
Abstract
Irruptive or boom-and-bust population dynamics, also known as ‘outbreaks’, are an important phenomenon that has been noted in biological invasions at least since Charles Elton’s classic book was published in 1958. Community-level consequences of irruptive dynamics are poorly documented and invasive species provide excellent systems for their study. African Jewelfish (Rubricatochromis letourneuxi, “jewelfish”) are omnivores that demonstrate opportunistic carnivory, first reported in Florida in the 1960s and in Everglades National Park (ENP) in 2000. Twelve years after invasion in ENP, jewelfish underwent a 25-fold increase in density in one year. By 2016, jewelfish represented 25–50% of fish biomass. Using a 43-year fish community dataset at two sites (1978–2021), and a 25-year dataset of fish and invertebrate communities from the same drainage (1996–2021), with additional spatial coverage, we quantified differences in fish and invertebrate communities during different phases of invasion. During jewelfish boom, abundant, native cyprinodontiform fishes decreased in density and drove changes in community structure as measured by similarity of relativized abundance. Density of two species declined by > 70%, while four declined by 50–62%. Following the jewelfish bust, some species recovered to pre-boom densities while others did not. Diversity of recovery times produced altered community structure that lagged for at least four years after the jewelfish population declined. Community structure is an index of ecological functions such as resilience, productivity, and species interaction webs; therefore, these results demonstrate that irruptive population dynamics can alter ecological functions of ecosystems mediated by community structure for years following that population’s decline.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Irruptive, or boom-and-bust, population dynamics are an ecological phenomenon that has been observed in a variety of taxa with implications for ecosystem function, conservation, and human health (Pimentel 1961; Myers 1998; McCann et al. 2000; Ma 2020; López-Mañas et al. 2022). Irruptive population dynamics are common for invasive species and provide an opportunity to increase our understanding of population outbreaks in general (Elton 1958). This opportunity stems from the observation that biological invasions and associated ecological theory are extensions or special cases of general ecological hypotheses (Jeschke 2014; Daly et al. 2023). A fundamental question for advancing our understanding of outbreaks and resilience is whether impacts from an invasive boom lag or extend after the bust (Elton 1958; Simberloff and Gibbons 2004; Aagaard and Lockwood 2016).
Population declines of invasive species without human intervention are poorly studied and infrequently documented (but see Szydlowski et al. 2023), which has led to a lack of understanding about the persistence of community impacts from pulsed invasion dynamics (Simberloff and Gibbons 2004; Aagaard and Lockwood 2016). Declines are sometimes the result of a self-induced negative feedback from effects of invasive populations on recipient ecosystems, such as depletion of resources or altered habitat (Tang et al. 2012; Lester and Gruber 2016; Vuorinen et al. 2021), but little is known about post-disturbance community resilience of native communities and ecosystems (Carpenter et al. 2001; Strayer et al. 2017). For managers to make informed decisions about control, eradication, and/or active restoration, a better understanding of the mechanisms and impacts of natural population declines of invasive species is needed (Simberloff and Gibbons 2004).
Non-native and invasive species that persist at low densities—“sleeper populations”, either from the onset of their invasion or following a natural population decline, may eventually undergo irruptive population growth as part of a boom-bust cycle extending their impacts (Strayer et al. 2017; Spear et al. 2021; Vuorinen et al. 2021). In some instances, effects of the invasive species persist beyond their declines when impacts are severe (e.g., extirpation of native species), there has been a shift between alternative stable states, or in the presence of additional invasions (Weber and Brown 2009; Hansen et al. 2013; Strayer et al. 2017). For example, after 33 years of declines of rusty crayfish (Faxonius rusticus), macrophyte abundance and richness recovered to levels present in low-crayfish references lakes. While snail abundance and richness recovered, their approach to pre-invasion abundance lagged behind macrophyte recovery and failed to reach levels of reference lakes (Szydlowski et al. 2023). Rusty crayfish effects on snail abundance and richness persisted after the bust because snails depend on macrophytes for habitat (Szydlowski et al. 2023). Conversely, zebra mussels (Dreissena spp.) in the Hudson River estuary undergo repeated boom-bust cycles that do not correlate with zooplankton biomass as one might expect (Pace et al. 2010). Effects of invasive species do not always correspond to their population size. The relationship between invasive species density and impact may be nonlinear because of shifts in traits in the invasive population, or because effects can be time-lagged, hysteretic (i.e., a new stable state has been reached), or irreversible (Yokomizo et al. 2009; Pace et al. 2010; Dostál et al. 2013; Strayer et al. 2017).
The greater Everglades ecosystem is undergoing the largest ecological restoration effort in history, with total costs expected to exceed $20 billion (Sklar et al. 2005). However, 17 non-native fishes have been found in the freshwaters of ENP, compared to 39 native species (Loftus 2000; Kline et al. 2013). Most of these non-native fishes persist at low densities (Trexler et al. 2002), but there are notable exceptions (Harrison et al. 2013, Pintar et al. 2023a, b). Until recently, planning for Everglades restoration has overlooked non-native and invasive fishes (National Academies of Sciences 2014), and lack of data has sometimes been conflated with lack of effects (Schofield and Loftus 2015). Everglades restoration aims to restore historic populations of iconic predators (i.e., wading birds and alligators) by implementing water-management policies that increase production of their prey (Trexler and Goss 2009). Invasive fishes may undermine these goals (Pintar et al. 2023a, b).
A particular invasive species of concern is the African Jewelfish, (Hemichromis letourneuxi, proposed placement in Rubricatochromis by Lamboj and Koblmüller (2022); hereafter “jewelfish”). Jewelfish are a mid-trophic level freshwater fish that arrived in Florida in the 1960s and invaded Everglades National Park (ENP) in 2000 (Kline et al. 2013). In mesocosms and in temporary seasonal refuges, jewelfish had deleterious effects on native fishes and invertebrates (Rehage et al. 2014; Schofield et al. 2014). Jewelfish existed at low densities in ENP as a sleeper population until rapidly increasing (> fivefold) in density after 2012 (ESM 1.3).
After adjusting for variable hydrodynamics of the Everglades, Pintar et al. (2023a) used data from 1996 to 2021 to demonstrate significant density reductions of four common native fishes associated with the rapid increase in jewelfish density (“boom”) from 2012 to 2017. Additionally, Pintar et al. (2023a) found that jewelfish density declined by 2018 (“bust”), and afterwards, modeled native fishes recovered to expected population density based on hydrologic models. Community structure of the full suite of fishes and invertebrates was not analyzed by Pintar et al. (2023a) though community structure based on species relative abundances have been appreciated in ecology for many decades (MacArthur 1960; Tokeshi 1993), and effects at a community scale might differ from previously documented population-level recovery.
In this study, we explored the effects of jewelfish boom-bust dynamics on native communities of fishes and macroinvertebrates in ENP. We incorporate a hydrologic covariate in statistical models at multiple spatial and temporal scales to control for the dynamic nature of the study system (e.g., Trexler et al. 2005). We used two long-term data sets: one of only fishes that was continuous from 1978 through 2021 (43 years), and a shorter time series from 1996 through 2021 (25 years) with greater spatial coverage that included both fishes and aquatic macroinvertebrates. These data presented a rare opportunity to compare communities in the pre-invasion period, post-invasion/pre-boom (low density—sleeper population) period, boom period, and bust period. They allowed us to search for both phase-dependency of impacts and recovery rate (resilience) of the native assemblages (Blossey 1999; Strayer et al. 2006; Strayer 2012). We predicted that (1) during the beginning of the jewelfish invasion prior to the boom, community structure would not deviate from patterns of historic variation, (2) that the jewelfish boom would alter fish and macroinvertebrate communities from low density and pre-invasion structure, (3) that after the jewelfish bust, fish and macroinvertebrate community structure would return to pre-jewelfish boom communities corresponding with recovery of individual populations (Pintar et al. 2023a), and (4) that time since hydrologic disturbance will play a larger role in community structure than biotic effects (i.e., jewelfish boom-bust dynamics), as shown by previous studies in this ecosystem (Trexler et al. 2005; Pintar et al. 2023a, b). We hypothesized that specific species that might decline from jewelfish effects would include native sunfishes, thought to be competitors with cichlids (Montaña and Winemiller 2013), and native cyprinodontiform fishes, Riverine grass shrimp (Palaemonetes paludosus), and odonate larvae, thought to be prey items of jewelfish based on studies in their native range and the Everglades (Hickley and Bailey 1987; Rehage et al. 2014; Schofield et al. 2014).
Methods
Study sites and sampling design
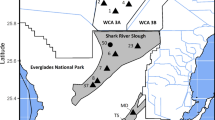
Monitoring sites were established in Everglades National Park in 1978 and expanded to a greater spatial coverage in 1996 to monitor responses of aquatic animals to changes in water management (Fig. 1; Loftus and Eklund 1994; Trexler et al. 2003). From July 1996 through December 2021 (25 years), fishes and invertebrates were collected using a 1-m2, 2-mm mesh throw trap (Loftus et al. 1990; Jordan et al. 1997). In each sampling period (five per year: February, April, July, October, and December), seven throw-trap samples were taken during daylight hours at three plots (A, B, C) at sites (21 throws per site) in the two major drainages of ENP, Shark River Slough (SRS) and Taylor Slough (TS), and in Water Conservation Areas 3A and 3B (Trexler et al. 2005). Fishes < 8 cm standard length (SL) were preserved, while larger fishes were identified, SL measured, and released. While plots were at fixed locations, throw-trap samples within a plot were taken at positions determined from a random number table (Wolski et al. 2004). After 1996, 367 samples were typically collected for each of the sampling periods (throws x plots x sites) and 1835 were taken annually; fewer samples were taken during drought years when some dry plots were unsamplable. We focus our analyses on sites in SRS where the jewelfish population demonstrated boom-bust dynamics (Pintar et al. 2023a). At two SRS sites (06 and 23), sampling began in 1978. However, from 1978 to 1985 sampling occurred monthly at one plot at each site, and the number of throw traps performed was determined by estimates of inter-sample variance (Kushlan 1974; Trexler et al. 2005). For inter-period comparisons, our analyses were performed using mean species density (individuals per m2) at each site per sampling period.

Map of long-term aquatic-animal monitoring sites in Shark River Slough, Everglades National Park (grey polygon). Additional sites were sampled but not included in this study. Each site (labeled on map) has multiple plots. At sites 6 and 23, plot A has been sampled continuously since 1978
Data analysis
To compare community structure through time, we first plotted jewelfish density and biomass (g ash-free dry mass (AFDM)/m2) at each site from 1996 through 2021 to delineate the jewelfish boom and bust. Biomass was calculated using length–weight relationships and standard conversions (Electronic Supplementary Material, ESM 1.5). Data prior to 1996 were not used here because jewelfish invaded ENP in 2000. We analyzed differences in fish-community structure from 1978 through 2021 and invertebrate-community structure from 1996 through 2021. Fish communities were analyzed at sites 06 and 23 throughout the entire time series, while fish (ESM 1.2) and invertebrate communities at all sites in Shark River Slough were analyzed from 1996 through 2021. Data were partitioned into time periods based on two factors: different hydrological regimes during the pre-invasion period that could influence community assemblage (ESM 1.1) and phases of invasion. There are five pre-invasion time periods (Pre-invasion 1 through 5) and three post-invasion time periods (low density (2000–2011), boom (2012–2017), and bust (2018–2021)). The five pre-invasion time periods facilitate quantifying variation in community structure based on changes in hydrology. Community assemblages were visualized with non-metric multidimensional scaling (NMDS) and compared among time periods using permutational multivariate analysis of variance (PERMANOVA) with Morisita-Horn dissimilarities (Jost et al. 2011). PERMANOVA models included both invasion status (“Status” in tables and figures) and a hydrologic measure of disturbance, DSD (days since dry, a measure of time since disturbance, where dry is defined as water depth < 5 cm). NMDS and PERMANOVA were repeated using Bray–Curtis dissimilarities for comparison (ESM). We use similarity percentages (SIMPER) analysis to identify species contributing to the top 95% of dissimilarity between phases of invasion. NMDS, PERMANOVA, and SIMPER were conducted in R using the ‘vegan’ package (Oksanen et al. 2022; R Core Team 2022) with and without jewelfish included in the community (results were nearly identical, so we report only those without jewelfish to focus on changes in native species). Post hoc pairwise PERMANOVAs were performed to determine which time periods were different from one another using pairwise.perm.manova function from the ‘RVAidMemoire’ package in R (Hervé 2022).
Results
At long-term monitoring sites in SRS, jewelfish demonstrated boom-bust dynamics over time. Jewelfish density and biomass rapidly increased starting in 2012 (boom) but substantially declined (bust) by 2018 (Fig. 2). PERMANOVA demonstrated that invasion status (Pseudo-F7 = 12.8, R2 = 0.22, p < 0.001), time since disturbance (DSD: Pseudo-F1 = 12.8, R2 = 0.14, p < 0.001), and their interaction (Pseudo-F7 = 6.8, R2 = 0.07, p < 0.001) had significant effects on fish-community structure. The significant interaction showed that the fish-community response to hydrologic-disturbance varied among phases of invasion. Pairwise PERMANOVAs indicated that fish communities during boom (2012–2017) and bust (2018–2021) were distinct from all other time periods (back to 1978), but not from one another (Fig. 3, Fig. S1, Table 1). Despite the decline in jewelfish density and biomass (Fig. 1), the fish community did not return to the jewelfish low density (2000–2011) nor any pre-invasion structure. The fish community from the jewelfish boom compared to that immediately before the jewelfish boom (low density) was characterized by notably fewer of the dominant cyprinodontiform fishes such as Eastern Mosquitofish (Gambusia holbrooki, change in mean density = -61%), Least Killifish (Heterandria formosa, -56%), Lake Chubsucker (Erimyzon succetta, -76%), Sailfin Molly (Poecilia latipinna, -70%), Golden Topminnow—(Fundulus chrysotus, -36%), and Flagfish (Jordanella floridae, -51%), and two small centrarchid species—Everglades Pygmy Sunfish (Elassoma evergladei, -52%) and Bluespotted Sunfish (Enneacanthus gloriosus, -17%; Figs. 4, 5, Table S3). A larger centrarchid, Spotted Sunfish (Lepomis punctatus), increased during the jewelfish boom (86%). Relative to before the jewelfish boom, declines of mean density of these species were maintained at similar or larger magnitudes after the jewelfish bust except for Bluespotted Sunfish, which increased after jewelfish declined (Table S3, Figs. S2–S4). Differences in community structure at a larger spatial but shorter temporal scale (all sites in SRS from 1996 to 2021), yielded similar results except for Everglades Pygmy Sunfish, which steadily increased in density during each phase of invasion (Fig. S5, Table S4).

Jewelfish relative abundance (individuals per m2) (A) and relative biomass (g AFDM per m2) (B) from 1996 through 2021. Vertical dashed lines represent jewelfish invasion of ENP in 2000 (black), boom phase of the invasion from 2012 through the end of 2017 (red)

NMDS (stress = 0.16, k = 2) of fish communities from different phases of the jewelfish invasion (Status) from 1978 through 2021 (see Electronic Supplemental Material for definitions of and rationale for each level of Status) at sites 06 and 23 in SRS. Pre-jewelfish invasion, fish communities oscillated through time along an axis from bottom left to top right. Severe drought in 1989–1990 caused the community to become more variable and move towards the top right. Post-jewelfish invasion, fish communities orthogonally diverged from the pre-invasion pattern (excluding that severe drought) towards the top left with nearly complete overlap between “Boom” and “Bust” communities. Ellipses represent 95% confidence intervals. Species names are abbreviated below illustrations as first three letters of the genus followed by first three letters of the species (Table S1). Jewelfish were omitted from analyses, but when included appeared in the top-left corner. The ten species with the highest mean density over the entire time series are plotted in NMDS space

Differences in average density of fishes between low density and jewelfish boom phases of invasion that explained 95% of the variance between these time periods. Species are ordered left to right from most to least abundant throughout the entire dataset and listed by common name (Table S1). Error bars represent two standard deviations. Pairwise comparisons for all time periods are in Fig. S4

Average density of eight fish species and total fish density at sites 06 and 23 during different water management regimes and phases of jewelfish invasion (Status) that explained the most community dissimilarity. Species are listed in order from highest (top left) to lowest (bottom right) mean density throughout the time series. Eastern Mosquitofish, Everglades Pygmy Sunfish, Golden Topminnow, Least Killifish, and Sailfin Molly have decreased in density since the jewelfish invasion and did not increase during the bust. Note that y-axis scales are different for each panel. Error bars represent upper bounds of two standard deviations and lower bounds were near zero for all species
Invertebrate communities during and after the jewelfish boom differed from pre-invasion and pre-boom invertebrate communities, but not from one another. PERMANOVA showed that invasion status (Pseudo-F3 = 24.2, R2 = 0.040, p < 0.001), DSD (Pseudo-F1 = 104, R2 = 0.056, p < 0.001), and their interaction (Pseudo-F3 = 2.68, R2 = 0.004, p = 0.023) were significant with DSD and invasion status explaining similar amounts of variation (Table 2A). However, the model explained relatively little of the overall variation in the data. Pairwise PERMANOVAs indicated that the only invertebrate communities that did not differ from one another were boom and bust communities (p = 0.061; Table 2B). This lack of difference was marginal (95% confidence ellipses barely overlapped); however, boom and bust communities are closer to each other than other groups in ordination space (Fig. 6). Relative to years of low densities of jewelfish beforehand, invertebrate communities during the jewelfish boom were characterized by decreases in mean density of pennant (Celithemus spp., -17%) and skimmer (Libellula spp., -24%) dragonfly larvae and planorbid snails (Planorbella spp., -65%). Each of those taxa underwent increases in mean density from the MDW period (1996–1999) to the low-density period (2000–2011). In contrast, other taxa increased during the jewelfish boom relative to low density, such as creeping water bugs (Pelocoris femoratus, 47%), giant water bugs (Belostoma spp., 77%), damselfly larvae (Coenagrionidae, 47%), beetle larvae (Coleoptera, 95%), and mayfly larvae (Ephemeroptera, 343%) (Fig. 7, Table S5).

NMDS (stress = 0.12, k = 2) of aquatic macroinvertebrate communities from different phases of the jewelfish invasion (Status) from 1996 through 2021 (see Supplemental Information for definitions of and rationale for each level of Status) across all sites in SRS. Ellipses represent 95% confidence intervals. The only two communities that have any ellipse overlap and did not statistically differ from each other were “boom” and “bust” (although this was marginal with p = 0.06, Table 2). Without the same amount of historic data that the fish community had at sites 06 and 23, it was impossible to determine long-term trends (prior to 1996) that provide context to pre- and post-invasion patterns. Taxa are abbreviated by either the first six letters of taxonomic group for groups not identified to species, or first three letters of genus and first three letters of species (Table S1)

Differences in average density of aquatic macroinvertebrate taxa between the low density and boom phases of jewelfish invasion during different phases of jewelfish invasion that explain 95% of the variance per comparisons. Taxa are ordered from most to least abundant throughout the entire dataset. Grass shrimp and creeping water bug were omitted because those taxa were several times more abundant than the next most abundant taxa. As a result, they also explained the largest portion of variance among time periods (Fig. S10). Pairwise comparisons between all time periods are in Fig. S10. Error bars represent two standard deviations
Discussion
Irruptive dynamics of a trophic generalist (e.g., jewelfish) were linked to a range of multi-directional impacts on community assembly that persisted after jewelfish population decline. These effects may be attributed to frequent disturbance, hysteresis, priority effects, and/or strength of effects even at low densities. Contrary to our hypothesis, hypothesized biotic effects from jewelfish explained more variance in community structure than time since hydrologic disturbance, which is commonly a strong parameter in analyses of dynamics of this community (e.g., Ruetz et al. 2005; Trexler et al. 2005; Banet and Trexler 2013). For the fish community, altered structure was observed at multiple spatial and temporal scales: two sites over a 43-year period and an additional four sites over a 25-year period. Differences in fish-community structure were driven by declines in some of the most abundant fishes (e.g., Eastern Mosquitofish, Least Killifish, Golden Topminnow, and Flagfish) that are also abundant species in diets of other fishes and wading birds (Klassen et al. 2016; Flood et al. 2023). Changes in invertebrate-community structure included declines in dragonfly larvae and planorbid snails along with increases in some predatory taxa like creeping water bugs and giant water bugs. These multispecies dynamics may have consequences for relative amounts of autotrophic versus heterotrophic energy flow (planorbid snails) or predation pressure (creeping water bugs and giant water bugs). Community structure is an index for ecological functions such as resilience, productivity, and species interaction networks (Mayfield et al. 2023). Therefore, these results demonstrate that irruptive population dynamics can be linked to disrupted ecological function of ecosystems for years following that population’s decline.
Several native species recovered after the jewelfish bust relative to predicted densities modeled based on hydrologic covariates (Pintar et al. 2023a), while a lack of community-wide recovery was observed based on relative abundances in this study. This result may have important implications for assessing invasive species and evaluating ecological theory. Impacts from invasive species are notoriously difficult to evaluate (Parker et al. 1999; Simberloff et al. 2013). One contributing factor is that many studies have information for one dimension of an invasive species’ potential impact(s), such as the effect of that invasive species on density of one or several native taxa, which can misrepresent the net effects of invasive species on recipient ecosystems at other levels of ecological organization (Flood et al. 2020; Crystal-Ornelas and Lockwood 2020). In the case of jewelfish in ENP, if study of this invasion had stopped at recovered densities of several native species (Pintar et al. 2023a), community-level effects would have gone undocumented, with potential implications for resources devoted to invasive-species management. This scenario is limited to neither jewelfish in ENP nor invasion biology; based on our results, we suggest that relative abundance should be considered in tandem with total abundance (e.g., density or biomass) for a more comprehensive understanding of both effects of invasive species and ecological theory across multiple ecological scales (i.e., populations—total abundance, communities—relative abundances, and ecosystems).
Altered community structure measured by relative abundance for both fishes and invertebrates persisted even after the jewelfish bust. Similar results were observed for invasive cane toads (Rhinella marina) in Australia, where the native community did not recover following cane toad declines (Brown and Shine 2019). There are several possible mechanisms for community recovery to lag population recovery (Pintar et al. 2023a): frequent disturbance (drying) may delay community assembly after droughts, community changes may require more energy or effort to reverse than was required to cause those changes (i.e., they are hysteretic), even at low densities jewelfish may exert negative effects on native species, and/or jewelfish invasion reset priority effects and interaction webs that maintain post-jewelfish boom relative abundances (Drake 1991). Priority effects usually benefit invasive species more than native species, and native species pay a higher cost for arriving late (Dickson et al. 2012; Stuble and Souza 2016; Weidlich et al. 2021). In mesocosms, experiments have demonstrated initial conditions can create alternate food-web stable states (Chase 2003). These studies suggest that altered initial conditions post-invasion reset priority effects and may contribute to changes at the community level and thus food-web structure (Vander Zanden et al. 2006). This idea is akin to the trophic cascade in the Greater Yellowstone Ecosystem that resulted from reintroduction of wolves. Wolf reintroduction created an alternate food-web stable state through altered herbivore behavior driving increases in riparian tree recruitment and improved bird nesting sites, among other effects at multiple trophic levels (Osborne and Kovacic 1993; Berger et al. 2001; Ripple et al. 2001). Additional research is needed to understand how jewelfish have altered energetic and dynamics linkages within this food web.
Even at low jewelfish densities, fish and invertebrate communities did not converge on pre-boom communities. The density threshold at which invasive species effects are detectable may be high relative to the densities where they impact interaction webs (Yokomizo et al. 2009; Parkos et al. 2019). For example, effects of Peacock Bass (Cichla monoculus) introduction to Lake Gatun observed by Zaret and Paine (1973) were the result of Peacock Bass densities of < 0.05 fish/m2. Native fish communities in Lake Gatun had not recovered 45 years later, despite Peacock Bass densities remaining relatively low (Sharpe et al. 2017). While the effects of jewelfish documented here are not as severe as effects of Peacock Bass demonstrated by Zaret and Paine (1973), our results emphasize that recovery of native taxa in the presence of a low-density invasive species is not guaranteed (Brown and Shine 2019).
We found support for our hypothesis that centrarchids would decrease in density post-jewelfish invasion, possibly because of competition that results from niche overlap (Montaña and Winemiller 2013) and similar body size. Smaller sunfish species (i.e., Bluespotted and Pygmy sunfishes) decreased in density during the jewelfish boom, while Spotted Sunfish, with a larger terminal size than jewelfish, increased in density. If exploitative competition was the primary driver of altered community structure, theory predicts that generalist species would persist (Holt et al. 1999). Our results regarding sunfishes are consistent with this prediction. In addition to being smaller, Bluespotted and Pygmy Sunfishes are also more specialized in their feeding, with diets consisting almost entirely of omnivorous invertebrates, while Spotted Sunfish are generalists that eat detritus, primary producers, and a variety of invertebrates and small fishes (Flood et al. 2023). Invasive species that displace native biota, such as jewelfish in this study, are predicted to not only be superior resource exploiters, but also exert strong interference effects on native fauna (Amarasekare 2002). Subordinate species often undergo niche contraction in the presence of a dominant competitor (Pianka 1974; Case and Gilpin 1974). Further study is required to determine how niches of native sunfishes have responded to jewelfish invasion to better understand relative importance of different competitive interactions between invasive species and native analogs.
Many native species that declined in density after jewelfish invasion are prey taxa based on previous work in the Everglades and their native range (Hickley and Bailey 1987; Rehage et al. 2014), suggesting that jewelfish are exerting top-down effects on native fauna. Despite the comparatively small size of jewelfish, the observed declines in native fauna are like those observed from invasive predators such as salmonids, centrarchids, and lake trout (Crowl et al. 1992; Vander Zanden et al. 1999; Koel et al. 2019). Each of those invasions has been responsible for trophic cascades and/or ecosystem-level impacts (Simon and Townsend 2003; Cucherousset and Olden 2011; Wainright et al. 2021). It remains to be seen if jewelfish have had similar effects in the Everglades. Nonetheless, these results support the idea that mid-trophic-level consumers can exert dramatic effects on ecosystems by having a range of trophic positions, rapid response to environmental change, intraguild predation, and relatively high numeric abundance (Taylor et al. 2001; Stewart et al. 2017). The central location of these taxa in the food web leads to multi-directional effects that can have consequences for ecosystem functions and services (Flood et al. 2020).
Small fishes that declined during jewelfish invasion are important diet items for wading birds (Boyle et al. 2012; Klassen et al. 2016). Snowy Egrets (Egretta thula) and Tricolored Herons (Egretta tricolor) rely heavily on Sailfin Molly and topminnows (Fundulus spp.), which declined by 70 and 36%, respectively, after the jewelfish boom (Boyle et al. 2012; Klassen et al. 2016). White Ibis (Eudocimus albus) prefer crayfish but switch to piscivory when fishes become seasonally concentrated (Kushlan 1979; Dorn et al. 2011; Boyle et al. 2012). Little Blue Heron (Egretta caerulea) diets from 2012 to 2014 (during the jewelfish boom) had jewelfish as the most abundant diet item, followed by Spotted Sunfish and Dollar Sunfish (Lepomis marginatus) (Klassen et al. 2016). Quantitative wading-bird diets prior to jewelfish invasion do not exist for many species (Klassen et al. 2016), so it is unclear whether dietary changes by wading birds have occured because of the jewelfish boom. An important element of Everglades restoration is to facilitate irruptive dynamics of breeding wading birds and the consistent return of large breeding colonies in ENP (Frederick et al. 2009; National Academies of Sciences Engineering and Medicine 2021). Declines in native-fish populations and altered fish and invertebrate community structure because of invasive fishes such as jewelfish may have negative effects on wading-bird breeding success despite tremendous effort and resources being devoted to hydrologic restoration.
At present, jewelfish represent a “sleeper” population, as they were during the first decade of their invasion in ENP, persisting at low abundances with potential to rapidly increase in density if triggered by disturbance or environmental change (Spear et al. 2021). Such low abundance populations of non-native species are often overlooked. However, low-abundance populations present an opportunity for more efficient and effective management actions when each individual represents a higher proportion of the population. Not attempting to remove potentially invasive species when they are at low abundance risks the population rebounding (Aagaard and Lockwood 2016). In the Everglades, frequent disturbance (Trexler et al. 2005) and additional invasive species expanding their range (Pintar et al. 2023b) elevate the risk of a sleeper population undergoing rapid population growth (Spear et al. 2021). Given the globally increased potential for anthropogenic disturbance and directional environmental change, coupled with the accelerating spread of non-native species, documenting and understanding impacts of irruptive or boom-and-bust population dynamics and associated sleeper populations, are critical for managing not just non-native and invasive species, but also native populations and communities that undergo outbreaks (Seebens et al. 2017; Strayer et al. 2017; Ratajczak et al. 2018; Spear et al. 2021; Pimentel 1961). In the case of jewelfish, even after population decline there remains ecological damage measured at multiple spatial and temporal scales with unknown consequences for ecosystem functions and services.
Data and code availability
Data and R code are archived at the Florida Coastal Everglades LTER website: https://doi.org/10.6073/pasta/91ccf52d9b3c4530736775f7923eb71f
References
Aagaard K, Lockwood JL (2016) Severe and rapid population declines in exotic birds. Biol Invasions 18:1667–1678. https://doi.org/10.1007/s10530-016-1109-2
Amarasekare P (2002) Interference competition and species coexistence. Proc R Soc B: Biol Sci 269:2541–2550. https://doi.org/10.1098/rspb.2002.2181
Banet AI, Trexler JC (2013) Space-for-time substitution works in Everglades ecological forecasting models. PLoS ONE 8(11):e81025
Berger J, Stacey PB, Bellis L, Johnson MP (2001) A mammalian predator-prey imbalance: grizzly bear and wolf extinction affect avian neotropical migrants. Ecol Appl 11:947–960. https://doi.org/10.1890/1051-0761(2001)011[0947:AMPPIG]2.0.CO;2
Blossey B (1999) Before, during and after: the need for long-term monitoring in invasive plant species management. Biol Invasions 1:301–311
Boyle RA, Dorn NJ, Cook MI (2012) Nestling diet of three sympatrically nesting wading bird species in the Florida Everglades. Waterbirds 35:154–159. https://doi.org/10.1675/063.035.0116
Brown GP, Shine R (2019) Using a natural population collapse of an invasive species to assess the benefits of invader control for native species. Biol Invasions 21:2781–2788. https://doi.org/10.1007/s10530-019-02015-8
Carpenter S, Walker B, Anderies JM, Abel N (2001) From metaphor to measurement: resilience of what to what? In: Ecosystems. pp 765–781. https://doi.org/10.1007/s10021-001-0045-9
Case TJ, Gilpin ME (1974) Interference Competition and Niche Theory (population dynamics/adaptive strategies/resource gradients). Proceedings of the National Academy of Sciences USA 71:3073–3077. https://doi.org/10.1073/pnas.71.8.3073
Chase JM (2003) Experimental evidence for alternative stable equilibria in a benthic pond food web. Ecol Lett 6:733–741. https://doi.org/10.1046/j.1461-0248.2003.00482.x
Crowl TA, Townsend CR, Mcintosh AR (1992) The impact of introduced brown and rainbow trout on native fish: the case of Australasia. Rev Fish Biol Fish 2:217–241. https://doi.org/10.1007/BF00045038
Crystal-Ornelas R, Lockwood JL (2020) The ‘known unknowns’ of invasive species impact measurement. Biol Invasions 22:1513–1525. https://doi.org/10.1007/s10530-020-02200-0
Cucherousset J, Olden JD (2011) Ecological impacts of non-native freshwater fishes. Fisheries (bethesda) 36:215–230. https://doi.org/10.1080/03632415.2011.574578
Daly EZ, Chabrerie O, Massol F et al (2023) A synthesis of biological invasion hypotheses associated with the introduction–naturalisation–invasion continuum. Oikos 5:e09645. https://doi.org/10.1111/oik.09645
Dickson TL, Hopwood JL, Wilsey BJ (2012) Do priority effects benefit invasive plants more than native plants? An experiment with six grassland species. Biol Invasions 14:2617–2624. https://doi.org/10.1007/s10530-012-0257-2
Dorn NJ, Cook MI, Herring G et al (2011) Aquatic prey switching and urban foraging by the White Ibis Eudocimus albus are determined by wetland hydrological conditions. Ibis 153:323–335. https://doi.org/10.1111/j.1474-919X.2011.01101.x
Dostál P, Müllerová J, Pyšek P et al (2013) The impact of an invasive plant changes over time. Ecol Lett 16:1277–1284. https://doi.org/10.1111/ele.12166
Drake JA (1991) Community-assembly mechanics and the structure of an experimental species ensemble. Am Nat 137:1–26
Elton CS (1958) The ecology of invasions by animals and plants. Methuen, London
Flood PJ, Duran A, Barton M et al (2020) Invasion impacts on functions and services of aquatic ecosystems. Hydrobiologia 847:1571–1586. https://doi.org/10.1007/s10750-020-04211-3
Flood PJ, Loftus WF, Trexler JC (2023) Fishes in a seasonally pulsed wetland show spatiotemporal shifts in diet and trophic niche but not shifts in trophic position. Food Webs 34:e00265. https://doi.org/10.1016/j.fooweb.2022.e00265
Frederick P, Gawlik DE, Ogden JC et al (2009) The white ibis and wood stork as indicators for restoration of the Everglades ecosystem. Ecol Indic 9:S83–S95. https://doi.org/10.1016/j.ecolind.2008.10.012
Hansen GJA, Vander Zanden MJ, Blum MJ et al (2013) Commonly rare and rarely common: comparing population abundance of invasive and native aquatic species. PLoS ONE 8:e77415. https://doi.org/10.1371/journal.pone.0077415
Harrison E, Lorenz JJ, Trexler JC (2013) Per capita effects of non-native mayan cichlids (cichlasoma urophthalmus; gunther) on native fish in the. Copeia 2013:80–96. https://doi.org/10.1643/CE-11-182
Hervé M (2022) RVAidMemoire: testing and plotting procedures for biostatistics. R package version 0.9-83
Hickley P, Bailey RG (1987) Food and feeding relationships of fish in the Sudd swamps (River Nile, southern Sudan). J Fish Biol 30:147–159. https://doi.org/10.1111/j.1095-8649.1987.tb05741.x
Holt RD, Lawton JH, Polis GA, Martinez ND (1999) Trophic rank and the species-area relationship. Ecology 80:1495–1504. https://doi.org/10.1890/0012-9658(1999)080[1495:TRATSA]2.0.CO;2
Jeschke JM (2014) General hypotheses in invasion ecology. Divers Distrib 20:1229–1234. https://doi.org/10.1111/ddi.12258
Jordan F, Coyne S, Trexler JC (1997) Sampling fishes in vegetated habitats: effects of habitat structure on sampling characteristics of the 1-m 2 throw trap. Trans Am Fish Soc 126:1012–1020. https://doi.org/10.1577/1548-8659(1997)126%3c1012:sfivhe%3e2.3.co;2
Jost L, Chao A, Chazdon RL (2011) Compositional similarity and beta diversity. In: Magurran AE, McGill BJ (eds) Biological diversity. Frontiers in measurement and assessment. Oxford University Press (OUP), Oxford, UK, pp 66–84
Klassen JA, Gawlik DE, Frederick PC (2016) Linking wading bird prey selection to number of nests. J Wildl Manag 80:1450–1460. https://doi.org/10.1002/jwmg.21141
Kline JL, Loftus WF, Kotun K et al (2013) Recent fish introductions into Everglades National Park: an unforeseen consequence of water management? Wetlands 34:S175–S187. https://doi.org/10.1007/s13157-012-0362-0
Koel TM, Tronstad LM, Arnold JL et al (2019) Predatory fish invasion induces within and across ecosystem effects in Yellowstone National Park. Sci Adv 5:eaav1139. https://doi.org/10.1126/sciadv.aav1139
Kushlan JA (1974) Quantitative sampling of fish populations in shallow, freshwater environments. Trans Am Fish Soc 103:348–352. https://doi.org/10.1577/1548-8659(1974)103%3c348:qsofpi%3e2.0.co;2
Kushlan JA (1979) Feeding ecology and prey selection in the white ibis. Condor 81:376–389. https://doi.org/10.2307/1366963
Lamboj A, Koblmüller S (2022) Molecular phylogeny and taxonomic revision of the cichlid genus Hemichromis (Teleostei, Cichliformes, Cichlidae), with description of a new genus and revalidation of H. angolensis. Hydrobiologia 850:2177–2195. https://doi.org/10.1007/s10750-022-05060-y
Lester PJ, Gruber MAM (2016) Booms, busts and population collapses in invasive ants. Biol Invasions 18:3091–3101. https://doi.org/10.1007/s10530-016-1214-2
Loftus WF (2000) Inventory of fishes of Everglades National Park. Biol Sci 63:27–47
Loftus WF, Chapman JD, Conrow R (1990) Hydroperiod effects on Everglades marsh food webs, with relation to marsh restoration efforts. In: Fisheries and Coastal Wetlands Research. Proceedings of the 1986 Conference on Science in National Parks, US NPS and The George Wright Society, pp 1–22
Loftus WF, Eklund AM (1994) Long-term dynamics of an Everglades small-fish assemblage. In: Davis SM, Ogden JC (eds) Everglades: the ecosystem and its restoration. St. Lucie Press, Delray Beach, FL, pp 461–483
López-Mañas R, Pascual-Díaz JP, García-Berro A et al (2022) Erratic spatiotemporal vegetation growth anomalies drive population outbreaks in a trans-Saharan insect migrant. Proc Natl Acad Sci USA 119:e2121249119. https://doi.org/10.1073/pnas.2121249119
Ma Z (2020) Predicting the outbreak risks and inflection points of COVID-19 pandemic with classic ecological theories. Adv Sci. https://doi.org/10.1002/advs.202001530
MacArthur R (1960) On the relative abundance of species. Am Nat 94:25–36
Mayfield MM, Lau JA, Tobias JA et al (2023) What can evolutionary history tell us about the functioning of ecological communities? ASN Presidential Debate Am Nat 202:587–603. https://doi.org/10.1086/726336
McCann K, Hastings A, Harrison S, Wilson W (2000) Population outbreaks in a discrete world. Theor Popul Biol 57:97–108. https://doi.org/10.1006/tpbi.1999.1444
Montaña CG, Winemiller KO (2013) Evolutionary convergence in Neotropical cichlids and Nearctic centrarchids: evidence from morphology, diet, and stable isotope analysis. Biol J Lin Soc 109:146–164. https://doi.org/10.1111/bij.12021
Myers JH (1998) Synchrony in outbreaks of forest Lepidoptera: a possible example of the Moran effect. Ecology 79:1111–1117. https://doi.org/10.1890/0012-9658(1998)079[1111:SIOOFL]2.0.CO;2
National Academies of Sciences E and M (2014) Progress toward restoring the everglades: the fifth biennial review, 2014. National Academies Press. Washington D.C
National Academies of Sciences E and M (2021) Progress toward restoring the Everglades: the eighth biennial review - 2020. National Academies Press, Washington, D.C
Oksanen J, Simpson G, Blanchet F, Kindt R, Legendre P, Minchin P, O'Hara R, Solymos P, Stevens M, Szoecs E, Wagner H, Barbour M, Bedward M, Bolker B, Borcard D, Carvalho G, Chirico M, De Caceres M, Durand S, Evangelista H, FitzJohn R, Friendly M, Furneaux B, Hannigan G, Hill M, Lahti L, McGlinn D, Ouellette M, Ribeiro Cunha E, Smith T, Stier A, Ter Braak C, Weedon J (2022) Vegan: community ecology package version 2.6-4
Osborne LL, Kovacic DA (1993) Riparian vegetated buffer strips in water-quality restoration and stream management. Freshw Biol 29:243–258. https://doi.org/10.1111/j.1365-2427.1993.tb00761.x
Pace ML, Strayer DL, Fischer D, Malcom HM (2010) Recovery of native zooplankton associated with increased mortality of an invasive mussel. Ecosphere 1:1–21. https://doi.org/10.1890/ES10-00002.1
Parker IM, Simberloff D, Lonsdale WM et al (1999) Impact: toward a framework for understanding the ecological effects of invaders. Biol Inv 22:1515–1525
Parkos JJ, Kline JL, Trexler JC (2019) Signal from the noise: model-based interpretation of variable correspondence between active and passive samplers. Ecosphere 10:e02858. https://doi.org/10.1002/ecs2.2858
Pianka ER (1974) Niche Overlap and Diffuse Competition (desert lizards/resource partitioning/community structure/species diversity). Proc Natl Acad Sci USA 71:2141–2145
Pimentel D (1961) Species diversity and insect population outbreaks. Ann Entomol Soc Am 54:76–86. https://doi.org/10.1093/aesa/54.1.76
Pintar MR, Dorn NJ, Kline JL, Trexler JC (2023a) Contrasting invasion histories and effects of three non-native fishes observed with long-term monitoring data. Biol Invasions. https://doi.org/10.1007/s10530-023-03146-9
Pintar MR, Dorn NJ, Kline JL, Trexler JC (2023b) Hydrology-mediated ecological function of a large wetland threatened by an invasive predator. Sci Total Environ 857:159245. https://doi.org/10.1016/j.scitotenv.2022.159245
R Core Team (2022) R: a language and enivronment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL https://www.R-project.org/
Ratajczak Z, Carpenter SR, Ives AR et al (2018) Abrupt change in ecological systems: inference and diagnosis. Trends Ecol Evol 33:513–526. https://doi.org/10.1016/j.tree.2018.04.013
Rehage JS, Liston SE, Dunker KJ, Loftus WF (2014) Fish community responses to the combined effects of decreased hydroperiod and nonnative fish invasions in a Karst wetland: are Everglades solution holes sinks for native fishes? Wetlands 34:S159–S173. https://doi.org/10.1007/s13157-012-0361-1
Ripple WJ, Larsen EJ, Renkin RA, Smith DW (2001) Trophic cascades among wolves, elk and aspen on Yellowstone National Park’s northern range. Biol Conserv 102:227–234. https://doi.org/10.1016/S0006-3207(01)00107-0
Ruetz CR, Trexler JC, Jordan F et al (2005) Population dynamics of wetland fishes: spatio-temporal patterns synchronized by hydrological disturbance? J Anim Ecol 74:322–332. https://doi.org/10.1111/j.1365-2656.2005.00926.x
Schofield PJ, Loftus WF (2015) Non-native fishes in Florida freshwaters: a literature review and synthesis. Rev Fish Biol Fish 25:117–145. https://doi.org/10.1007/s11160-014-9373-7
Schofield PJ, Slone DH, Gregoire DR, Loftus WF (2014) Effects of a non-native cichlid fish (African jewelfish, Hemichromis letourneuxi Sauvage 1880) on a simulated Everglades aquatic community. Hydrobiologia 722:171–182. https://doi.org/10.1007/s10750-013-1697-0
Seebens H, Blackburn TM, Dyer EE et al (2017) No saturation in the accumulation of alien species worldwide. Nat Commun 8:1–9. https://doi.org/10.1038/ncomms14435
Sharpe DMT, De León LF, González R, Torchin ME (2017) Tropical fish community does not recover 45 years after predator introduction. Ecology 98:412–424. https://doi.org/10.1002/ecy.1648
Simberloff D, Gibbons L (2004) Now you see them, now you don’t!-population crashes of established introduced species. Biol Invasions 6:161–172. https://doi.org/10.1023/B:BINV.0000022133.49752.46
Simberloff D, Martin JL, Genovesi P et al (2013) Impacts of biological invasions: what’s what and the way forward. Trends Ecol Evol 28:58–66. https://doi.org/10.1016/j.tree.2012.07.013
Simon KS, Townsend CR (2003) Impacts of freshwater invaders at different levels of ecological organisation, with emphasis on salmonids and ecosystem consequences. Freshw Biol 48:982–994. https://doi.org/10.1046/j.1365-2427.2003.01069.x
Sklar FH, Chimney MJ, Newman S et al (2005) The ecological - Societal underpinnings of Everglades restoration. Front Ecol Environ 3:161–169. https://doi.org/10.1890/1540-9295(2005)003[0161:TEUOER]2.0.CO;2
Spear MJ, Walsh JR, Ricciardi A, Vander Zanden MJ (2021) The invasion ecology of sleeper populations: prevalence, persistence, and abrupt shifts. Bioscience 71:357–369. https://doi.org/10.1093/biosci/biaa168
Stewart SD, Hamilton DP, Baisden WT et al (2017) Variable littoral-pelagic coupling as a food-web response to seasonal changes in pelagic primary production. Freshw Biol 62:2008–2025. https://doi.org/10.1111/fwb.13046
Strayer DL (2012) Eight questions about invasions and ecosystem functioning. Ecol Lett 15:1199–1210. https://doi.org/10.1111/j.1461-0248.2012.01817.x
Strayer DL, D’Antonio CM, Essl F et al (2017) Boom-bust dynamics in biological invasions: towards an improved application of the concept. Ecol Lett 20:1337–1350. https://doi.org/10.1111/ele.12822
Strayer DL, Eviner VT, Jeschke JM, Pace ML (2006) Understanding the long-term effects of species invasions. Trends Ecol Evol 21:645–651
Stuble KL, Souza L (2016) Priority effects: natives, but not exotics, pay to arrive late. J Ecol 104:987–993. https://doi.org/10.1111/1365-2745.12583
Szydlowski DK, Elgin AK, Lodge DM et al (2023) Long-term macrophyte and snail community responses to population declines of invasive rusty crayfish (Faxonius rusticus). Ecol Appl. https://doi.org/10.1002/eap.2818
Tang L, Gao Y, Wang C-H et al (2012) A plant invader declines through its modification to habitats: a case study of a 16-year chronosequence of Spartina alterniflora invasion in a salt marsh. Ecol Eng 49:181–185. https://doi.org/10.1016/j.ecoleng.2012.08.024
Taylor RC, Trexler JC, Loftus WF (2001) Separating the effects of intra- and interspecific age-structured interactions in an experimental fish assemblage. Oecologia 127:143–152. https://doi.org/10.1007/s004420000575
Tokeshi M (1993) Species abundance patterns and community structure. Advances in ecological research. Academic Press, Cambridge, pp 111–186
Trexler JC, Goss CW (2009) Aquatic fauna as indicators for Everglades restoration: applying dynamic targets in assessments. Ecol Indic 9:S108–S119. https://doi.org/10.1016/j.ecolind.2008.11.001
Trexler JC, Loftus WF, Jordan F, et al (2002) Ecological Scale and Its Implications for Freshwater Fishes in the Florida Everglades. In: Porter JW, Porter KG (eds) The Everglades, Florida Bay, and Coral Reefs of the Florida Keys: an ecosystem sourcebook. CRC Press, Boca Raton, FL, pp 153–184
Trexler JC, Loftus WF, Chick J (2003) Setting and monitoring restoration goals in the absence of historical data: the case of fishes in the Florida Everglades. In: Busch D, Trexler JC (eds) Monitoring Ecoregional Initiatives: Interdisciplinary Approaches for Determining Status and Trends of Ecosystem. Island Press, Washington, D.C., pp 351–376
Trexler JC, Loftus WF, Perry S (2005) Disturbance frequency and community structure in a twenty-five year intervention study. Oecologia 145:140–152. https://doi.org/10.1007/s00442-005
Vander Zanden MJ, Casselman JM, Rasmussen JB (1999) Stable isotope evidence for the food web consequences of species invasions in lakes. Nature 401:464–467. https://doi.org/10.1038/46762
Vander Zanden MJ, Olden JD, Gratton C (2006) Food-web approaches in restoration ecology. In: Falk DA, Zedler JB, Palmer MA (eds) Foundations of restoration ecology. Island Press, pp 165–189
Vuorinen KEM, Oksanen T, Oksanen L et al (2021) Why don’t all species overexploit? Oikos 130:1835–1848. https://doi.org/10.1111/oik.08358
Wainright CA, Muhlfeld CC, Elser JJ et al (2021) Species invasion progressively disrupts the trophic structure of native food webs. Proc Natl Acad Sci USA 118:e2102179118. https://doi.org/10.1073/pnas.2102179118
Weber MJ, Brown ML (2009) Effects of common carp on aquatic ecosystems 80 years after “Carp as a Dominant”: ecological insights for fisheries management. Rev Fish Sci 17:524–537. https://doi.org/10.1080/10641260903189243
Weidlich EWA, Nelson CR, Maron JL et al (2021) Priority effects and ecological restoration. Restor Ecol 29:e13317. https://doi.org/10.1111/rec.13317
Wolski LF, Trexler JC, Nelson EB, et al (2004) Assessing researcher impacts from a long-term sampling program of wetland communities in the Everglades National Park, Florida, U.S.A. Freshw Biol 49:1381–1390. https://doi.org/10.1111/j.1365-2427.2004.01256.x
Yokomizo H, Possingham HP, Thomas MB, Buckley YM (2009) Managing the impact of invasive species: the value of knowing the density-impact curve. Ecol Appl 19:376–386. https://doi.org/10.1890/08-0442.1
Zaret TM, Paine RT (1973) Species introduction in a tropical lake. Science 182:449–455. https://doi.org/10.1126/science.182.4111.449
Acknowledgements
We acknowledge the numerous technicians, graduate students, and postdocs who have worked to collect, process, and maintain these datasets over the past several decades. We would also like to specifically acknowledge the role played by David Sikkema, NPS (retired), for supporting establishment and continuation of this long-term monitoring program. This is publication number #1728 from the Institute of Environment at Florida International University.
Funding
PJF and JCT were supported by Cooperative Agreement P18AC01074 between Everglades National Park (ENP) and Florida International University (FIU) while working on this project. PJF was supported by the Everglades Foundation as an FIU ForEverglades Scholar. Data collection was supported by the Modified Water Deliveries program through a series of task agreements between ENP and FIU culminating in P21AC10856 under the South Florida and Caribbean Cooperative Ecosystems Studies Unit cooperative agreement P18AC00974, the Critical Ecosystem Studies Initiative and ENP base funding. This material is based upon work supported by the National Science Foundation (NSF) under grants no. HRD-1547798 and HRD-2111661. These NSF grants were awarded to FIU as part of the Centers of Research Excellence in Science and Technology (CREST) Program. This manuscript was developed in collaboration with the Florida Coastal Everglades Long-Term Ecological Research Program under NSF grants no. DEB-1237517 and DEB-2025954.
Author information
Authors and Affiliations
Contributions
JCT and WFL managed the project and data collection. PJF analyzed the data with input from JCT. PJF led manuscript writing and all authors contributed to revisions.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no relevant financial or non-financial interests to disclose.
Ethical approval
Florida International University’s IACUC committee most recently approved this work under #IACUC-22-047.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Additional information
Communicated by Catherine Price.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Flood, P.J., Loftus, W.F. & Trexler, J.C. Do community changes persist after irruptive population dynamics? A case study from an invasive species boom and bust. Oecologia 205, 445–459 (2024). https://doi.org/10.1007/s00442-024-05582-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-024-05582-3