
Abstract
Key message
Analyses of tree-ring anatomical features showed to be more sensitive to specific intra-seasonal environmental factors than tree-ring width in Tectona grandis and Pinus caribaea growing in a subtropical region.
Abstract
Earlywood vessels (EWV) and intra-annual density fluctuations (IADFs) in tree rings are influenced by intra-seasonal environmental factors, as have mostly been studied in temperate climate areas. However, it is not clear whether such anatomical traits can also be used as climate proxies in tropical regions. Therefore, the main objective of this study was to analyse the relationship between the anatomical features of two tropical species growing in Piracicaba, State of Sao Paulo, Southeast Brazil—EWV in Tectona grandis and IADFs in Pinus caribaea—and the climate. Wood cores were extracted by increment borer and processed using the classical methodology of dendrochronology. Chronologies of the mean vessel cross-sectional area of the first row of each tree ring (FRV) and the entire earlywood (EWV) in T. grandis, and of IADF in earlywood (IADF-E) and latewood (IADF-L) in P. caribaea were developed for the period 1988–2011 and 1982–2011, respectively. Our results showed a significant correlation of EWV and FRV with mean accumulated precipitation during the summer season (DJF) and with mean temperature in December. The IADF-Es in P. caribaea were related to the precipitation of DJF as well as the temperature of April, whereas IADF-Ls were linked to precipitation in fall (MAM). In general, the tree-ring anatomical variables were more influenced by intra-seasonal environmental factors than tree-ring width in both species, indicating their great potential for dendroecological studies in subtropical regions.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Tree rings are one of the most important proxies for reconstructing environmental data, since trees contain some of nature’s most accurate evidence of the past growth conditions at annual resolution (Fritts 1976; Schweingruber 1996). However, the influence of specific intra-seasonal environmental factors on tree growth is difficult to establish from tree rings (de Luis et al. 2011), as they integrate growth conditions over the entire growing season and partly respond with temporal delays to the controlling climatic factors (Eckstein 2004). In this context, one of the most important advances in dendrochronological studies has been the additional focus on anatomical features, i.e., tree-ring anatomy (Lindorf 1994). Indeed, some studies have shown that anatomical characteristics within tree rings may record potentially important information for ecological and climatological studies (Cherubini et al. 1996; Wimmer 2002; Fonti et al. 2010).
In angiosperms, studies of water-conducting elements in the xylem (vessels) are generally considered most interesting, whereas in conifers the same applies for intra-annual density fluctuations (IADFs). Characteristics of the vessels, such as the diameter, density and topology in a 3D network, play a fundamental role in regulating tree hydraulics and mechanical functioning (Tyree and Zimmermann 2002; Carlquist 2009; von Arx et al. 2013). In particular, plastic adjustments of vessel dimensions could be related to temperature and water availability (Fonti et al. 2007; Robert et al. 2009; von Arx et al. 2012; Fonti et al. 2013). IADFs—also referred to as “false rings”—are the results of changes in cambial activity induced by climate, which interrupt the normal course of growth during the growing season (Wimmer et al. 2000; Copenheaver et al. 2006; Edmonson 2010). This zone of altered growth in the tree ring is associated with an abrupt change in wood density (De Micco et al. 2014). False rings have been linked to drought and water stress, if occurring in the earlywood, and conditions stimulating growth, if occurring in the latewood, as evidenced in several studies of angiosperms in temperate (Masiokas and Villalba 2004; Campelo et al. 2007; Copenheaver et al. 2010; Battipaglia et al. 2014) and tropical climates (Priya and Bhat 1998). But most of the studies have considered the conifer genus Pinus because of the easy identification and high frequency of IADFs (e.g., De Luis et al. 2011; Rozas et al. 2011; Marchand and Filion 2012; Campelo et al. 2013).
Over the last decade, the analysis of ring width chronologies in the tropics has developed rapidly (Rozendaal and Zuidema 2011), whilst the use of tree-ring anatomical chronologies has still been comparatively rare. Accordingly, the numerous studies showing the high suitability of tree-ring width for evaluating the climate sensitivity of tropical tree growth (e.g., Brienen and Zuidema 2005; Soliz-Gamboa et al. 2011; Zuidema et al. 2012) profited from important methodological advances in the field (Rozendaal and Zuidema 2011). However, despite its rare use in the tropics, the variability in intra-ring features may record climate information that is potentially complementary to tree-ring width (Fonti et al. 2010), which would improve climate reconstructions in tropical and subtropical regions. The aim of this study was to investigate the relationship between local climate variability and tree-ring anatomy of two tropical species—earlywood vessels in Tectona grandis and IADFs in Pinus caribaea—and to compare it to the relationship of climate with tree-ring width. Furthermore, evidence for intra-seasonal phenotypic plasticity in xylem anatomy could be used as ecological proxies.
Materials and methods
Study area and species
The study was conducted in an Atlantic forest fragment of Piracicaba (14 ha), located in the western part of the State of São Paulo, Brazil (22°42′S, 47°37′W, 546 m), bordering on the Piracicamirim River. In this site, several exotic tree species, with predominance of T. grandis (Verbenaceae) and P. caribaea var. hondurensis (Pinaceae), were planted four to five decades ago. Whilst the dendrochronological potential of these two species has already been demonstrated in other regions by other authors (e.g., Worbes 1999; Shah et al. 2007; Cook et al. 2010; Olajidel et al. 2010), an evaluation of their potential for investigating climate–tree relationships in combination with specific wood anatomical features is still missing. In addition, due to the great adaptability to Brazilian soil and climatic conditions, plantations of teak and Caribbean pine have become more common in the last three decades. Furthermore, these species are found growing in natural forests comprising the local flora (Lilienfein et al. 2000; Schuhli and Paludzyszyn 2010).
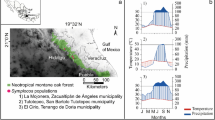
According to the Köppen classification, the climate in the study region is Cfa (humid subtropical) (Alvares et al. 2013). There is clear seasonality, with a warm and rainy period coinciding with a longer photoperiod (approximately from October to May) and cooler dry season coincident with a shorter photoperiod (from April to September). In the summer, the mean temperature is 24.5 °C and the accumulated precipitation is 630 mm, and in the winter 18.1 °C and 103 mm, respectively (Fig. 1).

Study area in Piracicaba, State of São Paulo, and climatic diagram of the meteorological station ESALQ-USP for the period between 1970 and 2011. The blue bars represent mean monthly precipitation sums and the red lines mean monthly temperature
Tectona grandis (teak) is a hardwood species originally growing in tropical Asia, which was introduced in Brazil due to its economic importance for timber production (Tomazello and Cardoso 1999). The species is deciduous and develops a rounded canopy with large leaves, and a smooth and cylindrical trunk of more than 25 m height (Pandey and Brown 2000). Teak tree rings are classified as ring-porous (Gottwald and Parameswaran 1980; Pumijumnong et al. 1995). P. caribaea (Caribbean pine) is a species originally occurring in Central America. Due to its great ecological plasticity, it became the most-used tropical pine species in reforestation programmes. Brazil features the largest planting area of P. caribaea, reflecting the species’ value for timber and numerous other products (Eguiluz 1982). The tree-ring boundaries between the latewood and earlywood are marked by the difference of cell-wall thickness and lumen area (Ferreira and Tomazello Filho 2009).
Sampling, wood preparation and building of tree-ring width chronologies
As the analysis of chronologies of vessel size and IADFs is a relatively recent technique, there is no consensus in the scientific dendrochronological community as to the best method to statistically assess the quality of these chronologies. Therefore, we created a tree-ring width (RW) chronology of both species to determine the cambial age and identify the tree rings to be used for subsequent analysis of the intra-annual anatomical features (vessels for T. grandis and IADFs for P. caribaea).
For tree-ring width analysis, we selected eight living trees of T. grandis and eight living and two dead trees of P. caribaea. Two to six wood cores from each tree were sampled at breast height, in May 2012, and the best two (and in a few cases three) cores per tree were selected and averaged to build the chronologies based on the criterion of entire sample and visible rings. The population of both species is very small; in the case of T. grandis, all trees were sampled and in P. caribaea, about 50 % were analysed, thereby avoiding individuals attacked by pathogens or having compression wood. Since the sampled trees were young, essentially all rings were included.
Wood cores were glued on a wooden holder and cross-section surfaces were polished with a series of 80–600 grit/mm2 sand paper to highlight the tree-ring boundaries. The demarcation, counting and measuring of the tree-ring widths were performed using the software Image-Pro Plus, version 4.5.0.29 (Media Cybernetics Inc, Rockville, MD, USA). The software COFECHA was used for crossdating tree rings (Holmes 1986), obtaining a 41-year-long chronology (1971–2011) for T. grandis and a 36-year-long chronology (1976–2011) for P. caribaea. Age- and size-related trends of trees (low-frequency variation) were standardised by detrending with a negative exponential function or a linear regression with negative slope using software TURBO ARSTAN 2.07 (Cook 1999). The indices were calculated as ratios between the actual and fitted values (growth trend) of tree-ring width. The standardised tree-ring series were averaged to produce a mean chronology (Fritts 1976). To avoid a potential bias in a way that could lead to inflated recent growth increase estimates (Cook and Peters 1997), residual chronology from the growth curve, rather than ratios, was applied to the climatological analysis.
The quality of each chronology was analysed by the mean correlation between series (rbs) and the expressed population signal (EPS). The chronologies were described by classical statistical parameters of dendrochronology (Fritts 1976), i.e., the average tree-ring width, standard deviation, mean sensitivity (MS) and first-order autocorrelation (AR).
Analysis of vessels in Tectona grandis
Cores with good correlation with the master chronology of COFECHA (greater than a correlation coefficient of 0.60 core/tree) were selected to analyse vessels in T. grandis, comprising 12 cores from 8 trees. When two cores were present in one tree, average measurements were used. The wood samples were scanned on a flatbed Epson Perfection V750 Pro scanner at 4800 DPI in 48-bit colour. The vessels were subsequently identified using Image-Pro Plus, by semi-automatic drawing of all the earlywood vessels in a region of 2 mm of tree-ring width, and their cross-sectional lumen area (henceforth “vessel size”) was subsequently measured. The variables analysed were average size of earlywood vessels (EWV) and average size of the vessels belonging to the first row of each tree ring (FRV) (Fig. 2). Earlywood vessels were visually distinguished from latewood vessels by the clear size difference known in ring-porous species. Standardised chronologies for each variable were also produced using the software TURBO ARSTAN 2.0.7.

Analysis of tree-ring vessels in T. grandis as manually demarcated using the software Image-Pro Plus, in a region of 2 mm of width and the total length of each tree ring, showing size of earlywood vessels (EWV) and vessels belonging to the first row of each tree-ring (FRV). Vertical bars in the left panel show the location of tree rings. The arrow indicates the direction of growth. Scale bar 1 mm
Analysis of IADFs in P. caribaea trees
For the creation of the IADF chronologies, we used only those cores from living trees that were used in the crossdating in COFECHA (18 cores from 8 trees). To avoid pseudo-replication, an average of IADF presence in the cores of the same tree was used. The IADFs were visually defined by one investigator to maintain consistency as tangential zones in the tree rings showing abnormal growth. In the earlywood, they were visible as bands consisting of tracheids with reduced lumen area and increased cell-wall thickness (IADF-E), whereas in the latewood earlywood-like tracheids characterised IADF-Ls (Fig. 3).

Examples of intra-annual wood density fluctuations (IADFs) in earlywood (E) and latewood (L) of P. caribaea. The horizontal arrows indicate the IADFs and vertical bars in the left panel show the extension of tree rings. The vertical arrow indicates the direction of growth. Scale bar 1 mm
The relative frequency of IADFs per year (F) was calculated using the following equation:
where N is the number of trees with presence of IADFs in a given year and n is the total number of trees sampled in that year. The changing sample depth (n) over time generated a bias for the frequency variance, which was corrected by calculating the stabilised IADF frequency as:
where f is the relative frequency of IADFs (Osborn et al. 1997). The significant vs random occurrence of IADFs in any given year was tested using the binominal distribution test adopted from Edmondson (2010), which calculates the 95 % confidence intervals (CI) around the proportion of false rings in a given year. To be significant, the IADF frequency and p-CI must exceed the random background frequency of IADF occurrence. This background value was calculated as the ratio between all rings with IADFs and the total number of dated rings, and represents the approximate frequency of an entirely random inter-annual occurrence in a given year.
Signal strength amongst tree-ring anatomical chronologies
The common signal between five chronologies (RW teak and pine, EWV, FRV and IADFs) was then checked in the overlapping period using several measures. The Gleichläufigkeit index (GLK), expressed as a percentage, was calculated to compare the interval trends between different chronologies and to examine how the chronologies followed each other in their growth patterns year-to-year (Schweingruber 1988). The Baillie Pilcher t values measured the strength of agreement between two chronologies using the correlation coefficient r, the coefficient of determination R 2 and the n years analysed (Baillie and Pilcher 1973). In addition, Pearson correlation coefficients were calculated.
Relationship of the tree-ring variables with climate
Correlation analysis was used to establish the relationship between climate and each tree-ring variable chronology (RW, EWV, FRV and IADFs). Therefore, each chronology was related to monthly average temperature and total monthly precipitation from previous June (before the growing season) to current May (end of the growing season). To include an additional intra-annual time scale, the same analysis was performed for each quarter of the year (JJA, SON, DJF and MAM), coinciding with the four seasons. For RW, EWV and FRV bootstrapped correlation coefficients were used. The climatic signals of IADFs were analysed only for the significant years according to the background value (see above). IADF-Es were investigated using Spearman’s correlation coefficients, whilst IADF-Ls were analysed with standard deviations (SD) due to the abundance of non-significant years. For this, climatic variables were normalised and the SD was calculated from years with and without significant IADF-Ls. SD values above 0.5 or below −0.5 were considered as significantly positive or negative anomalies, respectively (Masiokas and Villalba 2004; Rozas et al. 2011). To avoid potential problems with ontogenetic trends and lower values of the variables, the first 6 (P. caribaea) and 12 years (T. grandis) of growth were excluded from analysis (Fig. 4).

a Mean (+SD) frequency of size of earlywood vessels (EWV) in 12 cores of T. grandis and b mean (+SD) frequency of intra-annual density fluctuations (IADF) in 18 cores of P. caribaea in different tree-ring age classes. Inset tables contain main statistical parameters. Vertical dashed lines indicate the lower cut-off period used for analysis to avoid potential problems with ontogenetic trends
We used meteorological data from the ESALQ-USP station, Piracicaba-SP, which is within 250 m from the study area. The beginning of the growing season was considered in November for T. grandis (Tomazello and Cardoso 1999), and September (precipitation higher than 60 mm) for P. caribaea, according to Worbes (1995). Growing season typically ends in May for the two species. Statistical analysis was also performed using R 3.0.3 (http://cran.r-project.org/bin/windows/base/old/3.0.3/), package dplR (Bunn 2008) and package bootRes (Zang and Biondi 2013), and significance was assumed at p = 0.05.
Results
Tree-ring width chronologies
Table 1 shows a summary of the statistics used to evaluate the quality of the different chronologies obtained. The average tree-ring width was 6.4 mm for P. caribaea and 4.9 mm for T. grandis. The ring width chronologies of both species were not significantly related to each other according to the low values of GLK, t values and correlation coefficient (Table 2). These values may be influenced by divergent short-term variability in tree-ring growth between the species. In the first 15 overlapping years, P. caribaea tree-ring widths were wider and decreased thereafter with age, whilst in T. grandis tree-ring widths were wider in the last 20 years. However, a common decreasing tendency of tree-ring widths in the last few years, especially in 2011, was observed for both species (Fig. 5a).

a Mean tree-ring width in mm (±SE) vs growth ring year, and growth tendency according to quadratic regression of both series (significant models with p < 0.001), blue (T. grandis) and red line (P. caribaea). b The standard tree-ring chronology for the period 1976–2011 and 1971–2011 for T. grandis and P.caribaea, respectively. c Rbar and EPS statistics; the moving window used was 10 (12) years with an overlap of 5 (6) years between adjacent windows for T. grandis (P. caribaea); the dashed line indicates the acceptable EPS threshold of 0.85 (Wigley et al. 1984) and for Rbar, the vertical bars represent the two standard error limits
For both species, the series intercorrelation exceeded 0.64 (threshold for significance was 0.51) and T. grandis had a lower first-order autocorrelation coefficient. The MS values of T. grandis trees were greater than for P. caribaea, although both species proved to be sensitive with values higher than 0.30 (Grissino-Mayer 2001). The expressed population signal (EPS), was higher than 0.85, indicating that sample size was adequate for the period of the two species evaluated according to Wigley et al. (1984). However, Rbar and EPS (Fig. 5c) showed that sample quality varied across the length of the chronologies in the two species with low values in late 1980 s and early 1990 s for teak, associated with a decrease in the intercorrelation in this period, and in the last 6–7 years for Caribbean pine, due to the shorter tree-ring series obtained from the two dead trees analysed.
In the ring width chronology of P. caribaea, a dramatic decrease of growth in 1981 was observed (Fig. 5b) due to a fire in that year in this forest fragment (Catharino 1989). This radial growth behaviour is consistent with other fires studies dated by tree-ring analysis (Miller 2007) and ecological interactions with fire (O’Brien et al. 2008).
Characteristics of earlywood vessels in T. grandis and IADF in P. caribaea
Figure 6 compares the standardised chronologies of vessels and RW in T. grandis for the period 1988–2011, and IADFs and RW in P. caribaea for the period 1982–2011. In total, 9,146 earlywood vessels were analysed in the 12 cores of T. grandis, of which 30.6 % belonged to the first row of earlywood vessels (FRV). The average size of FRV was larger than EWV in the analysed period (37.0 and 35.0 × 103 µm2, respectively), and both had low mean sensitivity and first-order autocorrelation coefficients. Although Rbs and EPS were not significant for the two variables (Table 1), they were still high compared to other studies using anatomical variables (García-González and Fonti 2006; Campelo et al. 2010; Locosselli et al. 2013; Olano et al. 2013).

a Standardised chronologies of vessel characteristics (EWV and FRV) vs tree-ring width (RW) in T. grandis. b Intra-annual density fluctuations (IADFs) vs RW in P. caribaea. c Annual mean temperature vs annual total precipitation. To avoid potential problems with ontogenetic trends (see Fig. 4) and a disturbance through fire in 1981, we only considered the period 1988–2011 for T. grandis and 1982–2011 for P. caribaea. The asterisk symbol in the middle panel indicates significant occurrence of IADFs with respect to the random background value (69.0 %)
We counted 507 IADFs in 18 cores of the eight P. caribaea trees, obtaining a mean of 1.05 IADF per ring. Whilst 56.5 % of all tree rings showed IADFs, the frequency of type E was much more abundant (52.8 %) than type L (18.5 %). Considering the 30 years of analysis, 100 % of trees formed at least three rings with IADF-E, but only the 62.5 % of trees formed at least three rings with IADF-L. The co-occurrence of IADFs was statistically significant for 18 years of the analysed period, according to the randomised background frequency (Table 1).
Results of the correlations amongst the considered anatomical variables are shown in Table 2. T. grandis tree-ring widths were closely related to the two vessel variables indicated by the significant t values, GLK values and correlation coefficient. In addition, there was a very strong common signal between EWV and FRV.
Although RW of T. grandis and P. caribaea had no common signal (Fig. 5b), there was an association between IADFs and vessels variables. FRV and IADF-Es had a significant GLK value, showing that these two variables may be influenced by the same climatic parameters. The two vessel variables and IADF-Ls showed a significant negative correlation.
Relationship between tree-ring variables and climate
The climate-growth relationships for all anatomical features are summarised in Fig. 7 (T. grandis, 1988–2011) and Fig. 8 (P. caribaea 1982–2011). For precipitation, the ring width and two tree-ring anatomy chronologies of T. grandis were significantly correlated with total summer rainfall (DJF) of the current growing season, showing the strongest relationship (p < 0.01). These results were influenced by December and January in FRV and EWV, and January and February in RW. For average temperature, RW did not show any significant correlation with the quarterly period, whilst EWV was significantly positively correlated with temperatures of spring and summer and FRV with summer temperature. All variables were influenced by temperature of the current December, being highly significant for EWV (p < 0.01). In addition, May temperature had a negative relationship in the three variables.

a–c Bootstrapped correlations of EWV, FRV and RW chronologies of T. grandis with the accumulated precipitation (P) and average temperature (T) from June to May from 1988 to 2011. Bars on the right of the vertical line represent the 3-month seasons. Horizontal dashed line indicates the current growing season. Months followed by “+1” indicate year +1 due to biological age ≠ calendar age (e.g., the tree ring of 2011 was formed between November 2010 and May 2011). Significance level *p < 0.05 and **p < 0.01

a Standard deviations of IADFs type L chronology with accumulated precipitation (P) and average temperature (T), b Spearman’s correlations of IADFs type E chronologies with P and T and c bootstrapped correlations of RW chronologies of P. caribaea with P and T from June to May (17 years in IADF-E and 12 years in IADF-L) in the period 1982–2011. Bars on the right of the vertical line represent the 3-month seasons. Horizontal dashed line indicates current growing season. Months followed by “+1” indicate year +1 due to biological age ≠ calendar age (i.e., tree ring of year 2011 was formed between September 2010 and May 2011). Significance level: *p < 0.05 and **p < 0.01 to IADF-E and RW; filled circle SD > 0.5 to IADF-L
In P. caribaea, RW and IADF-L were significantly correlated with total precipitation of fall (MAM), whilst IADF-E was associated with total summer rainfall during the growing season. The monthly climatic analysis showed that RW and IADF-E were significantly correlated with current February precipitation, highlighting the strongest relationship of IADF-E (p < 0.01), whilst IADF-Ls were linked with the current March and May precipitation. Temperature of current April was significantly correlated with the formation of IADF-E. RW was negatively related to current March temperature and August temperature before the growing season.
Discussion
The objective of this study was to investigate whether the variation in anatomical features of tree rings—earlywood vessel size in T. grandis and IADFs in P. caribaea—was more sensitive to local climate than tree-ring width in a tropical region. Despite the relatively short tree-ring anatomical chronologies due to the young age of the sample trees, our results suggest that the vessel variables and IADFs are indeed more influenced by monthly and seasonal climate than ring width.
Plastic responses of earlywood vessel size in Tectona grandis to climate
The observed positive relationship between the earlywood vessels (FRV and EWV) and precipitation and temperature in summer can be explained by the need for a more efficient water transport due to the increased evapotranspiration. Current understanding is that narrower vessels are linked to drier conditions, thus increasing hydraulic safety. On the other hand, wider vessels are linked to moister conditions, thus increased hydraulic efficiency (Bhattacharyya et al. 2007; Giantomasi et al. 2009; Fonti et al. 2010; von Arx et al. 2012), which was also observed in this study. Whilst the plastic responses of vessel size were related to water limitation (Campelo et al. 2010), we found the plasticity to occur early in the season (December), a time when water was likely not limiting in Piracicaba (Fig. 1). Earlywood vessel size (EWV and FRV) thus seemed to increase in response to even more ample water, which would be a release response as opposed to a limitation response. This could be possibly explained by the need to exploit the soil water faster and more efficiently to successfully compete with other trees. Investing into a more efficient hydraulic system could, therefore, be a way to promote the observed “explosive growth” in early season (see below).
Temperature seems to have an influence on earlywood vessels size through a complex interaction with cell division and differentiation (Fonti and García-González 2004). Besides summer environment, we observed that spring conditions also had a direct influence on the formation of EWV; thus, we interpret that warmer temperature favours an efficient water-transporting system. Our results also coincide with Tomazello and Cardoso (1999) who studied seasonal variations of cambial activity in T. grandis plantations in the same study region. They found that differences in fusiform cells formed at the end of November–December and in the dormancy period reached their highest activity in January–February.
Intra-seasonal climate parameters cause the formation of false rings in P. caribaea
In the current season, IAFD-E formation is triggered primarily by summer precipitation, especially in February. This suggests that abundant precipitation combined with higher temperatures (it tended to be significant; Fig. 8a) during the rainy period induces the formation of IADF-Es mainly due to increased evapotranspiration rates (Campelo et al. 2013), which decreased cambial activity and produced latewood-like cells. However, such results could be interpreted differently. IADF-Es are a typical response triggered by drought stress associated with depletion of the soil water storage (Rozas et al. 2011; Campelo et al. 2013), in years with dry summers (Panayotov et al. 2013). Our study showed this response is associated with the rainy period, so the formation of narrower thicker-walled earlywood would be influenced by the water saturation in the soil (Young et al. 1993), caused by water saturation characteristic of the rainy season. Some studies indicated such changes in the wood anatomy can be attributed to flash-flood events (Ballesteros et al. 2010; Leoni et al. 2011). This interpretation may be supported by the soil characteristic in the study area, which is a shallow soil, classified as eutroferric cambisol type, influenced by the Piracicamirim River.
The formation of IADF-L and the RW were associated with rainy periods in MAMc, which is related to favourable conditions of soil water storage (De Luis et al. 2011), and so it causes a extended growing season in fall. Therefore, these results indicate that although at the end of the tree growth period, P. caribaea trees should enter into cambial dormancy, an increase in water availability in the soil can reactivate cambial cells, producing more xylem in that specific year. Such results also coincide with some other studies on Pinus sp. (Vieira et al. 2009; De Luis et al. 2011; Rozas et al. 2011; Campelo et al. 2013).
IADF-Es showed positive relationships with April temperature, when highest values increased the evapotranspiration. However, we observed a negative relationship between RW and current March temperature, which means that the increasing temperature towards the end of the growing season caused a negative effect on radial growth, triggering the formation of tracheids with thicker walls and reduced lumen area. This confirms the sensitivity of cell-wall thickness to temperature.
Our results suggest that, during the growing season, precipitation of the rainy period (DJF) and temperature of April are the main factors triggering the formation of IADF-E, whilst IADF-L formation is related to precipitation of fall season (MAM). In addition, the high frequency of IADFs found here for P. caribaea trees—notably the first report of IADFs in trees grown in Brazil—shows that the anomalies in radial growth may be key to future research in tropical pine planted in Brazil. The analysis of IADF dynamics can provide new perspectives on the role of climatic parameters in the ecology of inter-annual wood in tropical trees.
The influence of growth strategy on climate sensitivity
Our results are consistent with those by Tomazello Filho and Cardoso (1999), who observed teak trees in Brazil have the highest cell production in the cambium during the summer. One possible explanation lies in the fact that teak, as a deciduous species, defoliates between July and October in the study area. In November, during the sprouting of new leaves, rainfall plays an important role in FRV formation, as well as during the summer season (December–February), when greater radial growth and earlywood vessel formation were observed. These results are similar to those found by Pumijumnong (1997) for cambial development of teak trees in Thailand. The authors observed that the beginning of the rainy period determined the growing season and its cambial activity, which was restricted to a short period of 4 months. Subsequent to that period, narrower vessels are formed, constituting semi-ring-porous tree rings, a characteristic of the species and a result of seasonal fluctuations of cambial activity (Priya and Bhat 1999; Rao and Rajput 1999). A similar behaviour was observed at teak plantations in South America (Worbes 1999) and Africa (Olajidel et al. 2010).
Unlike teak trees that need a rainy summer, we can infer that the Caribbean pine trees cease growth in this season evidenced by the formation of IADF-E (Fig. 8b). Our results can indirectly estimate the key period of cambium reactivation, which is linked to precipitation during dry months before growing (JJA), with marginal significance to tree-ring width (Fig. 8c). Similar results were found by Singh and Venugopal (2011) for Pinus kesiya growing in a subtropical wet forest of Northeast India. In this sense, evergreen conifers are continuously regulated by primary productivity and only need the first rainfalls to reactivate the xylem formation (Jacoby and D’Arrigo 1995; Pumijumnong and Wanyaphet 2006). As P. caribaea trees never become leafless in the year due to a continuous production of new needles, the xylem production period is extended when compared to T. grandis.
In T. grandis trees, growth surges explosively in summer (rainy period) when new leaves are at their maximum photosynthetic capacity, thus showing wider vessels. Comparing secondary growth strategies of the two species, a phenological pattern is present irrespective of water availability, since species are growing under the same environmental conditions. The tropical evergreen species have higher water use and tree transpiration rates during the dry season. In contrast, deciduous trees (especially T. grandis) use less water during the dry season than the wet season, which can be explained by the reduced number of leaves during the dry season (Kunert et al. 2010). Wyka et al. (2012) observed that the maximum photosynthesis rate per unit of leaf mass is much greater in deciduous broadleaved angiosperms than evergreen conifers, since photosynthesis occurs only when the trees gain their full set of mature leaves—a result of a higher contribution of chloroplasts containing mesophyll tissue to leaf biomass. This is also supported by Die et al. (2012) who asserted that T. grandis trees must develop early growth for water transport, when transpiration, photosynthesis and radial growth increase since the trees have full foliage.
Potential of vessel size and IADFs in tropical trees for dendroclimatological studies
This pioneering work in Brazil showed that the anatomical parameters in both species were better, or at least well correlated with monthly and seasonal climate, as was tree-ring width. In this sense, vessel size and IADF analyses are very attractive alternatives to ring width analysis for climate studies in the tropics, particularly if different life-history strategies of species are considered. In addition, including these variables may accurately link tree growth to sub-annual climate condition, as they are directly affected by the conditions at the time of formation, whereas tree-ring width integrates conditions over the entire growing season. In tropical and subtropical regions in South America, there are few studies concerning climatic drivers that affect tree growth (e.g., Worbes 1999; Schongart et al. 2004; Brienen and Zuidema 2005; Oliveira et al. 2010), so that microanatomy chronologies allow us to better understand the effect of precipitation and temperature on cellular anatomy of the tree rings in those regions. Although we worked with exotic tropical species, results of our vessels size and IADF investigation may be relevant for future climatic studies in native tropical species with proven potential for dendroclimatology and dendroecology. For example, in Brazil, EWV and FRV could be useful for Cedrela odorata (Dunisch et al. 2003, Brienen and Zuidema, 2005) and Swietenia macrophylla (Worbes 1999; Dunisch et al. 2003), and IADF could be used in Araucaria angustifolia (Oliveira et al. 2010) and Podocarpus lambertii (Mattos et al. 2007).
In summary, under the current global climate change scenario, it is known that tropical trees will be amongst those strongly affected (Zeng et al. 2013). Therefore, studies of intra-annual tree growth are important to better understand the influence of intra-seasonal environmental factors, and to better connect them to the ecophysiology of tropical trees, which could be useful for climate change and tree biology studies.
Conclusions
This study explored an incipient research area in dendrochronology of tropical species showing that T. grandis and P. caribaea trees growing in southeast of Brazil are very sensitive to seasonal climatic parameters. Chronologies based on vessel size and IADF were better correlated with climate than tree-ring width. Since such anatomical traits contain important ecophysiological information, they may be used for integrated climate studies, in addition to the classical dendrochronological analysis.
Author contribution
The authors participated in drafting the article or revising it critically for important intellectual content, as well as approving the submitted and revised version. Acquisition of data: AVG; MPC analysis and interpretation of data: AVG; GVA; MPC; MTF manuscript drafting: AVG; GVA; MTF critical revision: AVG; GVA; MPC; MTF.
References
Alvares CA, Stape JL, Sentelhas PC et al (2013) Köppen’s climate classification map for Brazil. Meteorol Z 22(6):711–728. doi:10.1127/0941-2948/2013/0507
Baillie MGL, Pilcher JR (1973) A simple crossdating program for tree-ring research. Tree-Ring Bull 33:7–14
Ballesteros JA, Stoffel M, Bodoque JM, Bollschweiler M, Hitz O, Diez-Herrero A (2010) Changes in wood anatomy in tree rings of Pinus pinaster Ait. following wounding by flash floods. Tree-Ring Res 66:93–103. doi:10.3959/2009-4.1
Battipaglia G, De Micco V, Brand WA, Saurer M, Aronne G, Linke P, Cherubini P (2014) Drought impact on water use efficiency and intra-annual density fluctuations in Erica arborea on Elba (Italy). Plant Cell Environ 37:382–391. doi:10.1111/pce.12160
Bhattacharyya A, Eckstein D, Shah SK, Chaudhary V (2007) Analyses of climatic changes around Perambikulum, South India, based on early wood mean vessel area of teak. Curr Sci 93:1159–1164
Brienen R, Zuidema P (2005) Relating tree growth to rainfall in Bolivian rain forests: a test for six species using tree ring analysis. Oecologia 146:1–12. doi:10.1007/s00442-005-0160-y
Bunn AG (2008) A dendrochronology program library in R (dplR). Dendrochronologia 26:115–124. doi:10.1016/j.dendro.2008.01.002
Campelo F, Gutierrez E, Ribas M, Nabais C, Freitas H (2007) Relationships between climate and double rings in Quercus ilex from northeast Spain. Can J Forest Res 37:1915–1923. doi:10.1139/X07-050
Campelo F, Nabais C, Gutierrez E, Freitas H, García-González I (2010) Vessel features of Quercus ilex L. growing under Mediterranean climate have a better climatic signal than tree-ring width. Trees 24:463–470. doi:10.1007/s00468-010-0414-0
Campelo F, Vieira J, Nabais C (2013) Tree-ring growth and intra-annual density fluctuations of Pinus pinaster responses to climate: does size matter? Trees 27:763–772. doi:10.1007/s00468-012-0831-3
Carlquist S (2009) Non-random vessel distribution in woods: patterns, modes, diversity, correlations. ALISO 27:39–58. doi:10.5642/aliso.20092701.04
Catharino ELM (1989) Estudos fisionômico-florísticos e fitossociológico em matas residuais secundárias do município de Piracicaba, SP. Dissertation, University of Campinas
Cherubini P, Piussi P, Schweingruber FH (1996) Spatio temporal growth dynamics and disturbances in a subalpine spruce forest in the Alps: a dendroecological reconstruction. Can J Forest Res 26:991–1001. doi:10.1016/S1631-0691(03)00075-1
Cook ER (1999) TurboARSTAN program and reference manual V 2.0.7 February. Palisades, New York
Cook ER, Peters K (1997) Calculating unbiased tree-ring indices for the study of climatic and environmental change. The Holocene 7(3):361–370. doi:10.1177/095968369700700314
Cook ER, Anchukaitis KJ, Buckley BM et al (2010) Asian Monsoon failure and megadrought during the last millennium. Science 328:486–489. doi:10.1126/science.1185188
Copenheaver CA, Pokorski EA, Currie JE, Abrams MD (2006) Causation of false ring formation in Pinus banksiana: a comparison of age, canopy class, climate, and growth rate. For Ecol Manage 236:348–355. doi:10.1016/j.foreco.2006.09.020
Copenheaver CA, Gartner H, Schafer I, Vaccari FP, Cherubini P (2010) Drought-triggered false ring formation in a Mediterranean shrub. Botany 88:545–555. doi:10.1139/B10-029
De Luis M, Novak K, Raventós J et al (2011) Climate factors promoting intra-annual density fluctuations in Aleppo pine (Pinus halepensis) from semiarid sites. Dendrochronologia 29:163–169. doi:10.1016/j.dendro.2011.01.005
De Micco V, Battipaglia G, Cherubini P, Aronne G (2014) Comparing methods to analyse anatomical features of tree rings with and without intra-annual density fluctuations (IADFs). Dendrochronologia 32:1–6. doi:10.1016/j.dendro.2013.06.001
Die A, Kitin P, Kouame FN, Van den Bulcke J, Van Acker J, Beeckman H (2012) Fluctuations of cambial activity in relation to precipitation result in annual rings and intra annual growth zones of xylem and phloem in teak (Tectona grandis) in Ivory Coast. Ann Bot 110:861–873. doi:10.1093/aob/mcs145
Dunisch O, Montoia VR, Bauch J (2003) Dendroecological investigations on Swietenia macrophylla King and Cedrela odorata L. (Meliaceae) in the central Amazon. Trees 17:244–250. doi:10.1007/s00468-002-0230-2
Eckstein D (2004) Change in past environments: secrets of the tree hydrosystem. New Phytol 163:1–4. doi:10.1111/j.1469-8137.2004.01117.x
Edmondson JR (2010) The meteorological significance of false rings in eastern redcedar (Juniperus virginiana L.) from the Southern Great Plains. USA. Tree-Ring Res 66:19–34. doi:10.3959/2008-13.1
Eguiluz PT (1982) Clima y distribución del genero Pinus en México. Rev Mex Cien For 7:30–44
Ferreira ATB, Tomazello Filho M (2009) Caracterização dos anéis de crescimento de árvores de Pinus caribaea var. hondurensis Barr. et Golf. por densitometria de raios X. Sci For 37:287–298
Fonti P, García-González I (2004) Suitability of chestnut earlywood vessel chronologies for ecological studies. New Phytol 163:77–86. doi:10.1111/j.1469-8137.2004.01089.x
Fonti P, Solomonoff N, García-González I (2007) Earlywood vessels of Castanea sativa record temperature before their formation. New Phytol 173:562–570. doi:10.1111/j.1469-8137.2006.01945.x
Fonti P, von Arx G, Garcia-Gonzalez I, Eilmann B, Sass-Klaassen U, Gartner H, Eckstein D (2010) Studying global change through investigation of the plastic responses of xylem anatomy in tree rings. New Phytol 185:42–53. doi:10.1111/j.1469-8137.2009.03030.x
Fonti P, Bryukhanova M, Myglan V, Naumova O, Kirdyanov A, Vaganov E (2013) Temperature-induced responses of xylem structure of Larix sibirica (Pinaceae) from Russian Altay. Am J Bot 100:1332–1343. doi:10.3732/ajb.1200484
Fritts HC (1976) Tree rings and climate. Academic Press, New York
García-González I, Fonti P (2006) Selecting earlywood vessels to maximize their environmental signal. Tree Physiol 26:1289–1296. doi:10.1093/treephys/26.10.1289
Giantomasi MA, Junent FAR, Villagra PE, Srur AM (2009) Annual variation and influence of climate on the ring width and wood hydrosystem of Prosopis flexuosa DC trees using image analysis. Trees 23:117–126. doi:10.1007/s00468-008-0260-5
Gottwald H, Parameswaran N (1980) Anatomy of wood and bark of Tectona (Verbenaceae) in relation to taxonomy. Bot Jahrb Syst 101:363–384
Grissino-Mayer HD (2001) Evaluating crossdating accuracy: a manual and tutorial for the computer program COFECHA. Tree Ring Res 57:205–221
Holmes RL (1986) Quality control of crossdating and measuring: a user’s manual for program COFECHA. In: Holmes RL, Adams RK, Fritts HC (eds) Tree-ring chronologies of western North America: California, eastern Oregon and northern Great Basin. Tucson, Arizona, pp 41–49
Jacoby GC, D’Arrigo RD (1995) Tree ring width and density evidence of climatic and potential forest change in Alaska. Glob Biogeochem Cycles 9:227–234. doi:10.1029/95GB00321
Kunert N, Schwendenmann L, Hölscher D (2010) Seasonal dynamics of tree sap flux and water use in nine species in Panamanian forest plantations. Agric For Meteorol 150:411–419. doi:10.1016/j.agrformet.2010.01.006
Leoni JM, Fonseca SF, Schongart J (2011) Growth and population structure of the tree species Malouetia tamaquarina (Aubl.) (Apocynaceae) in the central Amazonian floodplain forests and their implication for management. For Ecol Manag 261:62–67. doi:10.1016/j.foreco.2010.09.025
Lilienfein J, Wilcke W, Ayarza MA, Vilela L, Lima SC, Zech W (2000) Soil acidification in Pinus caribaea forests on Brazilian savanna Oxisols. For Ecol Manag 128:145–157. doi:10.1016/S0378-1127(99)00143-7
Lindorf H (1994) Eco-anatomical wood features of species from a very dry tropical forest. IAWA J 15:361–376
Locosselli G, Buckeridge M, Moreira M, Ceccantini G (2013) A multi-proxy dendroecological analysis of two tropical species (Hymenaea spp., Leguminosae) growing in a vegetation mosaic. Trees 27:25–36. doi:10.1007/s00468-012-0764-x
Marchand N, Filion L (2012) False rings in the white pine (Pinus strobus) of the Outaouais Hills, Quebec (Canada), as indicators of water stress. Can J For Res 42:12–22. doi:10.1139/x11-151
Masiokas M, Villalba R (2004) Climatic significance of intra-annual bands in the wood of Nothofagus pumilio in southern Patagonia. Trees 18:696–704. doi:10.1007/s00468-004-0355-6
Mattos PP, Santos AT, Oliveira JM, Rosot MAD (2007) Dendrocronologia de espécies de Floresta Ombrófila Mista do munícipio de Candói, PR. Pesquisa Florestal 54:153–156
Miller A (2007) Fire history of Caribbean Pine (Pinus caribaea var. bahamensis (Griseb.) W.H. Barrett & Golfari) Forests on Abaco Island, the Bahamas. M.Sc. Thesis, University of Tennessee
O’Brien JJ, Hiers JK, Callaham MA, Mitchell RJ, Jack SB (2008) Interactions among overstory structure, seedling life-history traits, and fire in frequently burned neotropical pine forests. Ambio 37:542–547. doi:10.1579/0044-7447-37.7.542
Olajidel O, Ndaeyo NU, Ekongl AB (2010) Dendroclimatology of a Pinus caribaea plantation in the tropical rainforest area of Southweastern Nigeria. J Agric Biotechnol Ecol 3:92–97
Olano JM, Arzac A, Garcia-Cervigon AI, von Arx G, Rozas V (2013) New star on the stage: amount of ray parenchyma in tree rings shows a link to climate. New Phytol 198:486–495. doi:10.1111/nph.12113
Oliveira JM, Roig FA, Pillar VD (2010) Climatic signals in tree-rings of Araucaria angustifolia in the southern Brazilian highlands. Austral Ecol 35:134–147. doi:10.1111/j.1442-9993.2009.02018.x
Osborn TJ, Briffa KR, Jones PD (1997) Adjusting variance for sample-size in tree-ring chronologies and other regional mean time series. Dendrochronologia 15:89–99
Panayotov MP, Zafirov N, Cherubini P (2013) Fingerprints of extreme climate events in Pinus sylvestris tree rings from Bulgaria. Trees 27:211–227. doi:10.1007/s00468-012-0789-1
Pandey D, Brown C (2000) Teak: a global overview. Unasylva 51(201):3–13
Priya KM, Bhat KM (1998) False ring formation in teak (Tectona grandis L.f.) and the influence of environmental factors. For Ecol Manage 108:215–222. doi:10.1016/S0378-1127(98)00227-8
Priya KM, Bhat KM (1999) Influence of rainfall, irrigation and age on the growth periodicity and wood structure in teak (Tectona grandis). IAWA J 20:181–191
Pumijumnong N (1997) Cambium development of teak (Tectona grandis L.) in Thailand and its relationship to climate. Proceedings of the 97th international symposium on wood science and technology, wood-human-environment, October 23–24, 1997, Seoul, Korea, pp 61–72
Pumijumnong N, Wanyaphet T (2006) Seasonal cambial activity and tree-ring formation of Pinus merkusii and Pinus kesiya (Pinaceae) in Thailand in dependence on climate. For Ecol Manage 226:279–289. doi:10.1016/j.foreco.2006.01.040
Pumijumnong N, Eckstein D, Sass U (1995) Tree-ring research on Tectona grandis on northern Thailand. IAWA J 16:385–392
Rao KS, Rajput KS (1999) Seasonal behaviour of vascular cambium in teak (Tectona grandis L.f) growing in moist deciduous and dry deciduous forest of Gujarat. IAWA J 20:85–93
Robert EMR, Koedam N, Beeckman H, Schmitz N (2009) A safe hydraulic architecture as wood anatomical explanation for the difference in distribution of the mangroves Avicennia and Rhizophora. Funct Ecol 23:649–657. doi:10.1111/j.1365-2435.2009.01551.x
Rozas V, García-González I, Zas R (2011) Climatic control of intraannual wood density fluctuations of Pinus pinaster in NW Spain. Trees 25:443–453. doi:10.1007/s00468-010-0519-5
Rozendaal DMA, Zuidema PA (2011) Dendroecology in the tropics a review. Trees 25:3–16. doi:10.1007/s00468-010-0480-3
Schuhli GS, Paludzyszyn Filho E (2010) O cenário nacional da silvicultura de teca (Tectona grandis L.f.) e perspectivas de melhoramento. Pesqui Florest Bras 30(63):217–230. doi:10.4336/2010.pfb.30.63.217
Schweingruber FH (1988) Tree rings. Basics and applications of dendrochronology. Reidel, Dordrecht
Schweingruber FH (1996) Tree rings and environment: dendroecology. Berne, Switzerland
Shah SK, Bhattacharyya A, Chaudhary V (2007) Reconstruction of June–September precipitation based on tree-ring data of teak (Tectona grandis L.) from Hoshangabad, Madhya Pradesh. India. Dendrochronologia 25:57–64. doi:10.1016/j.dendro.2007.02.001
Schongart J, Junk WJ, Piedade MTF, Ayres JM, Huttermann A, Worbes M (2004) Teleconnection between tree growth in the Amazonian floodplains and the El Nino–Southern Oscillation effect. Glob Chang Biol 10:683–692
Singh ND, Venugopal N (2011) Cambial activity and annual rhythm of xylem production of Pinus kesiya Royle ex. Gordon (Pinaceae) in relation to phenology and climatic factors growing in sub-tropical wet forest of North East India. Flora 206:198–204. doi:10.1016/j.flora.2010.04.021
Soliz-Gamboa CC, Rozendaal DMA, Ceccantini G, Angyalossy V, van der Borg K, Zuidema PA (2011) Evaluating the annual nature of juvenile rings in Bolivian tropical rainforest trees. Trees 25:17–27. doi:10.1007/s00468-010-0468-z
Tomazello Filho M, Cardoso NS (1999) Seasonal variations of the vascular cambium of teak (Tectona grandis L.) in Brazil. In: Wimmer R, Vetter RE (eds) Tree-ring analysis: biological, methodological and environmental aspects. CAB International, London, pp 147–154
Tyree MT, Zimmermann MH (2002) Xylem structure and the ascent of sap. Springer Verlag, Berlin
Vieira J, Campelo F, Nabais C (2009) Age-dependent responses of tree-ring growth and intra-annual density fluctuations of Pinus pinaster to Mediterranean climate. Trees 23:257–265. doi:10.1007/s00468-008-0273-0
von Arx G, Archer SA, Hughes MK (2012) Long-term functional plasticity in plant hydraulic architecture in response to supplemental moisture. Ann Bot 109:1091–1100. doi:10.1093/aob/mcs030
von Arx G, Kueffer C, Fonti P (2013) Quantifying plasticity in vessel grouping—added value from the image analysis tool ROXAS. IAWA J 34:433–445. doi:10.1163/22941932-00000035
Wigley TML, Briffa KR, Jones D (1984) On the average value of correlated time series with application in dendroclimatology and hydrometereology. J Clim App Meteorol 23:201–221. doi:10.1175/1520-0450(1984)023<0201:OTAVOC>2.0.CO;2
Wimmer R (2002) Wood anatomical features in tree-rings as indicators of environmental change. Dendrochronologia 20:21–36. doi:10.1078/1125-7865-00005
Wimmer R, Strumia G, Holawe F (2000) Use of false rings in Austrian pine to reconstruct early growing season precipitation. Can J For Res 30:1691–1697. doi:10.1139/cjfr-30-11-1691
Worbes M (1995) How to measure growth dynamics in tropical trees, a review. IAWA J 16:337–351
Worbes M (1999) Annual growth rings, rainfall-dependent growth and long-term growth patterns of tropical trees from the Caparo Forest Reserve in Venezuela. J Ecol 87:391–403. doi:10.1046/j.1365-2745.1999.00361.x
Wyka TP, Oleksyn J, Zytkowiak R, Karolewski P, Jagodzinski AM, Reich PB (2012) Responses of leaf structure and photosynthetic properties to intra-canopy light gradients: a common garden test with four broadleaf deciduous angiosperm and seven evergreen conifer tree species. Oecologia 170:11–24. doi:10.1007/s00442-012-2279-y
Young PJ, Megonigal JP, Sharitz RR, Day FP (1993) False ring formation in bald cypress (Taxodium distichum) saplings under two flooding regimes. Wetlands 13:293–298. doi:10.1007/BF03161295
Zang C, Biondi F (2013) Dendroclimatic calibration in R: The bootRes package for response and correlation function analysis. Dendrochronologia 31(1):68–74. doi:10.1016/j.dendro.2012.08.001
Zeng ZZ, Piao SL, Chen AP, Lin X, Nan HJ, Li JS, Ciais P (2013) Committed changes in tropical tree cover under the projected 21st century climate change. Sci Rep 3:1951. doi:10.1038/srep01951
Zuidema PA, Brienen JW, Schöngart J (2012) Tropical forest warming: looking backwards for more insights. Trends Ecol Evol 27:193–194. doi:10.1016/j.tree.2011.12.007
Acknowledgments
We thank Priscila Amaral de Sá and MSc. Claudio Roberto Anholetto Júnior for field and laboratory support. We would also like to thank Dr. Ana Carolina Maioli Campos Barbosa and Dr. Clóvis Angeli Sansigolo for their contributions to this study, as well as the “National Counsel of Technological and Scientific Development” (CNPq) for the financial support (Project 561910/2010-3), its coordinator, Dr. Edson José Vidal da Silva, the National Commission for Scientific and Technological Research of Chile (CONICYT- PAI/INDUSTRIA 79090016) and Siria Rosales for English corrections and two anonymous reviewers for valuable feedback on and improvements to an earlier draft of this article.
Conflict of interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by S. Leavitt.
Rights and permissions
About this article

Cite this article
Venegas-González, A., von Arx, G., Chagas, M.P. et al. Plasticity in xylem anatomical traits of two tropical species in response to intra-seasonal climate variability. Trees 29, 423–435 (2015). https://doi.org/10.1007/s00468-014-1121-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-014-1121-z