol Polon 19:309–432
Ziembińska-Tworzydło M, Grabowska I, Kohlman-Adamska A, Skawińska K, Słodkowska B, Stuchlik L, Sadowska A, Ważyńska H (1994) Taxonomical revision of selected pollen and spores taxa from Neogene deposits. Acta Palaeobot Suppl 1:5–30
Acknowledgements
We thank Barbara Słodkowska for her assistance with the bibliographic search for this study, Nicholas Turland and Robert Fensome for their guidance in interpreting the International Code of Nomenclature for algae, fungi, and plants, as well as the editor, the reviewer Torsten Utescher and one anonymous reviewer for their useful comments. Ana María Vicente Montaña is thanked for her assistance during the SEM analysis.
Funding
This work has been partly funded by the Spanish project 20788/PI/18, ‘Iberian flora and vegetation during the last 66 Ma’ (Fundación Séneca, Murcia).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflicts of interest to declare that are relevant to the content of this article.
Additional information
Handling editor: Julius Jeiter.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Casas-Gallego, M., Barrón, E. The fossil pollen record of Hamamelidaceae in the Iberian Peninsula. Plant Syst Evol 307, 53 (2021). https://doi.org/10.1007/s00606-021-01772-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00606-021-01772-1