
Abstract
Under temperate climates, cattle are often at pasture in summer and are not necessarily provided with shade. We aimed at evaluating in a temperate region (Belgium) to what extent cattle may suffer from heat stress (measured through body temperature, respiration rate and panting score, cortisol or its metabolites in milk, and feces on hot days) and at assessing the potential benefits of shade. During the summer of 2012, 20 cows were kept on pasture without access to shade. During the summer of 2011, ten cows had access to shade (young trees with shade cloth hung between them), whereas ten cows had no access. Climatic conditions were quantified by the Heat Load Index (HLI). In animals without access to shade respiration rates, panting scores, rectal temperatures, and milk cortisol concentrations increased as HLI increased in both 2011 and 2012. Fecal cortisol metabolites varied with HLI in 2011 only. When cattle had access to shade, their use of shade increased as the HLI increased. This effect was more pronounced during the last part of the summer, possibly due to better acquaintance with the shade construction. In this case, shade use increased to 65% at the highest HLI (79). Shade tempered the effects on respiration, rectal temperature, and fecal cortisol metabolites. Milk cortisol was not influenced by HLI for cows using shade for > 10% of the day. Therefore, even in temperate areas, cattle may suffer from heat when they are at pasture in summer and providing shade can reduce such stress.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
In temperate regions, cattle are often kept on pasture during the warmer part of the year. This has several benefits for cattle health and welfare, including fewer claw and leg problems and more opportunities for natural behavior such as exploration and grazing, but also poses certain risks, such as exposure to insects and parasites, to predators, or to adverse weather conditions—cold, wet, or hot weather—(Haskell et al. 2006; Hernandez-Mendo et al. 2007; Phillips et al. 2013; Reijs et al. 2013). In hot climates, heat is known to induce adverse physiologic effects and to reduce productivity (Collier et al. 1982; Kadzere et al. 2002; Silanikove 2000). In such conditions, the provision of shade is generally regarded as one of the most efficient prevention strategies at pasture (Hansen 1990; Muller et al. 1994a; Muller et al. 1994b; Valtorta et al. 1997; Van Laer et al. 2015c).
In temperate regions, the importance of heat stress prevention for cattle is sometimes contested. Indeed shade is not necessarily provided to cows at pasture. However, high productive dairy cows (such as Holstein or Holstein-Friesian cows) are particularly susceptible to heat stress, because their high metabolic rate results in considerable metabolic heat, making it difficult for cows to maintain thermal balance under hot conditions (Kadzere et al. 2002; Van Laer et al. 2014). In addition, even in temperate regions such as Belgium, summer climatic conditions are occasionally above the thermo-neutral zone of dairy cows (Van Laer et al. 2014).
The experienced heat load is a combination of air temperature, relative air humidity, wind speed, and solar radiation (Gaughan et al. 2008). During high heat load, cattle seek cooler locations such as ridge tops, wind-exposed slopes, or shade (Schutz et al. 2014; Senft and Rittenhouse 1985; Van Laer et al. 2015a). They increase their water intake and reduce their general activity and feed intake (Silanikove 2000). A shift in respiration rate dynamics is observed under high heat load, from rapid panting to a slower, deep phase panting (Gaughan et al. 2000; Silanikove 2000). Such responses are controlled by body temperature information received by the hypothalamus from peripheral as well as central thermoreceptors. For instance, rectal temperature, indicating deep body temperature, is highly correlated to heat stress indicators such as the reduction in feed intake and milk yield (Johnson et al. 1963; Umphrey et al. 2001; West et al. 2003). Behavioral and physiological responses to heat load may nevertheless not prevent cattle from experiencing stress. As a matter of fact, heat load is known to activate the HPA axis, resulting in an increase of plasma cortisol concentrations, a response that reflects stress (Christison and Johnson 1972).
According to Mormède et al. (2007), milk cortisol (MC) concentrations are highly correlated to plasma cortisol concentrations sampled at the same time. In feces, not the native glucocorticoids but their metabolites (produced by the liver and excreted into the gut via the bile) are present (Mostl and Palme 2002). Concentrations of fecal cortisol metabolites (FCM) represent the cumulative secretion of glucocorticoids integrated over a few hours (Palme 2012). In addition, there is a time delay between increased plasma glucocorticoid levels and the excretion of FCM, i.e., the gut passage time from the duodenum to the rectum which is about 10 to 12 h in cattle (Palme 2012). FCM can thus provide an integrated measure of stress over several hours, whereas MC rather provides a measure of stress over the past minutes.
The current study had two objectives. The first objective was to study to what extent high producing dairy cows experience heat stress in a temperate climate area (here Belgium). Heat stress was assessed through modification of the respiration, rectal temperature, and cortisol release assessed through MC and FCM to avoid invasive measurements. The second objective was to assess the potential benefit of providing shade to dairy cows on reducing heat stress in such temperate conditions. The impact of heat stress and provision of shade on energy metabolism, milk yield, and milk composition have already been reported (Van Laer et al. 2015c) and will not be addressed here.
Materials and methods
Experimental setup, animals, and management
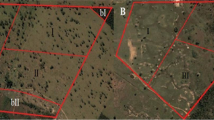
The study took place during the summers of 2011 and 2012. It was approved by the Animal Ethics Committee of the Flemish Institute for Agricultural and Fisheries Research (application nos. 2011/151 and 2011/151bis). The experiment was carried out on the pastures of the institute’s research farm (latitude 50° 59′ 1″ N, longitude 3° 46′ 49″ E) (Fig. 1). The Holstein dairy cows were—rotationally—kept on four pastures in 2011 and on two pastures in 2012. Each pasture was adjacent to a shaded area, especially arranged for the experiment, that was surrounded by an electric fence. The two shaded areas were 625 m2 each and were used by a maximum of 60 dairy cows on the adjacent pastures. They thus offered at least 10.5 m2 of shade per cow. This shaded area could be accessed from either of the two adjacent pastures through a 3 to 5 m wide passage. The shade was provided by young trees—up to 10 m high—and shade cloth (Duranet bvba, Ostend, Belgium) spanned between the trees (Fig. 1). The cloth blocks 80% of the incoming sun light. At least one open drinking trough was available in each pasture the cows were kept in. No drinking troughs were present in the shaded areas.

Experimental setup in 2011. The “shade” treatment and “no shade” treatment were kept on separate allotments, with and without access to a fenced shade area, respectively. To exclude possible effects of allotment on the response variables, the experimental groups and the passage to the shaded area were switched daily between allotments. The shade area consisted of a part of the pasture (25 m × 25 m) with evenly spaced (5 m) young willow (Salix alba), poplar (Populus alba) and alder (Alnus glutinosa) trees, and shade cloth with a shading percentage of 80% spanned up between them
In 2011, the herd consisted of 125 dairy cows, which were of 2.0 ± 1.6 (mean ± SD) parity, 169.1 ± 132.4 days in milk, and yielded 26.9 ± 11.7 L milk/day at the beginning of the experiment (8/6/2011). In 2012, the herd consisted of 66 dairy cows, which were of 2.9 ± 1.1 parity, 181 ± 123 days in milk, and yielded 30.9 ± 7.3 L milk/day at the beginning of the experiment (1/6/12). All cows were milked twice daily (starting at 05:30 and at 15:30) and received half of the daily ration of concentrates during each milking. After each milking, they were kept indoors (maximum 2 h indoors around each milking), and they were fed the daily mixed ration of corn silage (49 to 76%, 60% on average) and prewilted grass silage (9 to 29%, 21% on average), supplemented with a protein source (soybean meal or protected soybean meal) and wheat or corn cob mix. Additionally, during some periods, the ration was completed with pressed beet pulp (0 to 25%, 9% on average) or by-products from bio-ethanol or starch industry. This mixed ration was provided in free-access, custom-built, concrete feed troughs located in a loose housing barn. Water inside the barn was provided through a free-access water trough of 2 m long and a content of 135 L (Suevia), which was automatically filled.
During the summer of 2011, the dairy herd was divided into two groups of similar size which were as comparable as possible with regard to traits known to affect susceptibility to heat stress, namely productivity (Kadzere et al. 2002), parity (Gantner et al. 2012), age (Collier et al. 1982), and percentage of black coat (Becerril et al. 1993; Cena and Monteith 1975; Hansen 1990). One group (the “shade” treatment) was always granted access to the shaded area, whereas the other group (the “no shade” treatment) never had access to shade when on pasture (by opening and closing the appropriate gates to the shaded areas). In order to exclude potential confounding effects of allotment to either of the pastures available (e.g., pasture productivity or composition, location of drinking troughs) on the cows’ behavior or productivity, the “no shade” and “shade” groups were switched daily between allotments (Fig. 1). During the summer of 2012, the cows were kept on the same pastures as those used in 2011, but none of the cows had access to shade (“no shade” treatment).
We will report on data from 20 “focal cows,” i.e., always the same 20 cows, in each summer. The cows were chosen so as to reflect the normal variability within a herd in terms of percentage of black coat, productivity, parity, and age.
In 2011, each of the ten “focal cows” in the “shade” group formed a “matched pair” with a cow of the “no shade” group which was similar in terms of productivity, parity, age, and percentage of black coat. The data were collected on 11 days in 2011 and 8 days in 2012, spread over a range of Heat Load Index (HLI) from 50.8 to 83.2 corresponding in our experiment to a range of Thermal Humidity Index (THI) from 60 to 78.9 (Table 1). In the following sections, we consider HLI rather than THI because it reflects more precisely the heat perceived by animals, taking into account the intensity of solar radiation and the wind speed in addition to air temperature and humidity used in THI (Gaughan et al. 2008; Van Laer et al. 2015a; Van Laer et al. 2015b).
Climatic data
A custom-built Campbell Scientific BWS200 weather station (Campbell Scientific Inc., Logan, Utah, US) located in open pasture within 500 m of all pastures used in the trial, registered the average air temperature, air humidity, solar radiation, and wind speed every 15 min.
We first calculated the black globe temperature (Tbg) according to Hahn et al. (2003) formula:
with Ta, air temperature (in °C) and Rad, intensity of solar radiation (in W/m2).
Then, we calculated HLI according to Gaughan et al. (2008) formulas:
with RH, relative air humidity in % and WS, wind speed in m/s.
These indicators were calculated for every 15 min interval between 09:00 and 16:00; then averaged per day.
In addition, for each of the shaded areas, we directly measured air temperature and black globe temperature with Testo 400’s Wet Bulb Globe Temperature probe (Testo AG Inc., Lenzkirch, Germany), under shade and in open area nearby (20–50 m). These measurement were made on 7 days when the weather forecast had predicted daily maximum temperatures ≥ 25 °C (i.e., when weather conditions outside the thermoneutral zone for cattle could be expected), in order to evaluate the effect of shade on microclimate during hot days.
Use of shade
An unmanned camera (Sony HDR-CX220E) was used to record each individual cow’s passage to and from the shaded area between 10:00 and the evening milking at 15:00. The use of shade by cows could thus be detected. The percentage time spent in shade by individual cows was then estimated. Shade use probability reached 10% at HLI values beyond the threshold of 77, which Gaughan et al. (2008) used to define moderate heat. Therefore, we decided to use a cut-off of 10%, above which an individual cow is considered to use the shade to avoid thermal discomfort outside shade.
Respiration rate and panting score
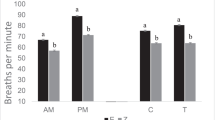
We monitored respiration rate and panting score, by means of hourly scans, between 10:00 and 15:00. The respiration rate was determined by timing five respirations (flank movements) and converting this to the breaths per minute. We estimated a panting score based on respiratory rate, deepness of panting, and the degree of drooling (Gaughan et al. 2008). Like in Tuyttens et al. (2014), panting was scored on a tagged scale, labeled with the descriptors of Gaughan et al. (2008) as shown in Fig. 2. The observers could choose values between tags so that the data were continuous. The respiration rates and panting scores were averaged per cow over the entire observation day.

Rectal temperature and HPA axis activity
At the end of each observation day, the focal cows were separated from the herd prior to milking for measuring rectal temperature and collecting a 20 mL milk sample. After milking, the focal cows were housed in a loose housing barn adjacent to the milking parlor, while the rest of the herd returned to pasture. At morning milking, around 06:00, a second 20 mL milk sample was collected. All milk samples were immediately frozen at − 20 °C until analysis. Following morning milking, a fecal sample of 20 g was taken directly from the rectum of the focal cows. Immediately afterwards, a well-mixed subsample of 2 g was taken from each sample and frozen at − 20 °C until analysis.
Fecal samples were analyzed for 11,17-dioxoandrostanes (a group of FCM) using an 11-oxoetiocholanolone enzyme immunoassay (EIA). Details of the extraction, the EIA, and its validation for use in cattle have been previously described in detail (Palme and Mostl 1997; Palme et al. 1999; Palme et al. 2013). Milk samples were thawed and defatted by centrifugation. Glucocorticoids were extracted with ethyl acetate, and MC concentrations were determined by radioimmunoassay as described in Veissier et al. (2008).
Data analysis
All data were analyzed with SAS 9.3 software. This paper examines the reactions of individual cows when they are exposed to a high heat load. Although the cows were in groups, we considered that the cow was the observation unit because we focused on the physiological reactions to heat and not on behavior. Therefore, we carried out longitudinal analyses, with the animals being their own controls. The reactions of cows were compared between days varying in HLI, and in 2011, we analyzed the effect of the use of shade by a given cow, taking into account the amount of shade use by each cow.
Effect of shade on microclimate
The air temperature and the black globe temperature were compared between measurement locations in open area and under shade, using a mixed model ANOVA. Measurement session and shade area were included as random intercept effects.
Use of shade
We examined the effect of HLI on probability of shade use (per 15 min) by means of a mixed model logistic regression. A random factor was included to correct for repeated measurements per cow. This was modeled both with the entire dataset (including all sampling days) and with a restricted dataset, including the four last sampling days of the summer of 2011 only. This decision was based on preliminary analyses of shade use, which showed that the daily average shade use exceeded 10% on only 1 day in June and July versus 4 days in August and September (Fig. 3). The logistic regression models yield the probability of shade use as outcome variable. This probability can be interpreted as the probability that an individual cow will use shade at a given HLI value, which is basically the same as the proportion of the group that can be expected to use shade at a given HLI value.

Boxplots showing the mean (black rectangle), the 25 and 75% quartile (box), the 5 and 95% percentiles (whiskers) and the outliers (black circle) for the shade use by the ten focal cows having access to shade in 2011. The daily average Heat Load Index (HLI) during the observation days is given below the X-axis
Respiration rate, panting score, rectal temperature, and fecal cortisol metabolites (FCM)
For cows with access to shade (2011 only), the daily percentage of time spent in shade varied strongly between cows within the same observation day, as well as between observation days. Therefore, the daily shade use percentage was divided into three categories: (a) shade used < 10% of the day, (b) shade used between 10 and 50% of the day, and (c) shade used > 50% of the day, to obtain a new variable which we named “shade use category.”
The daily average respiration rates and panting scores, the rectal temperature, and FCM were analyzed separately for 2011 and 2012, by means of mixed linear regressions (proc mixed, SAS 9.4). Again, a random factor was included to correct for repeated measurements per cow. These regressions modeled the rectal temperature and FCM as a function of (1) the daily average HLI, (2) the treatment (“shade” or “no shade” in 2011; not applicable in 2012), and (3) the effect of the shade use category (only applicable for “shade” cows in 2011), the interaction between (1) and (2), and the interaction between (1), (2), and (3).
Milk cortisol (MC)
The MC concentrations were analyzed for the effect of HLI, treatment, and of time of sampling. We aimed to compare the relation between HLI and evening as well as morning MC concentrations between animals that used shade to different degrees. Therefore, we modeled the effect of (1) the daily average HLI, (2) the time of sampling (morning or evening), and (3) the interaction between (1) and (2), separately per shade-use category within each “shade” treatment of 2011 (n = 3), separately for the “no shade” treatment in 2011 (n = 1) and separately for the “no shade” treatment in 2012.
We tried strictly linear models (with HLI included as a linear factor only) as well as polynomial models (with HLI as a quadratic factor too), to detect potential non-linear effects. In each group, we only report results from the best fitting model, i.e., the model which had lowest Corrected Akaike Information Criterium (AICC). This was always the strictly linear model, except for one group, i.e., the cows from the “no shade” treatment in 2011.
We tested whether milk production interacted with HLI to explain cow responses. We did not find any significant interactions (P > 0.05). When only main effects were tested, we found some main effects of milk production on cortisol levels only. However, we decided not to include the effect of milk production in our models because we included the animal as a random factor (nested within year) to correct for repeated measurements per cow. We considered that the effect of milk production was included in the animal effect.
In the next section, the results are shown with means ± standard error.
Results
Effect of shade on microclimate
During hot summer days (i.e., days when weather forecast had predicted daily maximum temperatures ≥ 25 °C), shade lowered the black globe temperature by 4.5 °C and the air temperature by 2.0 °C. Black globe temperature was 30.3 ± 2.0 °C outside shade and 25.8 ± 2.0 °C inside of it (P < 0.001). Air temperature was 25.8 ± 2.1 °C outside shade and 23.8 ± 2.1 °C inside of it (P < 0.001).
Use of shade
The use of shade by the ten focal cows from the “shade” treatment increased exponentially with increasing HLI (P < 0.001, Fig. 4). Over all sampling days, the shade use probability reached 10% at a HLI of 79 and reached 16% at a HLI of 85, which was the maximum HLI that occurred in 2011. During the last four sampling days in 2011, the increase of shade use with increasing HLI (P < 0.001) was more pronounced; the shade use probability reached 10% at a HLI of 68 and 65% at a HLI of 85.

Effect of the 15-min values of the Heat Load Index (HLI) on the probability of shade use, (1) over all sampling days from June to September: black rectangle, Y = − 13.60 (± 1.32) + 0.14 (± 0.02) × X (P < 0.0001) and (2) over the last four sampling days only: dash lines, Y = − 12.88 (± 1.57) + 0.16 (± 0.02) × X (P < 0.0001)
Respiration rate, panting score, and rectal temperature
The respiration rate significantly increased with HLI for cows without access to shade (in 2012 and 2011), whereas a mere trend was observed for cows with access to shade (Table 2). The panting score and the rectal temperature significantly increased with HLI, both for cows without access to shade and for cows with access to shade. Access to shade per se did not affect significantly the panting score (P > 0.10), whereas a slightly higher rectal temperature was noticed in cows with access to shade (P = 0.059). Nevertheless, the access to shade tempered the increases in panting score and rectal temperature with increasing HLI.
For cows with access to shade, the degree to which shade was used (shade use category) had no significant effect on the respiration rate (P = 0.088) or the panting score (P = 0.203). Only rectal temperature was influenced by the degree to which shade was used (P = 0.030). For cows that used shade for more than 10% of the day, the rectal temperature increased less with increasing HLI than for cows that used shade for less than 10% of the day (Table 2).
Fecal cortisol metabolites (FCM)
In 2012, FCM concentrations did not increase significantly with increasing HLI (Table 2). In 2011, FCM concentrations increased significantly with increasing HLI, for cows without as well as for cows with access to shade. Treatment per se had no significant effect, but access to shade tempered the increase of FCM with increasing HLI. The degree to which shade was used (shade use class) had no significant effect on FCM (P = 0.177). When HLI increased from 50 (i.e., the lowest observed daily average HLI value) to 79 (i.e., the highest observed daily average HLI in 2011), FCM concentrations increased from 9.8 to 39.7 ng/g for cows without access to shade and from 17.6 to 35.9 ng/g for cows with access to shade.
Milk cortisol (MC)
For cows without access to shade (all cows in 2012 and cows of the “no shade” treatment in 2011), MC concentrations increased with increasing HLI (Table 3). In 2012, this effect was also influenced by the time of sampling. At daily average HLI values < 70, MC concentrations were higher in the morning than in the evening. But as HLI increased, the evening concentrations increased more than concentrations on the next morning (Fig. 5).

Effects of the daily average Heat Load Index (HLI) and time of sampling (evening or next morning) on concentrations of milk cortisol, for cows without access to shade in 2012 (a) and 2011 (b), and for cows with access to shade that used it less than 10% of the day (SU < 10%) (c). The lines are regression lines
In 2011, the time of sampling affected MC concentrations only for cows that had access to shade but used it less than 10% of the day (Table 3). More specifically, evening concentrations were affected by HLI: when HLI was below 80, MC concentrations were lower in the evening than in the morning (Fig. 5) but when HLI was above 80, then MC concentrations exceeded the morning ones. For cows that used shade for more than 10% of the day, MC levels were unaffected by HLI.
Discussion
The main results of our study are that on days with a high HLI, cows reacted with increases in respiration rate, panting score, rectal temperature, and concentrations of milk cortisol (MC) and fecal cortisol metabolites (FCM) in 2011. When cows had access to shade, these changes were less pronounced.
According to Gaughan et al. (2008), an HLI below 70 indicates thermoneutral conditions for cattle; HLI between 70 and 77 indicate warm conditions, HLI between 77 and 85 indicate hot conditions, and an HLI above 85 indicates very hot conditions. Out of the 19 days that we studied, most of them corresponded to thermoneutrality (10 days), four corresponded to warm days, and four corresponded to hot days. In our study, the cows increased their respiratory rate and panting scores when HLI increased; they did not only pant but also drooled, which suggests severe discomfort. These are important adaptive responses of cattle to eliminate excessive body warmth (Silanikove 2000). These findings confirm earlier results obtained with an average HLI of 81in New Zealand by Schutz et al. (2014; 2010). Nevertheless, the abovementioned adaptive responses were not enough for cows to maintain their deep body temperature, as shown by an increased rectal temperature with increasing HLI when cows did not have access to shade. Indeed, we recorded rectal temperatures up to about 39.5 °C, similar to results from unshaded Holstein cows in South Africa (Muller et al. 1994a) and New Zealand (Kendall et al. 2006; Tucker et al. 2008). Given that dairy cows’ normal body temperature is between 38.5 and 39 °C (West et al. 2003), this is a strong indication that the hottest climatic conditions in our study exceeded the cattle’s thermoneutral zone (Silanikove 2000).
According to Gaughan et al. (2008), the hottest days of our studies (HLI > 77) were prone to induce moderate stress. This was confirmed by the activation of the HPA axis of cows that did not have access to shade or used it for less than 10% of the day. In feces taken on the morning, concentrations of FCM did not increase with increasing HLI on the previous day in 2012, but it did so in 2011. For cows without access to shade in 2011, FCM concentrations increased by 400% when HLI increased from 50 to 79. This finding confirms that FCM can reflect heat stress (Rees et al. 2016). Similarly, the MC levels increased in cows with no access to shade or cows that used shade for less than 10% of the day. The changes in MC was especially marked in the evening. In 2011, as the daily average HLI increased from 65 to 79, the evening MC concentrations increased from 0.45 to 0.82 ng/mL for cows without access to shade. The cortisol concentration in milk reflects closely that of blood (concentrations being about 10 times lower in milk), with no delay in the passage from blood to milk and no accumulation in milk (Mormède et al. 2007). By contrast, the passage through gut induces a delay of about 12 h for cortisol in blood to be reflected in FCM and those accumulate in feces. Taken together, the results on FCM and MC suggest that cows had been stressed for a prolonged period in the afternoon on hot days.
In transport experiments, Palme et al. (2000) reported an increase in FCM (11,17-dioxoandrostanes) of dairy cattle by 550 to 3910% following a 2-h transport over country roads, and Verkerk et al. (1998) reported an increase in MC concentrations from 0.1 to 12.0 ng/mL after a 45-min transport. These values are largely above those that we observed in our cows without shade on hot days. Thus, the degree of stress due to heat in our study seems rather moderate compared to that caused by transport. On the other hand, the stress of cows experiencing high heat load may be less expressed by cortisol release, since this hormone has a strong metabolic effect allowing the production of extra heat by the body (Mormède et al. 2007).
Concentrations of glucocorticoids in blood vary during the day, with generally higher levels in the morning (Thun et al. 1981). This was also the case in our study during thermoneutral conditions. In hot conditions, evening concentrations of cortisol in milk could exceed that of the next morning, bearing support to the fact that cows did suffer from heat, but this effect did not last until the next morning. In other words, the cows could recover from heat stress during the night.
When the cows were provided with shade (half the cows in 2011), they increased their use of shade when weather conditions became hotter. This confirms previous findings obtained in beef cattle in similar climate conditions which showed that White-Blue cattle use natural or artificial shelters at pasture when the ambient temperature, the solar radiation, or the relative humidity increase (Rosselle et al. 2013). However, although our cows provided with shade had free access to it, they made notably more use of it during the last part than during the first part of the summer. Averaged over all sampling days, the use of shade did not increase very steeply with increasing HLI, but on the last four sampling days it did (up to 65% at the highest observed HLI). One could conclude that at the beginning of the pasturing season, the cows were reluctant to use the shaded area because they had not been adequately habituated to it. Nevertheless, the trees had been on their pastures for 2 years, and the shade cloth was hung between trees 1 month before the start of the study. Therefore, it seems that cows need to learn that the shaded area provides them with a protection against high heat load and that the cloth was harmless before they use it efficiently.
The cows did not all use the shade to the same extent. This may be related to their temperament (e.g., cows can differ in their tendency to explore their environment) or their susceptibility to heat stress (due, e.g., to their production level or their percentage of black coat). The position in the hierarchy may also play a role although the shaded area was very large: if all cows were in the shaded area, the space allowance would have been 10.5 m2/cow which is large enough for subordinate cows to avoid the dominant ones. More investigations are necessary to understand why cows vary in their use of shade.
The cows that used the shaded area could maintain their body temperature on hot days. They still panted but to a lesser extent, and their respiratory rate was not significantly affected by HLI. In addition, evening MC levels of these cows were not affected by HLI. Their FCM still increased with HLI but to a lesser extent than in cows without access to shade. The use of shade by cows can thus help them avoid (or at least reduce) heat stress. The fact that the cows using the shade still display some signs of heat stress (panting and increase in FCM) may come from the fact that cows seek shade when the level of heat stress is getting too high and then it may take some time for them to recover. Alternatively, the shade we used may not be enough efficient to remove heat stress completely. Although this was not the case in our study, some types of artificial shelters can actually increase heat load due to thermal radiation from their roof or from the other animals under the shelter. Thus if shade is provided thanks to a shelter, it is essential that its roof is high enough (or insulated) and that the animal density remains low (Berman and Horovitz 2012). It is also essential to design the shelter so as to allow sufficient air movement and to benefit from prevailing winds (Nienaber and LeRoy Hahn 2004; Shoshani and Hetzroni 2013). Heat stress can be reduced by other (or additional) means than providing shelter. Because the digestion of fibers is a large source of heat production by cows, feed ingredients that have a high digestibility should be preferred during hot conditions (e.g., selection of high quality forages, lowering fibers in diet, inclusion of fat,…) (Yasothai 2014)). Therefore, to avoid heat stress in cows at pasture, one needs not only to provide them with shade but also to ensure that the shaded area is adequate and that the grass is of good quality.
In conclusion, weather conditions under a temperate climate—such as in Belgium—can occasionally exceed the cows’ thermal comfort zone on some days in summer, triggering adaptive reactions such as changes in respiration and stress reactions. It is thus necessary to consider taking measures to reduce heat stress, such as providing shade to cows. Additional research into optimal shade area design would be useful in order that cows get easily accustomed to it, and the shade provided is efficient in reducing heat load.
References
Becerril CM, Wilcox CJ, Lawlor TJ, Wiggans OR, Webb DW (1993) Effects of percentage of white coat color on Holstein production and reproduction in a subtropical environment. J Dairy Sci 76(8):2286–2291. https://doi.org/10.3168/jds.S0022-0302(93)77565-7
Berman A, Horovitz T (2012) Radiant heat loss, an unexploited path for heat stress reduction in shaded cattle. J Dairy Sci 95:3021–3031. https://doi.org/10.3168/jds.2011-4844
Cena K, Monteith JL (1975) Transfer processes in animal coats. I Radiative Transfer Proc R Soc Lond B Biol Sci 188(1093):377–393. https://doi.org/10.1098/rspb.1975.0026
Christison GI, Johnson HD (1972) Cortisol turnover in heat-stressed cows. J Anim Sci 35(5):1005–1010. https://doi.org/10.2527/jas1972.3551005x
Collier RJ, Doelger SG, Head HH, Thatcher WW, Wilcox CJ (1982) Effects of heat-stress during pregnancy on maternal hormone concentrations, calf birth-weight and postpartum milk-yield of Holstein cows. J Anim Sci 54(2):309–319. https://doi.org/10.2527/jas1982.542309x
Gantner V, Mijic P, Jovanovac S, Raguz N, Bobic T, Kuterovac K (2012) Influence of temperature-humidity index (THI) on daily production of dairy cows in Mediterranean region in Croatia. In: Casasús I, Rogošiç J, Rosati A, Štokoviç I, Gabiña D (eds) Animal Farming and Environmental Interactions in the Mediterranean Region, vol 131. EAAP, pp 71–78
Gaughan JB, Holt SM, Hahn GL, Mader TL, Eigenberg R (2000) Respiration rate—is it a good measure of heat stress in cattle? Asian Australas J Anim Sci 13:329–332
Gaughan JB, Mader TL, Holt SM, Lisle A (2008) A new heat load index for feedlot cattle. J Anim Sci 86(1):226–234. https://doi.org/10.2527/jas.2007-0305
Hahn GL, Mader TL, Eigenberg RA (2003) Perspective on development of thermal indices for animal studies and management. In: Lacettera N, Bermabucci U, Khalifa HH, Ronchi B, Nardone A (eds) Interactions between climate and animal production. EAAP technical series, vol 7. pp 31–44
Hansen PJ (1990) Effects of coat color on physiological-responses to solar-radiation in Holsteins. Vet Rec 127(13):333–334
Haskell MJ, Rennie LJ, Bowell VA, Bell MJ, Lawrence AB (2006) Housing system, milk production, and zero-grazing effects on lameness and leg injury in dairy cows. J Dairy Sci 89(11):4259–4266. https://doi.org/10.3168/jds.S0022-0302(06)72472-9
Hernandez-Mendo O, von Keyserlingk MAG, Veira DM, Weary DM (2007) Effects of pasture on lameness in dairy cows. J Dairy Sci 90(3):1209–1214. https://doi.org/10.3168/jds.S0022-0302(07)71608-9
Johnson H, Ragsdale A, Berry I, Shanklin M (1963 ) Temperature-humidity effects including influence of acclimation in feed and water consumption of Holstein cattle Missouri Agricultural Experiment Station Research Bulletin 846, Columbia
Kadzere CT, Murphy MR, Silanikove N, Maltz E (2002) Heat stress in lactating dairy cows: a review. Livest Prod Sci 77(1):59–91. https://doi.org/10.1016/s0301-6226(01)00330-x
Kendall PE, Nielsen PP, Webster JR, Verkerk GA, Littlejohn RP, Matthews LR (2006) The effects of providing shade to lactating dairy cows in a temperate climate. Livest Sci 103(1-2):148–157. https://doi.org/10.1016/j.livsci.2006.02.004
Mormède P, Andanson S, Aupérin B, Beerda B, Guémené D, Malmkvist J, Manteca X, Manteuffel G, Prunet P, van Reenen CG, Richard S, Veissier I (2007) Exploration of the hypothalamic-pituitary-adrenal function as a tool to evaluate animal welfare. Physiol Behav 92(3):317–339. https://doi.org/10.1016/j.physbeh.2006.12.003
Mostl E, Palme R (2002) Hormones as indicators of stress. Domest Anim Endocrin 23(1-2):67–74. https://doi.org/10.1016/s0739-7240(02)00146-7
Muller CJC, Botha JA, Coetzer WA, Smith WA (1994a) Effect of shade on various parameters of Friesian cows in a Mediterranean climate in South-Africa .2. Physiological-responses. S Afr J Anim Sci 24:56–60
Muller CJC, Botha JA, Smith WA (1994b) Effect of shade on various parameters of Friesian cows in a Mediterranean climate in South-Africa .1. Feed and water-intake, milk-production and milk-composition. S Afr J Anim Sci 24:49–55
Nienaber JA, LeRoy Hahn G Engineering and management practices to ameliorate livestock heat stress. In: International symposium of CIGR New Trends in Farm Buildings, Evora, 2004
Palme R (2012) Monitoring stress hormone metabolites as a useful, non-invasive tool for welfare assessment in farm animals. Anim Welfare 21:331–337. https://doi.org/10.7120/09627286.21.3.331
Palme R, Mostl E (1997) Measurement of cortisol metabolites in faeces of sheep as a parameter of cortisol concentration in blood. Z Sáugetierkunde 62:192–197
Palme R, Robia C, Baumgartner W, Mostl E (2000) Transport stress in cattle as reflected by an increase in faecal cortisol metabolite concentrations. Vet Rec 146(4):108–109
Palme R, Robia C, Messmann S, Hofer J, Möstl E (1999) Measurement of fecal cortisol metabolites in ruminants: a non-invasive parameter of adrenocortical function. Wien Tierarztl Monatsschr 86:237–241
Palme R, Touma C, Arias N, Dominchin MF, Lepschy M (2013) Steroid extraction: get the best out of faecal samples. Wien Tierarztl Monatsschr 100:238–246
Phillips CJC, Beerda B, Knierim U, Waiblinger S, Lidfors L, Krohn CC, Canali E, Valk H, Veissier I, Hopster H (2013) A review of the impact of housing on dairy cow behaviour, health and welfare. In: Aland A, Banhazi T (eds) Livestock housing: modern management to ensure optimal health and welfare of farm animals. Wageningen Academic Publishers, pp 37–54. https://doi.org/10.3920/978-90-8686-771-4_02
Rees A, Fischer-Tenhagen C, Heuwieser W (2016) Effect of heat stress on concentrations of faecal cortisol metabolites in dairy cows. Reprod Domest Anim 51(3):392–399. https://doi.org/10.1111/rda.12691
Reijs JW, Daatselaar CHG, Helming JFM, Jager J, Beldman ACG (2013) Grazing dairy cows in North-West Europe; economic farm performance and future developments with emphasis on the Dutch situation LEI Report 2013–001, p 124. http://library.wur.nl/WebQuery/wurpubs/fulltext/265398
Rosselle L, Permentier L, Verbeke G, Driessen B, Geers R (2013) Interactions between climatological variables and sheltering behavior of pastoral beef cattle during sunny weather in a temperate climate. J Anim Sci 91(2):943–949. https://doi.org/10.2527/jas.2012-5415
Schutz KE, Cox NR, Tucker CB (2014) A field study of the behavioral and physiological effects of varying amounts of shade for lactating cows at pasture. J Dairy Sci 97(6):3599–3605. https://doi.org/10.3168/jds.2013-7649
Schutz KE, Rogers AR, Poulouin YA, Cox NR, Tucker CB (2010) The amount of shade influences the behavior and physiology of dairy cattle. J Dairy Sci 93(1):125–133. https://doi.org/10.3168/jds.2009-2416
Senft RL, Rittenhouse LR (1985) A model of thermal-acclimation in cattle. J Anim Sci 61(2):297–306. https://doi.org/10.2527/jas1985.612297x
Shoshani E, Hetzroni A (2013) Optimal barn characteristics for high-yielding Holstein cows as derived by a new heat-stress model. Anim Behav Abst 7:176–182
Silanikove N (2000) Effects of heat stress on the welfare of extensively managed domestic ruminants. Livest Prod Sci 67:1–18
Thun R, Eggenberger E, Zerobin K, Luscher T, Vetter W (1981) 24-hour secretory pattern of cortisol in the bull—evidence of episodic secretion and circadian-rhythm. Endocrinology 109(6):2208–2212. https://doi.org/10.1210/endo-109-6-2208
Tucker CB, Rogers AR, Schutz KE (2008) Effect of solar radiation on dairy cattle behaviour, use of shade and body temperature in a pasture-based system. Appl Anim Behav Sci 109:141–154. https://doi.org/10.1016/j.applanim.2007.03.015
Tuyttens FAM, de Graaf S, Heerkens JLT, Jacobs L, Nalon E, Ott S, Stadig L, van Laer E, Ampe B (2014) Observer bias in animal behaviour research: can we believe what we score, if we score what we believe? Anim Behav 90:273–280. https://doi.org/10.1016/j.anbehav.2014.02.007
Umphrey JE, Moss BR, Wilcox CJ, Van Horn HH (2001) Interrelationships in lactating Holsteins of rectal and skin temperatures, milk yield and composition, dry matter intake, body weight, and feed efficiency in summer in Alabama. J Dairy Sci 84:2680–2685
Valtorta S, Leva P, Gallardo M (1997) Evaluation of different shades to improve dairy cattlewell-being in Argentina. Int J Biometeorol 41(2):65–67
Van Laer E, Moons CPH, Ampe B, Sonck B, Vandaele L, De Campeneere S, FAM T (2015a) Effect of summer conditions and shade on behavioural indicators of thermal discomfort in Holstein dairy and Belgian Blue beef cattle on pasture. Animal 9(09):1536–1546. https://doi.org/10.1017/S1751731115000804
Van Laer E, Moons CPH, Ampe B, Sonck B, Vangeyte J, FAM T (2015b) Summertime use of natural versus artificial shelter by cattle in nature reserves. Anim Welfare 24(3):345–356. https://doi.org/10.7120/09627286.24.3.345
Van Laer E, Moons CPH, Sonck B, FAM T (2014) Importance of outdoor shelter for cattle in temperate climates. Livest Sci 159:87–101. https://doi.org/10.1016/j.livsci.2013.11.003
Van Laer E, Tuyttens FAM, Ampe B, Sonck B, Vandaele L (2015c) Effect of summer conditions and shade on the production and metabolism of Holstein dairy cows on pasture in temperate climate. Animal 9:1547–1558
Veissier I, Andanson S, Dubroeucq H, Pomiès D (2008) The motivation of cows to walk as thwarted by tethering. J Anim Sci 86(10):2723–2729. https://doi.org/10.2527/jas.2008-1020
Verkerk GA, Phipps AM, Carragher JF, Matthews LR, Stelwagen K (1998) Characterization of milk cortisol concentrations as a measure of short-term stress responses in lactating dairy cows. Anim Welfare 7:77–86
West JW, Mullinix BG, Bernard JK (2003) Effects of hot, humid weather on milk temperature, dry matter intake, and milk yield of lactating dairy cows. J Dairy Sci 86:232–242. https://doi.org/10.3168/jds.S0022-0302(03)73602-9
Yasothai R (2014) Effect of climate on nutrient intake and metabolism and countering heat stress by nutritional manipulation. Int J Environ Sci Te 3:1685–1690
Acknowledgements
We thank the animal caretakers and technicians at the ILVO Animal Sciences Unit (especially Seppe Holemans, Sara Van Lembergen, and Thijs Decroos) and master student Evelien Van de Keere. Thanks also to Edith Klobetz-Rassam and to Christine Ravel for steroid analyses and to Miriam Levenson (ILVO) for English-language editing.
Funding
This study was funded by the Federal Public Service of Health, Food Chain Safety and Environment (contract RT 10/13 Pastress).
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article

Cite this article
Veissier, I., Van laer, E., Palme, R. et al. Heat stress in cows at pasture and benefit of shade in a temperate climate region. Int J Biometeorol 62, 585–595 (2018). https://doi.org/10.1007/s00484-017-1468-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00484-017-1468-0