
Abstract
Key message
For the first time the interaction of vegetation structure, wind speed and seed attributes on seed suspension on the plants was quantified by means of wind tunnel experiments.
Abstract
The seed hanging above ground, a part of the vertical distribution and a determinant of seed fate, may be affected by vegetation structure. However, how seed hanging on vegetation layers during secondary wind dispersal is linked to vegetation feature has rarely been studied. We investigated the effect of vegetation structure on seed hanging in a wind tunnel. The number of hanging seeds during secondary dispersal of 30 species with various attributes (mass, height, width, length, shape, projected area, wing loading, terminal velocity) was measured in four vegetation structures (pure herb one-layer vegetation, pure shrub one-layer vegetation, herb + shrub uniform two-layer vegetation, and herb + shrub aggregated two-layer vegetation). The proportions of seeds on vegetation layer in one-layer vegetation were significantly more than in two-layer vegetation. The correlation between seed morphological attributes (mass, height, width, length, shape index, and projected area) and the proportion of hanging seeds on vegetation layers differed between the herb and shrub layer but was not affected by vegetation's horizontal and vertical distribution. Our study indicates that the complexity of vegetation structure decreases the seed hanging on the vegetation layer, and the role of seed attributes in determining seed hanging on vegetation layer is modified more largely by the life-form rather than by the spatial structure of vegetation. It is the first systematical study on how vegetation structure affects seed hanging, deeply enhancing our understanding of the effects of vegetation structure on vertical seed distribution.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Diaspore dispersal by wind is one of the most common mechanisms of long-distance dispersal (often referred to as wind seed dispersal) (Leendert 1982; Bouman et al. 2000). After primary seed dispersal (seed movement to the ground from its mother plant), a seed may be blown further through the secondary dispersal process until it is permanently entrapped or germinates (Johnson and Fryer 1992; Greene and Johnson 1997). To a large extent, the final spatial distribution of wind-dispersed seeds is determined by secondary wind dispersal (Schurr et al. 2005). Plant populations and communities are shaped by both the horizontal and vertical distribution of seeds (Schupp 1995; Nathan and Muller-Landau 2000; Levin et al. 2003; Clobert et al. 2012). However, whether primary disperse seeds will form a soil seed bank near the mother plant or move long distance from the mother plant depends on horizontal and vertical movement, and this was one of the primary concerns of previous studies (Brown et al. 1979; Jianhua and Maun 1994; Cechin et al. 2020). Additionally, some diaspores may hang on the plant, which may delay their arrival on the ground and the possibility of the availability of seeds ready to germinate in the right place at the right time. Thus, exploring the effect of vegetation stratification on seed hanging is very important to better understand the vertical distribution of seeds.
Vegetation stratification refers to the vertical arrangement of vegetation in layers (Dierschke 1995). The plant life-forms and resultant stratified layers mainly determine vegetation structure, and the vertical classification of vegetation usually shows the herb, shrub, and tree layers (Dierschke 1995). Life-forms differ in their blade properties, canopy size, branch length, etc. Leaves of herbs, for example, are generally softer and more easily blown by the wind but are harder to intercept moving seeds than those of shrubs. So, there may be differences in seed-hanging ability among vegetation layers. Hence, knowing the seed hanging capacity of the vegetation layer is fundamental to evaluate the vertical distribution in the vegetation.
The vegetative structure mainly includes coverage, vertical (lifeform composition) and horizontal patterns (Oke 1987), which can change wind conditions (Stull 1988) and seed entrapment by plants during secondary wind dispersal (Bochet and Garca-Fayos 2004; Rey 2004). It is speculated that the seed-hanging ability of one vegetation layer would be changed by the structure of vegetation.
Previous studies (involving mainly seed dispersal distance, dispersal kernel, and seed burial) were carried out by comparing the dispersal results on a few types of vegetation (i.e., comparisons of grasslands with mown grassland in reserves, short grasslands with matorral, and open land with dense forest) (Fuentes et al. 1984; Nathan et al. 2002; Soons et al. 2004; Bullock et al. 2017). However, the vegetation used in previous comparative studies consisted of single life form or involved different life-forms but used a single plant configuration. These studies may not sufficiently explore the relationship between vegetation structure and seed dispersal. It is still hard to know the influential vegetation attributes and how they make a difference in seed dispersal processes. Quantifying the attributes of vegetation will help to analyze and predict the final fate of seed dispersal with more precision.
The distribution of seeds after secondary wind dispersal is determined not only by environmental factors (e.g. wind and ground surface) but also by seed attributes (Sheldon and Burrows 1973). Both the flight and entrapment of diaspores are necessary steps for diaspores hanging on the plants. The flight-related attributes of diaspores (like terminal velocity) may directly affect their likelihood of hanging on the vegetation by reflecting the balance between seed gravity and the lift force of the wind (Greene and Johnson 1989). In comparison, other attributes (such as size, shape, and type of appendage) can also indirectly affect their hanging by altering entrapment of seeds (Xueer et al. 2021). However, the exact relationship between seed attributes and seed-hanging events has always been neglected. Furthermore, knowing how the relationship is regulated by the vegetation structure helps us better understand the mechanisms of the vertical distributions of seeds.
For this study, different vertical and horizontal vegetation patterns were structured using an experimental field wind tunnel. Various seed attributes (length, width, height, mass, shape index, projected area, wing loading, and terminal velocity) of 30 species were evaluated to determine the seed-hanging ability of the vegetation layer during secondary dispersal by wind. Specifically, we address two questions: (1) how does vegetation structure influence the seed-hanging ability of the vegetation layer? (2) What is the relationship between seed attributes and the seed-hanging ability of the vegetation layer, and how does the vegetation structure modify this relationship?
Materials and methods
Seed selection
Seeds of 30 species were collected from July 2018 to August 2019. Seeds of different species were selected to represent diverse morphological attributes, not phylogeny. Their attributes were measured after being air dried naturally for three weeks. They were classified into five categories according to their appendage types, i.e., none, one or two wings, more than two wings, hooks, and hair. The winged seeds were classified into two categories because seeds with one or two wings are usually thin slices, while the seeds with more than three wings are usually near-spherical, which makes them behave differently during the hanging process.
Trait measurement
Seed attributes (fruit attributes for some species, but we used “seed attributes” uniformly in this study), such as length, width, height, shape index, mass, projected area, wing loading, and terminal velocity were used since they are generally accepted indicators for wind dispersal and seed attachment to objects (Casper and Grant 1988; Matlack 1992; Casseau et al. 2015). Twenty intact seeds of each species were used for trait measurements. The length, width, and height of the seeds were measured using vernier calipers (0.01 mm accuracy). Shape index (Vs) was calculated as per Thompson et al. (1993):
where N = 3, X1 = length/length, X2 = width/length, and X3 = height/length. Seed mass was measured with an electronic balance (0.01 mg accuracy). Projected area (PA) is the two-dimensional area measurement of a three-dimensional object by projecting its shape onto an arbitrary plane (Palmer 1999). It was calculated using the python software library Open CV (Open Source Computer Vision Library) after scanning with a digital scanner (Berg et al. 2005). Wing loading (W) was defined as the seed mass per projected area and calculated as W = m/p, where ‘m’ is the seed mass and 'p' is the projected area (Matlack 1992). Terminal velocity, a maximum velocity (speed) attainable by an object as it falls through a fluid (Green 1980), was measured by recording seed free fall with a high-speed video with 300 frames per second (Phantom VEO 640L, Ametek, USA) in a black-light-proof box described by Qin et al. (2020). The videos were analyzed using Phantom Camera Control software (https://www.phantomhighspeed.com) as in Liu et al. (2021). The mass range was 0.760–225.147 mg. The length, width, and height ranges were 2.154–33.329 mm 2.051–29.385 mm, and 0.490–28.318 mm, respectively. The projected area range was 3.453–503.932 mm2, and the shape index range was 0.3–1761.8 (Fig. A1 and Table A1).
Wind tunnel
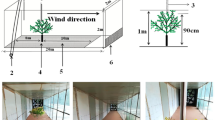
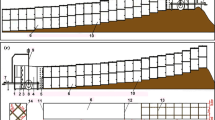
The wind tunnel was 2 m high and 2 m wide, with a test section of 22 m long (Fig. 1). Wind speed was monitored within the tunnel using a hot wire anemometer (AirPro Velocity Meter AP500, TSI, USA). The wind profile is consistent in the direction > 1 m away from the power system and > 8 m from the air outlet (Liu et al. 2015). The experiment surface was set 6 m away from the power system and 9.4 m away from the air outlet. Smooth and flat planks were paved between the power system and the experiment surface. The total planks were 6 m long and 2 m wide (Fig. 1). The anemometer was located 7.6 m from the power system. It was fixed on the roof, and wind speed was measured 1 m above the underlying surface.

Structure of the wind tunnel and the position of anemograph probe (a), the starting position of seeds in bared land or vegetation (b)
Vegetation arrangements
The underlying surface matrix consisted of aeolian sand from a moving sand dune in Inner Mongolia, China (105° 50′ E, 41° 02′ N). The sand was naturally air dried and then sieved with a mesh size of 5 mm to remove non-substrate impurities (such as stones and pieces of plants).
The sand matrix was manually made flat. Herbs and shrubs were manually fixed on the flat surface as the vegetation. These vegetations differed in their coverage, life-form composition, and horizontal patterns (Table 1 and Fig. 2).

Schematic for the size of the plants and their distribution in four vegetation structures
Artificial plastic herbs were used as experimental vegetation material to prevent damage from strong wind and to make uniformly sized replicas. The height, diameter, and projected area of the herb crown were 5.0 ± 0.32 cm, 5.0 ± 0.33 cm, and 18.08 ± 1.36 cm2, respectively. A centimeter-long nail was pierced through the center of the herb and the underlying surface was used to fix the herb. The whole crown of the herb was on the surface.
Convolvulus tragacanthoides was selected as the shrub model because it had thick and sturdy branches with few small leaves, which were easier to shape crown, set up replication, and prevent from being broken by the strong wind. They were dug in Inner Mongolia, China (106° 52′ E, 40° 08′ N). The clipped shrub crown's height, diameter, and projected area were 20.0 ± 1.52 cm, 22.0 ± 1.68 cm, and 320.0 ± 18.53 cm2, respectively. For each shrub, a roughly centimeter long taproot was kept and the leaves were removed. A 15 cm long hollow plastic pipe was pieced in the underlying surface to fix the shrub. The shrub root was put in the pipe's hole filled with sand, and the whole crown of the shrub was on the surface.
Measurement of hanging seed proportion
The wind velocity (measuring 1 m height from the ground) were set at 3.00, 5.25, 7.50, 9.75, and 12.00 m·s−1, increased by 2.25 m·s−1 between two adjacent wind speeds. Thirty species were used to conduct the experiments on vegetation. Seeds were put in the line in the middle of two rows of plants, and the seed position along the line was randomly selected. After landing, the diaspores were covered with an iron cover. The iron cover was immediately pulled up to release the seeds as the wind reached the targeted speed. Thirty seconds later, the wind tunnel was turned off. After seed dispersal, seed entrapped by plants and not touching the ground can be considered seed hanging on plants. The number of hanging seeds and plant life-form was recorded. The experiment was repeated five times for each species.
Data analysis
One-way ANOVA was performed using a rank sum test (Kruskal–Wallis) using IBM SPSS Statistics 22.0 (IBM Corporation 1989 and 2013, USA) to analyze changes in the proportions of hanging seeds between vegetation layers in each vegetation and wind velocity and seed appendage types in each vegetation layer and wind velocity. The correlation coefficients were calculated using the spearman coefficient (IBM SPSS Statistics 22.0, IBM Corporation 1989 and 2013, USA) to analyze the relationship between quantitative seed attributes and the proportions of hanging seeds in each vegetation layer and wind velocity.
Results
Relationship between vegetation structure and hanging seed proportion
The proportions of hanging seeds on herb layer in one-layer vegetation H(U) were consistently higher than that in two-layer vegetations H + S(U) and H + S(A) at each wind velocities (Fig. 3, left of the dashed line), and there was no significant difference between the proportions of hanging seeds on herb layer in H + S(U) and H + S(A) (Fig. 3, left of the dashed line). Like herb layer, it was the same for shrub layer (Fig. 3, right of the dashed line). Generally, the average proportions of hanging seeds for each wind velocity on herb layer or shrub layer in the one-layer vegetations [H(U) or S(U)] were significantly higher than that in the two-layer vegetations [H + S(U) and H + S(A)] (Fig. 3).

The proportion of seeds hanging on vegetation layer in vegetation differs in vertical (life-form composition) and horizontal patterns. Figures show the overall proportions found on the herb layer (left of the dashed line) and the shrub layer (right of the dashed line) in degrees of wind velocity and the average for each wind velocity (the rows)
There was no significant difference in the proportions of hanging seeds between herb layer in H(U) and shrub layer in S(U) (Fig. 4, left column), as well as between herb layer and shrub layer in H + S(U) and H + S(A) (Fig. 4, middle and right columns).

The proportion of seeds hanging on herb layer and shrub layer. Figures show the overall proportions found on the herb layer and shrub layer in H(U) and S(U) (the left column), H + S(U) (the middle column), and H + S(A) (the right column) in degrees of wind velocity and the average for each wind velocity (the rows)
Relationship between seed appendage type and hanging seed proportion
The orders of appendage types according to the hanging seed proportions were generally consistent across each wind velocity for both the herb and shrub layer (Fig. 5). However, when the wind velocity was 3.00 m•s−1, only the hairy seeds could be hung on the plant. The average proportion of hanging seeds was 15.39% and 8.93% for the herb and shrub layer, respectively (Fig. 5). After the wind velocity exceeded 5.25 m s−1, the order of appendage types was arranged as follows: none ≤ hooks < no more than two wings < more than two wings < hair (for the herb layer) and none ≤ hooks < no more than two wings = more than two wings < hair (for the shrub layer (Fig. 5).

The proportion of seeds with different appendages hanging on vegetation layer in vegetation differs in vertical (lifeform composition) and horizontal patterns. Figures show the overall proportions found on the herb layer (left of dashed line) and shrub layer (right of dashed line) and in degrees of wind velocity (the rows)
Relationship between quantitative seed attributes and hanging seed proportion
For the herb layer, wing loading and terminal velocity were always the most critical seed attribute determining seed hanging in all 15 treatments except for one treatment, vegetation H + S(A) under wind velocity of 12.00 m s−1 (Table 2). Seed mass, shape index, terminal velocity, and wing loading were negatively correlated with hanging seed proportions except for mass in the vegetation H + S(A) under wind velocity 12.00 m s−1. In contrast, projected area, length, width, and height were positively correlated with hanging seed proportions across all 15 treatments (Table 2).
Similarly, to the shrub layer, the wing loading and terminal velocity determined the seed hanging in the 15 treatments except for two treatments, that is, the one-layer S(U) and two-layer H + S(A) vegetation, respectively, under the wind speed of 9.75 m s−1 (Table 3). The seed mass and shape index were hardly significantly related to seed hanging. Terminal velocity and wing loading negatively correlated with hanging seed proportions, while projected area, length, width, and height positively correlated with hanging seed proportions across all 15 treatments (Table 3).
Discussion
Effect of vegetation structure on the seed-hanging ability of vegetation layer
The seed-hanging ability of vegetation layer in one-layer vegetation was always higher than in two-layer vegetation. In contrast to this, there were no significant differences in the seed-hanging capacity between herb layer and shrub layer, and between two-layer vegetation of uniform vegetation and aggregated vegetation. Consequently, the vertical complexity of the vegetation structure can weaken the seed-hanging ability of a vegetation layer no matter what the horizontal vegetation pattern and the plant life-form are.
When a vegetation layer's coverage remains unchanged, other vegetation layers' presence can increase the coverage of the entire vegetation. It is known that the horizontal and vertical wind speeds decrease with vegetation coverage (Nuttle and Haefner 2005). So, seeds will get less wind force to fly and hang on the vegetation layers as the vegetation is vertically stratified more complicatedly. In addition, once a flying seed hangs on a vegetation layer, it means its entrapment, and there is little possibility for the seed to move and hang on other vegetation layers. In other words, multiple layers coexisting in vegetation seem to compete for the priority of seed entrapment so that they decrease the seed hanging possibility of each other.
Horizontally, herb layer is able to hang more seeds than shrub layer due to its larger total perimeter. While vertically, shrub layer is able to hang more seeds than herb layer due to its greater average height, thereby explaining the overall absence of significant differences in seed hanging between herb and shrub layers.
In this study, the proportion of hanging seeds was approximately 8% of the total secondary wind dispersal seeds. Therefore, studying seed hanging mechanisms is worth understanding how seeds vertically distribute in vegetation. It could enrich our knowledge about seed dispersal and help make better predictions for vertical seed distribution according to the vegetation structure.
Effect of seed attributes on seed hanging
For the herb and shrub layers, the percentage of seed hanging on vegetation layers increases with the decrease of seed terminal velocity and wing loading. However, there are still differences in the attributes of hanging seeds in the canopy of different life forms. No matter what vertical stratification and horizontal vegetation patterns were, hanging on the herb layer tended to dominate by those seed attributes that were lightweight, spheroidal and bigger in length, width, height, and projected area. Hanging on the shrub layer was also dominated by similar seed attributes but with the minor role of seed weight and shape. Consequently, the relationship between seed attributes and seed hanging on the vegetation layer was not modified by the horizontal distribution and vertical complexity but by plants' life forms.
During secondary wind dispersal, flight and entrapment were the two decisive steps in seed hanging. Terminal velocity reflects the balance between vertical upward wind force and gravity (Greene and Johnson 1989), so the lower the terminal velocity, the lower the wind force can raise the seed to fly. Meanwhile, terminal velocity has shown a strong positive correlation to wing loading (Augspurger and Franson 1987; Green 1980). Consequently, terminal velocity and wind loading are effective indicators for predicting seed hanging.
The seeds hung in different layers of vegetation exhibited different properties. Compared to shrubs, the softer/smoother leaves and branches of herbs tend to have a much weaker load-bearing capacity. Therefore, the hanging of seeds on herbs is more sensitive to seed mass than on shrubs. Compared with herbs, wider gaps among branches of shrubs make them more difficult to entrapped tiny seeds. Thus, seeds hanging on shrubs are more sensitive to seed length, width, height, and projected area than that on herbs. Seeds with hooks or appendages could not hang on plants because they could not fly or be entrapped. Compared to seeds with one or two wings (which are usually thin slices and prolate), the spheroidal seeds with hairs or multiple wings are easy to fly and have more dimensionalities to hold. Although thin seeds hanging on herbs are less than seeds with hairs or multiple wings, they can be pierced by the thorny branches or other sharp structures of shrubs. Therefore, seed hanging on shrubs was not influenced by the shape index of the seed, while seed hanging on herbs did.
Seed entrapment involves direct contact between seeds and plants, so the possibility of seed entrapment is mainly determined by plant characteristics and seed morphological attributes (mass, shape, length, width, height, projected area, and appendage type) when the seeds encounter different plant life-form in vegetation structure. Seed flight is the necessary step in seed hanging. Therefore, the aerodynamic attributes (terminal velocity and wing loading) determining seed flight capacity always play crucial role in seed hanging in all types of vegetation. Consequently, it is the morphological attributes of seeds rather than the aerodynamic attributes that determines the plant life-form the seed tends to hang on.
Conclusion
The vertical complexity of vegetation structure can weaken the seed-hanging on vegetation layer during secondary wind dispersal. Compared to the spatial vegetation structure, the role of seed attributes in determining seed hanging on vegetation layer is more predominantly modified by plant life-form, and the plant life-forms seeds tend to hanging on are determined by seed morphological attributes. This study could help comprehend the relationship between seed attributes and seeds hanging in stratified vegetation, which eventually predict the vertical composition of seeds (species) in vegetation and their fate in population regeneration ecology.
Data availability
The data supporting the findings of this study are openly available in the Zenodo repository at https://zenodo.org/record/8319283.
References
Augspurger CK, Franson SE (1987) Wind dispersal of artificial fruits varying in mass, area, and morphology. Ecology 68:27–42
Berg AC, Berg TL, Malik J (2005) Shape matching and object recognition using low distortion correspondences. In: Proceedings—2005 IEEE computer society conference computer vision pattern recognition, CVPR 2005 I, pp 26–33
Bochet E, García-Fayos P (2004) Factors controlling vegetation establishment and water erosion on motorway slopes in Valencia, Spain. Restor Ecol 12:166–174
Bouman F, Boesewinkel FD, Bregman R et al (2000) Verspreiding van zaden. KNNV Uitgeverij, Utrecht
Brown JH, Reichman OJ, Davidson DW (1979) Granivory in desert ecosystems. Annu Rev Ecol Syst 10:201–227
Bullock JM, Mallada González L, Tamme R et al (2017) A synthesis of empirical plant dispersal kernels. J Ecol 105:6–19
Casper BB, Grant BW (1988) Morphology and dispersal of one- and two-seeded diaspores of Cryptantha flava. Am J Bot 75:859–863
Casseau V, De Croon G, Izzo D, Pandolfi C (2015) Morphologic and aerodynamic considerations regarding the plumed seeds of Tragopogon pratensis and their implications for seed dispersal. PLoS ONE 10:1–17
Cechin J, Schmitz MF, Hencks JR et al (2020) Burial depths favor Italian ryegrass persistence in the soil seed bank. Sci Agric 78
Clobert J, Baguette M, Benton TG, Bullock JM (2012) Dispersal ecology and evolution. Oxford University Press, New York
Fuentes ER, Otaiza RD, Alliende MC et al (1984) Shrub clumps of the Chilean matorral vegetation: structure and possible maintenance mechanisms. 62:405–411
Green DS (1980) The terminal velocity and dispersal of spinning samaras. Am J Bot 67:1218–1224
Greene DF, Johnson EA (1997) Secondary dispersal of tree seeds on snow. J Ecol 85:329–340
Greene DF, Johnson EA (1989) A model of wind dispersal of winged or plumed seeds. Ecology 70:339–347
Hartmut Dierschke (1995) Pflanzensoziologie-Grundlagen und Methoden. Stuttgart, German
Johnson EA, Fryer GI (1992) Physical characterization of seed microsites—movement on the ground. Br Ecol Soc 80:823–836
van der Pijl L (1982) Principles of dispersal in higher plants, 3rd edn. Springer, Berlin
Levin SA, Muller-Landau HC, Nathan R, Chave J (2003) The ecology and evolution of seed dispersal: a theoretical perspective. Annu Rev Ecol Evol Syst 34:575–604
Liu M, Zhu J, Xin Z et al (2015) A portable wind tunnel for studying seed dispersal by wind: test and evaluation. Chin J Ecol 34:1770–1778
Liu M, Xin Z, Su Z et al (2021) A video camera recording method for measuring terminal velocity of seed dispersal by wind. J for Res 32:81–90
Matlack GR (1992) Influence of fruit size and weight on wind dispersal in Betula lenta, a gap-colonizing tree species. Am Midl Nat 128:30–39
Nathan R, Horn HS, Chave J, Levin SA (2002) Mechanistic models for tree seed dispersal by wind in dense forests and open landscapes. Seed dispersal frugivory. Ecol Evol Conserv 69–82
Nathan R, Muller-Landau HC (2000) Spatial patterns of seed dispersal, their determinants and consequences for recruitment. Trends Ecol Evol 15:278–285
Nuttle T, Haefner JW (2005) Seed dispersal in heterogeneous environments: bridging the gap between mechanistic dispersal and forest dynamics models. Am Nat 165:336–349
Oke TR (1987) Boundary layer climates. Routledge, London
Palmer JM (1999) Radiometry and photometry FAQ. Radiom Radiom Photom FAQ 1–14
Qin X, Liu Z, Liu M, et al (2020) Shrub canopy interception of diaspores dispersed by wind. Seed Sci Res
Rey F (2004) Effectiveness of vegetation barriers for marly sediment trapping. Earth Surf Process Landf 29:1161–1169
Schupp EW (1995) Seed-seedling conflicts, habitat choice, and patterns of recruitment. Bot Soc Am 82:399–409
Schurr FM, Bond WJ, Midgley GF, Higgins SI (2005) A mechanistic model for secondary seed dispersal by wind and its experimental validation. J Ecol 93:1017–1028. https://doi.org/10.1111/j.1365-2745.2005.01018.x
Sheldon JC, Burrows FM (1973) the dispersal effectiveness of the achene-pappus units of selected compositae in steady winds with convection. New Phytol 72:665–675
Soons MB, Heil GW, Nathan R, Katul GG (2004) Determinants of long-distance seed dispersal by wind in grasslands. Ecology 85:3056–3068
Stull RB (1988) An introduction to boundary layer meteorology. Kluwer Academic Publishers, Dordrecht
Thompson K, Band SR, Hodgson JG (1993) Seed size and shape predict persistence in soil. Funct Ecol 7:236–241
Xueer W, Ke G, Guili J et al (2021) Effects of different animals on seed dispersal characteristics of Achnatherum inebrians. Acta Physiol Sin 29:2240–2246
Jianhua Z, Maun MA (1994) Potential for seed bank formation in seven Great Lakes sand dune species. Am J Bot 81:387–394. https://doi.org/10.1002/j.1537-2197.1994.tb15461.x
Acknowledgements
We thank the Experimental Center of Desert Forestry, Chinese Academy of Forestry and the First Forestry Station for providing accommodation during the experiment.
Funding
This work was supported by the National Natural Science Foundation of China (32171870, 42007427 and 31971732).
Author information
Authors and Affiliations
Contributions
ZL, QZ, WL and LT conceived the ideas and designed the study. LT, LZ, CB, XL, JW, SZ, ZX, ML, XQ and YL collected the data. LT analyzed the data and led the writing of the first draft of the manuscript. WL, SSP, and ZL revised several drafts of the manuscript. All authors gave final approval for publication.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflicts to declare.
Additional information
Communicated by Bräuning.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Tian, L., Liang, W., Liu, Z. et al. The complexity of vegetation structure decreases the seed-hanging ability of the vegetation layer during secondary wind dispersal. Trees 37, 1671–1680 (2023). https://doi.org/10.1007/s00468-023-02451-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-023-02451-z