
Abstract
Fucoxanthin and eicosapentaenoic acid (EPA) provide significant health benefits for human population. Diatom is a potential natural livestock for the combined production of EPA and fucoxanthin. In this study, first, the effects of three important parameters including light intensity, nitrogen concentration and salinity were evaluated for the production of EPA and fucoxanthin in two diatom strains Phaeodactylum tricornutum and Cylindrotheca fusiformis. And then, two steps method based on light intensity were applied to produce EPA and fucoxanthin in large scale. Higher light intensity was first adopted for the high growth rate and lipid content of diatom, and after a period of time, light intensity was lowered to enhance the accumulation of fucoxanthin and EPA. In final, the highest EPA yields were 62.55 and 27.32 mg L−1 for P. tricornutum and C. fusiformis, and the fucoxanthin yield reached 8.32 and 6.05 mg L−1, respectively.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
In recent years, the demand of fucoxanthin (a specific non-provitamin A carotenoid) as a nutraceutical is growing [1]. With an allenic carbon chain, an acyclic keto group, and a hydroxyl group at one of the β-ionone rings esterified with acetic acid [2], fucoxanthin possesses a wide range of remarkable biological properties, including anti-obesity, anti-diabetes, anti-cancer, anti-allergic, anti-inflammatory, anti-oxidation, and anti-osteoporotic activities [3,4,5].
Fucoxanthin is mainly naturally found in seaweeds and diatoms, where it is coupled to the thylakoid membrane to transfer excitation energy to the photosynthetic electron transport chain via chlorophyll a [6]. However, the fucoxanthin content ranges from 0.22 to 2.17% of dry weight in diatoms, much higher than that found in seaweeds [7]. Moreover, the marine microalgae diatoms can grow rapidly and perform robustly in controlled bioreactors, and are considered as the potential natural sources of fucoxanthin.
Besides fucoxanthin, diatoms are rich in lipid and some long-chain polyunsaturated fatty acids (LC-PUFA), such as eicosapentaenoic acid (EPA), which also provides significant health benefits in cardiovascular health in adults [8, 9]. In previous reports, the EPA content of diatom can reach up to 2.5 in 100 g dry weight cells (2.5%) at certain condition [10].
Growth and accumulation of fucoxanthin and EPA of diatoms may vary considerably across species and/or strains and are dependent on culture conditions such as nutrient concentrations and environmental factors [11, 12]. Nitrogen is a primary nutrient for microalgae growth and also plays an important role in regulating cellular metabolites including lipid and fucoxanthin accumulation [13]. Light is the primary source of energy that drives all biochemical processes in microalgae [6]. Moreover, salinity has previously been studied in several microalgal species and demonstrated that it can regulate some gene expression for FA synthesis [14], but the effect on fucoxanthin accumulation is still unknown.
In spite of being a promising source, there are only a limited number of reports about diatoms for the combined production of fucoxanthin and EPA. Moreover, the studies of fucoxanthin accumulation from diatoms were almost in bench scale in laboratory; the pilot scale of fucoxanthin and EPA production, to our knowledge, has not been done.
Phaeodactylum tricornutum and Cylindrotheca fusiformis are two advertised diatoms cultivated at a large scale for aquaculture and commercial lipid production. The main objective of the present study is to optimize several key parameters for improving EPA and fucoxanthin production of diatom under laboratory conditions, and examined the production yield of EPA and fucoxanthin in pilot scale. These results provide valuable implications into future exploration of diatom, especially P. tricornutum and C. fusiformis.
Materials and methods
Strains and culture conditions
Two diatom strains were investigated in this study. Strain P. tricornutum (UTEX 646) was obtained by the Culture Collection of Algae at The University of Texas at Austin, and strain C. fusiformis was kindly provided by Prof. Liang in Ocean University of China.
Tested strains were cultivated in 100 mL Erlenmeyer flask containing 30 mL of sterile f/2 medium. Cultures were incubated at 23 ± 1 °C under an illumination intensity of 30 µmol m−2 s−1 and were shaken manually twice a day.
Key culture conditions optimization in batch cultures
Three key parameters (nitrogen, light intensity and salinity) were optimized for improving EPA and fucoxanthin production of two diatom strains under laboratory conditions. In this section, strains were cultivated in 800 mL column photobioreactor containing 500 mL of sterile 2f (4 × f/2) enriched medium, and the experiments were carried out with constant aeration of 1% CO2 at 23 ± 1 °C room temperature.
For light intensity experiment, strains were grown in 2f enriched medium for about 8 days under different light intensities (30, 80, 120, 180 µmol photons m−2 s−1).
For nitrogen experiment, strains were grown in 2f enriched medium with different levels of NaNO3 supplementations (0-N, 0 mg L−1; f/2-N, 75 mg L−1; f-N, 150 mg L−1; 2f-N, 300 mg L−1) into stationary growth phase, and the experiments were carried out with 70 µmol photons m−2 s−1.
In salinity experiment, strains were grown in 2f enriched medium with different levels of salinity (5, 10, 20 and 30‰) under 70 µmol photons m−2 s−1 for 8 days.
Production of EPA and fucoxanthin in pilot scale
According to results of the above experiment, the two diatoms P. tricornutum and C. fusiformis were cultured in 2f-N enriched medium with 20‰ salinity. Higher light intensity (120 µmol photons m−2 s−1) was adopted for the high growth rate and lipid content, and then low light intensity (30 µmol photons m−2 s−1) was used to accumulate fucoxanthin for 3 days. For each strain, the experiment was carried out in hanging bag photobioreactor with a volume of 20 L at optimized light intensity with 1% CO2 at 23 ± 1 °C room temperature.
Determination of biomass concentration
A certain volume of microalgae culture was washed for twice with distilled water before being filtered on a pre-weighted Whatman GF/C filter. The filters containing the algae were dried in a vacuum oven at 105 °C overnight and subsequently cooled down to room temperature for weighing. The biomass concentration is expressed as cells dry weight per liter.
Analysis of lipid content and fatty acid profiles
The total lipid content was determined by gravimetric analysis according to modified Bligh and Dyer’s method [15]. Approximately, 50 mg dried algal pellet was ground with quart first and then mixed with 7.5 mL methanol/chloroform (2:1, v/v) at 37 °C overnight and then centrifuged. The supernatant was collected and residual biomass was extracted twice. The supernatants were combined, and chloroform and 1% sodium chloride solution were added to a final volume ratio of 1:1:0.9 (chloroform: methanol: water). The organic phase was carefully transferred to a vial and dried to constant weight with nitrogen flow. The lipid weight was the difference of the weights of vial with and without lipid, and the total lipid content was calculated as a percentage of the dry weight of the algae.
A 0.5 mg sample of lipid was dissolved in 1 mL heptane that contains 50 µg heptadecanoic acid methyl ester (C18H37 COOCH3) as internal standard for FAME analysis on a Varian 450GC (Varian Inc., USA) equipped with a flame ionization detector (FID) and Agilent HP-5 GC Capillary Column (30 m × 0.25 mm × 0.25 µm). Nitrogen was used as carrier gas. The injector temperature was set at 280 °C with an injection volume of 2 µL under splitmode (10:1). The detector temperature was set at 280 °C. The individual FAMEs were identified by chromatographic comparison with authentic standards (Sigma) and the quantities of individual FAMEs were calculated as reports by Chen et al. [16].
Pigments extraction and fucoxanthin analysis
For pigments extraction, 20 mg of lyophilized cell powder was grounded and then extracted with 2 mL cold methanol/acetone (1:1, v/v) under dim light. After shaking for 1 h, the whole solutions carrying grinded cells were centrifuged at 8000 r/min for 5 min at 4 °C. The supernatant was collected and the pellets were extracted for another time under the same conditions. All supernatants were combined and filtered through a 0.22 µm filter prior to HPLC analysis.
Fucoxanthin was detected using Waters 2695 HPLC systems (Waters, Milford, MA, USA) with a PDA detector and a C18 reverse phase bar (5 mm particle size, 250 mm × 4.6 mm ID). The mobile phase used was (A) 80% methanol, 20% 0.5 M ammonium acetate, (B) 90% acetonitrile, and (C) 100% ethyl acetate, with a flow rate of 0.8 mL min−1. The chromatogram was recorded at 449 nm. A purified fucoxanthin standard was used for quantification and identification of fucoxanthin by means of a calibration curve and comparison of retention time and ultraviolet–visible absorption spectra, respectively.
Statistical analysis
All cultivations were performed at lease in duplicate. All determinative data were collected from triplicate samples and the final values were expressed as mean value ± standard deviation. Statistical significance was evaluated by ANOVA and t test using SPSS programs (Version 19.0, IBM SPSS, Chicago, IL, USA) at a level of P < 0.05.
Results
Effect of light intensity on biomass, EPA and fucoxanthin accumulation in two diatoms
Results for dry weight, lipid and fucoxanthin contents of two diatom strains at an initial concentration of 0.15 g L−1 were shown in Fig. 1, and the fatty acid profiles were shown in Table 1. Cultures subjected to the lowest light intensity (30 µmol photons m−2 s−1) showed lower biomass concentrations than any higher light intensity tested for both P. tricornutum (Fig. 1a) and C. fusiformis (Fig. 1b). For P. tricornutum cultures, the similar growth rates were observed between 70 and 120 µmol photons m−2 s−1, although the highest biomass concentration reached the highest of 1.56 g L−1 under 70 µmol photons m−2 s−1 at day 8. For C. fusiformis cultures, a light intensity of 120 µmol photons m−2 s−1 resulted in the highest biomass concentration of 1.38 g L−1. However, at the highest light intensity of 180 µmol photons m−2 s−1, the cell grew slowly and the biomass concentrations were lower than 120 µmol photons m−2 s−1. Cultures that are light limited are expected to show an increase in biomass productivity upon increasing light intensities [17]. However, the results show that the biomass decreased when the cultures were exposed to 180 µmol photons m−2 s−1, this might be explained by photo inhibition that occurred. As soon as the photosynthetic machinery becomes inhibited, further increasing light intensity would not enhance the biomass concentration anymore [18]. Alma et al. [19] presented a light curve for P. tricornutum, and similar to our findings, they too reported that high light intensity decreases the growth of cells.

Effects of light intensity on biomass (a, b), lipid (c, d) and fucoxanthin accumulation (e, f) of P. tricornutum (a, c, e) and C. fusiformis (b, d, f)
From Fig. 1c, d, it could be noted that high light intensity enhanced the lipid accumulation of tested diatom strains. For two diatom strains, of all tested conditions, the cultures exposed to low light intensity (30 µmol photons m−2 s−1) reached the lowest lipid contents of 25.68 and 28.12%, respectively. Increasing the incident light intensity resulted in higher lipid content, and the highest lipid contents of P. tricornutum (41.48%) and C. fusiformis (44.82%) occurred in 180 and 120 µmol photons m−2 s−1, respectively. Table 1 illustrated the fatty acid composition of two diatoms strains cultured under different light intensities. The most abundant fatty acids in two diatom strains were found to be C16:1 (30.52–35.88%) and C16:0 (19.72–37.1%), that were in accordance with a previous study [20]. Besides, fractions of C20:4 (ARA) and C20:5 (EPA) were found in both two strains. As the increment of the light intensity, the content of EPA fraction (% of total fatty acids) reduced continuously, which is contrary to the change of lipid content. For P. tricornutum and C. fusiformis, the EPA content dropped from 17.15 to 13.83, 10.28 to 4.19% of the total fatty acids, respectively, from 30 to 180 µmol photons m−2 s−1. Literature reported that the culture exposed to high incident light intensity showed a decline in EPA content of fatty acid profiles [21]. Based on the lipid content of dry biomass and the EPA content of total fatty acids, the EPA content of dry biomass could be calculated. In final, with the tested light intensities, the EPA contents of biomass in P. tricornutum were 4.41, 4.77, 6.65 and 6.89%, respectively. While the EPA contents of biomass in C. fusiformis were 2.89, 2.53, 2.55 and 1.75%, much lower than those in P. tricornutum.
Contrary to cell growth and lipid accumulation, the highest values for fucoxanthin content in P. tricornutum and C. fusiformis were obtained under low light intensity (30 µmol photons m−2 s−1) (Fig. 1e, f). Under such conditions, 0.75 and 0.65% fucoxanthin (% of biomass) in P. tricornutum and C. fusiformis were produced on the 8th day of growth. Similar with the change in EPA content of fatty acid profiles, high light intensity reduced the biosynthesis of fucoxanthin. The fucoxanthin content of P. tricornutum and C. fusiformis dropped to 0.11 and 0.18%, respectively, under 180 µmol photons m−2 s−1. Guo et al. [7] reported that light intensity between 10 and 30 µmol photons m−2 s−1 promoted the fucoxanthin accumulation, while 40 µmol photons m−2 s−1 declined the fucoxanthin content, the conclusion of which was similar to ours. Thus, higher fucoxanthin production was obtained under low light intensities.
Effect of nitrogen concentration on biomass, EPA and fucoxanthin accumulation in two diatoms
Figure 2 illustrated the growth profile, lipid and fucoxanthin content of two diatom strains cultivated with various nitrogen concentrations in column photobioreactors. The cultures without nitrogen supply maintained relatively stable biomass concentrations, and the growth rates of two diatom strains were enhanced with an increase in higher nitrogen concentration (Fig. 2a, b). Consequently, the final biomass concentration reached 1.61 and 1.52 g L−1 on day 8 for the highest nitrogen concentration supplied cultures, 6.65- and 3.9-fold higher than those without nitrogen supply in P. tricornutum and C. fusiformis, respectively. This was in line with previous studies that high level of available nitrogen promoted biomass accumulation of diatoms as well as of other algae strains [22].

Effects of nitrogen concentration on biomass (a, b), lipid (c, d) and fucoxanthin accumulation (e, f) of P. tricornutum (a, c, e) and C. fusiformis (b, d, f)
Nitrogen concentration had a significantly negative effect on lipid accumulation. It was observed that P. tricornutum and C. fusiformis cultivated without nitrogen supply reached the highest lipid contents of 43.76 and 51.08%, respectively (Fig. 1c, d). As the increment of nitrogen concentration, the lipid contents of two diatom strains reduced remarkably. The highest concentration nitrogen-supplied cells of P. tricornutum and C. fusiformis declined to the lowest lipid content of 29.19 and 34.2%, respectively. Nitrogen is the most important element contributing to microalgal cells, its deprivation significantly changed the physiological and biochemical parameters. As protein is a nitrogen-rich compound, nitrogen deprivation can reduce protein content, extra energy flow would shift to carbohydrate and lipid metabolism [23].
The effect of nitrogen concentration on fatty acid compositions of two diatom strains was shown in Table 2. Similar to the above, the lipids of tested strains were primarily comprised of C16:0, C16:1 and C20:5. For P. tricornutum, the content of C16:0 had a slight increase from 20.72 to 24.4%, while there was a slight decrease from 34.35 to 31.77% in cells of C. fusiformis. However, in both strains, higher nitrogen concentration induced the lower content of C16:1. Noteworthy, as the increase of nitrogen supply, the contents of C20:5 (EPA) in P. tricornutum reached 16.83 from 13.65%, and that in C. fusiformis enhanced to 8.17 from 3.86%. Therefore, based on the data, nitrogen supply benefited the accumulation of EPA in total fatty acids in diatom. According to the lipid content and EPA ratio, the EPA contents of biomass in P. tricornutum were 5.97, 5.31, 5.05 and 4.91%, respectively, which were not significantly different. Moreover, the EPA contents of biomass in C. fusiformis were 1.97, 1.88, 2.41 and 2.79%, respectively. This finding revealed that the EPA production from P. tricornutum was better compared to that of C. fusiformis.
Nitrogen has an impact on both cell density and production of metabolites in microalgae. The further study on the impact of nitrogen concentration in fucoxanthin production was also investigated in this study (Fig. 2e, f). It was observed that the fucoxanthin contents with highest nitrogen concentration reached 0.54 and 0.61% at day 8, from 0.22 and 0.28% with nitrogen deprivation in P. tricornutum and C. fusiformis, respectively. Therefore, in the whole, nitrogen supply promoted the fucoxanthin biosynthesis in our study. The beneficial effect of nitrogen supply on fucoxanthin content and production was also observed in previous report [24]. The reason might be as analysis from Alipanah et al. [13]: nitrogen supply might upregulate the biosynthesis of chlorophyll and, consequently, promote fucoxanthin accumulation, as fucoxanthin is a core part of photosystem in diatoms.
Effect of salinity on biomass, EPA and fucoxanthin accumulation in two diatoms
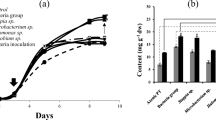
The biomass, lipid and fucoxanthin accumulation of two diatom strains cultivated at different salinities were compared (Fig. 3). The initial concentration in P. tricornutum was ca. 0.15 g L−1, which increased to 0.72, 1.12, 1.49 and 1.34 g L−1 at the salinity of 5, 10, 20 and 30‰, respectively (Fig. 3a). For C. fusiformis, the cultures grow slowly at the salinity of 5‰, while the other salinity had a significant effect on cell growth, as the biomass concentration reached 1.38, 1.53 and 1.64 g L−1 at the salinity of 10, 20 and 30‰ (Fig. 3b).

Effects of salinity on biomass (a, b), lipid (c, d) and fucoxanthin accumulation (e, f) of P. tricornutum (a, c, e) and C. fusiformis (b, d, f)
As illustrated in Fig. 3c, d, salinity showed different effects on lipid accumulation of two tested diatom strains. For P. tricornutum, quantitatively, the minimum lipid content of 33.88% was obtained at the salinity of 10‰, while the lipid content reached the maximum of 44.44% at the salinity of 20‰. This finding was different from the literature described by Ye et al. [25], in which the highest lipid content reached at the salinity of 30‰, higher and lower salinity decrease the lipid content. The reason might be that the strains of P. tricornutum they used were different and the their P. tricornutum cells were ultraviolet mutant. For C. fusiformis, except for the culture exposed to the salinity of 20‰, lipid accumulation has not significant difference for other salinity conditions. Specifically, the maximum lipid contents at the salinity of 20‰ were 43.99% C. fusiformis, ca. 12.5% higher than those of other salinity conditions. The fatty acid profiles of both diatoms strains with various salinities can be found in Table 3. As similar to lipid accumulation, the change of EPA ratio in total fatty acids differs from species to species. For P. tricornutum, the EPA content decreased to 12.54% from 16.46% as salinity increases, thus, low salinity benefits the EPA accumulation, which was in accordance with the previous report [25]. However, the EPA ratio in total fatty acids was maintained relatively stable, which was 5.94–6.87% of total fatty acids. Therefore, salinity had no significant effect on EPA accumulation in C. fusiformis. By contrast, C. fusiformis had a high percentage of C20:4 (5.96–9.94%), the ratio of which increased as salinity increased.
The salinities that rendered the fucoxanthin contents of the two diatom strains were shown in Fig. 3e, f. For P. tricornutum, the highest fucoxanthin content (0.74%) in P. tricornutum cultures was obtained at the salinity of 20‰, however, other salinities did not induce a significant effect on the fucoxanthin content (0.59–0.63%). However, for C. fusiformis, fucoxanthin accumulation also did not significantly differentiate for other salinity conditions except for the culture exposed to the salinity of 5‰. Under such conditions, 0.48% of fucoxanthin was produced on the 8th day of growth.
In conclusion, because salinity induced the change of biomass, lipid, EPA and fucoxanthin accumulation with different degrees, it is necessary to take into account the total EPA and fucoxanthin production of both microalgae species. According to the data given in Fig. 3 and Table 3, the highest EPA and fucoxanthin production were both obtained at the salinity of 20‰ for P. tricornutum. In addition, for C. fusiformis, the highest value for EPA production was also achieved at the salinity of 20‰. Although the maximum fucoxanthin production of C. fusiformis was 0.92 g L−1 at the salinity of 30‰, it was only 12% higher than that at the salinity of 20‰ (0.82 g L−1). Therefore, in total, among the tested salinities, 20‰ could be regarded as the ideal salinity for active substance production from diatom.
Production of EPA and fucoxanthin from two diatom strains in pilot scale
To assess EPA and fucoxanthin production scale-up feasibility, two diatom strains were cultured in a pilot scale hanging bag photobioreactor with a volume of 20 L in workshops. Based on the above result, 2f-N enriched medium with the salinity of 20‰ was adopted to cultivate the microalgal cells. In addition, it was demonstrated that high light intensity promoted the growth and lipid accumulation, while low light intensity benefited the fucoxanthin accumulation. Therefore, two steps method based on light intensity was applied to produce EPA and fucoxanthin from diatom. More concretely, higher light intensity (120 µmol photons m−2 s−1) was adopted for the high growth rate and lipid content, and then low light intensity (30 µmol photons m−2 s−1) was used to accumulate fucoxanthin for another 3 days. The concrete results were illustrated in Fig. 4.

The biomass (a), lipid content (b), EPA ratio (c), EPA yield (d), fucoxanthin content (e) and fucoxanthin yield (f) of P. tricornutum and C. fusiformis cultivated in pilot scale
Results showed the similar variation tendency of growth between two diatom strains, although the final biomass concentrations were different (Fig. 4a). The maximum biomass concentration of 1.3 g L−1 was obtained for P. tricornutum, which was higher than that of 1.14 g L−1 for C. fusiformis at day 18. As indicated in Fig. 4b, the lipid content of P. tricornutum showed an upward tendency with the extension of time, and that increased remarkably to 37.36 from 24.61% at day 12 from day 9. For C. fusiformis, the lipid content increased to 35.54% at day 15 from the initial of 22.49% under high light intensity, although it slightly decreased to 19.73% at day 3. It is worth mentioning that the lipid contents of two diatom strains still had slight increase even at low intensity from day 15 to day 18.
With the extension of time, EPA ratio of total fatty acids in two diatom strains exhibited irregular changes (Fig. 4c). EPA ratio of P. tricornutum increased to 11.18% at day 6, while a decline was observed at day 9 (9.52%), and then it reached its highest ratio (12.09%) at day 12. However, for C. fusiformis, in the initial stage of culture, EPA ratio decreased slightly from 5.32 to 4.66% at day 3, followed by a slow growing with the extension of culture time, and the final EPA ratio reached 6.39% at day 18. In spite of the irregular change of EPA ratio, EPA yields of two diatom strains increased continuously as the biomass concentration and lipid content improved (Fig. 4d). The EPA yield of P. tricornutum cultures reached 24.08 mg L−1 at day 9, followed by a sharp increase till 62.55 mg L−1 at day 18, 17.18-fold higher compared to the initial of 3.64 mg L−1. Moreover, for C. fusiformis, EPA yield showed a relatively linear increase tend from 2.19 to 27.32 mg L−1 during the culture time. However, the EPA yield of two diatom strains differed significantly, as EPA yield of P. tricornutum was higher than that in C. fusiformis.
Fucoxanthin content of P. tricornutum cultures decreased, as the culture enters the stationary phase during the first 15 days cultivation. As illustrated in Fig. 4e, fucoxanthin content in P. tricornutum cultures reached highest with 0.77% at growth phase, while there was a decline in stationary phase. Likewise, fucoxanthin content in C. fusiformis cultures was higher in the mid-linear growth phase (6 days) in comparison with the contents detected in stationary phase (15 days). In addition, when light intensity was lowered from day 15 to day 18, fucoxanthin contents in both strains improved at different degrees. The observation suggested that low light intensity led to a greater fucoxanthin accumulation in diatom. Besides, fucoxanthin yield of two diatom strains were calculated in terms of fucoxanthin content and biomass concentration and illustrated in Fig. 4f. Consequently, although fucoxanthin yield of P. tricornutum had slight decrease from day 12 to day 15, it still kept a stable increasing trend in both strains during the whole culture period. Consequently, fucoxanthin yield reached 8.32 and 6.05 mg L−1 from 0.99 and 0.68 mg L−1 in P. tricornutum and C. fusiformis, respectively.
Conclusions
With a range of light intensity, higher light intensity promoted the growth and lipid accumulation of two diatoms strains, but decreased the accumulation of fucoxanthin. Moreover, nitrogen supply benefited the accumulation of biomass concentration, EPA ratio and fucoxanthin accumulation. Among the tested salinities in the study, 20‰ was considered as the ideal salinity in terms of EPA and fucoxanthin yield. Finally, two steps method based on light intensity were applied to produce EPA and fucoxanthin from diatom, results showed that EPA and fucoxanthin yield of P. tricornutum were both higher than those of C. fusiformis.
References
Miyashita K, Nishikawa S, Beppu F, Tsukui T, Abe M, Hosokawa M (2011) The allenic carotenoid fucoxanthin, a novel marine nutraceutical from brown seaweeds. J Sci Food Agric 91:1166–1174
Ulrike E, Alexandros B, Jürgen B, Claudia B, Gerhard S (2016) Limitations in the biosynthesis of fucoxanthin as targets for genetic engineerng in Phaeodactylum tricornutum. J Appl Phycol 28:123–129
Maeda H, Hosokawa M, Sashima T, Murakami-Funayama K, Miyashita K (2009) Anti-obesity and anti-diabetic effects fucoxanthin on diet-induced obesity conditions in a marine model. Mol Med Rep 2:897–902
Tanaka T, Shnimizu M, Moriwaki H (2012) Cancer chemoprevention by carotenoids. Molecules 17:3202–3242
Yoshioka H, Ishida M, Nishi K, Oda H, Toyohara H, Sugahara T (2014) Studies on anti-allergic activity of Sargassum horneri extract. J Funct Foods 10:154–160
Mulder KJM, Lamers PP, Martens DE, Wijffels RH (2014) Phototrophic pigment production with microalgae: biological constraints and opportunities. J Phycol 50:229–242
Guo BB, Liu B, Yang B, Sun PP, Lu X, Liu J, Chen F (2016) Screening of diatom strains and characterization of Cyclotella cryptica as a potential fucoxanthin producer. Mar Drugs 14:125
Lee JH, O’Keefe JH, Lavie CJ, Harris WS (2009) Omega-3 fatty acids: cardiovascular benefits, sources and sustainability. Nat Rev Cardiol 6:753–758
Gharami K, Das M, Das S (2015) Essential role of docosahexaenoic acid towards development of a smarter brain. Neurochem Int 89:51–62
Kim SM, Kang SW, Kwon ON, Chung D, Pan CH (2012) Fucoxanthin as a major carotenoid in Isochrysis aff. galbana: characterization of extraction for commercial application. J Korean Soc Appl Biol Chem 55:477–483
Li Y, Horsman M, Wang B, Wu N, Lan CQ (2008) Effects of nitrogen sources on cell growth and lipid accumulation of green alga Neochloris oleoabundans. Appl Microbiol Biotechnol 81:629–636
Pahl SL, Lewis DM, King KD, Chen F (2012) Heterotrophic growth and nutritional aspects of the diatom Cyclotella cryptica (Bacillariophyceae): effect of nitrogen source and concentration. J Appl Phycol 24:301–307
Alipanah L, Rohloff J, Winge P, Bones AM, Brembu T (2015) Whole-cell response to nitrogen deprivation in the diatom Phaeodactylum tricornutum. J Exp Bot 66:6281–6296
Hernandez ML, Padilla MN, Sicardo MD, Mancha M, Martinez-Rivas JM (2011) Effect of different environmental stresses on the expression of oleate desaturase genes and fatty acid composition in olive fruit. Phytochemistry 72:178–187
Bligh EG, Dyer WJ (1959) A rapid method of total lipid extraction and purification. Can J Biochem Physiol 37:911–917
Chen L, Liu TZ, Zhang W, Chen XL, Wang JF (2012) Biodiesel production from algae oil high in free fatty acids by two-step catalytic conversion. Bioresour Technol 111:208–214
Breuer G, Lamers PP, Martens DE, Draaisma RB, Wijffesl RH (2013) Effect of light intensity, pH, and temperature on triacylglycerol (TAG) accumulation induced by nitrogen starvation in Scenedesmus obliquus. Bioresour Technol 143:1–9
Dillschneider R, Steinweg C, Rosello-Sastre R, Posten C (2013) Biofuels from microalgae: photoconversion efficiency during lipid accumulation. Bioresour Technol 142:647–654
Alma GL, Jorge B, Marco RP (2016) Growth kinetics and fucoxanthin production of Phaeodactylum tricornutum and Isochrysis galbana cultures at different light and agitation conditions. J Appl Phycol 28:849–860
Wu HL, Li T, Wang GH, Dai SK, He H, Xiang WZ (2016) A comparative analysis of fatty acid composition and fucoxanthin content in six Phaeodactylum tricornutum strains from different origins. Chin J Oceanol Limnol 34(2):391–398
Remmers IM, Martens DE, Wijffels RH, Lamers PP (2017) Dynamics of triacylglycerol and EPA production in Phaeodactylum tricornutum under nitrogen starvation at different light intensities. Plos One 12(4):e0175630
Xiao X, Si X, Yuan Z, Xu X, Li G (2012) Isolation of fucoxanthin from edible brown algae by microwave-assisted extraction coupled with high-speed countercurrent chromatography. J Sep Sci 35:2313–2317
Jiang Y, Yoshida T, Ouigg A (2012) Photosynthetic performance, lipid production and biomass composition in response to nitrogen limitation in marine microalgae. Plant Physiol Biochem 54:70–77
Xia S, Wang K, Wan LL, Li A, Hu Q, Zhang CW (2013) Production, characterization, and antioxidant activity of fucoxanthin from the marine diatom Odontella aurita. Mar Drugs 11:2667–2681
Ye L, Jiang XM, Mao XX, Gao XZ, Zhang ZL (2015) Effects of temperature, light intensity and salinity on the growth, total lipid and fatty acid of Phaeodactylum tricornutum mutant. Chin J Ecol 34(2):454–462
Acknowledgements
This work was supported by Shandong Provincial Natural Science Foundation, China (ZR2017QC007) and Marine economic innovation and development regional demonstration project of Qingdao.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Wang, H., Zhang, Y., Chen, L. et al. Combined production of fucoxanthin and EPA from two diatom strains Phaeodactylum tricornutum and Cylindrotheca fusiformis cultures. Bioprocess Biosyst Eng 41, 1061–1071 (2018). https://doi.org/10.1007/s00449-018-1935-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00449-018-1935-y