
Abstract
Vertical ectomycorrhizal (ECM) community composition was assessed on silver fir (Abies alba Mill.) in beech-silver fir forests in Bosnia and Herzegovina. Organic and upper mineral horizons were described by pedological analyses. Silver fir root tips were divided into vital ECM, old and non-mycorrhizal for each horizon separately. Morpho-anatomical classification of vital ECM root tips with an assessment of abundance was followed by ITS-based molecular characterization and classification into exploration types. The percentage of vital ECM root tips was not affected by the soil horizon. Altogether, 40 ECM taxa were recorded. Several taxa have not previously been reported for silver fir: Hebeloma laterinum, Inocybe fuscidula, I. glabripes, Lactarius acris, L. albocarneus, L. blennius, L. fluens, Ramaria bataillei, Russula badia, R. lutea, R. mairei, Sistotrema sp., Tarzetta catinus, Tomentella atroarenicolor, T. badia, T. cinerascens, T. bryophylla, and T. ramosissima, indicating high potential for diversity of ECM fungi in silver fir stands. No significant differences in community composition and species richness and diversity were detected between mineral and organic horizons. Community composition was affected by CaCO3, organic carbon concentration, organic carbon stock, total nitrogen stock, C/N ratio and soil bulk density. No significant effects of soil parameters were detected for exploration types. The contact exploration type was dominant in both soil horizons. Significantly different relative abundances of dominant taxa Tomentella stuposa, Cenococcum geophilum and Piloderma sp. 1 were detected in the two horizons. Twelve taxa were limited to the organic horizon and eight to the mineral horizon.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Studies to date of ectomycorrhizal (ECM) communities of silver fir have reported a total of 85 ECM taxa from silver fir in its native range and plantations outside the native range in eastern (Kowalski 2008; Rudawska et al. 2016; Ważny 2014; Ważny and Kowalski 2017), southern (Comandini et al. 2004; Comandini et al. 1998, 2001; Laganà et al. 2002) and central Europe (Cremer 2009; Schirkonyer 2013) but have not investigated any relationship between soil characteristics and community composition. Chemical and mineralogical properties of soils change with depth, creating a number of different habitats for microorganisms (Rosling et al. 2003).
Generally, studies investigating ECM fungal communities in both organic and mineral horizons are largely absent, probably due to the fact that the majority of fine root length is located in the organic horizon (e.g. Brunner and Godbold (2007)). In studying organic and mineral horizons as composite samples, we might miss important ecological information, such as niche differentiation and functional role of separate ECM taxa, while including only organic profiles may lead us to miss species richness of the site. According to Rosling et al. (2003), 50% of species richness can be attributed to mineral horizons in boreal coniferous forests. A few studies that differentiated between organic and mineral horizons in boreal ecosystems (Rosling et al. 2003; Tedersoo et al. 2003) and alpine forests (Baier et al. 2006; Scattolin et al. 2008) have revealed that several taxa might be restricted to mineral horizons of soil or at least show some preference for either organic or mineral horizons. Edaphic conditions structure the ECM community, resulting in very different ECM communities between organic and mineral horizons (Dickie et al. 2002; Rosling et al. 2003; Tedersoo et al. 2003). Chemical properties of the mineral soil were analysed, but no relationship was found between the species composition and the chemical properties of the mineral horizons in which they occurred in boreal Picea abies–Pinus sylvestris forest (Rosling et al. 2003). However, for an alpine Picea abies forest in Italy, it was demonstrated that ECM community composition is influenced by soil type, pH and nitrogen content (Scattolin et al. 2008). Differences in substrate preference could also be associated with different enzymatic capacities of ECM fungi to break down various substances or spreading strategies (Tedersoo et al. 2003).
As one of the representatives of Pinaceae, silver fir has very thick fine roots (Mrak and Gričar 2016). This results in low total length of roots per unit soil volume (Fitter 2002) and is supposed to be a conservative strategy beneficial in C-limited environments (e.g. shade) (Bauhus and Messier 1999) and soils with a near-homogeneous supply of nutrients, which could be created by recalcitrant coniferous leaf litter that accumulates over many years (Chen et al. 2016). Due to the thickness of their fine roots, ECM fungi might be highly important for silver fir nutrition because of their ability to take up nutrients and water from the root-inaccessible parts of soil through their small-diameter hyphae (Finlay 2008). Furthermore, different exploration types of ECM fungi may support silver fir nutrition to different extents, based on the presence or absence of extramatrical mycelium and physico-chemical soil characteristics. Extramatrical mycelium is a physical and functional extension of the root system (Duddridge et al. 1980; Finlay 2008) and increases the surface area across which nutrients can be taken up. Due to their small diameter, hyphae are able to penetrate soil microsites that are inaccessible to plant roots (Finlay 2008). In this way, extramatrical mycelium improves the uptake of mineral nutrients that are already in the soil solution as well as mobilizes nutrients from organic substrates, i.e. structural and other polymers (Finlay 2008). Variations in structure, function and activity of the extramatrical mycelium determine the potential resource utilization capacities of different fungal taxa (Kuyper and Landeweert 2002). Classification of ectomycorrhizae into exploration types (contact, short-distance, medium-distance: smooth, fringe and mat subtypes, long-distance), may help in the recognition of their distinct ecological functions (Agerer 2001). While some studies found no association of exploration types with soil horizon (Tedersoo et al. 2003), others have found evidence for it (Baier et al. 2006; Scattolin et al. 2008).
The contact exploration type is characterized by a smooth ectomycorrhizal mantle and only a few emanating hyphae. ECM mantles of the contact exploration types are hydrophilic and grow in close contact with the surrounding substrate (Agerer 2001). The contact exploration type is often associated with mineral horizons (Baier et al. 2006; Scattolin et al. 2008), where direct gain of nutrients via hydrophilic mantles that are in close contact with mineral particles (Scattolin et al. 2008) is beneficial due to the density and narrow pores of mineral soil horizons (Baier et al. 2006). With their lignolytic capabilities, contact exploration types might be able to access organic substances stabilized in Ca and Mg complexes (Baier et al. 2006).
Ectomycorrhizae of the short-distance exploration type are characterized by a voluminous envelope of emanating hyphae (Agerer 2001). They prefer the organic horizon, where they can make numerous contacts with loose organic residue by emanating hyphae (Baier et al. 2006).
Medium-distance exploration type ectomycorrhizae mainly form undifferentiated to slightly differentiated rhizomorphs, while the abundance of emanating hyphae depends on the subtype, with relatively few emanating hyphae in the smooth subtype to fringe and mat subtypes with abundant emanating hyphae (Agerer 2001). The fringe subtype is characterized by greater mycelial density compared with short-distance and long-distance exploration types, which allows for more intense resource gain per occupied space (Weigt et al. 2012).
The long-distance exploration type has rather smooth mantles with few highly differentiated rhizomorphs (Agerer 2001). The absence of cross cell walls in rhizomorphs speeds up transport of water (Duddridge et al. 1980) and solutes. The uptake zone is moved from the direct vicinity of ectomycorrhizae to more distant zones (Agerer 2001). Preference for mineral horizons was also reported for long-distance exploration types (Scattolin et al. 2008).
According to Chen et al. (2018), root diameter of temperate ECM tree species is significantly positively correlated with the proportion of medium-distance ECM fungal taxa and negatively with the proportion of contact ECM fungal taxa, while no correlation was found for short-distance and long-distance taxa. Through selection of an ECM community with appropriate exploration distance, trees would achieve root-fungal complementarity in nutrient foraging (Chen et al. 2018).
The aim of our study was to contribute to knowledge on ECM taxa diversity of silver fir and for the first time characterize the silver fir ECM community from the aspect of exploration type. The study was conducted in Bosnia and Herzegovina, and to our knowledge, it is the first report on ECM diversity for silver fir from this area and the first to study the ECM community composition of silver fir in organic and mineral soil horizons. The Balkan Peninsula is known for its high biodiversity due to its complex physical geography and the existence of refugia for several plant and animal species in the last glacial period (Griffiths et al. 2004), silver fir being one of them (Liepelt et al. 2009). Furthermore, we aimed to compare fine root density, vitality of ECM root tips and ECM communities in mineral and organic horizons and relate ECM community composition to physical-chemical characteristics of soil horizons. We hypothesized that the majority of fine roots, vital ECM root tips and ECM taxa will be located in the organic soil horizon, as this is the most dynamic part of soil. Differences in ECM communities of silver fir were expected between organic and mineral horizons. It was hypothesized that ECM exploration types with abundant emanating structures would dominate the silver fir ECM community to improve nutrient foraging of thick silver fir roots. We posited that exploration types would reflect different physical-chemical soil characteristics of organic and mineral horizons.
Material and methods
Study area and sampling design
The study was conducted in a climax beech-silver fir forest (Omphalodo-Fagetum Tregubov 1957 (Marinček et al. 1993)) at permanent research plots of the Faculty of Forestry of the University of Sarajevo at Bjelašnica Mountain in Bosnia and Herzegovina (Fig. 1), in autumn 2016. The research area is part of the large forested area of the mountains of Bjelašnica and Igman, with uneven aged and naturally regenerated stands due to close-to-nature forest management.

Study area and sampling design on unmanaged forest (UF) and managed forest (MF1 and MF2) plots in beech and silver fir forests at Mt Bjelašnica. Approximate position for examination of ectomycorrhizal community, soil type and soil samples for describing organic versus upper mineral soil on 20 × 20 m plots is shown
General data about the research site is given in Table 1. The prevailing geological substrate is limestone and dolostone, accompanied by soil types such as Leptosol, Rendzic Leptosol and Cambisol. Some properties characterizing these soils are shallowness (depth up to 50 cm), lytic contact with substrate, substantial stoniness (25–50%), high water permeability, susceptibility to drought and mesotrophicity (Topalović and Travar 1978).
Sampling and characterization of organic and mineral soil
The soil homogeneity within each plot was checked by augering down to 1 m (Pürckhauer auger) at four points per plot (Fig. 1). Soil type was determined according to IUSS Working Group WRB (2015) and humus form described (Zanella et al. 2011) at the same points. Depth (cm), structure, pH value, presence of earthworms and texture of top humus enriched mineral horizon (Ah) were determined. Additionally, the depth was measured for cambic horizon (Brz) and mean total soil depth was assessed.
More detailed analyses were conducted to describe the organic and upper mineral soil horizons considering soil factors possibly affecting ECM composition. Nine subsamples were mixed into three composite samples per plot per horizon. Composite samples of the organic (each taken from a 25 × 25 cm area) and mineral soil horizon (taken underneath the organic horizon into a cylinder of 7.0 cm diameter, up to 10 cm) were used for further examination. Thickness (cm), pH in 0.01 M CaCl2 dilution, carbonates (CaCO3) content (Scheibler unit, Eijkelkamp Soil & Water, Giesbeek, Nederland), phosphorous (mg P2O5 100 g−1 by AL method, only in the mineral horizon), potassium (mg K2O 100 g−1 by AL method, only in the mineral horizon), soil organic carbon (SOC), total nitrogen (TN) concentration (Elemental Analyser LECO CNS 2000, St. Joseph, MI, USA), C/N ratio and bulk density of fine earth (BD kg m−3 by gravimetric method, % content of gravel was assessed) were determined. In addition, SOC and TN stocks (kg m−2 × 10 = t ha−1) as the product of the SOC or TN concentration (g kg−1), BD (kg dm−3) and soil horizon depth (m), with a correction factor for rockiness (CFr = 100 − (% stones)/100), were calculated.
Sampling and characterization of ECM communities
Soil analyses for all three plots revealed that general soil properties are not significantly different among the plots and are therefore representative for the area (Table S1), while there was strong differentiation between soil horizons, as presented in ‘Results’.
One dominant silver fir (Abies alba) tree per plot was selected for sampling of ECM roots. Selected trees had minimum diameter at breast height of 50 cm and were surrounded by a clear area without other trees in a radius of 5 m. Five soil cores per plot (7.0 cm diameter) at a fixed depth of 20 cm depth were taken in a circle along the radius of the crown diameter of each tree. Samples were transported on ice and kept in plastic bags at 4 °C in the dark until further processing.
Each soil sample was separated into organic (combined Ol, Of and Oh) and mineral horizons. As one sample contained no silver fir roots, it was discarded, resulting in 14 subsamples for each horizon. The volume of each subsample was determined by submerging the soil subsample into a measuring cylinder filled with a known amount of water. Mean volume of organic and mineral subsamples was 90.4 ± 17.1 cm3 and 257 ± 24.5 cm3, respectively. Roots were cleaned by soaking samples in tap water. All roots longer than 0.5 cm or having at least one branch were separated. Roots of silver fir were distinguished from roots of beech by its greater diameter and structure of periderm (Mrak and Gričar 2016) and divided into vital ECM root tips, old ECM root tips and non-mycorrhizal root tips. Vital ECM were defined as root tips with a turgescent fungal mantle, old ECM as root tips with a non-turgescent or dry fungal mantle and non-mycorrhizal root tips as root tips with no visible ECM fungal mantle. Individual ECM morphotypes of each sample were isolated and categorized according to Agerer (1987–2012). Root tips of individual ECM morphotypes, old ECM roots and non-mycorrhizal roots were quantified with WinRhizo software (Regent Instruments Inc., Ville de Quebec, Canada) after scanning by Epson Perfection V700 Photo scanner (Seiko Epson Corp., Suwa, Nagano, Japan) in trays filled with water. For each morphotype, mantle preparation was prepared and anatomical characteristics determined under the Zeiss Axio Imager Z2 light microscope (Carl Zeiss Microscopy, Jena, Germany) using differential interference contrast following Agerer (1991) and DEEMY (Agerer and Rambold 2004–2017). Total DNA was extracted from a representative root tip for each morphotype from each soil subsample with DNeasy Plant Mini Kit (Qiagen, Hilden, Germany) following manufacturer’s instructions. From isolated DNA, the ITS region of nuclear ribosomal DNA was amplified using the fungus specific primers ITS1f and ITS4 (Gardes and Bruns 1993; White et al. 1990), following the modified procedure described in Sulzbacher et al. (2016). PCR products were run on 1.5% agarose gels in 0.5x TBE buffer and visualized with Gel Doc EQ Systems, PC (BioRad, USA). Amplified DNA fragments were cut out of agarose gels and purified with innuPrep DOUBLEpure Kit (Analytik Jena AG, Jena, Germany) following manufacturer’s instructions. Sequencing of pure DNA was performed by a commercial sequencing laboratory (Macrogen Inc., Seoul, South Korea). All ECM fungal morphotypes were sequenced in both directions with the primers ITS1f and ITS4 (Gardes and Bruns 1993; White et al. 1990), where all sequences were edited using Sequencher v 5.0 (Gene Codes Corp., Ann Arbor, MI, USA). Sequence identity searches were conducted using the BLASTN algorithm at the National Center for Biotechnology Information (NCBI) website (https://blast.ncbi.nlm.nih.gov/Blast.cgi) using default settings (Altschul et al. 1997). Sequences that belonged to saprotrophs or endophytes were discarded. Final criteria for a database match were as follows: query cover ≥ 80% and sequence similarity > 92% (representing approximate cut-off value at genus level) or sequence similarity ≥ 97–100% (representing approximate cut-off at species level) (Porras-Alfaro et al. 2014; Raja et al. 2017). Representative sequences were deposited in GenBank under accession numbers MN818523 to MN818560 (Table S2). Where sequencing was not successful, identification was performed by morphological and anatomical characteristics. In cases where no identification was possible, ECM root tips were classified as unidentified and their abundance was considered in calculations.
Percentages of vital and old ECM root tips were calculated for each subsample. Coverage, estimated species richness and diversity were calculated by SpadeR (Chao et al. 2015) using incidence-frequency data of 14 sampling units per soil horizon.
The relative abundances of individual ECM taxa were expressed as the number of root tips of individual ECM taxa divided by the total number of all ECM root tips. Exploration types were assigned to individual taxa following Agerer (2001, 2006) using our image data and morphotype descriptions, DEEMY and other authors’ descriptions of individual taxa, specifically for Tomentella taxa (e.g. Binder et al. 2013; Jakucs and Eros-Honti 2008; Jakucs et al. 2015, 2005).
Data analysis
Statistical analyses were performed in an R v. 3.5.2 environment (R Core Team 2018), unless otherwise stated.
A t test was used to determine differences between means of the organic vs. mineral soil horizon. Soil variables that did not show normal distribution and homogeneity of variance were sqrt transformed.
One-way ANOVA with soil horizon (organic/mineral) as a single variable and followed by the Tukey post hoc test was conducted in Dell Statistica version 13 data analysis software (Dell Inc., Tulsa, OK, USA) to reveal differences in root tip density, percentage of vital and old ECM root tips and exploration types. Homogeneity of variances was tested by Levene test. Data with P values less than 0.05 were considered as statistically significant.
Multivariate generalized linear models (MV-GLMs; Wang et al. (2012)) were used to examine the differences in ECM species number and abundances between organic and mineral soil horizon by using the ‘glm’ function. Multivariate and unadjusted univariate P-values were obtained by Wald tests, both using 10,000 Monte Carlo permutations. For visualization of ECM species abundances, which showed statistically significant differences in abundances between soil horizons, mean values and standard errors were calculated using the ‘ddply’ function from the package ‘plyr’ (Hadley 2011).
Bray–Curtis dissimilarity matrices were performed for taxa and soil condition data. The Bray–Curtis matrix was based on the relative abundance of each ECM taxon in each of 28 samples to account for differences in the organic vs. mineral horizon. All taxa were included in the analysis except unidentified species. To identify the main factors affecting community composition among soil horizons, nonmetric multidimensional scaling (NMDS) was performed. Soil variables (SOC and TN concentrations, C/N ratio, pH CaCl2, CaCO3 (%), SOC and TN stocks in organic and mineral horizons) as well as intrinsic variables (species driving site distribution pattern) were fitted to the NMDS by using the ‘envfit’ function from the ‘vegan’ package. Each soil variable was normalized. NMDS was also performed for ECM exploration types. Zero values were replaced with 0.001. To test the significance of NMDS and soil horizon we carried out a PERMANOVA test using the Adonis function from the ‘vegan’ package of R (Oksanen et al. 2012).
Results
Soil analysis
Calcic Chromic Cambisol (75%) was a predominant soil type of each plot (Table S1). The described humus form was mainly Oligomull (50%) accompanied by Dysmull (25%) and Eumacroamfi (25%). Soil on limestone and dolostone showed a usual well-built structure, with moderate to strong structured aggregates and presence of living earthworms as an indicator of porosity. The observed soil texture, thickness, pH values, amount of CaCO3, SOC and TN concentration, C/N ratio, BD, SOC stocks and TN stocks in both the organic and mineral horizon indicated mainly similar edaphic conditions among plots.
Organic and upper mineral soil horizons were significantly different in all analysed properties except for soil pH values (Table 2). The organic horizon was characterized by higher SOC and TN content and C/N ratio compared with mineral soil. Conversely, the mineral horizon was characterized by higher values of BD, SOC and TN stocks. Presence of CaCO3 was only observed in the mineral horizon.
Ectomycorrhizal communities
The organic horizon of soil samples contained 9.81 ± 1.99 root tips per cm3 of substrate, while the mineral part contained only 1.75 ± 0.51 root tips per cm3. Consequently, 56.9% of the investigated root tips and more than twice as many vital ECM root tips were found in the organic horizon, although the mean volume of the organic horizon was only 35.2% of the mean volume of the mineral horizon. Old and non-mycorrhizal root tips were more equally distributed between organic and mineral horizons, the organic part containing 52.3% of all old mycorrhizal root tips and 48.7% of all non-mycorrhizal root tips. The percentage of vital ECM root tips was 22.2 ± 3.94% and 12.8 ± 2.88% for the organic and mineral horizon, respectively, while the percentage of old ECM root tips was 52.9 ± 4.94% and 54.3 ± 5.13% for the organic and mineral horizon, respectively. However, the soil horizon had no significant effect on the percentage of vital (F = 3.71, P = 0.0651) and old (F = 0.04, P = 0.8468) ECM root tips. Morphotyping was performed on all vital ECM root tips, i.e. 10,543 from the organic and 5008 from the mineral horizon. A total of 407 out of 10,543 ECM root tips remained unidentified in the organic horizon, and 471 out of 5008 in the mineral horizon, but their abundance was considered in all analyses.
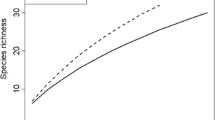
In the organic horizon, 77.0% of estimated diversity was recovered, and 57.9% in the mineral horizon. Estimated species richness (Chao 2) was 42.8 ± 7.11 for the organic horizon and 46.8 ± 11.9 for the mineral horizon, while the Shannon diversity index was 3.23 ± 0.10 for the organic and 3.25 ± 0.13 for the mineral horizon.
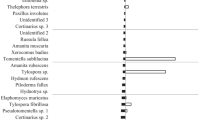
A total of 40 taxa were recorded for both soil horizons, with 32 being found in the organic and 28 in the mineral horizon (Fig. 2). Images and short morpho-anatomical descriptions are provided in Table S3. Taxonomically dominant orders were Russulales with 13 taxa, Thelephorales with 8 taxa and Agaricales with 8 taxa. Only three taxa out of 40 belonged to Ascomycota: Cenococcum geophilum, Tarzetta catinus and Tuber sp.

Comparison of ECM community structure of silver fir in organic and mineral soil horizons at Bjelašnica (BIH). Mean ± stderr values are shown, considering relative abundances in 14 samples for each soil horizon
Only six and nine taxa, respectively, were present with more than 5% mean relative abundance in organic and mineral horizons (Figs. 2 and 3): Tomentella stuposa, Russula sp. 1, Lactarius blennius, Clavulina cristata, Russula badia and Cenococcum geophillum for the organic horizon and Russula badia, Clavulina cristata, Piloderma sp. 1, Tarzetta catiunus, Russula sp. 1, Lactarius aurantiacus, Lactarius acris, Inocybe glabripes and Russula amethystina for the mineral horizon (Fig. 2). The most dominant ectomycorrhiza in the organic horizon was T. stuposa (9.83 ± 4.00%), while R. badia was dominant (7.93 ± 7.13%) in the mineral horizon. Among taxa with more than 5% relative abundance in the organic horizon, C. geophilum occurred in 57.1% (8/14) of samples, T. stuposa in 35.7% (5/14) and L. blennius, Russula sp. 1, R. badia and C. cristata in 14.3% (2/14) each. On the other hand, taxa with more than 5% abundance in the mineral horizon were present in a smaller percentage of samples (one or two out of 14), but with greater abundance there. Among taxa with more than 5% mean relative abundance in the organic horizon, only L. blennius occurred exclusively in the organic horizon, while among dominant taxa in the mineral horizon, I. glabripes, L. acris and T. catinus occurred exclusively in this soil horizon.

Selected dominant ECM taxa and their exploration types of silver fir in organic (a–c) and mineral horizon (d–f). aTomentella stuposa, medium-distance smooth exploration type. bRussula sp. 1, contact exploration type. cLactarius blennius, contact exploration type. dRussula badia, contact exploration type. eClavulina cristata, contact exploration type. fPiloderma sp. 1, medium-distance fringe exploration type
Among ECM taxa present in both soil horizons, significant differences in relative abundances of Amphinema byssoides, Boletus pruinatus, C. geophilum, Piloderma sp. 1, Tomentella ramosissima and T. stuposa were detected (Fig. 4; Table S4). Relative abundances of B. pruinatus and Piloderma sp. 1 were significantly higher in the mineral horizon, while those of A. byssoides, C. geophilum, T. ramosissima and T. stuposa were significantly higher in the organic horizon.

ECM taxa of silver fir from both soil horizons whose mean relative abundances differed significantly at P < 0.05 (MV-GLM), as marked by an asterisk
Of the 8 soil parameters fitted in NMDS, CaCO3, organic carbon, organic carbon stock, total nitrogen stock, C/N ratio and soil bulk density were significantly related to community composition (Fig. 5; Table 3).

Fungal community composition of Abies alba in organic and mineral soil horizons at Bjelašnica, BIH, displayed using NMDS with 95% confidence ellipses. Stress of the ordination is 0.1641. Variables used in NMDS are shown by arrows whose length is proportional to the strength of the correlation (Table 3). Soil factors were pH value, CaCO3 content, soil organic carbon (SOC), total nitrogen (TN), C/N ratio, bulk density of fine earth (BD), SOC stock and TN stock. Intrinsic ECM species (P < 0.001 and P < 0.01) are underlined and shown in bold
Species that may be driving the site distribution pattern, referred to as intrinsic variables, were C. geophilum, Inocybe fuscidula and Tomentella badia (all P < 0.001), followed by L. blennius, Piloderma sp. 2 (both P < 0.01), R. amethystina and Tomentella atroarenicolor (P < 0.05), Table S5.
Exploration types
Taxa in the organic horizon were ascribed to contact (43.8%), short- (28.1%), medium- (25.0%) and long- (3.13%) distance exploration types. In the mineral horizon, the majority of taxa were ascribed to contact and medium-distance exploration types (35.7% each), followed by short- (25.0%) and long-distance (3.57%) exploration types.
When percentages of exploration types were calculated by taking into account mean relative abundances, both soil horizons still had the highest proportion of contact exploration types, followed by the short, medium smooth, medium fringe and long-distance exploration types (Fig. 6). In the mineral horizon, the medium-distance mat exploration type, which was not detected in the organic horizon, was present. The two soil horizons did not differ significantly in proportions of exploration types (Table S6). When all exploration types with emanating hyphae were considered as one group, it accounted for 54.7 ± 9.65% in the organic horizon and 55.0 ± 9.87% in the mineral horizon versus 42.7 ± 9.47% for the contact exploration type in the organic and 38.7 ± 9.80% in the mineral horizon. There were no statistically significant differences between percentages of both exploration type groups by nonparametric test for dependent variables.

Mean ± stderr relative abundance of exploration types of silver fir ectomycorrhizae in organic and mineral horizons. C contact exploration type, SD short-distance exploration type, MS medium-distance smooth exploration type, MF medium-distance fringe exploration type, MM medium-distance mat exploration type, LD long-distance exploration type
Of taxa that had significantly different relative abundances in the two soil horizons and were at the same time the most abundant components of the ECM community, C. geophilum belongs to the short-distance exploration type, Piloderma sp. 1 to the medium-distance fringe exploration type and T. stuposa to the medium-distance smooth exploration type.
Of 8 variables fitted into NDMS ordination, no soil parameter was significantly related to exploration type composition (Fig. S1; Table S7).
Discussion
As expected, greater density of silver fir root tips in our study occurred in the organic soil horizon. This kind of distribution is generally reported and might be related to the higher amount of nutrients available for plant roots and fungi in the organic horizon (Hashimoto and Hyakumachi 1998) as a result of very active processes of mineralization and nitrogen mobilization there (Baier et al. 2006). Similarly, the majority of vital ECM root tips (> 67%) occurred in the organic horizon, as already noted by several authors (e.g. Dahlberg 2001; Baier et al. 2006). However, when considering relative abundances of vital ECM root tips in organic and mineral horizons, we found no significant differences between the two horizons, while Scattolin et al. (2008) reported greater relative abundance of vital ECM root tips with increasing soil depth.
Investigation of silver fir ectomycorrhizae at three locations at Bjelašnica (BIH) resulted in 40 ECM taxa. Several taxa identified in our study have not been previously reported for silver fir, namely Hebeloma laterinum, Inocybe fuscidula, I. glabripes, Lacatrius acris, L. albocarneus, L. blennius, L. fluens, Ramaria bataillei, Russula badia, R. lutea, R. mairei, Sistotrema sp., Tarzetta catinus and several Tomentella species (T. atroarenicolor, T. badia, T. cinerascens, T. bryophylla, T. ramosissima), indicating high potential for diversity of ECM fungi in silver fir stands, as already proposed by Unuk and Grebenc (2017). Due to the limited sampling effort of our study, we suggest that much higher biodiversity of silver fir ectomycorrhizae may be encountered at the location studied. For comparison, 48 morphotypes were recorded in five silver fir forests on ten sampling sites in Italy (Comandini et al. 2001), and 63 taxa in silver fir seedlings regenerating in three mature silver fir forests in the Polish Carpathians, with 30 seedlings sampled per forest with a sampling area 6 ha per forest (Ważny 2014).
The ECM community of both the organic and mineral horizon was composed of a few dominant species and a high number of rare species. This is a typical finding for ECM fungal communities elsewhere (Dahlberg 2001). The ECM community of silver fir in our study was mainly composed of generalist ECM taxa. A predominance of generalist fungi in mixed communities of ECM plants is commonly reported (Horton et al. 2005). The silver fir specialist Lactarius salmonicolor (Rudawska et al. 2016) was present only in the mineral soil horizon, while among species with silver fir preference—L. albocarneus, L. intermedius and Russula cavipes (Rudawska et al. 2016), L. albocarneus was identified in both soil horizons.
In our study, 20.0% of investigated taxa were present exclusively in the mineral soil horizon. In boreal forest ecosystems, the percentage of ectomycorrhizae present exclusively in mineral horizons may be as high as 50% (Rosling et al. 2003). The difference may be associated with the lower depth of sampling and lower sampling effort in our study, as our samples included horizons from 0 to 20 cm, while in the study by Rosling et al. (2003), sampling to a 53-cm depth was performed.
The dominant component of the organic soil horizon in our study, Tomentella stuposa, was reported to be a significant representative of ECM communities in temperate coniferous and broad-leaved forests (Jakucs et al. 2005). T. stuposa was also reported as the most dominant ectomycorrhiza of mature and juvenile silver firs in the Black Forest (Germany) (Cremer 2009) and of silver fir seedlings in the Polish Carpathians (Ważny 2014). In our study, two anatomically distinct types of T. stuposa were recorded: one with bottle-shaped cystidia with a straight or slightly curved neck and one without cystidia (Supplementary material, Table S3). In 100-year-old oak forest in northeastern France, T. stuposa contributed little to functional diversity in A1 and A2 horizons. As functional plasticity at the species level could be high and is related to the wide distribution of some ECM taxa (Rineau and Courty 2011), the role of T. stuposa in silver fir nutrition should be further investigated. It belongs to the medium-distance smooth exploration type shown to be positively correlated with C/N ratio in the study by Suz et al. (2014), which confirms its predominant occurrence in the organic horizon with a higher C/N ratio in our case.
Another dominant component of the organic horizon that also occurred with significantly higher abundance in the organic horizon was Cenococcum geophilum. C. geophilum is often reported to prefer but not limit itself to organic horizons (Rosling et al. 2003; Baier et al. 2006; Scattolin et al. 2008) due to its resistance to fluctuations in environmental conditions, such as soil moisture and temperature (Goodman and Trofymow 1998; Scattolin et al. 2008). C. geophilum was shown to correlate with increasing humus and N content and a higher C/N ratio, which are indicators of the organic horizon (Baier et al. 2006). As a representative of the short-distance exploration type, it can make multiple contacts to organic particles in the organic horizon (Baier et al. 2006).
On the other hand, the mean relative abundance of the most dominant component of the mineral horizon, Russula badia, did not differ significantly between the two soil horizons. As a representative of the contact exploration type with hydrophilic mantles, members of the Russula genus together with Lactarius often predominate in mineral horizons (Baier et al. 2006; Scattolin et al. 2008). However, in our case, Lactarius and Russula together were relatively abundant in the organic horizon, too. These diverse results indicate a need for investigation of ECM fungal communities of the mineral horizon in different soil systems and under different host taxa.
Piloderma sp. 1 was one of the most dominant species of the mineral horizon, whose mean relative abundance in the mineral horizon was significantly higher compared with the organic horizon. Piloderma species are symbiotic with a wide spectrum of conifer and hardwood tree species (Arocena et al. 2001). Piloderma spp. may increase cation exchange capacity (exchangeable Ca, Mg and K) of rhizosphere soil, most likely due to high production of organic acids (Arocena et al. 1999). It was characterized by simple root tip morphology and a thick transparent gelatinous plectenchymatic mantle (Supplementary material, Table S3). Emanating hyphae were covered by spherical warty structures that did not disappear in lactic acid. Presence of Piloderma in silver fir stands of Bjelašnica indicated low N deposition in those stands, as species with costly N-utilizing mechanisms from organic substrates disappear first when N deposition increases (Lilleskov et al. 2011; van der Linde et al. 2018). In the study by van der Linde et al. (2018), all Piloderma species that were negative indicators of N deposition were conifer specialists. Generally, differences in ECM community composition among organic and mineral soil horizons could be associated with differences in humus content, C/N ratio and N content, soil pH and K content (i.e. edaphic characteristics) (Baier et al. 2006). In our case, we could not confirm differences between ECM community composition in the organic and mineral horizons, but overall community composition was significantly related to CaCO3 content of soil, organic carbon, C/N ratio, organic carbon stock, total nitrogen stock and soil bulk density, while no effect on the structure of exploration type was shown for any soil parameter. Effects of CaCO3 on ECM community composition were well demonstrated in liming experiments to prevent acidification of forests (Monfort-Salvador et al. 2015). Although CaCO3 often induces an increase in soil pH (Monfort-Salvador et al. 2015), a higher percentage of CaCO3 in the mineral horizon of our plots was not associated with higher pH values. Besides pH changes, liming is associated with changes in the concentrations of Ca, Al or N (Bakker et al. 2000). Differences in occurrence of morphotypes related to liming were reported as well (Bakker et al. 2000), with a decrease in ectomycorrhizae with smooth mantles in favour of those with hairy mantles (Bakker et al. 2000; Monfort-Salvador et al. 2015). On the other hand, organic C and the C/N ratio was shown to be positively related to ectomycorrhizae with high biomass and presence of rhizomorphs, and total N stock to contact and medium-distance fringe exploration types (Suz et al. 2014). Effects of soil density on ECM communities are best represented by studies of soil compaction due to forestry operations or soil preparation methods in disturbed sites (e.g. Bauman et al. 2013). High soil bulk densities repress hyphal growth to a large extent (Skinner and Bowen 1974). Due to limited sampling effort of our study, including a low number of sampling locations, some patterns might remain hidden or overstated, preventing us from drawing sound conclusions.
The mineral horizon was characterized by high abundance of selected ECM taxa in a small number of samples, indicating a highly patchy distribution of ectomycorrhizae. Such a pattern is common for ECM fungi (Agerer and Göttlein 2003) and has also been reported for other temperate tree (F. sylvatica, Picea abies and Pinus sylvestris) ECM communities, not specifically dealing with vertical distribution (Rosinger et al. 2018). Taxa with high on-site abundance and low across site frequency dominate a niche and competitively exclude other taxa (Rosinger et al. 2018). This pattern may reflect a heterogeneous distribution of nutrients in soil (Agerer and Göttlein 2003), which is a result of patchy litter distribution and activity of animals (Chen et al. 2016), but on the other hand, a heterogeneous distribution of nutrients could be a result of ECM exudation of organic acids leading to acidification of the surrounding substrate and releasing of nutrients (Agerer and Göttlein 2003). Surprisingly, the pattern of patchy ECM distribution was more prevalent in the mineral horizon, which is considered more homogeneous (Duchaufour 1982; Marco et al. 2016), while the organic part is subjected to seasonal changes and disturbances (Bååth and Söderström 1982; Baldrian et al. 2010). Patchy distribution of ectomycorrhizae in the mineral horizon could also result from heterogeneous distribution of fungal inoculum. In addition to ECM roots, inoculum sources are extramatrical mycelium and fungal spores (Hashimoto and Hyakumachi 1998). Fungal spores could percolate into the mineral horizon by rainwater or be brought into the mineral horizon by the activity of soil animals, e.g. earthworms. Pore space in investigated plots was high, 60–70% gravimetrically.
Considering mean relative abundance, the prevailing exploration type of silver fir ECM community in our study was the contact exploration type, and no difference was detected between the two soil horizons studied. We could not confirm our hypothesis that silver fir relies mainly on ECM taxa with emanating structures since the mean relative abundances of contact exploration type and exploration types with emanating structures were not significantly different.
However, our limited sampling effort prevents us from drawing definitive conclusions about the specificities of silver fir exploration types since not only root diameter (Chen et al. 2018) but also soil structure (Baier et al. 2006), soil pH (Suz et al. 2014), quantity and availability of N sources (Suz et al. 2014), enzymatic capabilities of ECM taxa (Baier et al. 2006) and season (Unuk 2020) may play a role. Inclusion of different sites with various nutrient sources and soil structure profiles would be beneficial in explaining the role of exploration types to support growth of trees with thick fine roots.
Conclusions
The high number of ECM taxa found in our study and not previously recorded for silver fir indicates great potential for biodiversity of ectomycorrhizae in this tree species. Further studies should be conducted in hotspots of genetic diversity for silver fir to collect additional information on ECM biodiversity. This study could not detect significantly different species richness, species diversity or ECM community composition in the organic and mineral soil horizons. Overall, community composition was affected by CaCO3, organic carbon, organic carbon stock, total nitrogen stock, C/N ratio and soil bulk density, but no effect of soil parameters was confirmed for exploration type structure. Taxa that occurred with higher mean relative abundance and belonged to the dominant components of the community were Tomentella stuposa and Cenococcum geophilum in the organic horizon and Piloderma sp. 1 in the mineral horizon. These differences suggest some functional differentiation of ectomycorrhizal taxa related to soil horizon in silver fir nutrition. However, we could not confirm the greater importance of exploration types with emanating structures for silver fir, nor any significant differences in the occurrence of exploration types between the two soil horizons.
References
Agerer R (1991) Characterization of ectomycorrhiza. In: Norris JR, Read DJ, Varma AK (eds) Techniques for the study of mycorrhiza. Methods in microbiology, vol 23. Academic, London, pp 25–73
Agerer R (2001) Exploration types of ectomycorrhizae. Mycorrhiza 11:107–114
Agerer R (2006) Fungal relationships and structural identity of their ectomycorrhizae. Mycol Prog 5:67–107
Agerer R (1987-2012) Colour Atlas of Ectomycorrhizae, 1st – 15th edn. Einhorn-Verlag, Schwäbisch Gmünd
Agerer R, Göttlein A (2003) Correlations between projection area of ectomycorrhizae and H2O extractable nutrients in organic soil layers. Mycol Prog 2:45–51
Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res 25:3389–3402
Arocena J, Glowa K, Massicotte H, Lavkulich L, Lavkulich A (1999) Chemical and mineral composition of ectomycorrhizosphere soils of subalpine fir (Abies lasiocarpa (Hook.) Nutt.) in the Ae horizon of a Luvisol. Can J Soil Sci 79:25–35
Arocena JM, Glowa KR, Massicotte HB (2001) Calcium-rich hypha encrustations on Piloderma. Mycorrhiza 10:209–215
Bååth E, Söderström B (1982) Seasonal and spatial variation in fungal biomass in a forest soil. Soil Biol Biochem 14:353–358
Baier R, Ingenhaag J, Blaschke H, Gottlein A, Agerer R (2006) Vertical distribution of an ectomycorrhizal community in upper soil horizons of a young Norway spruce (Picea abies [L.] Karst.) stand of the Bavarian Limestone Alps. Mycorrhiza 16:197–206
Bakker MR, Garbaye J, Nys C (2000) Effect of liming on the ectomycorrhizal status of oak. Forest Ecol Manag 126:121–131
Baldrian P, Merhautová V, Petránková M, Cajthaml T, Šnajdr J (2010) Distribution of microbial biomass and activity of extracellular enzymes in a hardwood forest soil reflect soil moisture content. Appl Soil Ecol 46:177–182
Bauhus J, Messier C (1999) Soil exploitation strategies of fine roots in different tree species of the southern boreal forest of eastern Canada. Can J For Res 29:260–273
Bauman J, Keiffer C, Hiremath S, McCarthy B, Kardol P (2013) Soil preparation methods promoting ectomycorrhizal colonization and American chestnut Castanea dentata establishment in coal mine restoration. J Appl Ecol 50:721–729
Binder A, Persoh D, Yorou N, Verma R, Bässler C, Agerer R (2013) Ectomycorrhizae of Tomentella badia: description and molecular identification. Acta Mycol 48:153–169
Brunner I, Godbold DL (2007) Tree roots in a changing world. J For Res 12:78–82
Chao A, Ma KH, Hsieh TC, Chiu CH (2015) Online program SpadeR (species-richness prediction and diversity estimation in R). Program and User’s Guide published at http://chao.stat.nthu.edu.tw/wordpress/software_download/
Chen W, Koide RT, Adams TS, DeForest JL, Cheng L, Eissenstat DM (2016) Root morphology and mycorrhizal symbioses together shape nutrient foraging strategies of temperate trees. Proc Natl Acad Sci U S A 113:8741–8746
Chen W, Eissenstat DM, Koide RT (2018) Root diameter predicts the extramatrical hyphal exploration distance of the ectomycorrhizal fungal community. Ecosphere 9:e02202
Ćirić M (1966) Zemljište planinskog područja Igman-Bjelašnica. Radovi Šumarskog fakulteta i instituta za šumarstvo u Sarajevu 10:3–44
Comandini O, Pacioni G, Rinaldi A (1998) Fungi in ectomycorrhizal associations of silver fir (Abies alba Miller) in Central Italy. Mycorrhiza 7:323–328
Comandini O, Pacioni G, Rinaldi A (2001) An assessment of below-ground ectomycorrhizal diversity of Abies alba Miller in Central Italy. Plant Biosyst 135:337–350
Comandini O, Haug I, Rinaldi A, Kuyper T (2004) Uniting Tricholoma sulphureum and T. bufonium. Mycol Res 108:1162–1171
Cremer E (2009) Population genetics of silver fir (Abies alba Mill.) in the Northern Black Forest – preconditions for the recolonization of windthrow areas and associated ectomycorrhizal communities. Dissertation zur Erlangung des Doktorgrades der Naturwissenschaften (Dr. rer. nat.). Philipps-Universität Marburg
Dahlberg A (2001) Community ecology of ectomycorrhizal fungi: an advancing interdisciplinary field. New Phytol 150:555–562
DEEMY – an information system for characterization and determination of ectomycorrhizae. (2004–2017) www.deemy.de
Dickie IA, Xu B, Koide RT (2002) Vertical niche differentiation of ectomycorrhizal hyphae in soil as shown by T-RFLP analysis. New Phytol 156:527–535
Duchaufour P (1982) The dynamics of organic matter. In: Pedology: pedogenesis and classification. Springer Netherlands, Dordrecht, pp 28–66
Duddridge JA, Malibari A, Read DJ (1980) Structure and function of mycorrhizal rhizomorphs with special reference to their role in water transport. Nature 287:834–836
Finlay RD (2008) Ecological aspects of mycorrhizal symbiosis: with special emphasis on the functional diversity of interactions involving the extraradical mycelium. J Exp Bot 59:1115–1126
Fitter A (2002) Characteristics and functions of root systems. In: Waisel Y, Eshel A, Kafkafi U (eds) Plant roots: the hidden half. Third edition, revised and expanded edn. Marcel Dekker, New York
Gardes M, Bruns TD (1993) ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol Ecol 2:113–118
Goodman DM, Trofymow JA (1998) Distribution of ectomycorrhizas in micro-habitats in mature and old-growth stands of Douglas-fir on southeastern Vancouver Island. Soil Biol Biochem 30:2127–2138
Griffiths H, Kryštufek B, Reed J (2004) Balkan biodiversity: pattern and process in the European hotspot. Springer
Hadley W (2011) The split-spply-combine strategy for data analysis. J Stat Softw 40
Hashimoto Y, Hyakumachi M (1998) Distribution of ectomycorrhizas and ectomycorrhizal fungal inoculum with soil depth in a birch forest. J For Res 3:243–245
Horton TR, Molina R, Hood K (2005) Douglas-fir ectomycorrhizae in 40- and 400-year-old stands: mycobiont availability to late successional western hemlock. Mycorrhiza 15:393–403
IUSS Working Group WRB (2015) World Reference Base for Soil Resources 2014, update 2015. International soil classification system for naming soils and creating legends for soil maps. World Soil Resources Reports No. 106. Rome
Jakucs E, Eros-Honti Z (2008) Morphological-anatomical characterization and identification of Tomentella ectomycorrhizas. Mycorrhiza 18:277–285
Jakucs E, Kovács GM, Agerer R, Romsics C, Erős-Honti Z (2005) Morphological-anatomical characterization and molecular identification of Tomentella stuposa ectomycorrhizae and related anatomotypes. Mycorrhiza 15:247–258
Jakucs E, Eros-Honti Z, Seress D, Kovacs GM (2015) Enhancing our understanding of anatomical diversity in Tomentella ectomycorrhizas: characterization of six new morphotypes. Mycorrhiza 25:419–429
Kowalski S (2008) Mycorrhizae of the European silver fir (Abies alba Mill.) seedlings from natural and artificial regeneration in forests of the Karkonosze National Park. In: Silver fir in the Karkonosze National Park. KPN Jelenia Góra
Kuyper T, Landeweert R (2002) Vertical niche differentiation by hyphae of ectomycorrhizal fungi in soil. New Phytol 156:323–325
Laganà A, Angiolini C, Loppi S, Salerni E, Perini C, Barluzzi C, De Dominicis V (2002) Periodicity, fluctuations and successions of macrofungi in fir forests (Abies alba Miller) in Tuscany, Italy. Forest Ecol Manag 169:187–202
Liepelt S, Cheddadi R, de Beaulieu JL, Fady B, Gömöry D, Hussendörfer E, Konnert M, Litt T, Longauer R, Terhürne-Berson R, Ziegenhagen B (2009) Postglacial range expansion and its genetic imprints in Abies alba (Mill.) — a synthesis from palaeobotanic and genetic data. Rev Palaeobot Palyno 153:139–149
Lilleskov EA, Hobbie EA, Horton TR (2011) Conservation of ectomycorrhizal fungi: exploring the linkages between functional and taxonomic responses to anthropogenic N deposition. Fungal Ecol 4:174–183
Marco A, Fioretto A, Giordano M, Innangi M, Menta C, Papa S, Virzo De Santo A (2016) C stocks in forest floor and mineral soil of two Mediterranean beech forests. Forests 7:181
Marinček L, Mucina L, Zupančič M, Poldini L, Dakskobler I, Accetto M (1993) Nomenklatorische revision der illyrischen Buchenwälder (Verband Aremonio-Fagion). Stud Geobotanica 12:121–135
Monfort-Salvador I, García-Montero L, Grande-Ortíz M (2015) Impact of calcium associated to calcareous amendments on ectomycorrhizae in forests: a review. J Soil Sci Plant Nutr 15:217–231
Mrak T, Gričar J (2016) Atlas of woody plant roots. Morphology and anatomy with special emphasis on fine roots. Studia Forestalia Slovenica, vol 147, 1st edn. The Silva Slovenica Publishing Centre, Ljubljana
Oksanen J, Blanchet FG, Kindt R et al (2012) Vegan: community ecology package. R package version 2.0–5
Porras-Alfaro A, Liu KL, Kuske CR, Xie G (2014) From genus to phylum: large-subunit and internal transcribed spacer rRNA operon regions show similar classification accuracies influenced by database composition. Appl Environ Microbiol 80:829–840
R Core Team (2018) R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna
Raja HA, Miller AN, Pearce CJ, Oberlies NH (2017) Fungal identification using molecular tools: a primer for the natural products research community. J Nat Prod 80:756–770
Rineau F, Courty P-E (2011) Secreted enzymatic activities of ectomycorrhizal fungi as a case study of functional diversity and functional redundancy. Annal For Sci 68:69–80
Rosinger C, Sandén H, Matthews B, Mayer M, Godbold DL (2018) Patterns in ectomycorrhizal diversity, community composition, and exploration types in European beech, pine, and spruce forests. Forests 9:445
Rosling A, Landeweert R, Lindahl BD, Larsson K-H, Kuyper TW, Taylor AFS, Finlay RD (2003) Vertical distribution of ectomycorrhizal fungal taxa in a podzol soil profile. New Phytol 159:775–783
Rudawska M, Pietras M, Smutek I, Strzeliński P, Leski T (2016) Ectomycorrhizal fungal assemblages of Abies alba Mill. outside its native range in Poland. Mycorrhiza 26:57–65
Scattolin L, Montecchio L, Mosca E, Agerer R (2008) Vertical distribution of the ectomycorrhizal community in the top soil of Norway spruce stands. Eur J Forest Res 127:347–357
Schirkonyer U (2013) Ectomycorrhizal diversity at five different tree species in forests of the Taunus Mountains in Central Germany. Open J Ecol 3:66–81
Skinner MF, Bowen GD (1974) The penetration of soil by mycelial strands of ectomycorrhizal fungi. Soil Biol Biochem 6:57–61
Sulzbacher MA, Grebenc T, Cabral TS, Giachini AJ, Goto BT, Smith ME, Baseia IG (2016) Restingomyces, a new sequestrate genus from the Brazilian Atlantic rainforest that is phylogenetically related to early-diverging taxa in Trappeaceae (Phallales). Mycologia 108:954–966
Suz LM, Barsoum N, Benham S, Dietrich HP, Fetzer KD, Fischer R, García P, Gehrman J, Kristöfel F, Manninger M, Neagu S, Nicolas M, Oldenburger J, Raspe S, Sánchez G, Schröck HW, Schubert A, Verheyen K, Verstraeten A, Bidartondo MI (2014) Environmental drivers of ectomycorrhizal communities in Europe's temperate oak forests. Mol Ecol 23:5628–5644
Tedersoo L, Kõljalg U, Hallenberg N, Larsson K-H (2003) Fine scale distribution of ectomycorrhizal fungi and roots across substrate layers including coarse woody debris in a mixed forest. New Phytol 159:153–165
Topalović M, Travar J (1978) Tumač pedološke karte i tipološke karte gospodarskih jedinica “Igman”, “Zujevina” i “Mehina Luka”. Institut za šumarstvo i drvnu industriju. Zavod za šumarstvo i lovstvo, Beograd
Unuk T (2020) Ektomikorizni simbionti bele jelke (Abies alba Mill.): doktorska disertacija = Ectomycorrhizal symbionts of silver fir (Abies alba Mill.): doctoral dissertation. Ljubljana: [T. Unuk Nahberger], 169 pp., https://repozitorij.uni-lj.si/IzpisGradiva.php?id=114887
Unuk T, Grebenc T (2017) Silver fir (Abies alba Mill.) ectomycorrhiza across its areal. Folia Biol Geol 58:115–123
van der Linde S, Suz LM, Orme CDL, Cox F, Andreae H, Asi E, Atkinson B, Benham S, Carroll C, Cools N, de Vos B, Dietrich HP, Eichhorn J, Gehrmann J, Grebenc T, Gweon HS, Hansen K, Jacob F, Kristöfel F, Lech P, Manninger M, Martin J, Meesenburg H, Merilä P, Nicolas M, Pavlenda P, Rautio P, Schaub M, Schröck HW, Seidling W, Šrámek V, Thimonier A, Thomsen IM, Titeux H, Vanguelova E, Verstraeten A, Vesterdal L, Waldner P, Wijk S, Zhang Y, Žlindra D, Bidartondo MI (2018) Environment and host as large-scale controls of ectomycorrhizal fungi. Nature 558:243–248
Wang Y, Naumann U, Wright ST, Warton DI (2012) mvabund– an R package for model-based analysis of multivariate abundance data. Methods Ecol Evol 3:471–474
Ważny R (2014) Ectomycorrhizal communities associated with silver fir seedlings (Abies alba Mill.) differ largely in mature silver fir stands and in Scots pine forecrops. Ann For Sci 71:801–810
Ważny R, Kowalski S (2017) Ectomycorrhizal fungal communities of silver-fir seedlings regenerating in fir stands and larch forecrops. Trees 31:929–939
Weigt RB, Raidl S, Verma R, Agerer R (2012) Exploration type-specific standard values of extramatrical mycelium – a step towards quantifying ectomycorrhizal space occupation and biomass in natural soil. Mycol Prog 11:287–297
White T, Bruns T, Lee S, Taylor J, Innis M, Gelfand D, Sninsky J (1990) Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: PCR - protocols and applications - a laboratory manual. Academic Press
Zanella A, Jabiol B, Ponge JF, Sartori G, de Waal R, van Delft B, Graefe U, Cools N, Katzensteiner K, Hager H, Englisch M (2011) A European morpho-functional classification of humus forms. Geoderma 164:138–145
Acknowledgements
COST Action FP1305 Biolink, its chair Martin Lukač and STSM Managers Dr. Miglena Zhiyanski and Dr. Federica Ortelli are acknowledged for their support of the short-term scientific mission (STSM) titled ‘Linking mycorrhiza community structure to soil microclimate and organic matter in beech-fir forests: Case study in Bosnia’. We would like to thank Melita Hrenko from the Department of Forest Physiology and Genetics of the Slovenian Forestry Institute for cleaning of mycorrhizal roots and Emina Šunje from the Faculty of Natural Sciences and Mathematics, University of Sarajevo, for helping with statistical analyses.
Funding
The study was funded by the Slovenian Research Agency—Research Programme P4-0107 and Young Researcher Programme (T.U.N.). The visit by E.H. to the Slovenian Forestry Institute was supported by STSM of COST Action FP1305 Biolink (COST-STSM-FP1305-35140). E.H. was partly supported by the Ministry of Civil Affairs of Bosnia and Herzegovina within the Programme for preparation of projects and potential candidates for EU-FP7 funds for 2015 (Official Gazette of BiH No. 90/15).
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
ESM 1
(PDF 2319 kb)
Rights and permissions
About this article

Cite this article
Mrak, T., Hukić, E., Štraus, I. et al. Ectomycorrhizal community composition of organic and mineral soil horizons in silver fir (Abies alba Mill.) stands. Mycorrhiza 30, 541–553 (2020). https://doi.org/10.1007/s00572-020-00970-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00572-020-00970-y