
Abstract
Oxidative stress is an imbalance between oxidants and antioxidants. Chronic exercise can lead to an increase in the production of oxidants and the up-regulation of antioxidant defense mechanisms. In horses, there are several potential exercise-related mechanisms for oxidant production. Athletic horses are exposed to different amounts of stress according to the type of sport and physical exertion. The present study aimed to investigate the effect of type of sport, gender, and age on oxidant/antioxidant and enzymatic status in active athletic horses. Thirty-nine healthy athletic show-jumping, polo, tent-pegging, and normal work horses were evaluated for superoxide dismutase (SOD), catalase (CAT), malondialdehyde (MDA), total antioxidant capacity (TAC), lactate dehydrogenase (LDH), creatine kinase (CK), and alkaline phosphatase (ALP) at rest. A significant positive correlation between SOD and TAC was observed. CK was significantly higher in show-jumping horses compared to tent-pegging horses and higher in mares compared to stallions. ALP was significantly higher in polo horses compared to show-jumping horses. LDH was significantly higher in horses aged 11–14 compared to the 7–10-year group. Our findings support the occurrence of the adaptive response of oxidative/antioxidant status because of regular training in different equestrian sports disciplines, and stallions, mares, and geldings show similar adaptations to exercise-induced oxidative stress. Show-jumping seemed to cause more muscle damage than tent-pegging, especially in mares. A good physical condition may be observed in horses aged between 11 and 14 years.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Oxidative stress has been recently linked to a variety of systemic disorders (Yehia et al. 2020). It occurs because of an imbalance between antioxidants and oxidants. Antioxidant defense mechanisms can be divided into enzymatic and nonenzymatic groups. An enzymatic group, such as superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GPX). Nonenzymatic ones such as vitamins C and E. Antioxidants are essential in disabling harmful metal ions and their role in producing free radicals (Martinovic et al. 2009; Hadžović-Džuvo et al. 2014). Free radicals and reactive oxygen species (ROS) are produced as part of cellular metabolic processes. The harmful effects of free radicals are caused by the need to maintain electronic stability, so they react with the next stable molecule, taking its electron and forming a new free radical (Hadžović-Džuvo et al. 2014). Although the harmful effects of ROS, low levels of them are necessary for important physiological functions such as cell signaling, apoptosis, and immune response (Hadžović-Džuvo et al. 2014; Shankar et al. 2014).
Chronic exercise causes an increase in the production of oxidants, which leads to the up-regulation of antioxidant defense mechanisms (SOD, CAT, and GPx) in humans (Hadžović-Džuvo et al. 2014; D’Angelo and Rosa 2020). SOD and superoxide anion were the best indicators between volleyball player groups with different training experiences (Martinovic et al. 2009).
In horses, exercise has been shown to cause oxidative stress, which induces muscle damage and higher levels of lipid peroxidation (Kent et al. 2022). Several potential mechanisms exist for exercise-related oxidant production (Deaton and Marlin 2003). Horses have a special ability among sports animals to enhance their oxygen uptake by a factor of 60 during intense activity, which may boost ROS release during mitochondrial electron transfer. Regular training in equine athletes was found to stimulate the antioxidant defense mechanism (Andriichuk et al. 2016). There are possible pathways thought to be related to exercise-induced oxidative stress. (1) Inflammatory cell activation due to exercise-related injury and the subsequent production of ROS. (2) Exercise-induced hyperthermia causes an increase in lipid peroxidation in plasma and glutathione oxidation in red blood cell hemolysate. (3) The formation of superoxide due to anaerobic respiration of the hypoxic muscle fiber during stressful exercise (Deaton and Marlin 2003).
Trained horses had a significantly higher antioxidant barrier than untrained horses. As a result, frequent training helps horses adapt to larger amounts of oxidative substances during exercise, making them more resistant to oxidative stress and influencing oxidant/antioxidant balance (Fazio et al. 2016).
Creatinine kinase (CK) is a characteristic enzyme of muscular disorders that catalyzes the transfer of phosphate from phosphocreatine to ADP. Lactate dehydrogenase (LDH) is another enzyme found in muscle tissue that catalyzes the reversible conversion of pyruvate to lactate during glycolysis (Janicki et al. 2013). The intensity and duration of exercise are important factors in the onset of physiological fatigue. Muscle tissue may be damaged as a result of both metabolic and mechanical factors following intense prolonged training. As a result, this damage increases the permeability of the muscle membrane, which can result in the release of CK and all of the muscle damage biomarkers. (Mami et al. 2019).
Different anatomical structures could be exposed to a different amount of stress according to the type of sport and the amount and type of physical exertion (Murray et al. 2006; Bertuglia et al. 2014). The present study aimed to investigate the effect of type of sport, gender, and age on oxidant/antioxidant and enzymatic status in active athletic horses and to address the usefulness of these biomarkers in monitoring the health status of athletic horses.
Material and methods
Ethical statement
This study was conducted on horses belonging to a private governmental sector and was approved by the Institutional Animal Care and Use Committee (IACUC) of Veterinary Medicine, Cairo University, Certificate number Vet cut 20092022462.
Animal selection
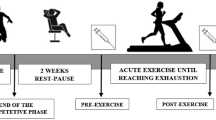
Thirty-nine apparently healthy athletic horses (12 stallions, 14 geldings, and 13 nonpregnant mares) in the period between January 2020 and January 2021 were evaluated for oxidative stress biomarkers and enzymatic activities at rest. All clinical data (including animal description, management history, previous disease history, and treatment strategies) were collected.
The inclusion criteria included horses with an average age of 7–20 years that played (7) show-jumping, (7) polo, (15) tent-pegging, and (10) normal work for at least more than 3 years, and all horses were kept in the same equestrian club, stable, and trained on the same track.
All horses were housed in the same stable in individual boxes (4.00 × 4.00 m). The diet consisted of barely, soybean meal (SBM), wheat bran, alfalfa hay, bean husks, molasses, limestone (CaCo3), and salt, which was administered three times daily (at 7 AM, 1 PM, and 7 PM). Feed was analyzed by Regional Center for Food & Feed (RCFF), Giza Governorate, Egypt. The composition and the type of offered diet adapted to provide crude protein 12.6% DM, crude fiber 22.6% DM, and fats 4.39% DM. Water was available ad libitum.
All horses were receiving moxidectin PO (Equest®). Administer gel by inserting the syringe applicator into the animal’s mouth depositing the gel in the back of the mouth near the base of the tongue. Once the syringe is removed, the horse’s head is raised to insure proper swallowing of the gel.
Horses were assembled according to age into 4 groups, 7–10 years (7 horses), 11–14 years (9 horses), 15–17 years (13 horses), and > 18 years (10 horses) to estimate the differences in enzymatic activities and oxidative stress biomarkers between age groups.
Blood sampling and biochemical analysis
One blood sample was collected from each animal. A single examiner performed the blood sampling of 39 active athletic horses during the study period. Blood was collected from a jugular vein via vein puncture. Blood samples were obtained for serum separation in plain tubes and on heparin anticoagulant for plasma and erythrocyte lysate preparation, which was obtained via centrifugation at 4000 rpm for 10 min, and the plasma was collected. The cells were rinsed three times with four volumes of cold saline, and the resulting red cell pellets were lysed by adding four volumes of cold deionized water to the estimated pellet volume. Centrifugation at 4000 rpm for 10 min at 4 °C was used to remove the red cell stroma. The resulting clarified supernatant was utilized to evaluate SOD using colorimetric test kits (Bio-Diagnostic Company, Egypt) according to the manufacturer’s instructions (Paglia and Valentine 1967; Nishikimi et al. 1972), respectively. Samples were assayed on the same day. Serum was used to estimate serum parameters TAC and ALP using commercial colorimetric kits (Bio-Diagnostic, Egypt), CK, and LDH using enzymatic kinetic specified test kits using Bio-Diagnostic (Egypt) and Salucea (Italy), respectively.
CAT and MDA levels in heparinized plasma samples were determined using corresponding test kits (Bio-Diagnostic Company, Egypt) and made according to the manufacturer’s instructions (Aebi 1984; Begenik et al. 2013).
Statistical analysis
Data are subjected to analysis using SPSS (version 25.0, 2016). Simple one-way ANOVA was used to study the effect of kind of sport, age, and gender on oxidative stress and blood biochemical parameters. Duncan’s multiple range test was used to differentiate between significant means at P < 0.05. Pearson’s correlation coefficient was used to correlate variables with each other.
Results
There was a significant positive correlation between SOD and TAC (r = 0.36; P = 0. 0.026).
Oxidative stress biomarkers and enzymatic activities in different kinds of sports are shown in Table 1. Mean CK concentration was significantly higher in show-jumping compared to tent-pegging horses. Mean ALP concentration was significantly higher in polo horses compared to show-jumping horses. There was no significant difference in mean SOD, CAT, MDA, TAC, and LDH between show-jumping, polo, tent-pegging, and normal work horses.
Oxidative stress biomarkers and enzymatic activities in different genders are shown in Table 2. Mean CK concentration was significantly higher in mares compared to stallions. There was no significant difference in mean SOD, CAT, MDA, TAC, ALP, and LDH between males, females, and geldings.
Oxidative stress biomarkers and enzymatic activities in different age groups are shown in Table 3. Mean LDH concentration was significantly higher in group 11–14-year horses compared to 7–10-year horses. There was no significant difference in means SOD, CAT, MDA, TAC, CK, and ALP between different age groups.
Discussion
Different studies investigated the differences in oxidative stress parameters in athletic horses before and after exercise (Andriichuk et al. 2016; Fazio et al. 2016; Ono et al. 1990; Williams et al. 2005; Kirschvink et al. 2006). The impact of exercise on oxidative/antioxidant balance is conflicting in the previous literature. In our study, we evaluated the oxidative stress status in equine athletes involved in different kinds of equestrian disciplines, including tent-pegging, polo, and show-jumping.
The correlation results between the serum enzymatic activities and oxidative stress biomarkers revealed a significant positive correlation between SOD and TAC in the current study. In parallel with our findings, positive correlations between TAC and the activities of all antioxidant substances were observed (Fazio et al. 2016). TAC is the sum of the total antioxidant capacity of the nonspecific polo of antioxidants, which includes nonspecific antioxidants, metal chelators, and antioxidant enzymes (GPX, CAT, and SOD) (Finsterer 2012).
Also, our findings revealed that there was no significant difference in mean SOD, CAT, MDA, and TAC between horse sports groups. In line with our findings, no change in SOD or CAT activity pre- and post-exercise was also observed in thoroughbred racehorses (Ono et al. 1990). In previous studies, glutathione peroxidase and total glutathione levels decreased in endurance horses after an 80 km (Hargreaves et al. 2002) and a 140 km race (Marlin et al. 2002). Plasma MDA levels in Marwari horses increased immediately after exercise and then significantly decreased after 10 days, indicating the activation of the antioxidant defense mechanism due to moderate exercise (Dedar et al. 2017). Contradicting our findings, a significant increase in SOD and CAT activity and also a decrease in lipid peroxidation activity (MDA) due to regular exercise have been previously observed (Andriichuk et al. 2016; Fazio et al. 2016). The conflicting findings in different previous reports may be explained by the variation in breeds, gender, type of exercise, and amount of training load (Andriichuk et al. 2016; Kirschvink et al. 2006). So, our findings support the occurrence of the adaptive response of oxidative/antioxidant status due to regular training in different equestrian sports disciplines (Andriichuk et al. 2014). The oxidative stress-induced adaptation process of exercise is based on reduced oxidative stress as a result of improved antioxidant defenses, decreased basal oxidant generation, and reduced radical leak during oxidative phosphorylation (Andriichuk et al. 2014).
There was no significant difference in mean SOD, CAT, MDA, and TAC between different sex groups. Similarly, it has been shown that males and females show similar exercise-induced adaptations in antioxidant defenses at the same absolute workload (Andriichuk et al. 2014; Pepe et al. 2009). Also, no difference between men and women in SOD and CAT activities at different running distances over time was reported due to a nonsignificant gender–time interaction (Pepe et al. 2009). There were no gender-based differences in lipid peroxidation or muscle damage levels caused by vigorous exercise (Kaikkonen et al. 2002). On the contrary, total SOD and CAT activity were significantly higher in physically active male rats than in physically active female rats (Yamamoto et al. 2002). Our findings support that stallions, mares, and geldings show similar adaptations to exercise-induced oxidative stress over time.
Concerning the effect of sport type, gender, and age on enzymatic activities in this study, LDH concentration was significantly higher in group 11–14-year horses compared to 7–10-year horses but remained within the reference range. LDH levels in horses at rest gradually decreased as the animals’ training condition advanced, and a relatively low increase in LDH occurred in more hard exercises if the animals were in good physical condition (Santos et al. 2015). Contrary to our results, LDH did not show significant age-related changes in Arabian mares, Bulgarian, and Murinsulaner horses (Gurgoze and Icen 2010; Prvanović Babić et al. 2019; Popova et al. 2020).
Higher CK activity in show-jumping horses compared to tent-pegging horses and significantly higher CK in mares compared to stallions was observed. All CK results are within the reference values previously reported by Knottenbelt and Malalana (2014). Physical exercise has been proven to produce physiological responses to meet increased metabolic demands during activities (Andriichuk et al. 2014). Show-jumping training with obstacles of 1 m triggered a metabolic change in CK (Carvalho et al. 2019). In the same line, significant increases in CK and LDH levels were observed in athletic mares after exercise, and no changes were detected in stallions (Filippo et al. 2016). Higher CK activity in mares may be attributed to more muscle damage than in stallions. The type, intensity, and duration of exercise all influence the profile of increased CK activity in the serum (Janicki et al. 2013). CK activity could be part of a pre-race evaluation of the horse’s suitability to compete in that race (Williams et al. 2005).
ALP concentration was significantly higher in polo horses compared to show jumpers in the present study. ALP levels were found to be lower than the previously reported reference range (Knottenbelt and Malalana 2014; Satué et al. 2022). Different studies reported that ALP decreases with age (Gurgoze and Icen 2010; Muñoz et al. 2012). There was no significant difference in ALP results between the exercising and resting groups or pre- and post-exercise samples in Marwari horses (Dedar et al. 2017). The clinical significance of low serum ALP levels is unknown, so practitioners may underestimate them (Riancho-Zarrabeitia et al. 2016). Low ALP activity could be caused by vitamin C, zinc, and magnesium deficiency (Schmidt et al. 2021). Zinc and magnesium are necessary for adequate ALP activity in serum; so, low levels of these essential minerals may cause a reduction in ALP (Nanji 1982).
Conclusions
Our findings support the occurrence of the adaptive response of oxidative/antioxidant status due to regular training in different equestrian sports disciplines, and stallions, mares, and geldings show similar adaptations to exercise-induced oxidative stress over time. Show-jumping seemed to cause more muscle damage than tent-pegging, especially in mares. A good physical condition may be observed in horses aged between 11 and 14 years. A further understanding of the usefulness of oxidative stress biomarkers for monitoring the health status of athletic horses still needs more investigations on a larger number of athletic horses with different types of sports, genders, and age ranges.
Data accessibility
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Code availability
Not applicable.
References
Aebi H (1984) Catalase in vitro. Methods Enzymol. Academic Press, New York, pp 121–126
Andriichuk A, Tkachenko H, Kurhaluk N (2014) Gender differences of oxidative stress biomarkers and erythrocyte damage in well-trained horses during exercise. J Equine Vet Sci 34(8):978–985. https://doi.org/10.1016/j.jevs.2014.05.005
Andriichuk A, Tkachenko H, Tkachova I (2016) Oxidative stress biomarkers and erythrocytes hemolysis in well-trained equine athletes before and after exercise. J Equine Vet Sci 36:32–43. https://doi.org/10.1016/j.jevs.2015.09.011
Begenik H, Soyoral YU, Erkoc R, Emre H, Taskın A, Tasdemir M, Aslan M (2013) Serum malondialdehyde levels, myeloperoxidase and catalase activities in patients with nephrotic syndrome. Redox Rep 18(3):107–112. https://doi.org/10.1179/1351000213y.0000000048
Bertuglia A, Bullone M, Rossotto F, Gasparini M (2014) Epidemiology of musculoskeletal injuries in a population of harness Standardbred racehorses in training. BMC Vet Res 10(1):1–10
Carvalho WP, Fonseca LA, Girardi FM, Bento LD, Souto PC, Orozco AM (2019) Serum amyloid A and muscle activity biomarkers in horses submitted to equestrian show jumping. Pesqui Vet Bras 39:668–671. https://doi.org/10.1590/1678-5150-PVB-6218
D’Angelo S, Rosa R (2020) Oxidative stress and sport performance. Sport Sci 13:18–22
Dedar R, Kumar V, Bala P, Legha R, Tripathi B (2017) Oxidative stress and hemato-chemical biochemical indices in Marwari horses during moderate intensity field exercise. Vet Pract 22
Deaton CM, Marlin DJ (2003) Exercise-associated oxidative stress. Clin Tech Equine Pract 2(3):278–291. https://doi.org/10.1053/S1534-7516(03)00070-2
Di Filippo PA, Martins LP, Meireles MAD, de Lannes ST, Peçanha RMS, Graça FAS (2016) Gender differences–induced changes in serum hematologic and biochemical variables in mangalarga marchador horses after a marcha gait competition. J Equine Vet Sci 43:18–22. https://doi.org/10.1016/j.jevs.2016.04.093
Dos Santos VP, Gonzalez FD, da Costa Castro Jr JF, Castro TF (2015) Hemato-biochemical response to exercise with ergometric treadmill, mount training and competition in jumping horses. Arch Vet Sci 20(2). https://doi.org/10.5380/avs.v20i2.36617
Fazio F, Cecchini S, Saoca C, Caputo AR, Lancellotti M, Piccione G (2016) Relationship of some oxidative stress biomarkers in jumper horses after regular training program. J Equine Vet Sci 47:20–24. https://doi.org/10.1016/j.jevs.2016.07.014
Finsterer J (2012) Biomarkers of peripheral muscle fatigue during exercise. BMC Musculoskelet Disord 13(1):1–13. https://doi.org/10.1186/1471-2474-13-218
Gurgoze SY, Icen H (2010) The influence of age on clinical biochemical parameters in pure-bred Arabian mares. J Equine Vet Sci 30(10):569–574. https://doi.org/10.1016/j.jevs.2010.09.006
Hadžović-Džuvo A, Valjevac A, Lepara O, Pjanić S, Hadžimuratović A, Mekić A (2014) Oxidative stress status in elite athletes engaged in different sport disciplines. Bosn J Basic Med Sci 14(2):56
Hargreaves BJ, Kronfeld DS, Waldron JN, Lopes MA, Gay LS, Saker KE, Cooper WL et al (2002) Antioxidant status of horses during two 80-km endurance races. J Nutr 132(6):1781S-1783S. https://doi.org/10.1093/jn/132.6.1781S
Janicki B, Kochowicz A, Buzała M, Krumrych W (2013) Variability of selected biochemical parameters in young stallions during the 100-day performance test. J Equine Vet Sci 33(12):1136–1141. https://doi.org/10.1016/j.jevs.2013.04.017
Kaikkonen J, Porkkala-Sarataho E, Tuomainen T, Nyyssönen K, Kosonen L, Ristonmaa U, Lakka H et al (2002) Exhaustive exercise increases plasma/serum total oxidation resistance in moderately trained men and women, whereas their VLDL+ LDL lipoprotein fraction is more susceptible to oxidation. Scand J Clin Lab Invest 62(8):599–607. https://doi.org/10.1080/003655102764654330
Kent E, Coleman S, Bruemmer J, Casagrande RR, Levihn C, Romo G, Herkelman K et al (2022) Comparison of an antioxidant source and antioxidant plus BCAA on athletic performance and post exercise recovery of horses. J Equine Vet Sci 104200
Kirschvink N, de Moffarts B, Farnir F, Pincemail J, Lekeux P (2006) Investigation of blood oxidant/antioxidant markers in healthy competition horses of different breeds. Equine Vet J 38(S36):239–244. https://doi.org/10.1111/j.2042-3306.2006.tb05546.x
Knottenbelt DC, Malalana F (2014) Plasma and serum biochemistry. In: Saunders Equine Formulary 2nd edn. Elsevier Health Sciences. pp 10–22
Mami S, Khaje G, Shahriari A, Gooraninejad S (2019) Evaluation of biological indicators of fatigue and muscle damage in arabian horses after race. J Equine Vet Sci 78:74–78. https://doi.org/10.1016/j.jevs.2019.04.007
Marlin DJ, Fenn K, Smith N, Deaton CD, Roberts CA, Harris PA, Dunster C et al (2002) Changes in circulatory antioxidant status in horses during prolonged exercise. J Nutr 132(6):1622S-1627S. https://doi.org/10.1093/jn/132.6.1622s
Martinovic J, Dopsaj V, Dopsaj M, Kotur-Stevuljevic J, Vujovic A, Stefanovic A, Nesic G (2009) Long-term effects of oxidative stress in volleyball players. Int J Sports Med 30(12):851–856. https://doi.org/10.1055/s-0029-1238289
Muñoz A, Riber C, Trigo P, Castejón F (2012) Age-and gender-related variations in hematology, clinical biochemistry, and hormones in Spanish fillies and colts. Res Vet Sci 93(2):943–949. https://doi.org/10.1016/j.rvsc.2011.11.009
Murray R, Dyson S, Tranquille C, Adams V (2006) Association of type of sport and performance level with anatomical site of orthopaedic injury diagnosis. Equine Vet J 38(S36):411–416. https://doi.org/10.1111/j.2042-3306.2006.tb05578.x
Nanji A (1982) Decreased serum alkaline phosphatase activity in hypothyroidism: possible relationship to low serum zinc and magnesium. Clin Chem 28(7):1711–1712
Nishikimi M, Rao NA, Yagi K (1972) The occurrence of superoxide anion in the reaction of reduced phenazine methosulfate and molecular oxygen. Biochem Biophys Res Commun 46(2):849–854. https://doi.org/10.1016/s0006-291x(72)80218-3
Ono K, Inui K, Hasegawa T, Matsuki N, Watanabe H, Takagi S, Hasegawa A et al (1990) The changes of antioxidative enzyme activities in equine erythrocytes following exercise. Jpn J Vet Sci 52(4):759–765. https://doi.org/10.1292/jvms1939.52.759
Paglia DE, Valentine WN (1967) Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J Lab Clin Med 70(1):158–169
Pepe H, Balci ŞS, Revan S, Akalin PP, Kurtoğlu F (2009) Comparison of oxidative stress and antioxidant capacity before and after running exercises in both sexes. Gend Med 6(4):587–595. https://doi.org/10.1016/j.genm.2009.10.001
Popova M, Malinova R, Nikolov V, Georgiev B, Taushanova P, Ivanova M (2020) Influence of the breed and age on hematological and biochemical indicators of mares from purebred Arabian and Eastbulgarian breeds. Sci Papers: Ser D, Anim Sci-Int Sess Sci Commun Fac Anim Sci 63(1)
Prvanović Babić N, Kostelić A, Novak B, Šalamon D, Tariba B, Maćešić N, Karadjole T et al (2019) Reference values and influence of sex and age on hemogram and clinical biochemistry in protected and endangered Murinsulaner horses. Vet Arh 89(1):25–42. https://doi.org/10.24099/vet.arhiv.0479
Riancho-Zarrabeitia L, García-Unzueta M, Tenorio JA, Gómez-Gerique JA, Pérez VLR, Heath KE, Lapunzina P et al (2016) Clinical, biochemical and genetic spectrum of low alkaline phosphatase levels in adults. Eur J Intern Med 29:40–45. https://doi.org/10.1016/j.ejim.2015.12.019
Satué K, Miguel-Pastor L, Chicharro D, Gardón JC (2022) Hepatic enzyme profile in horses. Animals 12(7):861
Schmidt T, Schmidt C, Amling M, Kramer J, Barvencik F (2021) Prevalence of low alkaline phosphatase activity in laboratory assessment: is hypophosphatasia an underdiagnosed disease? Orphanet J Rare Dis 16(1):1–8. https://doi.org/10.1186/s13023-021-02084-w
Shankar K, Mehendale HM (2014) Oxidative stress. In: Encyclopedia of Toxicology (Third Edition, (ed) Wexler. Academic Press, Oxford, pp 735–737
Williams C, Kronfeld D, Hess T, Saker K, Waldron J, Crandell K, Harris P (2005) Comparison of oxidative stress and antioxidant status in endurance horses in three 80-km races. Comp Exerc Physiol 2(3):153–157. https://doi.org/10.1079/ECP200562
Yamamoto T, Ohkuwa T, Itoh H, Sato Y, Naoi M (2002) Effect of gender differences and voluntary exercise on antioxidant capacity in rats. Comp Biochem Physiol C Toxicol Pharmacol 132(4):437–444. https://doi.org/10.1016/s1532-0456(02)00097-2
Yehia SG, Ramadan ES, Megahed EA, Salem NY (2020) Effect of parity on metabolic and oxidative stress profiles in Holstein dairy cows. Vet. World 13(12):2780
Author information
Authors and Affiliations
Contributions
All authors contributed to the conception, design, and drafting of the study.
Corresponding author
Ethics declarations
Funding
This study was not supported by any funding.
Conflict of interest
The authors declare that they have no conflict of interest.
Ethics approval
This study was conducted on horses belonging to a private governmental sector. This study was approved by Institutional Animal Care and Use Committee (IACUC) of Veterinary Medicine, Cairo University, Certificate number Vet cut 20092022462.
Informed consent
For this type of study informed consent is not required.
Consent for publication
For this type of study consent for publication is not required.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Limitations
The study was conducted on a small number of horses. Further studies are needed on a larger number.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Zohier, A.S., Baraka, T.A., Abdelgalil, A.I. et al. Oxidative stress biomarkers status in selected equine sports. Comp Clin Pathol 32, 461–466 (2023). https://doi.org/10.1007/s00580-023-03457-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00580-023-03457-2