
Abstract
Key message
Pinus sylvestris tree-ring δ13C and δ18O records from locally moist sites in central and northern Sweden contain consistently stronger climate signals than their dry site counterparts.
Abstract
We produced twentieth century stable isotope data from Pinus sylvestris trees near lakeshores and inland sites in northern Sweden (near Kiruna) and central Sweden (near Stockholm) to evaluate the influence of changing microsite conditions on the climate sensitivity of tree-ring δ13C and δ18O. The data reveal a latitudinal trend towards lower C and O isotope values near the Arctic tree line (− 0.8‰ for δ13C and − 2.4‰ for δ18O relative to central Sweden) reflecting widely recognized atmospheric changes. At the microsite scale, δ13C decreases from the dry inland to the moist lakeshore sites (− 0.7‰ in Kiruna and − 1.2‰ in Stockholm), evidence of the importance of groundwater access to this proxy. While all isotope records from northern and central Sweden correlate significantly against temperature, precipitation, cloud cover and/or drought data, climate signals in the records from moist microsites are consistently stronger, which emphasizes the importance of site selection when producing stable isotope chronologies. Overall strongest correlations are found with summer temperature, except for δ18O from Stockholm correlating best with instrumental drought indices. These findings are complemented by significant positive correlations with temperature-sensitive ring width data in Kiruna, and inverse (or absent) correlations with precipitation-sensitive ring width data in Stockholm. A conclusive differentiation between leading and co-varying forcings is challenging based on only the calibration against often defective instrumental climate data, and would require an improved understanding of the physiological processes that control isotope fractionation at varying microsites and joined application of forward modelling.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Stable carbon and oxygen isotope ratios (δ13C and δ18O values) are valuable proxies for reconstructing long-term climatic changes at annual resolution (Leavitt 2010). Compared to classical tree-ring width (TRW) and maximum latewood density data (MXD), which predominantly record temperature changes in cold environments (Esper et al. 2014; Luterbacher et al. 2016; Wilson et al. 2016) and hydroclimatic changes in dry environments (Cook et al. 2015; Esper et al. 2007; Tejedor et al. 2017), climate signals in stable isotopes appear to be less dependent on the ecoclimatic settings of the sampled trees (Esper et al. 2016; Frank et al. 2015; Gagen et al. 2004; Saurer et al. 2008; Treydte et al. 2007). This independency enables the use of tree-ring stable isotope records from lowland, non-boundary environments, where classical tree-ring parameters typically fail to record significant climate signals (Cernusak and English 2015; Friedrichs et al. 2008; Hartl-Meier et al. 2015) to assess long-term climatic changes at annual resolution (overview in Treydte et al. 2007, 2014).
In their analysis of a network of 23 δ13C chronologies from Europe, Frank et al. (2015) showed that the maximum climatic response exhibited by that proxy encompasses a broad range of climatic parameters including relative humidity, potential evapotranspiration, maximum temperature, vapor pressure deficit and precipitation during different months and seasons during the year of ring formation. Formal reconstructions based on δ13C include estimates of summer temperature variability in Central Asia (Treydte et al. 2009), winter-to-spring temperature in Turkey (Heinrich et al. 2013), summer drought in the European Alps (Kress et al. 2010), and river flow in west Siberia (Waterhouse et al. 2000). The climate sensitivity of tree-ring δ18O is typically (but not always) slightly weaker compared to δ13C, and includes precipitation, drought and humidity signals as mechanistic drivers in addition to widely recognized temperature signals (Saurer et al. 2008; Treydte et al. 2007). Formal reconstructions based on δ18O include estimates of annual precipitation in the Karakorum mountains (Treydte et al. 2006), May–August precipitation in northwest China (Liu et al. 2008) and southern England (Rinne et al. 2013), the summer standardized precipitation evapotranspiration index (SPEI) in western France (Labuhn et al. 2016), and April–July minimum temperatures in northwest Canada (Porter et al. 2014). Saurer et al. (2008) suggested the combination of tree-ring δ13C and δ18O may enhance the common climatic signals, an approach that was successfully carried out in northeast Canada to reconstruct summer maximum temperatures (Bégin et al. 2015).
In northern Europe, Scots pine (Pinus sylvestris L.) is the most common tree species and has been used in several stable isotope studies. Seftigen et al. (2011) analyzed δ13C and δ18O from sites in the Scandinavian mountains, and reported positive correlations with warm season temperatures, and reversed but weaker associations with precipitation. Similarly, Hilasvuori et al. (2009) found July temperature signals in δ13C (stronger) and δ18O (weaker) in northern and eastern Finland, as well as July precipitation signals in δ18O of eastern Finland. More recently, δ13C chronologies from Scandinavian pine sites were used to reconstruct sunshine hour (Loader et al. 2013) and cloud cover changes (Gagen et al. 2011). In all these assessments, the covariance among temperature, precipitation, drought indices, cloud cover and sunshine hour data was recognized as a complicating data property that makes it difficult to differentiate between directly environmental forcing and their co-variates (Frank et al. 2015).
δ13C and δ18O interseries correlations and climatic signals were shown to be particularly strong in the high-frequency, inter-annual domain (Konter et al. 2014). The proxies are good estimators for the reconstruction of extreme events (Treydte et al. 2001; Kress et al. 2009), but typically express less coherent lower frequency trends (Treydte et al. 2007). Work on the preservation of low frequency variance in tree-ring δ13C and δ18O of Pinus uncinata showed that both parameters contain biological age trends beyond the widely recognized juvenile effects over the first 20–30 years (Esper et al. 2010). This finding was recently confirmed for δ13C using a large compilation of modern tree and sub-fossil Pinus sylvestris samples from Finland (Helama et al. 2015), questioning the application of such data to reconstruct low frequency climate variability (Esper et al. 2015a).
Tree-ring stable isotopes reflect the plant physiological response to climate and other environmental variables (Gessler et al. 2014; Zeng et al. 2017). δ13C depends on factors affecting the photosynthetic uptake of CO2 and is primarily controlled by stomatal conductance and the rate of carboxylation during photosynthesis (Farquhar et al. 1989). Warm and dry conditions typically reduce stomatal conductance and discrimination against 13C, thus producing higher δ13C values (Leavitt and Long 1989; Saurer et al. 1995). In light-limited habitats, however, carbon isotope fractionation is dominated by photo-assimilation, and high δ13C values may primarily be associated with high photosynthetic activity in warm and sunny conditions (Loader et al. 2013).
Tree-ring δ18O integrates the stomatal response to atmospheric vapour pressure deficit and related climate variables, such as temperature and relative humidity, via leaf water 18O enrichment. Transpiration also regulates source water uptake by the roots (Barbour 2007; Roden and Ehleringer 1999), which typically originates from precipitation carrying a specific atmospheric δ18O signal controlled by air mass temperature (Dansgaard 1964; Rozanski et al. 1992). This precipitation signal can be mitigated, lagged or even masked, depending on the temporal variation of the amount and isotopic composition of infiltrated water (Treydte et al. 2014), the evaporative enrichment of soil water, and the influence of ground water (Ehleringer and Dawson 1992). High soil water availability in correspondence with warm and dry conditions enhances evaporative processes at the leaf level (Darling and Talbot 2003), which may lead to high δ18O tree-ring values and partly be modified by post-photosynthetic biochemical fractionation processes (Gessler et al. 2014).
Considering these specific sensitivities, and by employing a network of locally moist and dry Pinus sylvestris sites in central and northern Sweden, we address the following hypotheses:
(H1)
Tree-ring δ13C and δ18O values are lower in the colder environment of northern Sweden due to reduced photosynthesis and enzymatic fractionation (δ13C), relatively low transpiration and temperature effects on source water isotope fractionation (δ18O).
(H2)
At both sites, δ13C and δ18O values are lower in lakeshore trees due to higher soil moisture and stomatal conductance in dry periods (δ13C) and unhindered access to depleted groundwater (δ18O).
(H3)
δ13C and δ18O records from locally moist sites contain stronger climate signals because of increased metabolic activity and hence, isotope fractionation being more closely controlled by atmospheric weather processes. The signals are stronger in the warmer Stockholm region with larger implications of groundwater access, relative to Kiruna.
In this paper, we describe the microsite sampling scheme in central and northern Sweden, the procedures of tree-ring δ18O and δ13C measurement, and the statistical methods used to treat stable isotope data and compare these with instrumental climate data. We produce maps showing the spatial patterns of correlation between stable isotope and climate variables, and discuss differences in climate signals with particular focus on microsite effects. δ18O and δ13C chronologies are compared with TRW chronologies to support the interpretation of signal differences between locally moist and dry sites.
Materials and methods
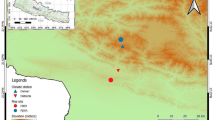
The Pinus sylvestris sampling sites are located near Kiruna (hereafter, “Kir”) at 68°N and ~ 1000 km north of the Stockholm (“Sto”) site at 59°N (Fig. 1). Kir and Sto are underlain by a Quaternary moraine including granite blocks of up to 1 m in size. The loose substrate is 0–25 cm thick and the soil type is a sandy podsol covered by a thin (< 3 cm) litter layer. Both sites are situated in the rain shadow of the Scandes, where summer precipitation approaches only 200 mm, but temperatures are 7–9 °C colder at the Arctic tree line in Kir compared to Sto (Table 1). The temperature difference affects both evaporative demand (Farquhar and Roderick 2007) and species composition, so that Kir is composed of a monotonic pine forest, whereas Sto hosts a mixed forest including Picea abies, Quercus robur, Fagus sylvatica, and Pinus sylvestris. The vegetation period in Kir is 60 days shorter compared to Sto (Swedish Meteorological and Hydrological Institute, SMHI), but seasonal daylight variations are much more accentuated in northern than central Sweden. The distances between trees range from 7 to 10 m in Sto and from 3 to 10 m in Kir, and the hydroclimatic conditions during the vegetation period are classified as “weakly humid” in Kir (positive water balance of 0 to + 50 mm) and “dry” in Sto (negative water balance of − 50 to 0 mm; Swedish University of Agricultural Sciences).

Tree-ring stable isotope setup in northern (Kir) and central (Sto) Sweden. In each region, four Pinus sylvestris trees were sampled in locally moist (at lakeshore) and dry (several decameters inland) microsites. Values are the mean carbon (green) and oxygen isotope ratios (brown) of four trees in each microsite, as well as the difference between Kir and Sto. Standard deviations range from 0.18 (Kir-D) to 0.65 (Sto-D) for δ18O, and from 0.09 (Kir-M) to 0.46 (Sto-M) for δ13C
At both sites, Kir and Sto, we used 5-mm increment borers to sample 40 pine trees growing in moist conditions within 5 m to the lakeshore (Kir-M and Sto-M) and several tens of meters inland in locally dry conditions considering all age classes in pre-defined plots (Kir-D and Sto-D; Düthorn et al. 2015, 2016). The dry microsites are located 15–30 m upslope (Kir microsites in 450 and 465 m, Sto microsites in 20 and 50 m a.s.l.) where tree roots are detached from lake filtration and phreatic water (Fig. 1). This change also impacts ground vegetation as Kir-M includes 90% bryophytes and 10% other species (Calluna vulgaris, Vaccinium vitis-idaea, Arctophylos uva-ursi, Empetrum nigrum, Vaccinium myrtillus), whereas Kir-D hosts no bryophytes but 50% lichen species. Vaccinium vitis-idaea and various lichen species are also present in Sto-D, while Sto-M is additionally covered by Vaccinium myrtillus and Calluna vulgaris. All increment core samples from these microsites were cut using a microtome, TRW measured at 1/100 mm resolution using a LINTAB station (Rinn 2005), and samples crossdated considering standard dendrochronological techniques (Holmes 1983). Mean growth rates range from 0.83 mm/year in Kir-M to 0.97 mm/year in Kir-D, and from 1.00 mm/year in Sto-D to 1.15 mm/year in Sto-M, and mean tree ages range from 77 years in Kir-D to 83 years in Kir-M, and from 93 years in Sto-D to 119 years in Sto-M (for more details see Düthorn et al. 2013).
In each microsite, Kir-M, Kir-D, Sto-M and Sto-D, four trees (two cores per tree, pooled) were selected for stable isotope measurement considering the criteria of homogeneous growth, absence of missing rings, ring width > 0.01 mm, well-defined annual rings, and minimum age of 150 years. Hydrophilic extractives and lipophilic compounds were removed by purging the increment cores with distilled water and water-free ethanol for 8–24 h at 60 °C (Sidorova et al. 2008). Annual tree-rings covering the period 1901–2009 were truncated with a scalpel, and whole wood samples (Mischel et al. 2015; Riechelmann et al. 2014, 2016) of 300–3000 µg packed in tin capsules for stable carbon isotope measurement, and 250–300 µg samples in silver capsules for stable oxygen isotope measurement. The tin capsules were combusted in an elemental analyzer at 1150 °C, 13C/12C measured using an IsoPrime IRMS, and δ13C expressed in parts per thousand (‰) with respect to an established reference material (Farquhar et al. 1982). For δ18O, the silver capsules were pyrolized at 1450 °C, 18O/16O quantified in an IRMS and expressed in ‰ relative to the standard isotopic composition of ocean water (Leavitt 2010). The analytical uncertainties of these data are ± 0.3‰ for δ13C and ± 0.5‰ for δ18O, and the microsite chronologies denoted Kir13D, Kir13M, Kir18D, etc.
Numerous studies have applied δ13C and δ18O values in tree-ring material such as cellulose, lignin, whole wood or lignin methoxyl groups for climatic and environmental studies (Barbour et al. 2001; Edwards and Fritz 1986; Gori et al. 2013; Loader et al. 2003; Mischel et al. 2015; Riechelmann et al. 2016). Acknowledging the findings of Wilson and Grinsted (1977) that tree-ring components have different isotopic ratios, most research has been carried out on single components, usually cellulose, and on single isotope proxies preferentially δ13C and δ18O (Boettger et al. 2007). However, the time-consuming cellulose extraction steps are still a disadvantage particularly in view of the need to develop long-term chronologies for climatic reconstruction. Alternatively, in recent years a number of studies have demonstrated the usefulness and effectiveness of δ13C, δ2H and δ18O of whole wood as a tree-ring climate proxy (Cullen and Grierson 2006; Gori et al. 2013; Loader et al. 2003; Mischel et al. 2015; Verheyden et al. 2005; Weigt et al. 2015). Gori et al. (2013) investigated Picea abies in alpine Italy and demonstrated that δ18O and δ2H from whole wood samples preserve the best temperature signals. Schleser et al. (2015) found a high correlation between δ13C values from whole wood and extracted cellulose of tropical Cariniana micrantha over the past two and a half centuries, and Mischel et al. (2015) report on differences between whole wood and cellulose δ13C and δ18O timeseries of Pinus sylvestris supporting the choice to analyse whole wood instead of cellulose. For practical reasons and to substantially reduce costs, we measured δ13C and δ18O of the bulk tree-ring material of Pinus sylvestris.
Each δ13C series was corrected for the Suess effect to remove a long-term declining trend since the mid-nineteenth century due to the combustion of fossil carbon and atmospheric enrichment of depleted CO2 (Farquhar et al. 1989; Treydte et al. 2009). In addition, all isotopic data (including δ18O) were detrended using 30-year spline high-pass filters (Cook and Peters 1981) to remove level differences among single-tree isotope series and long-term trends that are potentially unrelated to climatic forcings. We use both versions of the data, the non-detrended (“raw”) and the spline-detrended (“spl”) stable isotope timeseries for calibration against instrumental climate data and report the highest agreements (Table 2). Gridded monthly mean temperatures, precipitation sums, cloud cover changes (Harris et al. 2014), and Palmer Drought Severity Index (PDSI; van der Schrier et al. 2006) reaching back to 1901 were used for calibration against tree-ring δ18O and δ13C using Pearson correlations. The gridded products were accessed through the KNMI climate explorer (Trouet and van Oldenborgh 2013; van Oldenborgh and Burgers 2005) at 0.5° × 0.5° resolution and were employed to evaluate proxy climate signals over two overlapping intervals: 1901–2009 and 1951–2009. The latter period is used to evaluate the influence of potentially uncertain early instrumental data (Frank et al. 2007) as well as changes in proxy signal strength (Briffa et al. 1998b). Maps are produced highlighting correlations exceeding p < 0.05 to evaluate spatial patterns of climate signals. We also produced crossplots and linear regressions of selected isotope-climate associations and calculated 30-year running correlations to emphasize temporal changes in proxy climate signals.
Results and discussion
Average isotope ratios and covariance
On average, stable isotope ratios of tree-ring bulk wood increase by 0.8‰ (δ13C) and 2.4‰ (δ18O) from the Arctic tree line in northern Sweden towards Stockholm ~ 1000 km south of the ecotone, thereby verifying H1 for both proxies (Fig. 1). For oxygen, this offset is likely reflecting the temperature difference between northern and central Sweden (up to 10 °C), which controls the isotopic composition of atmospheric water supply (Kortelainen and Karhu 2004). Tree-ring δ18O does, however, not change between the moist and dry microsites, disproving H2 on the influence of depleted groundwater for this proxy: immediate access to lake filtration water has no effect on average oxygen isotope signatures of pine tree-rings. Snowmelt as an important water source, particularly for trees at dry microsites and reaching far into the growing season (Treydte et al. 2014), likely affects the isotopic signature and differentiation between lakeshore and inland sites. This situation is different for δ13C, which is not only depleted in 13C towards the artic tree line (Hilasvuori et al. 2009; Stuiver and Braziunas 1987), but also from the dry to the moist microsites. The large-scale change is likely a representation of increased photosynthetic assimilation in Sto, as reflected by wider tree-rings (average growth rate over the first 100 years of tree age in Sto = 1.10 mm, in Kir = 0.78 mm), regulating sub-stomatal CO2 concentrations and forcing the leaf enzyme Rubisco to catalyze more of the heavy 13C isotope (Saurer et al. 2004). At the local scale, the δ13C offset is likely caused by prolonged stomatal resistance in the dry microsites during warm season drought periods of several days to weeks (Farquhar et al. 1989). The effect seems to be slightly stronger in the warmer Sto site, in which the offset between dry and wet microsites reaches 1.2‰, compared to only 0.7‰ in Kir.
The δ13C offset between wet and dry microsites is also apparent in the annually resolved data covering the 20th and early twenty-first centuries (Fig. 2), though particularly in Kir-M (top panel) the differences between single trees are additionally noticeable. Besides these offsets, the annually resolved isotope data reveal highly coherent co-variability among trees and microsites, a key characteristic of these proxies building the foundation for high-resolution climate reconstruction (Frank et al. 2015; Kress et al. 2010; Treydte et al. 2006). Interseries correlations over 1901–2009 range from 0.48 to 0.69 for single trees and 0.74–0.85 for microsite chronologies (values shown in Fig. 2) substantiating the impact of common (climatic) forcings on inter-annual stable isotope variations. Interestingly, the mean interseries correlation is slightly higher among δ13C and δ18O series from wet microsites, though the difference is not substantial (0.08 across all sites) and not consistent (δ18O in Sto).

Tree-ring δ13C and δ18O data 1901–2009. a δ13C series of four trees in moist conditions (blue) and four trees in dry conditions (red) shown together with their mean chronologies (bottom panel). Dashed curve is the mean of only three trees excluding the data of one tree showing substantially less depleted values (top blue curve in top panel). Values are the inter-series correlations among trees at moist (blue) and dry (red) sites, as well as between the microsite chronologies (black). b–d Same as in a, but for δ13C from Sto (b), δ18O from Kir (c), and δ18O from Sto (d)
To emphasize the high frequency (inter-annual to decadal) patterns in the isotope data, we detrended the δ13C and δ18O series using 30-year splines, and consider both the non-detrended and spl-detrended data for covariance and climate signal assessments. The detrending removes multi-decadal scale variability from the data (Fig. 3), and increases, on average, the covariance among microsites and across proxies (i.e. between δ13C and δ18O) from 0.30 considering all non-detrended chronologies to 0.35 considering all detrended chronologies (Fig. 4). Besides the striking correlation between neighboring microsites, the matrix reveals significant correlations between carbon and oxygen isotopes within the study regions (Kir and Sto). Covariance between δ13C and δ18O is larger in Sto (r = 0.39) compared to Kir (r = 0.28), and particularly strong in Sto when considering only the spl-detrended chronologies (r = 0.48; the orange and yellow boxes in the two bottom rows of Fig. 4). The increased covariance in Sto reveals stomatal conductance, rather than photosynthesis, is the key driver of isotope fractionation in central Sweden. In comparison, the correlation between regions (Kir and Sto) for each proxy is relatively low (r = 0.28 for δ18O and only 0.18 for δ13C) indicating that the data from central and northern Sweden are controlled by regionally differing climatic forcings. The overall higher correlations among the spl-detrended chronologies demonstrate that cross-proxy and cross-microsite variance is most coherent at inter-annual timescale, and supports findings indicating strong climate signals in the high frequency domain across Europe (Treydte et al. 2007).

Effects of detrending tree-ring stable isotope data. a δ18O series of two trees from Sto-D (thin curves) shown together with their 30-spline low pass filters (thick curves). b Detrended δ18O series of the two trees derived from calculating ratios between the original measurements and spline filters shown in a. c Mean δ18O raw (black curve) and detrended (grey curve) chronologies of four trees at Sto-D. d 30-year low pass filters of the detrended and non-detrended chronologies

Correlation coefficients between δ13C and δ18O microsite chronologies from Kiruna and Stockholm using the non-detrended (Raw) and spline-detrended (Spl) data from 1901 to 2009. Considering the varying lag-1 autocorrelations of these data, p < 0.05 is reached at r ≈ 0.22 for the non-detrended and r ≈ 0.19 for the detrended data
Climate signals
The removal of trends from proxy timeseries is also known to potentially affect the climate signals estimated via the calibration against instrumental climate data (Fritts 1976). The modification caused by detrending can be small, such as expressed in a spatially expanded temperature correlation field recorded for Sto δ13C over the late 1951–2009 calibration period (Fig. 5c, d), but might also radically change the putative signal, from insignificant in the vicinity of the proxy location to a noticeable temperature response after detrending (Fig. 5a, b). In this latter example of δ13C from Kir, the recent negative trend in the raw data accidentally resulted in the detection of inverse temperature correlations in southern Europe, reminding of the danger of picking up spurious patterns when using auto-correlated data (Esper et al. 2005; von Storch et al. 2004).

Effects of stable isotope detrending on the calibration against instrumental climate data. a Spatial patterns of the correlation of the non-detrended Kir δ13C site chronology (blue curve) against gridded instrumental JJA temperatures from 1901 to 2009 (top left) and 1951–2009 (top right). b–d Same as in a, but for the detrended Kir δ13C (c), non-detrended Sto δ13C (c), and detrended δ13C chronologies (d). Significant correlation p < 0.10 displayed
Bearing these modifications related to timeseries detrending in mind, we calibrated all δ13C and δ18O microsite chronologies against gridded temperature, precipitation, cloud cover, and PDSI data over the 1901–2009 and 1951–2009 periods using Pearson correlations and illustrate the most significant fields (Figs. 6, 7). In most cases, highest correlations are achieved when averaging the instrumental data over the summer months (JJA), expect for the temperature-versus-δ18O pair, which is showing strongest responses during July in Kir and January–August in Sto (Table 2). Similarly, the spl-detrended stable isotopes more often correlate superior with instrumental fields, i.e. in only 5 of 16 cases the non-detrended data return stronger signals. While not highly significant, this finding indicates that the coherence with climate parameters can increase when removing inter-decadal scale variance from stable isotope records.

Climate signals in δ13C data from locally dry and locally moist sites in Kir and Sto. Correlation fields for the periods 1901–2009 and 1951–2009 (columns) for temperature, precipitation, cloud cover, and PDSI (rows) are shown. For each combination, only the best-correlating season and data (detrended or non-detrended) are illustrated. Significant correlation p < 0.10 displayed. See Table 2 for details

Climate signals in δ18O data from locally moist and locally wet sites in Kir and Sto. Correlation fields for the periods 1901–2009 and 1951–2009 (columns) for temperature, precipitation, cloud cover, and PDSI (rows) are shown. For each combination, only the best-correlating season and data (detrended or non-detrended) are illustrated. Significant correlation p < 0.10 displayed. See Table 2 for details
Even though the strength and spatial extent of correlation differ among the climate parameters tested here, significant fields are present for almost all proxy-climate pairs (Figs. 6, 7). This correlation with multiple instrumental datasets is seemingly affected by the covariance among climate parameters, making it difficult to differentiate between truly controlling and co-varying forcings (Biondi and Waikul 2004; Fritts et al. 1971). Comparisons between PDSI and Kir13M and Kir13D are non-significant when calculated over the long 1901–2009 calibration period, but the significant association evident for the 1951–2009 period (bottom-left in Fig. 6) implies potential deficiencies in the early instrumental network over northern Sweden. In other cases, the correlation fields do not match the tree sites, but are situated northeast (Kir δ13C and cloud cover) and east (Sto δ13C and cloud cover) of the sampling locations. These spatially disconnected patterns might reflect deficiencies in the cloud data network, since remote influences of cloud-controlled irradiance on tree-ring isotope formation are unlikely. An extreme case is the temperature pattern associated with Kir δ18O (top left in Fig. 7), where the correlation field is centered ~ 1000 km south of the tree site. The argument that instrumental data are defective might be somewhat circumstantial here (since temperature readings are typically more robust compared to hydroclimatic and insolation parameters; Auer et al. 2007; Böhm et al. 2001), and the remote correlation field might rather reflect a true signal related to prevailing air mass trajectories and their impact on precipitation δ18O signatures (see next section).
After considering all proxy-instrumental data combinations, it is evident that the highest correlations are recorded for warm season temperatures, except for Sto δ18O correlating best with PDSI (Fig. 8). Importantly, in almost all cases, the stable isotope records from moist microsites correlate better with climate parameters compared to the data from their neighboring dry sites. The few exceptions include temperature signals in Kir18D and Sto18D from 1951 to 2009 and the (overall weak) PDSI signal in Kir13D, which are slightly stronger compared to their moist counterparts. The correlation difference between moist and dry microsites is mostly insignificant and reaches, on average, 0.07 (0.12) for δ13C and 0.05 (0.05) for δ18O over the 1901–2009 (1951–2009) calibration period. This small, but fairly systematic, modification between moist and dry microsites likely originates from differentiating effects of groundwater access on the climatic signal strength of carbon and oxygen isotopes as formulated in hypothesis H3.

Stable isotope climate signals. a Pearson correlation coefficients of δ13C and δ18O microsite chronologies (moist sites in blue, dry sites in red) from Kiruna (dark colors) and Stockholm (light colors) against instrumental temperature, precipitation, cloud cover, and PDSI data (seasons specified in Table 2) considering the best-correlating grid-points shown in Figs. 6 and 7 from 1901 to 2009. Considering varying lag-1 autocorrelations, p < 0.05 is reached at r ≈ 0.19. b Same as in a, but over the 1951–2009 period. p < 0.05 is reached at r ≈ 0.27
These effects appear to be stronger in Sto compared to Kir (particularly δ18O from 1951 to 2009) as well as in δ13C compared to δ18O. Access to lake infiltration water allows for stronger stomatal conductance even during warm and dry periods, a process that not only reinforces atmospheric control of oxygen and carbon fractionation (Darling and Talbot 2003; Leavitt and Long 1989; Saurer et al. 1995), but is also more effective at the warmer Sto site in central Sweden compared to Kir at the Arctic tree line. As a consequence, isotope-based climate reconstructions of warm season temperature (but also precipitation, cloud cover, and PDSI) would explain more variance of the targeted climate variable, if trees from locally moist microsites were considered. When developing composite chronologies integrating isotopic data from living and sub-fossil trees (e.g., Hangartner et al. 2012), it seems therefore important to avoid changes in microsite habitats. Mixing microsites in millennium-scale chronologies could otherwise result in temporal changes in reconstruction skill that seem difficult to be statistically estimated.
Correlation with TRW data
While we acknowledge the difficulty of differentiating between leading and co-varying climate variables controlling stable isotope composition (Frank et al. 2015), comparison with TRW data from the same trees might offer some support to our interpretation of δ13C and δ18O records. Figure 9 shows the correlation of Kir13M and Kir18M with KirTRWM, and the anti-correlation between TRW from the dry microsite (StoTRWD) with Sto13D and Sto13M from 1901 to 2009. The coherent decadal scale variance and significant correlation between stable isotopes and TRW data in the Arctic Kir-M environment (Kir18M/KirTRWM = 0.44, Kir13M/KirTRWM = 0.33; Fig. 9a) is in line with the calibration against warm season temperatures (Büntgen et al. 2011; Esper et al. 2013, 2014; Linderholm et al. 2015). Interestingly, cross-proxy correlation in this cold environment is stronger in the moist microsite as both the lakeshore stable isotope (Fig. 8) and TRW data (Düthorn et al. 2016) contain stronger temperature signals, compared to their inland counterparts.

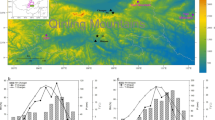
Stable isotope versus tree-ring width comparison. a Kir13M and Kir18M chronologies shown together with the TRW chronology from the same four trees in locally moist conditions (KirTRWM). All data detrended using 30-year spline high-pass filters. Grey and yellow boxes mark periods of reduced (1923–1927) and increased (1933–1937) covariance, respectively. b Same as in a, but for the Sto13M, Sto13D, and StoTRWD chronologies (TRW axis on the right reversed). Grey and yellow boxes highlight 1902–1906 and 1955–1959. C Maps showing the correlation between KirTRWM and July temperature (left), StoTRWD and MJJ precipitation, and StoTRWM and MJJ precipitation over the 1901–2009 period
This situation is different in the overall warmer Sto site, where temperature-controlled δ13C (from moist and dry microsites) correlates negatively with precipitation-controlled TRW from the dry microsite (Fig. 9b, r = − 0.38 for Sto13M, and r = − 0.45 for Sto13D). TRW data from the moist microsite in Sto contains no clear precipitation signal (compare middle and right panels in Fig. 9c), likely because groundwater access degrades the influence of rainfall events on pine cambial activity (Düthorn et al. 2015, 2016). As a consequence, StoTRWM and StoTRWD correlate at only r = 0.42, whereas KirTRWM and KirTRWD, located north of the Arctic Circle, correlate at r = 0.84. Finally, it seems noticeable that lag-1 autocorrelations differ systematically between the proxies displayed in Fig. 9 ranging from < 0.05 for all isotope chronologies to 0.30 in KirTRWM and 0.43 in StoTRWD (all after spl-detrending). High serial correlation is characteristic for TRW chronologies (Matalas 1962; Meko 1981) and limits the ability to properly assess distinct temperature extremes (Esper et al. 2015b). This situation is seemingly different in δ13C and δ18O, which could offer a pathway to improve the reconstruction of cooling events following large volcanic eruptions and other abrupt climate changes (Briffa et al. 1998a; Esper et al. 2013; Schneider et al. 2015).
Temperature and cloud cover signals
Two obvious applications arising from our results would be, first, the reconstruction of large-scale temperature patterns from δ18O, and second, estimates of past changes in insolation and cloud cover. The notable temperature correlation centered over northern Germany, reflected in both Kir13M and Sto13M, is coupled with significant, but inverse correlation fields in the North Atlantic and west Siberia (Fig. 10). These associations are physically meaningful as surface temperatures in these regions (North Atlantic, central Europe, west Siberia) are teleconnected via the westerlies steered by the polar jet stream (Ahrens 2012; Flohn 1950). The prevailing vectors of this planetary wind circulation (shown in grey in Fig. 10) are closely linked with the position and strength of the semi-permanent Icelandic low and Russian high. The combination of δ18O data from Kir-M and Sto-M thereby offers the opportunity to reconstruct changes in the strength and position of these synoptic systems relevant to weather and climate of a large fraction of the Northern Hemisphere (Wanner et al. 2001). We assume that a stronger zonal component of the planetary circulation would be reflected by a weakening of the anti-correlation among the Atlantic, European, Siberian sectors, whereas a stronger meridional component would strengthen these teleconnections. Such temporal changes could be studied using split calibration windows as shown in the bottom panel of Fig. 10a, but clearly more proxy data are needed to explore the potential of reconstructing such atmospheric dynamics with statistical skill (Luterbacher et al. 2002; Trouet et al. 2009, 2016).

Large-scale δ18O correlation patterns. a Correlation fields of the mean oxygen-moist chronology (Kir18M + Sto18M) against gridded instrumental JJA land and sea surface temperatures (SST) from 1901 to 2009 (top panel). Bottom panel shows the fields for 1901–1935, 1936–1972, and 1973–2009. Significant correlations p < 0.10 are displayed. b Mean δ18O-moist chronology (blue curve) scaled to JJA temperatures in Europe (black curve; 53°N, 12°E) together with a scatter plot and linear regression of the data (top panel). Middle and bottom panels show the δ18O-moist data scaled to SSTs from the North Atlantic (58°N, 36°W) and land temperatures from west Siberia (58°N, 60°E)
The insolation signal inherent to stable carbon isotopes from northern Scandinavia has been thoroughly explored by calibrating against local sunshine data recorded in Abisko in northern Sweden (Loader et al. 2013). Loader et al. (2013) show that in northern boreal forests stomatal control of CO2 diffusion into the leafs is less affected by moisture stress, but photosynthetically active radiation is the primary limiting factor controlling carbon isotope fractionation. The extremely high correlation between sunshine hour and cloud cover (typically > 0.90; Barbaro et al. 1981; Ododo et al. 1996; Reddy 1974) is the main justification for merging these different observations of solar insolation into gridded products covering global land areas (New et al. 2002). Our δ13C data from Kir-M and Sto-M revealed that the centers of correlation with cloud cover/sunshine hour data are displaced and shifted towards northeast (Kir13M) and east (Sto13M). Interestingly, the sunshine hour data from the Abisko climate station also correlate with gridded cloud data centered in northwest Finland (Fig. 11a). These remote correlation fields seen in both proxy and instrumental data likely indicate paucities in the gridded cloud cover network (New et al. 2002), limiting efforts to reconstruct long-term insolation variability based on tree-ring stable isotopes. This conclusion is supported by temporal correlation changes signifying skill deterioration in the gridded cloud cover data before 1960 (Fig. 11b). In addition, since both Kir13M and δ13C from Loader et al. (2013) reveal a correlation drop against Abisko sunshine hours centered in the 1950s, the validation of proxy insolation signals based on the calibration against observational data remains challenging. The main reason for this failure is likely the low and temporally varying quality of observational cloud cover and sunshine hour data. This being said, the previously detailed temperature and PDSI signals (Figs. 6, 7, 8) might merely appear superior because thermometer readings contain fewer biases compared to cloud cover, sunshine hour, and precipitation data (Auer et al. 2007).

Stable isotope cloud cover and sunshine hour signals. a Spatial pattern of the correlation between Kir13M and gridded JJA cloud cover data from 1913 to 2006 (top panel). Middle panel shows the correlation pattern for Sto13M, and bottom panel shows the pattern for JJA sunshine hour data recorded at the Abisko climate station from 1913 to 2006. The crosses indicate the grid point used for calculating running correlations shown in b. b 30-year running correlations between Kir13M and JJA cloud cover at 69°N and 28°E (see cross in a, top panel) in dark blue, Sto13M and JJA cloud cover at 61°N and 22°E (cross in a, middle panel) in light blue, and Abisko JJA sunshine hours with JJA cloud cover at 69°N and 28°E (cross in a, bottom panel) in black. Bottom panel shows the 30-year running correlations between Kir13M and the nearby JJA sunshine hour data recorded in Abisko (1913–2006) in dark blue, and between the Loader et al. (2013) δ13C data from Torneträsk and Abisko JJA sunshine hours
Conclusions
The analysis of pine tree-ring δ13C and δ18O data from moist and dry microsites in Scandinavia revealed small but systematic differences in climate signal strength. Stable isotopes from moist microsites correlate better (on average by 0.09 for δ13C and 0.05 for δ18O) against gridded climate data compared to their dry counterparts, a finding that is largely robust for temperature, precipitation, cloud cover, and PDSI. Access to lake filtration water also depleted δ13C by ~ 1‰, but did not affect δ18O. In contrast, both isotope values increased from the warmer sampling site near Stockholm to the colder site near the Arctic tree line located ~ 1000 km north of the Swedish capital (δ13C by 0.8‰ and δ18O by 2.4‰). These large-scale gradients are caused by temperature controlled changes in the isotopic composition of meteoric water (δ18O) and assimilation controlled enzymatic fractionation processes (δ13C).
We found significant correlations between all stable isotope microsite chronologies and all tested climatic parameters. In most microsites, δ13C and δ18O agreed best with warm season temperature. An exception is δ18O from the dry microsite in central Sweden, where needle aperture in trees with limited soil water access more closely regulates CO2, a mechanism that is best reflected in PDSI data (van der Schrier et al. 2006). The overall strong climatic forcing synchronized stable isotope variance among single trees and microsites. Covariance even increased after high-pass filtering the proxy data, reinforcing signal strength of tree-ring δ13C and δ18O chronologies for reconstructing abrupt climatic changes. Covariance is also substantial between δ13C and δ18O chronologies within each region (central and northern Sweden) underpinning that similar climate forcings influence these proxies (Treydte et al. 2007; Saurer et al. 2008).
A conclusive differentiation between forcing and co-varying climate parameters seemed challenging based on only the calibration against instrumental climate data. Covariance of δ13C and δ18O with (temperature controlled) TRW data in northern Sweden, and δ13C with (drought controlled) TRW data in central Sweden, however, supported the climatic interpretation of tree-ring stable isotopes as temperature and PDSI proxies, particularly in the high frequency domain. On the other hand, a physiologically perhaps more meaningful cloud cover reconstruction appeared to be constrained by the mediocrity of twentieth century observational data precluding signal strength assessments particularly in the low frequency domain.
Author contribution statement
The study was conducted in collaboration between all authors: JE and SH designed the study and carried out most calculations. KT, UB, CHM, SSG and DFCR supported the interpretation, and BS and FK contributed to the laboratory work. The discussion and conclusions were a joint work among all authors. The manuscript was written by JE and revised by all other authors.
References
Ahrens CD (2012) Meteorology today: an introduction to weather, climate, and the environment. Brooks/Cole, Belmont
Auer I et al (2007) HISTALP—historical instrumental climatological surface time series of the greater Alpine region 1760–2003. Int J Climatol 27:17–46
Barbaro S, Cannata G, Coppolino S, Leone C, Sinagra E (1981) Correlation between relative sunshine and state of the sky. Sol Energy 26:537–550
Barbour MM (2007) Stable oxygen isotope composition of plant tissue: a review. Funct Plant Biol 34:83–94
Barbour MM, Andrews TJ, Farquhar GD (2001) Correlations between oxygen isotope ratios of wood constituents of Quercus and Pinus samples from around the world. Funct Plant Biol 28:335–348
Bégin C, Gingras M, Savard MM, Marion J, Nicault A, Bégin Y (2015) Assessing tree-ring carbon and oxygen stable isotopes for climate reconstruction in the Canadian northeastern boreal forest. Palaeogeogr Palaeoclim Palaeoecol 423:91–101
Biondi F, Waikul K (2004) Dendroclim2002: a C++ program for statistical calibration of climate signals in tree-ring chronologies. Computers Geosci 30:303–311
Boettger T et al (2007) Wood cellulose preparation methods and mass spectrometric analyses of δ13C, δ18O and non-exchangeable δ2H values in cellulose, sugar, and starch: an inter-laboratory comparison. Anal Chem 79:4603–4612
Böhm R, Auer I, Brunetti M, Maugeri M, Nanni T, Schöner W (2001) Regional temperature variability in the European Alps 1760–1998 from homogenized instrumental time series. Int J Climatol 21:1779–1801
Briffa KR, Jones PD, Schweingruber FH, Osborn TJ (1998a) Influence of volcanic eruptions on Northern Hemisphere summer temperature over the past 600 years. Nature 393:450–455
Briffa KR, Schweingruber FH, Jones PD, Osborn TJ, Shiyatov SG, Vaganov EA (1998b) Reduced sensitivity of recent tree-growth to temperature at high northern latitudes. Nature 391:678–682
Büntgen U et al (2011) Causes and consequences of past and projected Scandinavian summer temperatures, 500–2100 AD. Plos One. https://doi.org/10.1371/journal.pone.0025133
Cernusak LA, English NB (2015) Beyond tree-ring widths: stable isotopes sharpen the focus on climate responses of temperate forest trees. Tree Phys. https://doi.org/10.1093/treephys/tpu115
Cook ER, Peters K (1981) The smoothing spline: a new approach to standardizing forest interior tree-ring width series for dendroclimatic studies. Tree-Ring Bull 41:45–54
Cook ER et al (2015) Old world megadroughts and pluvials during the Common Era. Sci Adv. https://doi.org/10.1126/sciadv.1500561
Cullen LE, Grierson PF (2006) Is cellulose extraction necessary for developing stable carbon and oxygen isotopes chronologies from Callitris glaucophylla? Palaeogeogr Palaeoclim Palaeoecol 236:206–216
Dansgaard W (1964) Stable isotopes in precipitation. Tellus 16:436–468
Darling WG, Talbot JC (2003) The O & H stable isotopic composition of fresh waters in the British Isles. 1. Rainfall. Hydrol Earth Syst Sci 7:163–181
Düthorn E, Holzkämper S, Timonen M, Esper J (2013) Influence of micro-site conditions on tree-ring climate signals and trends in Central and Northern Sweden. Trees 27:1395–1404
Düthorn E, Schneider L, Konter O, Schön P, Timonen M, Esper J (2015) On the hidden significance of differing micro-sites in dendroclimatology. Silva Fenn. https://doi.org/10.14214/sf.1220
Düthorn E, Schneider L, Günther B, Gläser S, Esper J (2016) Ecological and climatological signals in tree-ring width and density chronologies along a latitudinal boreal transect. Scand J For Res 31:750–757
Edwards TWD, Fritz P (1986) Assessing meteoric water composition and relative humidity from 18O and 2H in wood cellulose: paleoclimatic implications for southern Ontario, Canada. Appl Geochem 1:715–723
Ehleringer JR, Dawson TE (1992) Water-uptake by plants—perspectives from stable isotope composition. Plant Cell Environ 15:1073–1082
Esper J, Frank DC, Wilson RJS, Briffa KR (2005) Effect of scaling and regression on reconstructed temperature amplitude for the past millennium. Geophys Res Lett. https://doi.org/10.1029/2004GL021236
Esper J, Frank DC, Büntgen U, Verstege A, Luterbacher J, Xoplaki E (2007) Long-term drought severity variations in Morocco. Geophys Res Lett. https://doi.org/10.1029/2007GL030844
Esper J, Frank DC, Battipaglia G, Büntgen U, Holert C, Treydte K, Siegwolf R, Saurer M (2010) Low-frequency noise in δ13C and δ18O tree ring data: a case study of Pinus uncinata in the Spanish Pyrenees. Glob Biogeochem Cycl. https://doi.org/10.1029/2010GB0037772
Esper J, Schneider L, Krusic PJ, Luterbacher J, Büntgen U, Timonen M, Sirocko F, Zorita E (2013) European summer temperature response to annually dated volcanic eruptions over the past nine centuries. Bull Volcanol. https://doi.org/10.1007/s00445-013-0736-z
Esper J, Düthorn E, Krusic P, Timonen M, Büntgen U (2014) Northern European summer temperature variations over the Common Era from integrated tree-ring density records. J Quat Sci 29:487–494
Esper J, Konter O, Krusic P, Saurer M, Holzkämper S, Büntgen U (2015a) Long-term summer temperature variations in the Pyrenees from detrended stable carbon isotopes. Geochronom 42:53–59
Esper J, Schneider L, Smerdon J, Schöne B, Büntgen U (2015b) Signals and memory in tree-ring width and density data. Dendrochronologia 35:62–70
Esper J et al (2016) Review of tree-ring based temperature reconstructions of the past millennium. Quat Sci Rev 145:134–151
Farquhar G, Roderick M (2007) Worldwide changes in evaporative demand. Water Environ Scripta Varia 108:81–103
Farquhar GD, O’Leary MH, Berry JA (1982) On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Funct Plant Biol 9:121–137
Farquhar GD, Ehleringer R, Hubic KT (1989) Carbon isotope discrimination and photosynthesis. Annu Rev Plant Physiol Plant Mol Bioi 40:503–537
Flohn H (1950) Neue Anschauungen über die allgemeine Zirkulation der Atmosphäre und ihre klimatische Bedeutung. Erdkunde 141–162
Frank D, Büntgen U, Böhm R, Maugeri M, Esper J (2007) Warmer early instrumental measurements versus colder reconstructed temperatures: shooting at a moving target. Quat Sci Rev 26:3298–3310
Frank DC et al (2015) Water use efficiency and transpiration across European forests during the Anthropocene. Nature Clim Change 5:579–583
Friedrichs DA, Büntgen U, Frank DC, Esper J, Neuwirth B, Löffler J (2008) Complex climate controls on 20th century oak growth in Central-West Germany. Tree Phys 29:39–51
Fritts HC (1976) Tree rings and climate. Academic, New York
Fritts HC, Blasing TJ, Hayden BP, Kutzbach JE (1971) Multivariate techniques for specifying tree-growth and climate relationships and for reconstructing anomalies in paleoclimate. J Appl Meteorol 10:845–864
Gagen M, McCarroll D, Edouard JL (2004) Latewood width, maximum density, and stable carbon isotope ratios of pine as climate indicators in a dry subalpine environment, French Alps. Arct Antarct Alp Res 36:166–171
Gagen M, Zorita E, McCarroll D, Young GH, Grudd H, Jalkanen R, Loader NJ, Robertson I, Kirchhefer A (2011) Cloud response to summer temperatures in Fennoscandia over the last thousand years. Geophys Res Lett. https://doi.org/10.1029/2010GL046216
Gessler A, Ferrio JP, Hommel R, Treydte K, Werner RA, Monson RK (2014) Stable isotopes in tree rings: towards a mechanistic understanding of isotope fractionation and mixing processes from the leaves to the wood. Tree Phys 34:796–818
Gori YURI., Wehrens R, Greule M, Keppler F, Ziller L, La Porta N, Camin F (2013) Carbon, hydrogen and oxygen stable isotope ratios of whole wood, cellulose and lignin methoxyl groups of Picea abies as climate proxies. Rap Comm Mass Spectr 27:265–275
Hangartner S, Kress A, Saurer M, Frank D, Leuenberger M (2012) Methods to merge overlapping tree-ring isotope series to generate multi-centennial chronologies. Chem Geolog 294:127–134
Harris I, Jones PD, Osborn TJ, Lister DH (2014) Updated high-resolution grids of monthly climatic observations—the CRU TS3.10 dataset. Int J Climatol 34:623–642
Hartl-Meier C, Zang C, Büntgen U, Esper J, Rothe A, Göttlein A, Dirnböck T, Treydte K (2015) Uniform climate sensitivity in tree-ring stable isotopes across species and sites in a mid-latitude temperate forest. Tree Phys 35:4–15
Heinrich I, Touchan R, Dorado Linan I, Vos H, Helle G (2013) Winter-to-spring temperature dynamics in Turkey derived from tree rings since AD 1125. Clim Dyn 41:1685–1701
Helama S, Arppe L, Timonen M, Mielikäinen K, Oinonen M (2015) Age-related trends in subfossil tree-ring δ13C data. Chem Geol 416:28–35
Hilasvuori E, Berninger F, Sonninen E, Tuomenvirta H, Jungner H (2009) Stability of climate signal in carbon and oxygen isotope records and ring width from Scots pine (Pinus sylvestris L.) in Finland. J Quat Sci 24:469–480
Holmes RL (1983) Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull 43:69–78
Konter O, Holzkämper S, Helle G, Büntgen U, Saurer M, Esper J (2014) Climate sensitivity and parameter coherency in annually resolved δ13C and δ18O from Pinus uncinata tree-ring data in the Spanish Pyrenees. Chem Geol 377:12–19
Kortelainen NM, Karhu JA (2004) Regional and seasonal trends in the oxygen and hydrogen isotope ratios of Finnish ground waters: a key for mean annual precipitation. J Hydrol 285:143–157
Kress A, Saurer M, Büntgen U, Treydte KS, Bugmann H, Siegwolf TW (2009) Summer temperature dependency of larch budmoth outbreaks revealed by Alpine tree-ring isotope chronologies. Oecologia 160:353–365
Kress A, Saurer M, Siegwolf RTW, Frank DC, Esper J, Bugmann H (2010) A 350 year drought reconstruction from Alpine tree-ring stable isotopes. Glob Biogeo Cyc. https://doi.org/10.1029/2009GB003613
Labuhn I, Daux V, Girardclos O, Stievenard M, Pierre M, Masson-Delmotte V (2016) French summer droughts since 1326 CE: a reconstruction based on tree ring cellulose δ18O. Clim Past 12:1101–1117
Leavitt SW (2010) Tree-ring C–H–O isotope variability and sampling. Sci Tot Environ 8:5244–5253
Leavitt SW, Long A (1989) Drought indicated in carbon-13/carbon-12 ratios of southwestern tree rings. Water Resour Bull 25:341–347
Linderholm HW, Björklund J, Seftigen K, Gunnarson BE, Fuentes M (2015) Fennoscandia revisited: a spatially improved tree-ring reconstruction of summer temperatures for the last 900 years. Clim Dyn 45:933–947
Liu Y, Cai Q, Liu W, Yang Y, Sun J, Song H, Li X (2008) Monsoon precipitation variation recorded by tree-ring δ18O in arid Northwest China since AD 1878. Chem Geol 252:56–61
Loader NJ, Robertson I, McCarroll D (2003) Comparison of stable carbon isotope ratios in the whole wood, cellulose and lignin of oak tree-rings. Palaeogeogr Palaeoclim Palaeoecol 196:395–407
Loader NJ, Young GHF, Grudd H, McCarroll D (2013) Stable carbon isotopes from Torneträsk, northern Sweden provide a millennial length reconstruction of summer sunshine and its relationship to Arctic circulation. Quat Sci Rev 62:97–113
Luterbacher J, Xoplaki E, Dietrich D, Rickli R, Jacobeit J, Beck C, Gyalistras D, Schmutz C, Wanner H (2002) Reconstruction of sea level pressure fields over the Eastern North Atlantic and Europe back to 1500. Clim Dyn 18:545–561
Luterbacher J et al (2016) European summer temperatures since Roman times. Environ Res Lett. https://doi.org/10.1088/1748-9326/11/2/024001
Matalas NC (1962) Statistical properties of tree ring data. Hydrol Sci J 7:39–47
Meko DM (1981) Applications of Box-Jenkins methods of time series analysis to the reconstruction of drought from tree rings. PhD Dissertation, Tucson
Mischel M, Esper J, Keppler F, Greule M, Werner W (2015) δ2H, δ13C and δ18O from whole wood, a-cellulose and lignin methoxyl groups in Pinus sylvestris: a multi-parameter approach. Isotop Environ Health Stud 51:553–568
New M, Hulme M, Jones P (1999) Representing twentieth-century space–time climate variability. Part I: development of a 1961–90 mean monthly terrestrial climatology. J Clim 12:829–856
New M, Lister D, Hulme M, Makin I (2002) A high-resolution data set of surface climate over global land areas. Clim Res 21:1–25
Ododo JC, Agbakwuru JA, Ogbu FA (1996) Correlation of solar radiation with cloud cover and relative sunshine duration. Energy Convers Manag 37:1555–1559
Porter TJ, Pisaric MF, Field RD, Kokelj SV, Edwards TW, Healy R, LeGrande AN (2014) Spring–summer temperatures since AD 1780 reconstructed from stable oxygen isotope ratios in white spruce tree-rings from the Mackenzie Delta, northwestern Canada. Clim Dyn 42:771–785
Reddy SJ (1974) An empirical method for estimating sunshine from total cloud amount. Sol Energy 15:281–285
Riechelmann DFC, Maus M, Dindorf W, Schöne B, Scholz D, Esper J (2014) Sensitivity of whole wood stable carbon and oxygen istotope values to milling procedures. Rap Comm Mass Spectr 28:1371–1375
Riechelmann DFC, Maus M, Dindorf W, Konter O, Schöne B, Esper J (2016) Comparison of δ13C and δ18O from cellulose, whole wood, from an old high elevation Pinus uncinata in the Spanish Central Pyrenees. Isotop Environ Health Stud. https://doi.org/10.1080/10256016.2016.1161622
Rinn F (2005) TSAP Win—time series analysis and presentation for dendrochronology and related applications. User reference, Heidelberg
Rinne KT, Loader NJ, Switsur VR, Waterhouse JS (2013) 400-year May–August precipitation reconstruction for Southern England using oxygen isotopes in tree rings. Quat Sci Rev 60:13–25
Roden JS, Ehleringer JR (1999) Observations of hydrogen and oxygen isotopes in leaf water confirm the Craig-Gordon model under wide-ranging environmental conditions. Plant Physiol 120:1165–1173
Rozanski K, Araguas-Araguas L, Gonfiantini R (1992) Relation between long-term trends of oxygen-18 isotope composition of precipitation and climate. Science 258:981–985
Saurer M, Siegenthaler U, Schweingruber F (1995) The climate-carbon isotope relationship in tree-rings and the significance of site conditions. Tellus B 47:320–330
Saurer M, Siegwolf R, Schweingruber F (2004) Carbon isotope discrimination indicates improving water-use efficiency of trees in northern Eurasia over the last 100 years. Glob Change Biol 10:2109–2120
Saurer M, Cherubini P, Reynolds-Henne CE, Treydte KS, Anderson WT, Siegwolf RTW (2008) An investigation of the common signal in tree ring stable isotope chronologies at temperate sites. J Geophys Res. https://doi.org/10.1029/2008JG000689
Schleser GH, Anhuf D, Helle G, Vos H (2015) A remarkable relationship of the stable carbon isotopic compositions of wood and cellulose in tree-rings of the tropical species Cariniana micrantha (Ducke) from Brazil. Chem Geol 401:59–66
Schneider L, Smerdon J, Büntgen U, Wilson R, Myglan VS, Kirdyanov A, Esper J (2015) Revising midlatitude summer temperatures back to AD 600 based on a wood density network. Geophys Res Lett. https://doi.org/10.1002/2015GL063956
Seftigen K, Linderholm HW, Loader NJ, Liu Y, Young GH (2011) The influence of climate on 13C/12C and 18O/16O ratios in tree ring cellulose of Pinus sylvestris L. growing in the central Scandinavian Mountains. Chem Geol 286:84–93
Sidorova OV, Siegwolf RT, Saurer M, Naurzbaev MM, Vaganov EA (2008) Isotopic composition (δ13C, δ18O) in wood and cellulose of Siberian larch trees for early Medieval and recent periods. J Geophys Res Biogeosci. https://doi.org/10.1029/2007JG000473
Stuiver M, Braziunas TF (1987) Tree cellulose 13C/12C isotope ratios and climatic change. Nature 328:58–60
Tejedor E, Saz MA, Cuadrat JM, Esper J, de Luis M (2017) Summer drought reconstruction in Northeastern Spain inferred from a tree-ring latewood network since 1734. Geophys Res Lett. https://doi.org/10.1002/2017GL074748
Treydte K, Schleser GH, Schweingruber FH, Winiger M (2001) The climatic significance of δ13C in subalpine spruces (Lötschental/Swiss Alps)—a case study with respect to altitude, exposure and soil moisture. Tellus B 53:593–611
Treydte K, Schleser GH, Helle G, Frank DC, Winiger M, Haug GH, Esper J (2006) The twentieth century was the wettest period in Northern Pakistan over the past millennium. Nature 440:1179–1182
Treydte K et al (2007) Signal strength and climate calibration of a European tree ring isotope network. Geophys Res Lett. https://doi.org/10.1029/2007GL031106
Treydte K, Frank DC, Saurer M, Helle G, Schleser G, Esper J (2009) Impact of climate and CO2 on a millennium-long tree-ring carbon isotope record. Geochim Cosmochim Act 73:4635–4647
Treydte K, Boda S, Graf Pannatier E, Fonti P, Frank D, Ullrich B, Saurer M, Siegwolf R, Battipaglia G, Werner W, Gessler A (2014) Seasonal transfer of oxygen isotopes from precipitation and soil to the tree ring: source water versus needle water enrichment. New Phytol 202:772–783
Trouet V, van Oldenborgh GJ (2013) KNMI climate explorer: a web-based research tool for high-resolution paleoclimatology. Tree Ring Res 69:1–13
Trouet V, Esper J, Graham NE, Baker A, Scourse JD, Frank DC (2009) Persistent positive North Atlantic oscillation mode dominated the Medieval Climate Anomaly. Science 324:78–80
Trouet V, Harley GL, Domínguez-Delmás M (2016) Shipwreck rates reveal Caribbean tropical cyclone response to past radiative forcing. Proc Nat Acad Sci 13:3169–3174
van Oldenborgh GJ, Burgers G (2005) Searching for decadal variations in ENSO precipitation teleconnections. Geophys Res Lett. https://doi.org/10.1029/2005GL023110
van der Schrier G, Briffa KR, Jones PD, Osborn TJ (2006) Summer moisture variability across Europe. J Clim 19:2818–2834
Verheyden A, Roggeman M, Bouillon S, Elskens M, Beeckman H, Koedam N (2005) Comparison between δ13C of α-cellulose and bulk wood in the mangrove tree Rhizophora mucronata: implications for dendrochemistry. Chem Geol 219:275–282
von Storch H, Zorita E, Jones J, Dimitriev Y, Gonzalez-Rouco JF, Tett SFB (2004) Reconstructing past climate from noise data. Science 306:679–682
Wanner H, Brönnimann S, Casty C, Gyalistras D, Luterbacher J, Schmutz C, Stephenson DB, Xoplaki E (2001) North Atlantic oscillation—concepts and studies. Surveys Geophys 22:321–381
Waterhouse JS, Barker AC, Carter AHC, Agafonov LI, Loader NJ (2000) Stable carbon isotopes in Scots pine tree rings preserve a record of flow of the river Ob. Geophys Res Lett 27:3529–3532
Weigt RB et al (2015) Comparison of δ13C and δ18O values between tree-ring whole wood and cellulose in five species growing under two different site conditions. Rapid Commun Mass Spectr 29:2233–2244
Wilson AT, Grinsted MJ (1977) 12C/13C in cellulose and lignin as palaeothermometers. Nature 265:133
Wilson RJS et al (2016) Last millennium Northern Hemisphere summer temperatures from tree rings. Part I: the long term context. Quat Sci Rev 134:1–18
Zeng X, Liu X, Treydte K, Evans MN, Wang W, An W, Sun W, Xu G, Wu G, Zhang X (2017) Climate signals in tree-ring δ18O and δ13C from southeastern Tibet: insights from observations and forward modelling of intra- to interdecadal variability. New Phytol. https://doi.org/10.1111/nph.14750
Acknowledgements
We thank Florian Benninghoff, Willy Dindorf, Elisabeth Düthorn, Susanne Koch, Markus Kochbeck, Oliver Konter, Michael Maus, Maria Mischel and Jutta Sonnberg for field and laboratory support.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
All authors declare no conflict of interest.
Additional information
Communicated by A. Gessler.
Rights and permissions
About this article

Cite this article
Esper, J., Holzkämper, S., Büntgen, U. et al. Site-specific climatic signals in stable isotope records from Swedish pine forests. Trees 32, 855–869 (2018). https://doi.org/10.1007/s00468-018-1678-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-018-1678-z