
Abstract
Key message
In the center of the species’ distribution range in Germany, European beech stands show continued climate-change-related growth decline since the 1980s at low elevations, but growth increase at high elevations.
Abstract
Contradicting reports exist about the climate-change sensitivity of European beech (Fagus sylvatica), showing either a sensitive response of radial growth to dry and hot summer episodes and long-term growth decline with recent warming, or apparent insensitivity of growth and a remarkable potential for post-stress recovery. With a dendroecological study along an altitudinal transect in the center of the species’ distribution range in south-western Germany, we analyzed the climate response of radial growth with variation in altitude (110–1230 m) and associated temperature (10.6–3.5 °C MAT) and precipitation change (755–1788 mm year−1 MAP). Climate sensitivity analysis showed that annual stem increment was strongly limited at low elevations (110–300 m) by low precipitation in April/May, but by low summer temperatures at 1230 m. At intermediate elevation (640 m), indications of both moisture and temperature limitation were found. The differences in climate sensitivity were linked to contrasting long-term growth trends. At 110–300 m, radial growth has continually decreased since about the 1980s, while it has increased at 1230 m. Our results from the four stands suggest for the study region that the abiotic control of beech radial growth switches from moisture to temperature limitation at a threshold situated between 160 and 235 mm of precipitation in April/May (which corresponds to 200 and 313 mm of precipitation in June–August), in accordance with dendroecological results from other Central European lowland regions. This indicates that, with further warming and drying of the climate, beech may suffer in lowland and lower montane regions of Central Europe from reduced vitality and productivity, whereas it may profit from warming in montane to upper montane elevation. We conclude that climate-change-related growth decline is more widespread in the center of the species’ distribution range than previously thought, which is highly relevant for forestry planning.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
European beech (Fagus sylvatica L.) is the most widespread and abundant tree species of the natural forest vegetation of Central Europe and one of the economically most important timber trees of this region (Ellenberg and Leuschner 2010). The species’ distribution range lies in the more oceanic parts of Europe, and numerous dendrochronological and ecophysiological studies suggest that European beech is more drought sensitive than other Central European broad-leaved tree species, such as the oaks (Quercus petraea (Matt.) Liebl. and Q. robur L.), hornbeam (Carpinus betulus L.), ash (Fraxinus excelsior L.), linden (Tilia cordata L.), or Norway spruce (Picea abies (H.) Karst.) (Köcher et al. 2009; Zang et al. 2014; Zimmermann et al. 2015). Since global warming has already resulted in higher growing season temperatures and, in various regions of Central Europe, reduced summer precipitation during the last decades (Schönwiese and Janoschitz 2008; IPCC 2013; Lindner et al. 2014), and ongoing climate change is predicted to further increase the severity and frequency of hot and dry summers (Bindoff et al. 2013), the tolerance of European beech to summer drought and heat has raised some concern about its future in Central European forests (e.g., Gessler et al. 2007; Hacket-Pain et al. 2016).
European beech has been found to respond to soil water shortage with reductions of stomatal conductance, sap flow rate, drops in minimum and maximum leaf water potentials, and increased fine root mortality that are more severe than in the other widespread European broad-leaved species (Leuschner et al. 2001; Köcher et al. 2009). In Central and Western European broad-leaf mixed forests with beech, oak, hornbeam, maple, or linden, climate-change-related reductions in stem increment were more pronounced in beech than in other co-occurring tree species (Cavin et al. 2013; Zimmermann et al. 2015). Climate–growth analysis in beech forests of Western, Central, and South-eastern Europe revealed a consistent positive response to high precipitation in the growing season (mainly May to July) and a negative effect of high temperatures in the growing season and the previous summer (Jump et al. 2006; Scharnweber et al. 2011; Zimmermann et al. 2015; Hacket-Pain et al. 2016). In addition, direct measurement of the diurnal variation in radial stem increment indicated that the stem cambial activity of beech is highest in periods of high atmospheric humidity (Köcher et al. 2012), suggesting that increases in the atmospheric vapor pressure saturation deficit (VPD) with increasing summer temperatures may negatively influence radial growth rate.
Recent observations of long-term growth decline in beech forests at the southern edge of beech distribution have been related to drought effects associated with climate change (Piovesan et al. 2003, 2008; Jump et al. 2006). Drought-induced stand-level tree mortality has recently been observed in Hungary at the south-eastern distribution limit of Fagus in Europe (Lakatos and Molnár 2009). In certain regions of Central Europe, growth declines have been reported for beech stands since about the 1980s, e.g., in north-eastern France and Belgium (Charru et al. 2010; Kint et al. 2012), and central-eastern and north-eastern Germany (Scharnweber et al. 2011; Zimmermann et al. 2015). However, these reports contrast with several other studies which found no evidence of recent growth decline in beech in the context of climate change, or even documented recent growth increase (e.g., Friedrichs et al. 2009, Tegel et al. 2014). Various authors emphasize the remarkable potential for recovery after drought stress of Fagus (e.g., Elling et al. 2007).
In Central European beech forests, it appears that coarse wood production has generally increased since the late 19th century (Pretzsch et al. 2014), and Ammer et al. (2005) and Mette et al. (2013) assume that beech is not threatened by climate change in Central Europe in the next decades. In contrast, Cavin and Jump (2016) found in 46 Western European beech stands (UK to Spain) a higher drought sensitivity of radial growth at more northerly sites than close the southern range edge. It thus remains unclear whether long-term growth decline (or drought-induced mortality) is a local phenomenon in Central European beech forests or is more widespread in the center of the species’ distribution range.
In the Mediterranean regions of Spain and Italy, recent decreases in growth rate were most pronounced at lower elevations, but were visible also in montane forests at elevations of 1000–1800 m a.s.l. (Piovesan et al. 2003, 2008; Jump et al. 2006). For Central Europe, there is consistent evidence that beech radial growth at high elevations is predominantly controlled by temperature, while low-elevation productivity is mostly driven by summer rainfall and negatively affected by high summer temperatures (e.g., di Filippo et al. 2007; Babst et al. 2013). However, it is less clear whether these different modes of growth control already have translated in Central Europe into contrasting long-term growth trends at low and high elevations, i.e., widespread growth decline at lower elevation and increase at higher elevation driven by the recent warming. If such a pattern does emerge, the search for climatic thresholds of diverging beech growth responses would be of high importance to support foresters with sound information on future climate-change-related risks in beech production forests (Czucz et al. 2011).
To address this topic, we conducted a dendrochronological case study in south-western Germany in a region, where European beech occurs from lowland to montane elevation, extending over more than 1000 altitudinal meters. Four beech forests were selected along an altitudinal transect from 110 to 1230 m a.s.l., covering an air temperature gradient from 10.6 to 3.5 °C mean annual temperature (MAT) and a precipitation gradient (mean annual precipitation, MAP) from 755 to 1788 mm year−1. We tested the hypothesis that the recorded warming in the second half of the 20th century has resulted in a reduction of beech radial growth at lower elevations, but increased stem increment at higher elevations. A second goal was to search for the climatic tipping point at which beech radial increment is shifting from apparent drought limitation to low-temperature limitation in this region.
Materials and methods
Study area
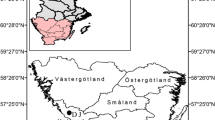
Field work was carried out in November and December 2013 in the federal state of Baden-Württemberg near Karlsruhe (49°4′8″N, 8°26′47″E, 110 m a.s.l.), Weinheim (49°28′45″N, 8°42′50″E, 300 m a.s.l.), Hausach (48°19′24″N, 8°8′36″E, 640 m a.s.l.), and Schönau (47°50′6″N, 7°50′43″E, 1230 m a.s.l.) the sites are labeled as 110 K, 300 W, 640H, and 1230S, respectively, throughout the paper (Fig. 1). Karlsruhe is situated in the middle Upper Rhine Plain, as is Weinheim at the western edge of the Odenwald Mountains east of the northern Upper Rhine Plain. Hausach and Schönau are located in the middle and southern Black Forest, respectively, a major mountain massif (highest summit Mt. Feldberg 1493 m a.s.l.) that emerges east of the southern Upper Rhine Plain. Both the Black Forest and the Odenwald Mountains are dominated by crystalline basement rocks, including granite and gneiss in the Black Forest, and granite and granodiorite in the Odenwald Mountains. The Upper Rhine Plain is dominated by quaternary alluvial deposits.

Study area in south-western Germany with the location of the sample plots (110 K, Karlsruhe; 300 W, Weinheim; 640H, Hausach; 1230S, Schönau)
Climate
Climate data were obtained from the weather stations Karlsruhe (49°2′17″N, 8°21′51″E; 110 m a.s.l.) for site 110 K, Heidelberg (49°25′14″N, 8°40′3″E; 110 m a.s.l.) for site 300 W, Wolfach (48°17′43″N, 8°14′21″E; 290 m a.s.l.) for site 640H, and the summit of Mt. Feldberg (47°52′30″N, 8°0′14″E; 1490 m a.s.l.) for site 1230S (Table 1). These meteorological stations were selected because of their proximity to the sample plots (Fig. 1), but deviated (except for Karlsruhe) markedly with respect to elevation from the plots. Using the data from Heidelberg for site 300 W and from Wolfach for site 640H likely results in higher temperatures and lower precipitation than found at the site of sampling. The deviation must have been largest at site 640H, where mean annual temperature was probably 2.1 K lower than in Wolfach, assuming a temperature lapse rate of 0.6 K 100 m−1. Site 300 W can be assumed to be c. 1.1 K colder than at the station Heidelberg. Concerning site 1230S, use of the data from the Mt. Feldberg station led probably to an underestimation of MAT by 1.6 K as well as to an overestimation of mean annual precipitation. The main purpose of inspecting the climate data was to search for long-term climate trends at the different elevations and to analyze climate–growth relationships. Weather data (temperature/precipitation) were available for the station Karlsruhe since 1851/1876, from Heidelberg since 1871/1871, from Wolfach since 1957/1936, and from Mt. Feldberg since 1921/1926. MAT showed the adiabatic decrease with elevation at the meteorological stations, whereas MAP increased. At lowland elevation (110 m a.s.l.), precipitation was higher in Karlsruhe than in Heidelberg. Seasonal variability of precipitation is relatively small with maxima in summer at the three lower stations and in summer and autumn at the highest elevation.
The standardized precipitation evapotranspiration index (SPEI) was calculated with R 2.15 (R Development Core Team 2008) using the R package “SPEI” v. 1.6 to quantify drought intensity (Vincente-Serrano et al. 2009). Measured values of monthly precipitation and calculated values of potential evapotranspiration (PET) were included in the calculation. PET was computed after Thornthwaite (1948) using monthly mean temperature and a correlation coefficient derived from latitude to calculate day length. SPEI was calculated on a monthly timescale using indices with a log-logistic probability distribution and parameter fitting based on unbiased probability-weighted moments. This time scale was used for identifying drought episodes in the last decades, which could affect drought-sensitive tree species on shallow soils.
Field and laboratory work for tree-ring analysis
The four stands are European beech forests used for timber production. Existing permanent plots of 20 m × 20 m size (site 110 K) or 20 m × 24 m size (other sites) were used to collect tree-ring samples at the four study sites. Beech was the only tree species in stand 300 W and dominated the canopy of the other stands (Table 2). In stand 110 K, beech was associated with six species of native and introduced broad-leaved trees (Prunus padus, Robinia pseudacacia, Acer pseudoplatanus, Carpinus betulus, Quercus petraea, and Q. rubra). In stand 640H, Abies alba, Quercus petraea, and Acer pseudoplantanus co-occurred with beech. In the high-montane forest at site 1230H, beech was mixed with Picea abies and Abies alba. While P. abies formed roughly 30% of the upper canopy in stand 1230H, where it was represented with c. 110- to 120-year-old individuals of large stem diameter, the majority of other admixed species were mostly found in few individuals that were usually smaller than beech and often in a subdominant or suppressed social position. Beech showed no consistent trend for a decrease (or increase) in stem diameter, tree height, stem density, or stand basal area with elevation (Table 2). While stand density was lower in the stands 640H and 1230S than at lower elevation, stand basal area was more similar across the transect with the highest value observed in stand 640H. Tree height was determined with a Vertex IV ultrasonic clinometer and T3 transponder (Haglöf, Långsele, Sweden). Stem diameter was calculated from stem circumference that was measured with a diameter tape.
Fifteen dominant or co-dominant beech trees reaching the upper canopy were randomly selected from each sample plot. Wood cores were collected at breast height (dbh; 1.3 m) using an increment borer of 5 mm inner diameter. The borer was driven into the wood parallel to the contour lines of the mountain slope to avoid compression wood. In the level stand 110 K in the Upper Rhine Plain, samples were taken in north–south direction. Wood cores were mounted on wooden stripes with glue, cut lengthwise, and the cut surface was polished before variation in wood density was made visible with chalk. Annual ring widths were measured with a precision of 10 µm on a movable object table (Lintab 6, Rinntech, Heidelberg, Germany), the movements of which are electronically transmitted to a computer system equipped with the TSAP (Time Series Analysis and Presentation)-Win software (Rinntech).
Analysis of tree-ring data
Most of the calculations for the evaluation of tree-ring data were conducted with the TSAP-Win software. Tree-ring series were controlled for missing rings and false rings during cross-dating. Cross-dating was based on the use of two parameters: coefficient of agreement (‘Gleichläufigkeit’ [GL]) (Eckstein and Bauch 1969) and (standard) t values. The GL and t values measure the similarity between tree-ring series in the high- and low-frequency domains, respectively. Tree-ring series used for the calculation of means had GL values >65% (P ≤ 0.05) and t values >3.5. Tree-ring series were smoothed using 5-year moving averages. Standardized tree-ring series were used in correlation analyses with climate parameters. Standardization is principally achieved by dividing the observed tree-ring width (w i ) through the expected annual increment. The annual tree-ring index (z i ) of year i was calculated using the equation z i = w i /m i with m i being the 5-year or 10-year moving averages of year i. This procedure was applied, since we aimed at keeping as much of the inter-annual (high-frequency) variation as possible, because this is crucial for any climate-response analysis.
Age is generally specified as the age of the oldest tree ring (‘cambial age’) at the sampling height of 1.3 m; c. 10 (at most 20) years should be added to deduce tree age from these age specifications. Tree-ring series were representative of the studied stand as shown by the calculation of the expressed population signal (EPS) using sums of squares of within-core variation and error sums of squares calculated in a two-way analysis of variance (ANOVA) (Wigley et al. 1984). The EPS calculated separately for the age groups and habitats (forest interior vs. edge) clearly exceeded the 0.85 threshold both in middle-aged and old trees. The calculation of EPS values was computed with the package “Dendrochronology Program Library” in R (dplR) 1.5.5 (Bunn 2008) in R 2.15.
Year-to-year variability of tree-ring width was expressed as mean sensitivity, as was the influence of the increment in the previous year on the increment of the current year as first-order autocorrelation coefficient (Fritts 1976). Mean sensitivity and autocorrelation coefficients were calculated over the whole lifespan of each sample tree using the TSAP software. Relationships of tree-ring width with monthly climate data were analyzed with multiple regression analyses using the SAS 9.4 software (SAS Institute Inc., Cary, North Carolina, USA). The climate data were related to standardized tree-ring chronologies. Monthly and seasonal mean temperatures and precipitation of the current and the preceding year of tree-ring formation were included in the climate-response analysis. The regression analyses covered different periods, depending on the availability of climate data for the different localities (see above). Standardized β coefficients were used to quantify the influence of individual months in the climate-response analysis.
The resilience of annual stem increment to individual summer drought events was analyzed by dividing the non-standardized mean tree-ring width in the 9 years after the drought year by the mean tree-ring width in the 9 years prior to the drought year (Lloret et al. 2011). Depending on the intensity and the temporal course of the drought episode, drought can reduce the stem increment in trees either in the year of the drought event or in the subsequent year. We selected growth depressions in 1976, 1989, 1992, 2000, and 2004 for our analysis of the resilience of wood formation to drought. The growth depressions in 1976 and 1989 were due to hot and dry summers in the same year; the drought in 1976 was more pronounced than in 1989. Low growth in 1992 followed a hot and dry summer in 1991, while 1992 was hot but less dry. The year 2000 was hot already early in the growing season, but precipitation was moderate. The growth depression in 2004 was the result of the excessive summer drought of 2003, which occurred in mid-summer at a point in time when most of the annual wood production of 2003 was already completed. The interval of 9 years was chosen, because it was the longest possible interval after 2004, given the wood core sampling in 2013.
Superposed epoch analysis (SEA) was calculated using dplR (Bunn 2008) to test for the mean growth response to the driest summers in the observation period. The SEA calculates the significance of departure of the tree-ring index from the mean for event years and the years immediately preceding and following the event year (superposed epoch), by comparing the value of the superposed epoch to randomly selected epochs, which are selected from the data set using bootstrap resampling (1000 resamples). Event years were deduced from minima of the SPEI value for the period from May to August separately for each study site (Table S1).
Statistical analysis
Arithmetic means ± standard errors are given throughout the paper. Multiple comparisons of mean values were made using Duncan’s multiple range test (significance level: P < 0.05), after testing for normal distribution with the Shapiro–Wilk test; degrees of freedom (df) are given as df model, df error. The statistical analyses were conducted with the SAS 9.4 software.
Results
Climate trends
All four weather stations showed significant MAT increases during the periods, where data were available. Since 1960, temperature increased by 1.95 K (or 0.40 K decade−1) in Karlsruhe, 0.75 K (or 0.14 K decade−1) in Heidelberg, 1.31 K (or 0.24 K decade−1) in Wolfach, and 1.67 K (or 0.31 K decade−1) on Mt. Feldberg (Fig. S1). Temperature increased primarily in spring and summer (Fig. S2), whereas values remained more or less constant in the cold seasons (data not shown), except for Heidelberg, where significant increases were found in both autumn (y = 0.01x + 5.97, r = 0.34, P ≤ 0.001) and winter (y = 0.01x + 3.30, r = 0.20, P ≤ 0.05). Spring (March–May) warming has strongly accelerated since the 1980s with rates varying between 0.51 and 1.04 K for a 34-year period (Table S2; Fig. S2). Summer temperature has increased at lower rates of 0.21–0.40 K since the 1970s (Table S2; Fig. S2).
Annual precipitation decreased across the entire measuring periods in Karlsruhe and on Mt. Feldberg, but remained constant at the two other stations (Fig. S3). Physiologically more important is the spring and summer rainfall decrease, which is visible at all four stations (Fig. S4). In absolute numbers, June–August precipitation was by 36, 29, 23, and 35 mm lower on average in Karlsruhe, Heidelberg, Wolfach, and Mt. Feldberg in the period 1970–2013 than in the 65- to 135-year-long measuring periods since installation of the stations. July rainfall has decreased by 21, 19, 16, and 19 mm, respectively, i.e., by about 27, 23, 14, and 12% at the four stations. Summer drought intensity, as quantified by the mean standardized precipitation/evapotranspiration index (SPEI), increased in all four localities. This was especially true for spring (April to May) and summer (June to August) (Fig. 2). Significant linear trends for SPEI increase were found in both low-elevation stations (Karlsruhe, Heidelberg) in spring and summer. In Wolfach, significant increases in SPEI were found for May and July. On Mt. Feldberg, SPEI increased in May and in the summer months (June to August).

Mean standardized precipitation/evapotranspiration index (SPEI) for the study sites a 110 K (April–May; June–August), b 300 W (April–May; June–August), c 640H (May; July), and d 1230S (May; June–August). Negative values indicate water deficits. Significant trend lines are indicated. Dark gray stands for April–May and light gray for June–August
Annual stem increment
Mean cambial tree age of the sampled dominant and co-dominant beech trees varied between 102 and 176 years. There was no significant correlation of mean tree age with altitude, but the sample trees in the two high-elevation stands were older (640H: 176 ± 7 years, 1230S: 157 ± 3 years) than in the two lowland stands (110 K: 103 ± 2 years, 300 W: 114 ± 2 years). All means from the four sites differed significantly from one another (P ≤ 0.05, Duncan’s multiple range test, df model, error = 3, 56). Maximum age was also higher at high (640H: 235 years, 1230S: 189 years) than low elevation (110 K: 125 years, 300 W: 139 years).
Mean annual radial stem increment was twice as high in the two low-elevation stands than at high elevation (Fig. 3a; Fig. S5). First-order autocorrelation of tree-ring width was lower (Fig. 3b) and mean sensitivity higher at the highest site 1230S (Fig. 3c) than in the other stands. From 110 to 640 m a.s.l., first-order autocorrelation and mean sensitivity were not correlated with altitude. At low elevation (sites 110 K and 300 W), mean sensitivity was significantly higher after 2000 than in the intervals from 1980 to 1999 and from 1960 to 1979 (Table S3). At site 640H, there was an insignificant trend for recently increased mean sensitivity, whereas no such trend existed at the high-elevation site 1230S.

a Mean annual stem increment, b mean sensitivity, and c first-order autocorrelation coefficients in the four study sites. Means sharing the same letter do not differ significantly (P ≤ 0.05, Duncan’s multiple range test, df model, error = 3, 56). Site 110 K (Karlsruhe), 300 W (Weinheim), 640H (Hausach), 1230S (Schönau)
Stem increment showed high synchronicity between the two low-elevation sites 110 K and 300 W (Figs. 4a, b, 5a, b). This assessment is supported by a high coefficient of agreement between the two chronologies of 73% and a high t value of 10.3. The beech trees at the low-elevation sites 110 K and 300 W showed a strong decrease of tree-ring width since the 1980s (Fig. 4a, b; Fig. S5). At site 640H, tree-ring width exhibited a similar but weaker trend for decrease (Fig. 4c), whereas at the high-montane site 1230S, annual stem increment has strongly increased since the 1980s (Fig. 4d).

Mean tree-ring width and number of sample trees at the sites a 110 K (EPS ≥ 0.92 since 1909), b 300 W (EPS ≥ 0.86 since 1909, ≥0.91 since 1924), c 640H (EPS ≥ 0.92 since 1849), and d 1230S (EPS ≥ 0.95 since 1849). Note different scale of y-axes

Mean tree-ring index at the sites a 110 K, b 300 W, c 640H, and d 1230S
Climate-response analysis
The results of the climate-response analysis clearly differ between the low-elevation sites 110 K and 300 W, on the one hand, and the montane to upper montane sites 640H and 1230S, on the other hand. The greatest contrasts exist between the uppermost stand 1230S and the low-elevation sites 110 K and 300 W, while stand 640H occupies an intermediate position not only in terms of elevation, but also with respect to the effect of inter-annual climate variability on annual stem increment.
At the sites 110 K and 300 W (Fig. 6a, b), the tree-ring index is negatively correlated with the summer temperature of the year prior to tree-ring formation and positively correlated with the autumn temperature of the previous year and with precipitation at the start of the current growing season (110 K: April and May; 300 W: May). At site 110 K, the tree-ring index also increases with summer precipitation of the previous year.

Response of the tree-ring index in beech trees from sites a 110 K and b 300 W to monthly and seasonal temperatures and precipitation of the year of, and the year prior to, tree-ring formation. Beta coefficients resulting from multiple regression analysis (*P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001) (n = 15 trees per site)
The climate response of annual stem increment at site 1230S is characterized by a positive correlation of the tree-ring index with summer temperature in the year of tree-ring formation (Fig. 7b), which is in contrast to the results from sites 110 K and 300 W. Nonetheless, the tree-ring index also decreased with the previous year’s June temperature and increased with the previous year’s June precipitation. A marginally significant positive correlation (P < 0.10) was found between the tree-ring index and the previous year’s autumn temperature. Furthermore, correlations with the weather conditions in the winter preceding the current growing season were found; the tree-ring index decreased with January precipitation (P ≤ 0.05) and January and February temperature (P ≤ 0.10).

Response of the tree-ring index in beech trees from sites a 640H and b 1230S to monthly and seasonal temperatures and precipitation of the year of, and the year prior to, tree-ring formation. Beta coefficients resulting from multiple regression analysis (*P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001)
Trees at site 640H (Fig. 7a) showed a positive correlation of the tree-ring index with June temperature of the current year, but lacked the correlation with the temperature of the whole current summer that was found at site 1230S. In addition, annual stem increment at site 640H decreased with the previous year’s summer temperature and increased with the previous year’s May precipitation. Furthermore, the temperature of the February prior to the current-year’s growing season was negatively correlated with the tree-ring index.
Resilience of annual stem increment to individual drought events
In the years following the growth depression years 1989, 1992, 2000, and 2004, the annual stem increment of beech did not fully recover at the sites 110 K, 300 W, and 640H (Fig. S6). The resilience at these sites was <1 in the 9 years after the drought event, reflecting lower stem growth after than prior to the drought event. At the high-elevation site 1230S, the resilience of wood formation to drought was significantly higher than at the other sites and exceeded 1. In 1976, growth resilience was not dependent on elevation.
Superposed epoch analysis (SEA) showed a stronger growth reduction in the year after the drought than in the drought year itself (Fig. 8). This was true for the low-elevation sites 110 K and 300 W as well as for the upland site 640H. At the uppermost site 1230S, however, SEA gave no significant result.

Superposed epoch analysis of the most severe drought years (see Table S1) in the study sites a 110 K, b 300 W, c 640H, and d 1230S. Asterisks indicate significance: (*) P ≤ 0.10, *P ≤ 0.05, **P ≤ 0.01
Discussion
Causes of growth decline at lower elevations
Climate warming in the study region in south-western Germany markedly exceeded the global average of 0.12 K from 1951 to 2012 (IPCC 2013). Only at 300 m elevation in Heidelberg (near site 300 W), the temperature increase was in the range of the global mean increase, but it has to be noted that slightly different periods (since 1960 in our study, since 1951 in IPCC 2013) are compared. The increase in temperature was most pronounced in spring and summer, i.e., the seasons with the highest physiological activity and thus biological relevance. Especially spring warming has accelerated since the 1980s. Compared to earlier periods, summer rainfall (June–August) has decreased at all stations in the last four decades, while spring rainfall (April, May) showed only minor long-term change, as did annual precipitation.
In the two lowest sites at 110 and 300 m a.s.l. in or near the Upper Rhine Plain, the stem increment of beech was significantly promoted when rainfall was higher in April and May. The apparent dependence of growth on sufficient moisture in spring and early summer in these sites coincides with a marked decline of tree-ring width in the two stands since the 1970s and 1980s. This decline is probably due to decreased spring and summer precipitation (Fig. S4) and increased spring and summer temperatures (Fig. S2). Temperature can be effective at reducing stem increment via various pathways, through its influence on the atmospheric evaporative demand (Kint et al. 2012; Köcher et al. 2012), a direct negative effect of higher temperatures on photosynthetic activity and the carbon balance (Filewod 2011), or the assumed promotion of masting frequency in beech by elevated summer temperatures (Drobyshev et al. 2010; Müller-Hauboldt et al. 2015; Hacket-Pain et al. 2016; see below).
Our observation of long-term growth decline of beech at the two sites at 110–300 m a.s.l. is in line with the results from other tree-ring studies on the climate sensitivity of dominant canopy trees of Fagus in various lowland or lower montane regions of Central Europe, including north-eastern and central-eastern Germany (Scharnweber et al. 2011; Zimmermann et al. 2015), Belgium (Kint et al. 2012), Luxembourg (Härdtle et al. 2013), north-eastern France (Bontemps et al. 2010; Charru et al. 2010), and Switzerland (Braun et al. 2015). Corresponding growth trends have also been reported for montane beech stands at the southern distribution limit of beech in Italy (Piovesan et al. 2003) and northern Spain (Jump et al. 2006). The evidence from Central Europe suggests that the observed recent vitality decline of Fagus in the center of its distribution range is not a local phenomenon but more widespread. Moreover, it cannot be explained as the response solely to certain severe drought events or as the outcome of demographic processes, because the growth decline started consistently in the 1970s or 1980s and continued until today, excluding causation by a single extreme event as the 1976 drought. Since only dominant trees in the upper canopy were sampled in most of these studies, shifts in the Fagus population structure can be ruled out as a cause as well. A simple age effect is also not plausible in our study and in those of Scharnweber et al. (2011) and Zimmermann et al. (2015), because the growth decline was observed only in certain stands, but not in others of similar age. The fact that the resilience of wood formation to individual drought episodes was <1 and thus relatively low in the sites at 110 and 300 m a.s.l. during the drought events in 1989, 1992, 2000, and 2004, but not in 1976, suggests that a negative impact of climate change on beech growth would have developed only since the 1980s. This pattern coincides with the increased growth sensitivity in the 2000–2013 interval compared to earlier decades in the 110 K and 300 W stands.
In the two stands with continued growth reduction, both spring and summer precipitation has declined in the last decades. However, the climate-response analysis shows sensitivity only to spring (March–May) precipitation and not to summer rainfall of the current year. This highlights the importance of a favorable water status during the period of early-wood formation for annual ring width suggesting that assessments of climate-change effects on the radial growth of beech must consider long-term trends in spring precipitation rather than summer or annual precipitation. The climate-response analysis further shows a negative impact of high temperatures in previous year’s June or July on radial growth. One possible explanation is reduced carbon gain in hot mid-summer periods and hence reduced carbohydrate availability for wood production in the subsequent year.
Growth stimulation at higher elevations
The situation is different at higher elevations. In our study region, the radial growth of the highest studied forest at 1230 m a.s.l. shows pronounced temperature limitation, and annual stem increment thus benefitted from the summer temperature increase in the last decades. In this stand at upper montane elevation, low summer temperatures limited wood formation despite the significant reduction in spring (May) precipitation observed during the last decades, i.e., in the physiologically most relevant precipitation component. Shortage of moisture in the period of main cambial activity cannot be an influential factor at this elevation, as the resilience of wood formation to individual drought events has remained invariably high during the last decades in the 1230S stand. We do not know to what extent increased atmospheric loads of reactive nitrogen have contributed to the recent increase in tree-ring width in this stand (de Vries et al. 2009), but the highly significant positive correlation of current-year summer temperature (June–August) with the tree-ring index in the climate-response analysis suggests that climate change is at least one of the factors significantly contributing to the recent increase in stem increment of the beeches at this elevation. Limitation of tree productivity by low summer temperatures is a characteristic of high-elevation forests around the world (Grace et al. 2002). Our conclusion is in agreement with dendroclimatological results of Dittmar (1999), di Filippo et al. (2007), and Babst et al. (2013) who found that F. sylvatica switches at montane elevation from moisture to temperature limitation of growth, which is at about 600–800 m a.s.l. in southern Germany.
The beech stand at 640 m a.s.l. is apparently close to the elevation, where apparent moisture or high-temperature limitation of growth switches to low-temperature limitation at higher elevations. As in most other temperate mountains, temperature and the evaporative demand decrease with elevation in the Black Forest, while precipitation does increase. The recent negative radial growth trend at 640 m a.s.l., which was weaker than in the low-elevation sites at 110 and 300 m a.s.l., suggests moisture limitation of stemwood formation also at this elevation, yet to a smaller extent than in the lowlands. However, in the climate-response analysis, negative correlation of the tree-ring index was found only with the previous year’s summer temperature, whereas summer (July) temperature of the present year was not negatively, but positively, correlated with the index. The latter correlation points to temperature limitation, but apparently to a much weaker extent than at 1230 m a.s.l., where the tree-ring index increased with the mean temperature of mid-summer (June–August) and with July temperature. Since stem increment is decreasing in recent time, moisture limitation seems to be more important for stemwood formation at 640 m a.s.l. than temperature limitation. The low resilience of tree-ring width to recent drought events at 640 m a.s.l., ranging in the same order of magnitude as in the low-elevation sites at 110 and 300 m a.s.l., also points to a dominant role of moisture limitation at this elevation.
The negative response of the tree-ring index to the previous year’s mid-summer (June–August) temperature in the stands at 110, 300, and 640 m a.s.l. and to previous year’s early summer (June) temperature in the high-elevation stand at 1230 m a.s.l. likely is attributable to the known induction of masting in F. sylvatica by hot summers with high radiation and/or temperature in the previous year being an important cue (Drobyshev et al. 2010; Müller-Hauboldt et al. 2015). A full mast consumes up to 60% of annual carbon gain in beech and results in pronounced decreases in stem growth in the year of, and the year following a masting event (Müller-Haubold et al. 2013, 2015). More frequent masting due to higher frequency of hot summers may indirectly cause reduction in stem increment, in addition to direct negative drought effects on growth, thereby reducing the competitiveness of beech. Increased fine root production under reduced precipitation and soil moisture as found by Hertel et al. (2013) may represent another driver of growth reductions in F. sylvatica in a warming climate.
Positive correlations between previous year’s autumn temperature and ring width may indicate that warmer autumn months extend the growing season and thus boost photosynthetic carbon gain in a season of reduced carbon demand for growth, allowing to store larger amounts of carbohydrates that are available for wood formation in the subsequent growing season (Dittmar and Elling 1999; Hacket-Pain et al. 2016). Higher spring precipitation at low, but not at high, elevation, where precipitation in this season is associated with cold weather and mostly falls as snow, also favors stemwood production, explaining the positive correlation of the tree-ring index with April and May precipitation at 110 m a.s.l. and with May precipitation at 300 m a.s.l., while this association does not exist at higher elevation. Reduced annual stem increment in years with low January and February temperatures at 1230 m a.s.l., and under low February temperatures at 640 m a.s.l., might be attributable to frost damage to the stem cambium or the root system in very cold winters (Dittmar et al. 2006). High radial growth rates in the 110 K and 300 W stands in the period 1950–1970/80 may be the result of selective logging activity, which reduced competition.
In our region, precipitation in April/May apparently was more influential than June–August precipitation as was found in other regions of Central Europe. Our data indicate a precipitation threshold range between 160 (sites 110 K and 300 W) and 235 mm (640H) of April/May precipitation below which temperature limitation of growth seems to switch to moisture limitation. These rainfall amounts correspond to a summer precipitation (June–August) range of ca. 200 mm (station Weinheim) to 313 mm (Hausach); negative growth trends due to moisture limitation may thus be expected at roughly 250 mm (or less) of summer precipitation. In central-eastern Germany, Zimmermann et al. (2015) derived from dendroclimatological data a precipitation limit for F. sylvatica in lowland and lower montane elevations in the range of c. 190 mm of June–August rainfall, corresponding to 600–640 mm year−1 MAP. This would indicate a somewhat higher precipitation threshold in south-western than in central-eastern Germany, which may be explicable by the higher summer temperatures and higher evaporative demand in the former region. Because we found significant precipitation–growth relationships only for 1 or 2 months of the year in spring, it is advisable to define precipitation limits for beech not on the basis of summer (or annual) precipitation but by referring to the sensitive months. Precipitation thresholds given by other authors based on mean annual precipitation are more variable (500–900 mm year−1; Czucz et al. 2011). Data from additional stands are needed to define the hygric threshold more precisely.
In the beech stands at lower elevation, the future development in the face of climatic warming is unclear. In unmanaged stands, possible consequences could be a demographic shift in the beech populations with gradual replacement of the currently dominant, exposed trees in the upper canopy by more vigorous, less climate-change-affected beech individuals from sub-canopy layers, increased mortality of large beech trees, or, in the long run, partial replacement of beech by more drought-tolerant species, such as Quercus petraea (Cavin et al. 2013). In many Central European lowland and lower montane regions, beech forest management likely will face a growing risk of productivity and vitality decline and possibly also higher mortality rate under a warming climate. This is indicated by dendroclimatological results from Western and South-western Europe, where drought sensitivity was higher in more northern beech forests than in stands at the southern range edge (Cavin and Jump 2016). In correspondence, Weber et al. (2013) reported a higher drought sensitivity of beech forests on mesic than on dry sites. Possible explanations are a better drought and heat adaptation of southern marginal populations or the selection of local sites with particularly favorable moisture regimes by the southern populations, i.e., avoidance of climate stress through habitat selection (Hacket-Pain et al. 2016).
These results may conflict with the intention of forestry politics in many countries to increase the share of beech in production forests owing to its dominant role in the natural forest vegetation (e.g., Rennenberg et al. 2004; Bayerische Staatsforsten 2011; Niedersächsische Landesforsten 2011). Long-term dendrochronological and demographic studies in forest reserves at different elevations and along precipitation gradients are needed to reach at reliable predictions about the future of monospecific and mixed beech forests in Central Europe (di Filippo et al. 2007). In particular, possible linkages between long-term growth decline and mortality rate in beech have to be analyzed.
Conclusions
Dendroclimatological evidence from this study and other stands in the center of the distribution range of F. sylvatica in western Central Europe with a beech-friendly sub-oceanic climate indicates a critical altitudinal range at c. 600 m a.s.l., where moisture limitation of radial growth at lower elevations switches to temperature limitation at higher elevations. Our results suggest that this threshold range is found in south-western Germany between c. 160 and 235 mm of April/May precipitation. In support of our hypothesis, a strong reduction of annual stem increment is visible at 110 and 300 m a.s.l. since more than 30 years, while the decline is less pronounced at 640 m a.s.l., and a growth increase has been recorded at 1230 m a.s.l. This response pattern cannot be attributed to an age effect or demographic processes, but must be caused by directly or indirectly acting climate-change-related factors, which are affecting the four stands differently. Reported growth trends from at least six Central European regions between north-eastern Germany and north-eastern France and Switzerland consistently evidence a more general vitality reduction of beech at lowland to lower montane elevations, which likely are caused by recent climate change, even though the physiological mechanisms in most cases are not fully understood. Given that the trend of summer warming will continue and summer rainfall is predicted to decrease in the future in various regions of Central Europe (Jacob 2009; IPCC 2013), the likely drivers of the vitality change will be strengthened and the growth decline may gain in momentum in the next decades.
Author contribution statement
CD and CL designed the study. CD (tree-ring sampling), GK, and MR (collection of stand data) performed field work. CD processed the tree-ring samples in the laboratory and performed the statistical analyses. CD, CL, and MH wrote the paper.
References
Ammer C, Albrecht L, Borchert H et al (2005) Zur Zukunft der Buche (Fagus sylvatica L.) in Mitteleuropa. Allg Forst- Jagdztg 176:60–67
Babst F, Poulter B, Trouet V, Tan K, Neuwirth B, Wilson R, Carrer M, Grabner M, Tegel W, Levanic T, Panayotov M, Urbinati C, Bouriaud O, Ciais P, Frank D (2013) Site- and species-specific responses of forest growth to climate across the European continent. Glob Ecol Biogeogr 22:706–717
Bayerische Staatsforsten (2011) Waldbauhandbuch Bayerische Staatsforsten. Grundsätze für die Bewirtschaftung von Buchen- und Buchenmischbeständen im Bayerischen Staatswald. Bavarian Ministry for Food, Agriculture and Forestry, Munich
Bindoff NL, Stott PA, AchutaRao KM et al (2013) Detection and Attribution of Climate Change: from Global to Regional. In: Stocker, T.F., D. Qin, G.-K. Plattner, M. Tignor, S.K. Allen, J. Boschung, A. Nauels, Y. Xia, V. Bex and P.M. Midgley (eds.) Climate change 2013: the physical science basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, New York
Bontemps JD, Herve JC, Dhote JF (2010) Dominant radial and height growth reveal comparable historical variations for common beech in north-eastern France. For Ecol Manage 259:1455–1463
Braun S, Remund J, Rihm B (2015) Indikatoren zur Schätzung des Trockenheitsrisikos in Buchen- und Fichtenwäldern. Schweiz Zschr Forstwes 166:361–371
Bunn AG (2008) A dendrochronology program library in R (dplR). Dendrochronologia 26:115–124
Cavin L, Jump AS (2016) Highest drought sensitivity and lowest resistance to growth suppression are found in the range core of the tree Fagus sylvatica L. not the equatorial edge. Glob Change Biol. doi:10.1111/gcb.13366 (online)
Cavin L, Mountford EP, Peterken GF, Jump AS (2013) Extreme drought alters competitive dominance within and between species in a mixed forest stand. Funct Ecol 27:1424–1435
Charru M, Seynave I, Morneau F, Bontemps J-D (2010) Recent changes in forest productivity: an analysis of national forest inventory data for common beech (Fagus sylvatica L.) in north-eastern France. For Ecol Manage 260:864–874
Czucz B, Galhidy L, Matyas C (2011) Present and forecasted xeric climatic limits of beech and sessile oak distribution at low altitudes in Central Europe. Ann For Sci 68:99–108
de Vries W, Solberg S, Dobbertin M, Sterba H, Laubhann D, van Oijen M, Evans C, Gundersen P, Kros J, Wamelink GWW, Reinds GJ, Sutton MA (2009) The impact of nitrogen deposition on carbon sequestration by European forests and heathlands. For Ecol Manage 258:1814–1823
di Filippo A, Biondi F, Cufar K, de Luis M, Grabner M, Maugeri M, Saba EP, Schirone B, Piovesan G (2007) Bioclimatology of beech (Fagus sylvatica L.) in the Eastern Alps: spatial and altitudinal climatic signals identified through a tree-ring network. J Biogeogr 34:1873–1892
Dittmar C (1999) Radialzuwachs der Rotbuche (Fagus sylvatica L.) auf unterschiedlich immissionsbelasteten Standorten in Europa. Bayreuther Bodenkundl Ber 67:208
Dittmar C, Elling W (1999) Jahrringbreite von Fichte und Buche in Abhängigkeit von Witterung und Höhenlage. Forstwiss Centralbl 118:251–270
Dittmar C, Fricke W, Elling W (2006) Impact of late frost events on radial growth of common beech (Fagus sylvatica L.) in southern Germany. Eur J For Res 125:249–259
Drobyshev I, Overgaard R, Saygin I, Niklasson M, Hickler T, Karlsson M, Sykes MT (2010) Masting behavior and dendrochronology of European beech (Fagus sylvatica L.) in southern Sweden. For Ecol Manage 259:2160–2171
Eckstein D, Bauch J (1969) Beitrag zur Rationalisierung eines dendrochronologischen Verfahrens und zur Analyse seiner Aussagesicherheit. Forstwiss Centralbl 88:230–250
Ellenberg H, Leuschner C (2010) Vegetation Mitteleuropas mit den Alpen in ökologischer, dynamischer und historischer Sicht, 6th edn. Ulmer, Stuttgart
Elling W, Heber U, Polle A, Beese F (2007) Schädigung von Waldökosystemen. Spektrum Verlag, Heidelberg
Filewod B (2011) Effects of an early season heat wave on eco-physiological parameters related to productivity in Sugar maple (Acer saccharum Marsh.). MSc Thesis, Faculty of Forestry, University of Toronto, Canada
Friedrichs DA, Trouet V, Büntgen U, Frank DC, Esper J, Neuwirth B, Löffler J (2009) Species-specific climate sensitivity of tree growth in Central-West Germany. Trees 23:729–739
Fritts HC (1976) Tree Rings and Climate. Elsevier
Gessler A, Keitel C, Kreuzwieser J, Matyssek R, Seiler W, Rennenberg H (2007) Potential risks for European beech (Fagus sylvatica L.) in a changing climate. Trees 21:1–11
Grace J, Berninger F, Nagy L (2002) Impacts of climate change on the tree line. Ann Bot 90:537–544
Hacket-Pain AJ, Cavin L, Friend AD, Jump AS (2016) Consistent limitation of growth by high temperature and low precipitation from range core to southern edge of European beech indicates widespread vulnerability to changing climate. Eur J For Res. doi:10.1007/s10342-016-0982-7 (online)
Härdtle W, Niemeyer T, Assmann T, Aulinger A, Fichtner A, Lang A, Leuschner C, Neuwirth B, Pfister L, Quante M, Ries C, Schuldt A, von Oheimb G (2013) Climate responses of tree-ring width and δ13C signatures of sessile oak (Quercus petraea Liebl.) on soils with contrasting water supply. Plant Ecol 214:1147–1156
Hertel D, Strecker T, Müller-Haubold H, Leuschner C (2013) Fine root biomass and dynamics in beech forests across a precipitation gradient–is optimal resource partitioning theory applicable to water-limited mature trees? J Ecol 101:1183–1200
IPCC (2013) Climate change 2013: the physical science basis. Contribution of working group I to the fifth assessment report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge
Jacob D (2009) Regionalisierte Szenarien des Klimawandels. Raumforsch Raumordn 67:89–96
Jump AS, Hunt JM, Peñuelas J (2006) Rapid climate change-related growth decline at the southern range edge of Fagus sylvatica. Glob Change Biol 12:2163–2174
Kint V, Aertsen W, Campioli M, Vansteenkiste D, Delcloo A, Muys B (2012) Radial growth change of temperate tree species in response to altered regional climate and air quality in the period 1901–2008. Clim Change 115:343–363
Köcher P, Gebauer T, Horna V, Leuschner C (2009) Leaf water status and stem xylem flux in relation to soil drought in five temperate broad-leaved tree species with contrasting water use strategies. Ann For Sci 66:11
Köcher P, Horna V, Leuschner C (2012) Environmental control of daily stem growth patterns in five temperate broad-leaved tree species. Tree Physiol 32:1021–1032
Lakatos F, Molnár M (2009) Mass mortality of beech (Fagus sylvatica L.) in south-west Hungary. Acta Silva Lign Hungarica 5:75–82
Leuschner C, Hertel D, Coners H, Büttner V (2001) Root competition between beech and oak: a hypothesis. Oecologia 126:276–284
Lindner M, Fitzgerald JB, Zimmermann NE, Reyer C, Delzon S, van der Maaten E, Schelhaas M-J, Lasch P, Eggers J, van der Maaten-Theunissen M, Suckow F, Psomas A, Poulter B, Hanewinkel M (2014) Climate change and European forests: What do we know, what are the uncertainties, and what are the implications for forest management? J Environ Manage 146:69–83
Lloret F, Keeling EG, Sala A (2011) Components of tree resilience: effects of successive low-growth episodes in old ponderosa pine forests. Oikos 120:1909–1920
Mette T, Dolos K, Meinardus C, Bräuning A, Reineking B, Blascke M, Pretzsch H, Beierkuhnlein C, Gohlke A, Wellsetin C (2013) Climatic turning points for beech and oak under climate change in Central Europe. Ecosphere 4:1–19 (Article 145)
Müller-Haubold H, Hertel D, Seidel D, Knutzen F, Leuschner C (2013) Climate responses of aboveground productivity and allocation in Fagus sylvatica: a transect study in mature forests. Ecosystems 16:1498–1516
Müller-Hauboldt H, Hertel D, Leuschner C (2015) Climatic drivers of mast fruiting in European beech and resulting C and N allocation shifts. Ecosystems 18:1083–1100
Niedersächsische Landesforsten (2011) The LÖWE Programme. 20 years of long-term ecological forest development. Lower Saxony State Forest, Braunschweig, Germany
Piovesan G, Bernabei M, Di Filipo A, Romagnoli M, Schirone B (2003) A long-term tree ring beech chronology from a high-elevation old-growth forest of Central Italy. Dendrochronologia 21:13–22
Piovesan G, Biondi F, Di Filipo A, Maugeri M (2008) Drought-driven growth reduction in old beech (Fagus sylvatica L.) forests of the central Apennines, Italy. Glob Change Biol 14:1265–1281
Pretzsch H, Bieber P, Schütze G, Uhl E, Rötzer T (2014) Forest stand growth dynamics in Central Europe have accelerated since 1870. Nature Comm 5 (article no. 4967)
R Development Core Team (2008) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org. ISBN 3-900051-07-0
Rennenberg H, Seiler W, Matyssek R, Gessler A, Kreuzwieser J (2004) Die Buche (Fagus sylvatica L.)–ein Waldbaum ohne Zukunft im südlichen Mitteleuropa? Allg Forst- Jagdztg 175:210–224
Scharnweber T, Manthey M, Criegee C, Bauwe A, Schröder A, Wilmking M (2011) Drought matters–declining precipitation influences growth of Fagus sylvatica L. and Quercus robur L. in north-eastern Germany. For Ecol Manage 262:947–961
Schönwiese CD, Janoschitz R (2008) Klima-Trendatlas Deutschland 1901–2000. Berichte des Instituts für Atmosphäre und Umwelt der Universität Frankfurt/Main 4:1–64
Tegel W, Seim A, Hakelberg D, Hoffmann D, Panev M, Westphal T, Büntgen U (2014) A recent growth increase of European beech (Fagus sylvatica L.) at its Mediterranean distribution limit contradicts drought stress. Eur J For Res 133:61–71
Thornthwaite CW (1948) An approach toward a rational classification of climate. Geogr Rev 38:55–94
Vincente-Serrano SM, Begueria S, Lopez-Moreno JI (2009) A multiscalar drought index sensitive to global warming: the standardized precipitation evapotranspiration index. J Climate 23:1696–1718
Weber P, Bugmann H, Pluess AR, Walthert L, Rigling A (2013) Drought response and changing mean sensitivity of European beech close to the dry distribution limit. Trees 27:171–181
Wigley TML, Briffa KR, Jones PD (1984) On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. J Appl Meteor Clim 23:201–213
Zang C, Hartl-Meier C, Dittmar C, Rothe A, Menzel A (2014) Patterns of drought tolerance in major European temperate forest trees: climatic drivers and levels of variability. Glob Change Biol 20:3767–3779
Zimmermann J, Hauck M, Dulamsuren Ch, Leuschner C (2015) Climate warming-related growth decline affects Fagus sylvatica, but not other broad-leaved tree species in Central European mixed forests. Ecosystems 18:560–572
Acknowledgements
The study was funded by the Landesanstalt für Umwelt, Messungen und Naturschutz Baden-Württemberg. We are grateful to Stefan Kaufmann (University of Göttingen) for his help during the field work and Jorma Zimmermann (University of Göttingen) for his support with the SPEI and SEA calculations. Bernd Raufeisen (University of Göttingen) prepared the map of the study area. We thank two anonymous reviewers for their constructive criticism of an earlier version of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by G. Wieser.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Dulamsuren, C., Hauck, M., Kopp, G. et al. European beech responds to climate change with growth decline at lower, and growth increase at higher elevations in the center of its distribution range (SW Germany). Trees 31, 673–686 (2017). https://doi.org/10.1007/s00468-016-1499-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-016-1499-x