
Abstract
The principal focus of this study was to describe how the coat characteristics could affect the heat exchanges in animals managed in a hot environment. The Morada Nova ewes were monitored once a month, during 10 consecutive months, in three commercial flocks. Initially, an analysis was performed to measure the differences regarding the coat color in the thermoregulation mechanisms. The animals were grouped into 4 different groups according to coat tonality, as follows: dark red animals (group 1, N = 23), intermediate red color (group 2, N = 27), light red animals (group 3, N = 30), and white-coated animals (group 4, N = 30). The data were collected from 1100 to 1400 h, after the animals were exposed to 30 min of direct sunlight. The cluster analysis was performed considering the hair structural characteristics such as coat thickness (CT, cm), hair length (HL, mm), hair diameter (HD, m), and number of hairs (NH, hairs per unit area), after that these clusters were compared in relation to thermoregulatory mechanisms that include rectal temperature (RT, °C), respiratory rate (RR, breaths min−1), cutaneous evaporation (CE, °C), and respiratory evaporation (RE, W m−2). The groups were characterized and compared using mean and standard deviation, and the differences between the clusters were compared using the Tukey test with a 5% probability of error. In relation to coat color, no differences were found in groups 1, 2, and 3 regarding the activation of the thermoregulation mechanisms. The most different was observed in the totally white coat that presented different thermoregulatory responses as the highest sweating rate. White-coated animals showed a non-pigmented epidermis, and the hair structure is responsible to promote skin protection as necessary, such as a dense coat (1242.7 hair cm−2), long hair (14.2 mm), and thicker coat (7.38 mm). In red-coated animals, the hair structure favored heat loss to the environment, such as short hairs, less thick coat, and less hairs per square centimeter. All evaluated animals showed the rectal temperature within the reference limits for the ovine species, regardless of the coat color. In the analysis of clusters related to the physical structure of hair, it was possible to observe that the animals with thick hair, short hair, and less dense coat tended to have a higher capacity to eliminate heat through their respiratory rate and showed less intense heat loss by cutaneous evaporation. We verified that coat color presents a direct influence on the hair structure and the activation of mechanisms related to thermoregulation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The external traits of domestic animals’ coats, such as coat color and physical characteristics of the hair, are determinants of their adaptive capacity (Silva and Maia 2013). Changes occur as a function of the animals’ environment; animals raised in cold weather usually have specific characteristics for such environmental conditions, like a thicker coat, for example, which reduces heat loss from the animal to the environment. On the other hand, animals from regions with high air temperatures and thermal radiation tend to exhibit characteristics that favor thermolysis. Maia et al. (2003) showed differences in coat characteristics in cattle of the same breed raised in the northern and southern hemispheres, as well as differences in coat color, comparing totally white- and black-coated animals, and stated the strong influence of the environment on the formation of coat characteristics; their findings can be verified in other species.
Most domestic sheep species have wool as a body surface coating, and several studies have been conducted to evaluate its importance in the processes related to thermal insulation (McFardare 1968; Piccione et al. 2002; Suhair and Abdalla 2013). Holfman and Riegle (1977) verified differences in shorn and unshorn sheep, reporting a higher rectal temperature and respiratory evaporation, as well as a decline in the arterial P(CO2), in animals evaluated under different air temperatures, such as 20, 30, and 40 °C. Given these studies, it is possible to suppose a high adaptive capacity to hot environments by the external coat in hair sheep. In Brazil, domestic animals were brought by the colonizers, and for many years, the use of wool animals was predominant until new racial types were formed by natural selection; some animals changed to grow a surface covered by hair, as in the case of the Morada Nova breed, which is locally adapted to the semiarid region of Brazil and presents a skin surface covered by hair, as it occurs in cattle. Information on the adaptive advantage of this type of coat in sheep is not clear. McManus et al. (2011) evaluated differences in the physiological responses of sheep with wool and hair coats and found that wool animals had characteristics that were less favorable to their production under the natural conditions of tropical environments, reflecting directly on their rectal temperature, respiratory rate, and hematological characteristics.
In an equatorial region, characterized by high air temperatures and low humidity, the most efficient mechanism of heat loss is evaporation, either cutaneous or respiratory, since it does not depend on a temperature gradient between the surface of the animal and the environment (Silva and Maia 2013). In these conditions, sensitive thermolysis mechanisms may be less expressive or even an additional source of thermal energy gain from the environment (Maia et al. 2005). Thus, animals that have high adaptability to a hot environment should have low resistance to heat dissipation through evaporation, which can be influenced directly by the ambient conditions, such as temperature, humidity, and radiation, as well as the coat characteristics, like a pigmented epidermis; coat color; and coat morphological characteristics, such as hair length, thickness and density (Bianchini et al. 2006; McManus et al. 2011), and the functionality of the sweat gland activity (Costa et al. 2014).
Coat characteristics can be considered important phenotypic markers, capable of being introduced as criterion for the selection of animals; however, it is necessary to know the relation of these characteristics with the thermoregulatory capacity and homeostasis of the animals (Acharya et al. 1995; Bertipaglia et al. 2008; Façanha et al. 2013). In the work conducted by Bertipaglia et al. (2008), the authors found an important relationship between coat characteristics and the interval between calving in beef cattle, such that the selection for animals with shorter hair was correlated with a shorter calving interval. Leite et al. (2018) evaluated coat traits in the Morada Nova sheep in order to select animals with more favorable coat characteristics for heat dissipation, and found that animals who presented greater difficulty in heat elimination, like longer hair and thicker coat, also showed a reduction in T4 concentration. In this way, it is possible to verify the importance of the hair structure for the adaptive capacity of the animals. Thus, the present work was elaborated to verify the relation between the coat characteristics, such as coat color and hair morphological structure, and their influences in the thermoregulatory mechanisms, such as cutaneous and respiratory evaporation, in hair sheep under a hot environment.
Materials and methods
The experiment was conducted in three commercial flocks in an equatorial region (5° 09′ 02″ S), located in Ceará State, Brazil, over 10 consecutive months, from March to December. The climate type is BWSh according to the Koppen classification (Alvares et al. 2014). The animals were raised in an extensive system with the exclusive feeding of Caatinga vegetation.
Environmental characterization
We recorded information about the thermal environment which the animals were exposed. The thermal environment was monitored once a month in each herd. Environmental data were always recorded together with the physiological data and hair sample. The data were recorded from 1100 to 1400 h. The animals remained under direct sunlight for 30 min before being evaluated. The following parameters were registered: air temperature (AirT, °C), wet-bulb temperature (Tw, °C) obtained with a ventilated psychrometer, black globe temperature (TG, °C) was taken by a 15-cm-diameter copper black globe positioned in the same location as the animals, wind speed (w, m s−1) registered by a digital anemometer (LUTRON AM-4201), and radiant heat load (RHL, W m−2) measured according to Silva (2008). Average values of those measurements are shown in Table 1.
Animal characterization
Animal care procedures throughout the study followed protocols approved by the University Ethics Committee, CEUA-UFERSA, number 23091003895/2014-71. Forty apparently healthy Morada Nova ewes were evaluated in each flock during 10 consecutive months. The measurements were taken always between 1100 and 1400 h after animals were exposed to 30 min of direct sunlight. The animals were divided into four groups according to coat color, as follows: group 1 (dark red animals, N = 23), group 2 (animals with intermediate red color; N = 27), group 3 (light red animals, N = 30), and group 4 (white-coated animals, N = 30) (Fig. 1). The animals were monitored once a month, during ten consecutive month, in three commercial flocks with a total of 971 observations.

Coat color tonality in Morada Nova breed. Group 1 is formed by animals with darker coat; group 2 is formed by animals with red intermediate tonality; group 3 consists of light red animals; and group 4 is composed of totally white animals
Physiological measurement
The parameters evaluated on the ewes were rectal temperature (RT, °C) using a digital clinical thermometer inserted into the rectum of each animal, respiratory rate (RR, breaths/min) recorded by counting flank movements on 60 s, skin temperature (ST, °C) obtained by an infrared thermometer (MasTec Dt 8550) on the flank region body, and cutaneous evaporation (CE; W m−2) was measured with a ventilated capsule coupled to a tube containing moisture absorbent according to Leite et al. (2018). The data were recorded from 1100 to 1400 h.
As for the heat loss by respiratory evaporation (RE; W m−2), the method described by Silva et al. (2002) was used:
where A (m2) is the body surface area of animal, which is estimated as A = 0.6265e0,013P; P is body weight of the animal (kg); λ is the latent heat of vaporization of water (J g−1); ρ is the air density (g m−3); \( \dot{\mathsf{m}} \) is the mass flow rate (g s−1), estimated as \( \dot{\mathsf{m}}={V}_{\mathrm{RC}}\rho \left(\frac{RR}{60}\right) \) (g s−1), where VRC is the tidal volume, obtained according to Silva et al. (2002); and RR is respiratory rate (breaths min−1). The absolute humidity of the atmosphere (ΨA, g m−3) and of the expired air (훹EXP, g m−3), are given, respectively by:
where PP{TA} is partial vapor pressure (kPa) of the ambient air and PS{TEXP}is the saturation pressure of expired air; TA is air temperature (°K) and TEXP is expired air temperature (°K). The TEXP was determined according to Silva et al. (2002) specifically for sheep.
The rate of convective heat flow from the coat surface to the surrounding air is given by the equation:
where TA and TS are the air and coat surface temperature, respectively, while rH (s m−1) is the boundary layer resistance to convective heat transfer.
where d (m) is the characteristic dimension of the body that was estimated as the square root of the body surface area, determined from the body weight (kg) as AS = 0.171W0.503 after Bennett (1973); ρ (g m−3) is the density; cρ (J g−1 K−1) is the specific heat; and κ (W m−1 K−1) the thermal conductivity of the air (Silva and Maia 2013). The Nusselt number (NU) was estimated as for a horizontal cylinder by Churchill and Bernstein (1977) equation; Reynolds, Prandtl, and Grashof numbers were obtained as given by Silva and Maia (2013).
The heat exchange by radiation was estimated by follow equation:
where ST and MRT are body surface temperature (°K) and mean radiant temperature (°K).
where ε is emissivity thermal radiation of animal surface, ε = 0.98. The σ is the Stefan-Boltzmann constant, σ = 567,051 × 10−8 W m−2 k−1. The Tm = 0.5 (ST + MRT).
Coat characteristics
Coat thickness was determined in situ in the middle of the thorax of each animal, about 20 cm below the dorsal line, with a thin metal rule. Hair samples were taken from the same region where coat thickness (CT, cm) was measured; hair was removed using pliers (Silva 2008). The sample hair was removed and stored in plastic envelopes and measurements of hair length (HL; mm), hair diameter (HD, μm), and hair density (NH, hair/cm2) were recorded in the laboratory. Hair density was obtained by counting the number of hairs removed in 0.1399 cm2 using pliers, according to Lee et al. (1974), and then converted into 1 cm2 (Maia et al. 2003). Hair length was taken as the average length of the 10 longest hairs in the sample, according to Udo’s method (1978). The number of hairs per unit area was obtained by directly counting all hairs in the sample. A digital micrometer (Mitutoyo model) with a scale of 0 to 25 μm was used to measure the hair diameter (μm); were measured in the same hair sample as those used for the measurement of hair length.
Statistical analysis
Statistical analysis combined the multivariate approaches to describe whole response of animals (principal component analysis, discriminant analysis and cluster analysis), and general linear model was used to contrast the hypothesis of the effect of tonality on morphophysiological and thermoregulation response of animals.
Principal component analysis (PCA) was run to evaluate the relative influence of morphological and physiological variables as explanatory factors of animal behavior. PCA allows us to characterize and classify whole response of animals and the relationship with different color coats through new and synthetic variables (principal components as synthesis of original variables). Influence of each variable is evaluated through component matrix of the two first principal components that show correlation between each original variable and each factor as measure of relative importance of each variable in response characterization. KMO and Bartlett Sphericity tests were used to prove analysis and sampling adequacy.
In order to identify variables that determines the differences among animals grouped by coat colors (dark, intermediate, light, and totally white) and how strong these differences are, linear discriminant analysis (LDA) and hierarchical cluster analysis (HCA) were performed with within-animal average of independent variables (average of repeated measurements of each animal over time). The LDA model was stablished through variable inclusion procedure based on Wilks’ lambda test in order to simplify the model. Box’s M test was applied to check the equality of variance-covariance matrices. The relative importance of each independent variables was evaluated through standardized coefficients of canonical discriminant functions and the discriminant capacity of these functions, through canonical correlations and explained variance. The HCA provided the most appropriate number of clusters according to Ward’s method. The following clusters were formed: hair diameter (HD, μm), cluster 1 ≤ 4.3, cluster 2 > 4.3 ≤ 5.1, cluster 3 > 5.1 μm; hair length (HL, mm), cluster 1 ≤ 11.5, cluster 2 > 11.5 ≤ 13.5, cluster 3 ≥ 13.5; hair density (NH, number of hair per cm2), cluster 1 ≤ 950, cluster 2 > 950 ≤ 1300, cluster 3 > 1300. Each cluster of hair morphological traits was compared using the mean test (Tukey test) in relation to cutaneous evaporation and respiratory rate.
Repeated measures general linear model was run. Months (March to December) and coat color group (dark, intermediate, light, and fully white) were included as fixed factors and morphological and physiological traits as dependent variables. Box-Cox transformation was applied to ensure normality and homoscedasticity of residuals.
All statistical analyses were performed with the Statistical Package for the Social Sciences (SPSS), version 20, 2010.
Results
The thermal environment in which the animals were managed was characterized by high air temperature and levels of thermal radiation (Table 1). The lowest air temperature recorded was 29.0 °C and maximum of 37.8 °C. The radiant heat load (RHL) was always high above the 500.0 W m−2. Regardless of the month of data collection, in the year, the animals were exposed to environmental conditions that could represent thermal stress.
Initially, analysis was performed in order to determine the differences in relation in activation of the thermoregulatory mechanisms between the animals with different coat color in the Morada Nova breed. Discriminant analysis showed the differences between white and red coat color (Fig. 2). High percentage of correctly classified animals, mainly due to totally white animals (Table 2) and high canonical correlation of first function (ρ1 = 0.945; 97.9% of variance) point out the strong discriminant capacity of independent variables. White coat animals showed negative value in canonical function 1; therefore, they tend to have lower respiratory rate (standardized coefficient: 0.371), coat surface temperature (standardized coefficient: 0.363), and respiratory evaporation (standardized coefficient: 0.444) but higher hair length (standardized coefficient: − 0.412), hair density (standardized coefficient: − 0.445), and cutaneous evaporation (standardized coefficient: − 0.371). The red coat animals (groups 1, 2m and 3) did not show differences in physical hair traits and in activation of the thermoregulatory mechanisms, regardless of the tonality.

Discriminant analysis plot of the four groups of coat color of Morada Nova breed, evaluated for heat tolerance in semiarid region of Brazil. Independent variables are (cutaneous evaporation (CE, W m−2), respiratory evaporation (RE, W m−2), skin surface temperature (ST, °C), respiratory rate (breaths min−1), hair length (HL, mm), and hair density (NH, hair per unit area). Explained variance of two first canonical functions: 99.7% (first function: 97.9%; second function: 1.8%). Canonical correlation indexes are 0.945 and 0.365 for the first and second functions, respectively, 1 represents dark red coat, 2 is intermediate tonality of red, 3 is light red tonality, and 4 represents totally white sheep
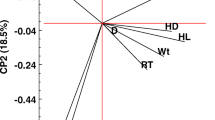
A principal component analysis was performed to assess the interaction between thermoregulation and hair characteristics in each coat color group. In white sheep, the animals tended to present hair characteristics that favor the protection of the epidermis, like longer hair, and a thick and dense coat (Fig. 3). No distinction was observed between the animals in groups 1, 2, and 3. In these groups, the animals had short hair, and a coat with less numerous and more seated characteristics that may favor thermolysis.

Principal component analysis based on thermoregulatory and hair traits in each group of coat color. a Score of each variable included in PCA analysis. b Score of each animals according to coat color group included in PCA analysis. Abbreviations are explained in Table 3
The animals with totally white coats did not have a pigmented epidermis. For this reason, they tended to present hair characteristics that promoted a more intensive protection of the epidermis, as well as a high-density coat (1242.7 hairs cm−2), long hair (14.2 mm), and a thicker coat (7.38 mm). On the other hand, animals with red coats had pigmented epidermis. They presented a greater heating of the coat, leading to a necessity to elevate the respiratory rate (RR) as an immediate heat loss mechanism to maintain homeothermic conditions (Table 3). In this case, it is necessary that these animals present a hair structure that favors heat loss to the environment; this was verified in animals which had a short hair, less thick coat, and less hairs per square centimeter. The important finding was that, regardless of the color and hair structure, there was no difference in the animals’ rectal temperature and all animals were able to maintain homoeothermic conditions, for that, each group triggered different heat loss mechanisms.
In addition to the coat color, the morphological hair structure was also evaluated. The coat traits may be directly related to the animal’s ability to lose heat; animals with thick, short, and less dense hair tend to have a higher capacity to dissipate heat through their respiratory rate and show less intense heat loss by cutaneous evaporation (Fig. 4). The animals that presented the thickest hairs (clusters 2 and 3) tended to have a lower respiratory rate, a coat composed by short hair (< 11.5 mm) and less dense coat (< 950.0 hairs per cm2), corresponding to the animals with lower cutaneous evaporation (CE, W m−2). These characteristics could facilitate the heat dissipation from the animal to the environment.

Mean of cutaneous evaporation (CE) and respiratory rate (RR) in relation to different coat characteristics relation to capacity of heat dissipation, like hair diameter (HD), hair length (HL), and hair density (NH) in hair sheep managed under hot environment
Discussion
The environmental conditions were monitored during 10 consecutive months and indicated that the location where the animals grazed could represent a stressful condition during all months of the year. The minimum value of air temperature (AirT) and radiant heat load (RHL) was 29 °C and 510 W m−2 (Cezar et al. 2004), respectively, which may represent a stressful condition for the animals and create discomfort. When skin temperature is close to the air temperature, the sensible mechanisms are impaired and heat loss depends directly on the capacity of the animals to trigger latent mechanisms, such as an elevation of the respiratory rate and cutaneous evaporation (Table 1). The minimum AirT registered during the experimental period was higher than 25 °C, which is considered the thermal comfort zone for sheep hair, according to Eustáquio Filho et al. (2011). Macías-Cruz et al. (2016) observed that animals exposed to an average AirT of 34 °C had an altered physiological and metabolic status, with changes to the concentration of some blood metabolites and electrolytic adjustments. These changes were also reported by Nardone et al. (2010), who described the negative effects of heat stress on ruminants, warning of the need to preserve breeds that are more adapted to environments with high air temperatures. Thus, under these natural environmental conditions, animals need to present a good adaptive capacity, mainly when they are managed under an extensive system with no access to environmental improvements; thus, animals must have the morphological and physiological structures to adapt to these climatic conditions, such as a hair structure that may favor heat dissipation to maintain homeostasis.
Table 2 classifies the animals according to the canonical discriminant analysis, as a function of their thermoregulatory and hair characteristics. The greatest misclassification was observed between groups 1, 2, and 3, which represent the red variety of the Morada Nova breed. Some animals were not classified correctly in their corresponding group due to homogeneity in their hair characteristics, with the only differences between them being coat color, which does not affect the thermoregulatory mechanisms. The classification error depends on the characteristics used in the analyses. Correa et al. (2013) noted a higher misclassification when considering few characteristics, like anatomical and physiological characteristics; however, in the present study, we analyzed the thermoregulatory mechanisms and hair traits in each group, and verified that red-coated animals did not show differences in relation to their adaptive capacity to the environment in which they are managed.
There was a high difference in white-coated animals, with most differences being with larger animals, denser coat, longer hair, and more heat loss by cutaneous evaporation; these characteristics were different from the red-coated animals, independent of tonality (Fig. 3). The same results were reported by Ferreira et al. (2014) that evaluated genetic diversity and population structure of different varieties of the Morada Nova hair sheep and concluded that the red and white varieties were genetically different and can be considered distinct genetic group. Castanheira et al. (2010) also evaluated the influences of coat color in different genetic groups of sheep and identified that black and brown animals were closest on the tree diagram, probably due to the higher amount of melanin in the skin and coat of these animals, separating them from the white animals. McManus et al. (2009) evaluated the different coat colors of sheep hair managed in a hot environment and concluded that white animals were better adapted to heat stress, with lower heart and breathing rates as well as lower rectal temperatures, concluding that coat color was more important than skin color in terms of heat stress. In red Morada Nova sheep, differences were verified among tonalities, which does not justify discarding animals with darker coats, a practice that has been reducing the genetic variability of the breed (Shiotsuki et al. 2016; Leite et al. 2018).
The assessment of characteristics related to the adaptive capacity of the animals should consider physiological parameters, as measured frequently using the rectal temperature (RT) and RR, as well as add evaluations of the mechanisms of thermoregulation and the coat characteristics that aid with thermal exchange; the joint evaluation of these characteristics favors better clarification of the adjustments to unfavorable environmental conditions and the adaptive capacity of the animals in such situations (Façanha et al. 2013). Thus, we verified that the main differences between the coat color groups were in relation to weight, skin, and hair characteristics and cutaneous evaporation, which were higher in fully white animals. A thicker and denser coat, composed of longer hairs, may have reflected in greater heat dissipation by CE, which was important to maintain rectal temperature within a normal range (Fig. 3). We observed that all tonalities of coat color have the plasticity to adapt to a hot environment, and a change in hair structure could help the formation of this naturally. Correa et al. (2013) also evaluated the importance of adequate coat characteristics in relation to the capacity for heat tolerance in three genetic groups with different hair structures; they reported that the animals that presented the thickest and densest coats with the longest hairs were classified as less adapted because they had more difficulty in releasing heat and failed to maintain homeotermic conditions.
The coat characteristics of white animals promote the protection of the epidermis by blocking the ultraviolet short-wave rays, which is important due to the depigmentation of the epidermis, leading animals to be prone to erythema, burns, and neoplasms. Thus, the physical characteristics of white coats, composed of long hair, and thick and dense coats, acted as a barrier that reduced the absorption of intense direct solar radiation, favoring their production in a hot environment (Table 3). Gebremedhin et al. (1997) suggested that cattle with no pigmented epidermis should have denser coat and long hairs, characteristics that were also noted in white Morada Nova sheep. On the other hand, the physical structure of the hairs of red-coated animals presented differences related to the greater absorption of thermal energy, leading to more heat on the body surface; as such, the characteristics like short-haired and less dense coat are favorable to the dissipation of thermal energy to the environment through convective thermolysis and evaporation on the body surface (Silva 1999).
The relationship between coat characteristics and the activation of the respiratory rate (RR) and cutaneous evaporation (CE) suggests that animals with long hair and denser coat presented greater difficulty in losing heat by convective thermolysis and evaporation on the skin surface; this difficulty on heat elimination can impair the adaptive capacity of animals (Fig. 4). The evaluation of the exterior characteristics of the animals, such as the coat and hair structure, is very important in animals that live in a tropical region; they need high protection against intense solar radiation, as well as a good capacity to eliminate body heat excess through the skin surface, to avoid changes in the physiological characteristics that may have an impact of the homeostasis conditions (Marai et al. 2007). On the other hand, animals that exhibit different hair characteristics than those considered ideal for this region may present greater difficulty on dissipating heat. Thus, the identification of coat characteristics that can help in the selection of animals with better adaptive attributes is of high importance in animals that graze in equatorial regions, which are constantly exposed to high AirT and RHL.
Animals that had a less dense coat showed less cutaneous evaporation (Fig. 4), which may suggest a better adaptive capacity, such that a less dense coat (DN) is ideal for animals raised in environments with high air temperatures and thermal radiation, since it is easier for wind to penetrate inside the coat and remove air trapped between the hairs, which can favor thermal changes.
McManus et al. (2011) evaluated differences in the external coat characteristics of sheep and reported that animals with long hair, and a thick and dark coat showed a higher sweating rate, rectal temperature, and respiratory frequency; however, even the animals that used latent mechanisms of heat loss could not avoid elevations in their internal temperature. Bertipaglia et al. (2008) evaluated the influence of hair characteristics, together with reproductive indices, and verified a high and positive genetic correlation between hair length and the interval between deliveries (0.71) of Braford cattle raised in a tropical environment; this study suggests the inclusion of coat characteristics as a phenotypic marker of adaptability to the selection of better adapted animals, resulting in improved reproductive efficiency, such that animals with short hairs had smaller intervals between deliveries. Thus, it is necessary to consider not only the color but also the structure of the coat in the selection of animals with better adaptive capacities to warm environments.
The influence of coat color and hair structure on the adaptive capacity of animals was verified. There was plasticity in the maintenance of homeothermic conditions, even when animals presented characteristics that were less favorable to being raised in regions with high air temperatures.
Conclusion
In the Morada Nova breed, differences in skin characteristics are due to the pigmentation of the epidermis. In animals with a fully white coat, the protection of the non-pigmented epidermis is necessary, whereas in the red variety, the external coat characteristics promote heat dissipation to the environment.
Hair traits can be used as phenotypic markers in the selection of better adapted animals, mainly the length and number of hairs per square centimeter, which are characteristics associated with thermoregulatory mechanisms.
References
Acharya RM, Gupta RM, Sehgal JPE, Singh M (1995) Coat characteristic of goats in relation to heat tolerance in the hot tropics. Small Rumin Res 18:245–248
Alvares CA, Stape JL, Sentelhas PC, Gonçalves JLM, Sparovek G (2014) Köppen’s climate classification map for Brazil. Meteorol Z 22(6):711–728
Bennett JW (1973) Regional body surface area of sheep. J Agric Sci 81:429–432
Bertipaglia ECA, Silva RG, Cardoso V, Fries LA (2008) Desempenho reprodutivo, características do pelame e taxa de sudação em vacas da raça Braford. Rev Bras Zootec 37:1573–1583
Bianchini E, McManus C, Lucci CM, Fernandes MCB, Prescott E, Mariante AS, Egito AA (2006) Caracteristicas corporais associadas com a adaptação ao calor em bovinos naturalizados brasileiros. Pesq Agropecuaria Bras 1:1443–1448
Castanheira M, Paiva SR, Louvandini H, Landim A, Fiorvanti MCS, Dallago BS, Correa OS, McManus C (2010) Use of heat tolerance traits in discriminating between groups of sheep in central Brazil. Trop Anim Health Prod 42:1821–1828. https://doi.org/10.1007/s11250-010-9643-x
Cezar MF, Souza BB, Souza WH (2004) Evaluation of physiological parameters of sheep from Dorper, Santa Inês and their crosses in climatic conditions of northeast semi-arid. Ciênc Agrotecnol 28:614–620
Churchill SW, Bernstein M (1977) Correlating equation for forced convection from gases and liquids to a circular cylinder in crossflow. J Heat Transf 94:300–305
Correa MPC, Dallago BSL, Paiva SR, Canozzi MEA, Louvandini H, Barcellos JJ, McManus C (2013) Multivariate analysis of heat tolerance characteristics in Santa Inês and crossbreed lamb in the Federal District of Brazil. Trop Anim Health Prod 45:1407–1414
Costa CCM, MaiaASC, Fontenele Neto JD,Oliveira SEO,Queiroz JPAF (2014) Latent heat loss and sweat gland histology of male goats in an equatorial semi-arid environment. Int J Biometeorol 58:179–184
Eustáquio Filho A, Teodoro SM, Chaves MA, Santos PEF, Silva MWR, Murta RM, Carvalho GGP, Souza LEB (2011) Thermal comfort zone of Santa Inês sheep on the basis of physiological responses. Rev Bras Zootec 40:1907–1814
Façanha DAE, Chaves DF, Morais JHG, Vasconcelos AM, Costa WP, Guilhermino MM (2013) Tendências metodológicas para avaliação da adaptabilidade ao ambiente tropical. Rev Bras Saúde Prod Anim 14:91–103
Ferreira JSB, Paiva SR, Silva EC, McManus CM, Caetano AR, Façanha DAE, Sousa MAN (2014) Genetic diversity and population structure of different varieties of Morada Nova hair sheep from Brazil. Genet Mol Res 13:2480–2490
Gebremedhin KG, Ni H, Hillian PE, (1997) Temperature profile and heat flux through irradiated fur layer. In: Proceeding of the international Livestock Environment Symposium, Bloomington, MN. 1:226–233
Holfman WF, Riegle GD (1977) Thermorespiratory responses of shorn and unshorn sheep to mild heat stress. Respir Physiol 30:327–338
Lee JA, Roussel JD, Beatty JF (1974) Effect of temperature season on bovine adrenal cortical function, blood cell profile, and milk production. J Dairy Sci Champaign 59:104–108
Leite JHGM, DaSilva RG, Silva WST, Silva WE, Paiva RDM, Sousa JER, Bermejo LA, Façanha DAE (2018) Locally adapted Brazilian ewes with different coat colors maintain homeothermy during the year in an equatorial semiarid environment. Int J Biometeorol 62:1635–1644
Macías-Cruz U, López-Baca MA, Vicente R, Mejía A, Álvarez FD, Correa-Calderón A, Meza-Herrera CA, Mellado M, Guerra-Liera JE, Avendaño-Reyes L (2016) Effects of seasonal ambient heat stress (spring vs. summer) on physiological and metabolic variables in hair sheep located in an arid region. Int J Biometeorol 60:1279–1286. https://doi.org/10.1007/s00484-015-1123-6
Maia ASC, Silva RG, Bertipaglia EA (2003) Características de pelame de vacas Holandesas em ambiente tropical: um estudo genético e adaptativo. Rev Bras Zootec 38:843–853
Maia ASC, Silva RG, Bertipaglia EA (2005) Environmental and genetic variation of the effective radiative properties of the coat of Holstein cows under tropical conditions. Livest Prod Sci 92:307–315
Marai IFM, El-Darawany AA, Fadiel A, Abdel-Hafez MAM (2007) Physiological traits as affected by heat stress in sheep–a review. Small Rumin Res 71:1–12
McFardare WV (1968) Adaptation of ruminants to Tropics and deserts. In: Hafez ESE (ed) Adaptation of domestic animals. Lea and Febiger, Philadelphia, pp 164–182
McManus C, Louvandini H, Gugel R, Sasaki LCB, Bianchini E, Bernal FEM, Paiva SR, Paim TP (2011) Skin and coat traits in sheep in Brazil and their relation with heat tolerance. Trop Anim Health Prod. Roslin 43(1):121–126
McManus C, Paludo GR, Louvandini H, Gugel R, Sasaki LCB, Paiva SR (2009) Heat tolerance in Brazilian sheep: physiological and blood parameters. Trop Anim Health Prod 41:95–101
Nardone A, Ronchi B, Lacetera N, Ranieri MS, Bernabucci U (2010) Effects of climate changes on animal production and sustainability of livestock systems. Livest Sci 130:57–69
Piccione G, Caola G, Refinetti R (2002) Effect of shearing on the core temperature of three breeds of Mediterranean sheep. Small Ruin Res 46:211–215
Shiotsuki L, Silva PHT, Silva KM, Landim AV, Morais OR, Facó O (2016) The impact of racial pattern on the genetic improvement of Morada Nova sheep. Anim Genet Resour Food Agric Organ U N 1–10
Silva RG (1999) Estimativa do balanço térmico por radiação em vacas Holandesas expostas ao sol e à sombra em ambiente tropical. Rev Bra Zootech 28:1403–1411
Silva RG (2008) Biofísica ambiental: os animais e seu ambiente. Jaboticabal: FUNEP-SP 393 p
Silva RG, Lascala JRN, Lima Filho AE (2002) Respiratory heat loss in the sheep: a comprehensive model. Int J Biometeorol 46:136–140
Silva RG, Maia ASC (2013) Principles of animal biometeorology, 1st edn. Springer, Hedelberg 261p
Suhair MS, Abdalla AM (2013) Effect of season changes and shearing on thermoregulation, blood constituents and semen characteristics of desert ram (Ovis aries). Park J Biolo Sci 16(24):1884–1893
Udo HM (1978) Hair coat characteristics in Friesian Heifers in the Netherlands and Kenya. Meded. Landbouwhogeschool Wagneingen, Wagneingen 135 p
Funding
The authors acknowledge EMBRAPA—Goat and Sheep (process number: 99999010837/2014-00) for the financial Support and CAPES—Ciência sem Fronteiras Program for the partnership with the doctor Luis Alberto Bermejo Asensio (process number: 88881030352/2013-1).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Animal care procedures throughout the study followed protocols approved by the University Ethics Committee, CEUA-UFERSA, number 23091003895/2014-71.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Leite, J.H.G.M., Da Silva, R.G., Asensio, L.A.B. et al. Coat color and morphological hair traits influence on the mechanisms related to the heat tolerance in hair sheep. Int J Biometeorol 64, 2185–2194 (2020). https://doi.org/10.1007/s00484-020-02014-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00484-020-02014-8