
Abstract
The ability of non-native species to successfully invade new ecosystems sometimes involves evolutionary processes such as hybridization. Hybridization can produce individuals with superior traits that give them a competitive advantage over their parent species, allowing for rapid spread. Here we assess growth, functional morphology, and species interactions between two non-native beachgrass species (Ammophila arenaria and A. breviligulata) and their recently discovered hybrid (A. arenaria × A. breviligulata) on the U.S. Pacific Northwest coast. We asked whether the hybrid beachgrass differs from its parent species in morphology and growth, whether it competes with its parent species, and, if so, what are the potential mechanisms of competition. Plant taxa were grown in low- and high-density monocultures and in two-way interactions in a common garden environment. We show that the hybrid grew taller and more densely, with greater total biomass, than either parent species. The hybrid was also the better competitor, resulting in the model prediction of competitive exclusion against A. breviligulata and, depending on its relative abundance, A. arenaria. The hybrid displays a mixed ‘guerilla–phalanx’ growth form that allows it to spread laterally and achieve high shoot densities, giving it a competitive advantage. Given the current dominance of A. breviligulata compared to A. arenaria in most of the region where these taxa co-occur, we suggest that the hybrid will grow, compete, and spread quickly with potentially widespread consequences for the two non-native Ammophila congeners and the dunes they build.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The ability of non-native species to successfully invade new ecosystems sometimes involves strong competitive interactions, especially with closely related species (Pyšek and Richardson 2007; Theoharides and Dukes 2007; Kuester et al. 2014). Hybridization is one mechanism by which non-native species have been shown to evolve a competitive advantage over other species to become successful invaders (Stebbins 1985; Abbott 1992; Ellstrand and Schierenbeck 2000; Vilà et al. 2000; Lee 2002; Schierenbeck and Ellstrand 2009). Hybridization can produce unique genetic combinations and greater genetic diversity, both of which may be beneficial in novel or highly variable environments (Anderson and Stebbins 1954; Stebbins 1959; Rieseberg et al. 1999; Ellstrand and Schierenbeck 2000; Hovick et al. 2012).
One possible result of hybridization is hybrid vigor, which is the tendency of first-generation hybrids (F1) to have higher growth rates and biomass than their parent species. Some well-studied invasive plant hybrids, including cordgrass (Spartina anglica) and knotweed (Fallopia x bohemica), are first-generation crosses that display strong hybrid vigor and successfully outcompete their parent species (Ainouche et al. 2004; Comai 2005; Parepa et al. 2014). Other examples of non-native hybrids are reviewed in Hovick and Whitney (2014), a meta-analysis that considered observational and experimental studies to link hybridization with invasion success. Their analysis showed that invasive hybrids tend to be larger and more fecund than their parents and have traits that facilitate their dominance and spread. However, there was significant variation among studies depending on factors, such as the focal parent species used for comparisons, the generation class of the hybrid (F1 versus later generations), the phenotypic traits measured, and the environmental context. Thus, establishing a causal relationship between hybridization and invasion success has been challenging and few studies have tested hybrid performance under controlled experimental conditions (Hovick and Whitney 2014).
In this study, we use a common garden experiment to examine whether a newly discovered beachgrass hybrid (Ammophila arenaria × A. breviligulata) has the potential to grow faster, compete with its parent species, and eventually dominate U.S. Pacific Northwest dune ecosystems. The hybrid beachgrass was first discovered in 2012 and was morphologically, cytologically, and genetically verified in 2021 (Mostow et al. 2021). Recent sampling of hybrid individuals shows that a hybrid swarm has developed with F1 and F2 individuals and one case of backcrossing with a parent species (Mostow 2022). The hybrid is the product of a cross between two non-native beachgrasses, Ammophila arenaria (European beachgrass) and Ammophila breviligulata (American beachgrass), that were intentionally introduced to the Pacific coast in the early twentieth century to stabilize sand dunes (Wiedemann and Pickart 1996; Hacker et al. 2012). As these ecosystem-engineering grasses spread, they stabilized the naturally shifting sand and built tall, linear foredunes, outcompeting some native plants and decreasing habitat for some native bird species (Zarnetske et al. 2010; Hacker et al. 2012; Biel et al. 2017). Ammophila arenaria was introduced to the Pacific coast first and spread widely from Mexico to Canada. A second introduction of A. breviligulata in the 1930s led to its spread from Washington to central Oregon, where it now dominates but coexists with A. arenaria (Hacker et al. 2012). Common garden experiments, coupled with competition models, show that A. breviligulata is the better competitor compared to A. arenaria with coexistence maintained under high sand supply (Zarnetske et al. 2013) and warming conditions (Biel and Hacker 2021). The two beachgrass species also differ in their morphology and growth patterns, with A. arenaria having higher shoot densities and more vertically growing rhizomes that form tall and narrow dunes (Hacker et al. 2012, 2019). Ammophila breviligulata has lower shoot densities and more horizontally growing rhizomes that produce shorter and wider dunes, which provide less coastal protection (Seabloom et al. 2013).
The hybrid beachgrass has been found in the range overlap of the two Ammophila species but is most abundant from northern Oregon to southern Washington on dunes dominated by A. breviligulata with patches of A. arenaria (Mostow et al. 2021; Askerooth et al. 2024). Field observations show that hybrid individuals are intermediate in some morphological characters (e.g., ligule length, shoot weight, and leaf area) but equal or exceed both parents in shoot height, shoot density, and belowground biomass (Mostow et al. 2021; Askerooth et al. 2024). Thus, these differences in functional morphology may change patterns of sand deposition, resulting in dunes that are taller, wider, and/or more stable than those produced by the parent species, which could have implications for coastal protection, carbon storage, and biodiversity (Zarnetske et al. 2012; Seabloom et al. 2013; Biel et al. 2017, 2019; Stepanek 2023).
To predict the invasion potential of the novel Ammophila hybrid, we determined growth, functional morphology, and species interactions of the first-generation hybrid compared to its parent species using a common garden experiment. We used large sand-filled planters to measure the growth, morphological response, and species interactions of the hybrid and its parent species. Our design included both high- and low-density monocultures to measure density-dependent effects on growth and performance, in addition to pairs of taxa in polyculture to measure two-way interactions of the three beachgrass taxa (Inouye 2001). In particular, we asked: (1) Does the growth and morphology of the Ammophila hybrid differ from that of its parents under controlled, common garden conditions? Based on field data (Mostow et al. 2021), we expected that the hybrid would grow taller, denser, and have greater total biomass than either parent, and that all other characteristics would be intermediate between the parents. (2) Does the Ammophila hybrid compete with its parent species and, if so, what is the predicted outcome of those interactions? We hypothesized that the hybrid would outcompete A. arenaria (similar to A. breviligulata) given that many more hybrid traits exceed those of the European species compared to the American species. Alternatively, the hybrid may outcompete both parent species given a unique combination of traits that increases competitive ability. (3) If the taxa do compete, what mechanisms are important to competition and how does the morphology of the three Ammophila taxa change? Based on a previous interaction experiment (Biel and Hacker 2021), we expected competition would decrease biomass and shoot production of all taxa but that leaf and shoot traits would be unaffected.
Materials and methods
Common garden experiment
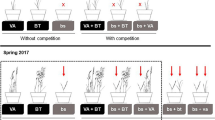
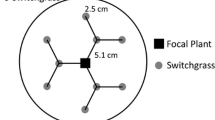
We initiated an 18-month common garden experiment on an open sandy area outside Hatfield Marine Science Center (HMSC), Newport, OR, in March 2020 (Fig. S1) to assess the growth, morphology, and interactions of the two beachgrass parent species (Ammophila arenaria and A. breviligulata) and their first-generation hybrid (A. arenaria × A. breviligulata). We set up 54 1 m2 (base) × 2 m (height) permeable geotextile bags (Flexible Intermediate Bulk Container; Bag Corp, Richardson, TX) in a 9 × 6 grid and filled the bags with 20 cm of locally obtained beach sand (Fig. S1). The bags formed an experimental array with nine treatments per row (or block) and six replicate rows. Treatments were randomly assigned positions within each replicate row. Six of the treatments were monocultures of each beachgrass taxon, planted at two densities (low density = 6 plants per bag and high density = 12 plants per bag) to test for the lack of interactions (low-density treatment) and for intraspecific interactions (high-density treatment). The remaining three treatments were two-species interaction combinations planted at high density (A6B6 = 6 A. arenaria and 6 A. breviligulata plants, A6H6 = 6 A. arenaria and 6 hybrid plants, or B6H6 = 6 A. breviligulata and 6 hybrid plants) to test for interspecific interactions.
In March 2020, we collected individual A. arenaria, A. breviligulata, and hybrid plants (defined as a single rhizome and its attached shoot(s)) from Sunset Beach, Oregon, USA, a site where all three taxa co-occur. Five hybrid individuals collected from the same site were genetically sequenced (SNP genotyped) and all were F1 individuals. Plants were transported to HMSC and planted the following day in the experimental array described above (Fig. S1). We also planted several bags with extra plants of each taxon to be used as replacements for plants that died from transplantation. Prior to planting, we sprayed the rhizomes of each plant with a dilute solution of 1:20 TakeRootR rooting hormone to initiate root formation. The plants were maintained with watering (sprinkled for 2 h, 3 times per week) throughout the first three months (March–June 2020) to reduce transplant shock and stimulate growth.
A single sand deposition regime was established in the bags to maintain the growth of the plants. Each month, two centimeters of sand was gently added to the initial sand depth of each bag [19.9 ± 0.2 cm (mean ± standard error)], resulting in a final sand depth of 45.1 ± 0.9 cm after 18 months of deposition and sand settling. The annual sand deposition rate was 17.4 ± 0.2 cm per year, which closely reflects the median field deposition rate of 18 cm per year measured by Biel et al. (2019) for Washington and Oregon coastal foredunes.
One-month post-transplantation, dead plants (5 A. arenaria, 5 A. breviligulata, and 12 hybrid plants) were replaced with healthy plants from the extra bags. By April 2020 all the plants had successfully established. We then counted the total number of shoots per bag every month from April 2020 to September 2021. The initial shoot counts were roughly equivalent among taxa and the high-density treatment was twice that of the low-density treatment (Table S1).
Starting in late September 2021, the experiment was terminated, and the grasses were removed from the bags, taking special care to preserve the connections between the roots, rhizomes, and shoots (Fig. S2). We first cut away the geotextile bag and then removed the plants from the sand by brushing away the outer layer of sand and then washing down the inner layers of sand with an open hose. Once liberated from the sand, the plants were rinsed in water to remove the remaining sand. Back at the laboratory, the plants were spread out on tables and air dried at room temperature (21 °C) until completely dry (2–3 weeks).
Beachgrass response variables
The dried plant samples per bag were separated by taxon and then into aboveground biomass (material 1 cm above the primary node of the rhizome) and belowground biomass (the rest of the plant) (Fig. S3). For each taxon, we counted the number of shoots and weighed the aboveground and belowground biomass (g). We also measured the total rhizome length (cm) (in monoculture treatments), which is the sum of the lengths of all rhizomes contained within a bag. Additionally, 5 individual plants (defined as a rhizome with a variable number of attached shoots) of each taxon were subsampled within each bag and the following characteristics were measured: total shoots per plant, vertical and lateral shoots per plant, shoot weight (primary shoot) (g), shoot height (primary shoot) (cm), shoot leaf number (primary shoot), and shoot leaf width (second leaf on the primary shoot) (mm) (Fig. S3). Finally, we measured total rhizome length per plant (cm), rhizome distance between the primary and each of the secondary, tertiary, and quaternary shoots, and the rhizome width (below the primary shoot) (cm) (Fig. S3). For each plant, we calculated shoot leaf area (cm2) of the primary shoot as half shoot length (cm) × leaf width (cm) × leaf number and standardized rhizome length of each plant by dividing total rhizome length by shoots per plant.
Statistical analyses
All analyses were preformed using R (R Development Core Team 2019). All response variables were tested for normality and homoscedasticity and outliers were visually assessed. Using only the low-density monoculture results, we conducted one-factor ANOVAs (followed by Tukey’s honestly significant difference [HSD] post hoc tests, when statistically significant) or Kruskal–Wallis tests (followed by pairwise Wilcoxon rank-sum post hoc tests, when statistically significant) to determine taxon-specific differences in whole bag traits (i.e., total biomass, aboveground biomass, belowground biomass, total shoot number, and total rhizome length) and plant, shoot, and rhizome morphological traits (i.e., shoots per plant, proportion vertical shoots per plant, shoot weight, shoot height, leaf number, leaf width, leaf area, rhizome length per plant, standardized rhizome length, rhizome distance between primary, secondary, tertiary, and quaternary shoots, and rhizome width). We also used principal component analysis (PCA) to determine if the three taxa differed on the combined suite of plant, shoot, and rhizome response variables. A scree plot was run and principal components with eigenvalues > 1 were used in the PCA.
To compare the effect of intraspecific and interspecific interactions on the three species, we used two-way ANOVAs to determine whether whole-bag traits (i.e., total biomass, aboveground biomass, belowground biomass, and total shoot number) and plant, shoot, and rhizome morphological traits (i.e., shoots per plant, shoot weight, shoot height, leaf area, and standardized rhizome length) differed based on taxon, experimental treatment (interactions versus no interactions), and their possible interaction. Tukey HSD post hoc tests were conducted on significant factors unless interactions were found, in which case one-way ANOVAs and Tukey HSD post hoc comparisons were conducted between levels of each factor (Underwood 1997). Whole-bag traits (i.e., biomass and shoot totals) of high-density monoculture treatments (12 single taxon plants per bag) were divided by two such that they could be compared relative to the six focal plants of each taxon in the other treatments.
To describe the magnitude of the effect of intraspecific and interspecific interactions on the same whole-bag traits and plant, shoot, and rhizome morphological traits as above, we calculated the log response ratio (LRR) for each interaction treatment (i.e., the high-density monoculture and pairwise taxa treatments) as:
where the no interaction treatment for each taxon is the low-density monoculture. We then conducted two-way ANOVAs (including Tukey HSD post hoc tests and one-way ANOVAs, when appropriate) for taxon, experimental treatment, and their possible interaction. One sample two-tailed Student’s t-tests were also used to test whether mean LRR values were different from zero (no effect).
To assess the outcomes of interspecific interactions, and to estimate and compare growth parameters for each taxon, we used linear mixed models (with the R package lme4) to parameterize the Ricker model, a density-dependent logistic growth model previously used by Biel and Hacker (2021) for beachgrass interactions. The Ricker model describing a generic two-species interaction is written as:
in which N1 and N2 are the total shoot numbers for two interacting species, r1 and K1 are the intrinsic growth rate and carrying capacity of the species of interest, and α1,2 is the competition coefficient representing the effect of species 2 on species 1. N1,final is the total shoot number of species 1 at the end of the experiment while N1,initial is the initial planted shoot number of species 1. The response variable, the log ratio of N1,final and N1,initial, represents the log response ratio of the effect of intraspecific and interspecific competition on total shoot number of the focal species. Expressed as a linear mixed model, this equation takes the form:
We evaluated the Ricker model for each taxon in each two-way interaction. For example, we first evaluated the two-way interaction between A. arenaria as species 1 and A. breviligulata as species 2 and then the reciprocal interaction with A. breviligulata as species 1 and A. arenaria as species 2. We then repeated the analyses for the interactions between the hybrid and each of its parent species for a total of six separate models. After parameterizing the linear models for each two-way interaction, we estimated the intrinsic growth rate (r), carrying capacity (K), and competition coefficient (α) for each species using the following derivations of the equations:
Finally, we used the parameter estimates (Table S2) to graph the zero net growth isoclines of each species (i.e., line in species space at which dN/dt = 0, or there is no change in population size), to identify any equilibrium points of these population curves in species space, and to evaluate the stability of those equilibria. For example, the zero net growth isocline for species 1 is:
where the carrying capacity for species 1 (K1) is the intercept of the line and the interaction effect of species 2 on species 1 (α1, 2) is the slope of the line.
Results
Taxon-specific differences in growth and morphology
We found that under common garden conditions the hybrid beachgrass had equal or greater biomass and shoot density compared to its parent species (Figs. 1, 2, Table 1, Table S3). Specifically, within the monoculture bags, the hybrid had greater total biomass, aboveground biomass, and belowground biomass compared to A. arenaria, and the hybrid was equal across those measures when compared to A. breviligulata (Fig. 2, Table 1, Table S3). Moreover, the hybrid greatly exceeded both its parent species in shoot density (by 1.6-fold), but there was no difference in density between the two parent congeners by the end of the experiment. We also found a strong seasonal and species-specific component to shoot production over time (Fig. 1). Ammophila arenaria had its highest shoot production in late summer to fall (August to November 2020) and A. breviligulata, in winter to early summer (January to June 2021). Interestingly, the hybrid had greater shoot production rates both in late summer to fall (similar to A. arenaria) and in winter to early summer (similar to A. breviligulata), mirroring patterns of production seen for both parent species. Finally, shoot production per plant was greater in low-density bags and the seasonal patterns of production were the same between the two density treatments (Fig. 1).

Smoothed mean shoot density (per m2 bag) in high-density (solid) and low-density (dashed) monoculture beachgrass treatments over the 18-month experiment (April 2020–September 2021). Taxa are Ammophila arenaria (AMAR; red), A. breviligulata (AMBR; blue), and hybrid (HYBR; purple)

Comparison of whole-bag traits (per m2 bag) under low-density monoculture (no interaction) and species interaction treatments. Box plots show median, 1st and 3rd quartile box, and 1.5 interquartile range whiskers. Taxa are Ammophila arenaria (AMAR), A. breviligulata (AMBR), and hybrid (HYBR). Letters represent significant differences among taxa in low-density monoculture treatment (Tukey post hoc test, p < 0.05; Table S3). Number of stars represent significant differences among taxa from interaction treatments (Tukey post hoc test, p < 0.05; Table S5)
Plant- and shoot-level morphological differences determined from field samples (Mostow et al. 2021) were largely maintained or expanded under common garden, monoculture conditions (Fig. 3, Table 1, Table S3). The hybrid beachgrass had more shoots per plant and taller shoots compared to both parent species. Moreover, hybrid shoots were heavier, and leaves were wider than those of A. arenaria but similar to those of A. breviligulata. Finally, A. breviligulata had the widest leaves, greatest leaf area, and longest total rhizome length of the three taxa and A. arenaria had the greatest standardized rhizome length as well as rhizome distances between primary, secondary, tertiary, and quaternary shoots.

Comparison of plant, shoot, and rhizome traits under low-density monoculture (no interaction) and species interaction treatments. Box plots show median, 1st and 3rd quartile box, and 1.5 interquartile range whiskers. Taxa are Ammophila arenaria (AMAR), A. breviligulata (AMBR), and hybrid (HYBR). Letters represent significant differences among taxa in low-density monoculture treatment (Tukey post hoc test, p < 0.05; Table S3). Number of stars represent significant differences among taxa from interaction treatments (Tukey post hoc test, p < 0.05; Table S5)
A principal component analysis on the suite of plant, shoot, and rhizome traits showed that samples grouped by taxon, with the hybrid clustering between the two Ammophila species and nested within the A. breviligulata cluster, which had the most variation in morphological trait values of the three taxa (Fig. S4). The first principal component (PC1) explained the greatest variation (40.6%) and is most correlated with plant size (i.e., total biomass, aboveground biomass, belowground biomass, total shoot number, and total rhizome length) while PC2 explained 19.3% variation and is most correlated with shoot characters (i.e., shoot weight, shoot leaf number, and shoot leaf width) (Fig. S4, Table S4). Two additional PCs with eigenvalues slightly greater than 1 explained 11.6% (PC3) and 8.4% (PC4) of the variation and were correlated with shoot and rhizome traits (Table S4).
Competition among the beachgrass taxa
The species interaction treatments showed that the hybrid beachgrass grew larger and was more productive than its parent species under all treatments (Fig. 2). Two-way ANOVAs revealed that total biomass, aboveground biomass, belowground biomass, and total shoot number varied by taxon but not by treatment or their interaction (Table S5). Post hoc tests showed a competitive hierarchy with the hybrid beachgrass having the highest biomass and total shoot number, followed by A. breviligulata, and then A. arenaria. However, A. arenaria and A. breviligulata did not differ in total shoot number (Table S5).
Log response ratios (LRRs) for the whole-bag response variables illustrate the effect of the interaction treatments on plant biomass and shoot production (Fig. S6). First, none of the LRRs varied by treatment and only the LRR of total shoot number varied by taxon (Table S6). Post hoc tests revealed that A. breviligulata experienced the strongest negative effects of competition on total shoot number, followed by A. arenaria. Second, the LRRs for total biomass, aboveground biomass, belowground biomass, and total shoot number under the various interaction treatments were negative and the majority were significantly different from zero, indicating intraspecific and interspecific competition (Fig. S5, Table S7). The largest negative interaction for all four production traits occurred when A. breviligulata grew with the hybrid (Table S7).
Finally, the parameter estimates for the Ricker model showed that the strongest competition occurred between A. breviligulata and A. arenaria (αA,B = 1.72) and between the hybrid and A. breviligulata (αB,H = 1.56) and the weakest occurred between A. arenaria and the hybrid (αH,A = 0.50) (Table S2). The hybrid had the highest estimated intrinsic growth rate and A. arenaria had the lowest. Ammophila arenaria had the highest carrying capacity when grown with A. breviligulata (K = 61.9), followed by the hybrid growing with A. breviligulata (K = 55.0).
The state space graphs, with zero net growth isoclines for each species parameterized with the Ricker model values (Table S2), projected an unstable equilibrium between A. arenaria with A. breviligulata (Fig. 4a) and A. arenaria with the hybrid (Fig. 4b). The state space graph of A. breviligulata with the hybrid indicates that the hybrid would competitively exclude A. breviligulata at equilibrium regardless of starting shoot densities (Fig. 4c).

State space diagrams showing the zero-net growth isoclines for the two-way interactions of Ammophila arenaria (NA; red line), A. breviligulata (NB; blue line), and the hybrid (NH; purple line), calculated using the Ricker model. a Unstable coexistence between A. arenaria and A. breviligulata. b Unstable coexistence between A. arenaria and the hybrid. c Competitive exclusion of A. breviligulata by the hybrid. See Table S2 for model parameter estimates
Effects of competition on morphology of hybrid beachgrass and its parent species
Plant, shoot, and rhizome traits all differed by taxa, and there was an additional treatment effect and/or interactive effect for some shoot specific traits (i.e., shoot number, weight, and height) (Fig. 3, Table S5). Post hoc tests revealed that for shoot number and shoot height, the hybrid typically exceeded its parents when grown alone or with another parent species. Shoot number did not differ among the treatments for the hybrid, but it did for the two parent species with higher shoot numbers under monoculture compared to interspecific competition. Likewise, shoot height did not differ for the hybrid or A. arenaria among the treatments, but shoots of A. breviligulata were the tallest when grown in high-density monoculture, the shortest when grown with the hybrid, and intermediate in height when grown with A. arenaria.
Comparisons of shoot weight among the taxa showed that A. breviligulata had the heaviest shoots, followed by the hybrid, and then A. arenaria (Fig. 3, Table S5). Ammophila arenaria shoot weight did not vary by treatment, but A. breviligulata shoot mass was significantly decreased by growing with the hybrid. The hybrid’s shoot mass was the greatest when grown with conspecifics, the lowest when grown with A. arenaria, and intermediate when grown with A. breviligulata. Leaf area was the largest for A. breviligulata, followed by the hybrid, and then A. arenaria and standardized rhizome length was the longest for A. arenaria and then A. breviligulata and the hybrid, which did not differ (Fig. 3, Table S5).
The LRRs of morphological traits did not vary by treatment, but there was a treatment by taxon interaction for shoots per plant (Fig. 5, Table S6). The hybrid strongly negatively affected the number of shoots per plant of A. arenaria and A. breviligulata but not itself in monoculture. The LRRs of shoot height and standardized rhizome length varied by taxon but post hoc tests revealed different effects. Ammophila breviligulata experienced the strongest negative effects of competition on shoot height, followed by the hybrid. Conversely, A. breviligulata experienced the strongest positive effects on standardized rhizome length. However, most LRRs for plant, shoot, and rhizome traits did not differ from zero and the ones that did were small and negative (Fig. 5, Table S7), especially compared to those measured for biomass and shoot numbers (Fig. S5). For example, the hybrid caused a decline in shoots per plant, shoot weight, and shoot height in A. breviligulata, and the leaf area of A. arenaria, but had no effect on itself. Ammophila arenaria caused a decline in shoots per plant and shoot height and an increase in the standardized rhizome length in A. breviligulata, but it had no effect on the hybrid or itself. Finally, A. breviligulata had no effect on the morphology of any taxa.

Mean (± SE) log response ratio (LRR) of plant, shoot, and rhizome traits under interaction treatment conditions. Taxa are Ammophila arenaria (AMAR), A. breviligulata (AMBR), and hybrid (HYBR). For trait and taxon differences, see Supplementary Table 6. Stars represent a significant difference from zero (one-sample Student’s t-test, two-tailed, true mean = 0, p < 0.05; Supplementary Table 7)
Discussion
The goal of this study was to describe growth, functional morphology, and species interactions of the newly discovered Ammophila hybrid and its parent species using a common garden experiment to predict the invasion potential of the hybrid in areas where the three taxa co-occur. In monoculture, although the hybrid displayed some intermediate characteristics, such as shoot weight and rhizome length, it also exhibited hybrid vigor by producing more shoots that were taller than either parent (Figs. 1, 2, 3, Table 1). In addition, the hybrid negatively affected both A. breviligulata and A. arenaria, and it experienced the least intraspecific and interspecific competition of the three congeners (Figs. 2, 3, S5). There was also strong support for the previous finding (Zarnetske et al. 2013; Biel and Hacker 2021) that A. breviligulata is a better competitor to A. arenaria. Overall, based on the Ricker model, our results indicate that the hybrid competitively excludes A. breviligulata under all density conditions (Fig. 4, Table S2). However, A. arenaria coexists with the hybrid and with A. breviligulata in an unstable equilibrium, indicating a competitive hierarchy in which the hybrid outcompetes A. breviligulata and potentially A. arenaria depending on the relative abundances of the two taxa. Given that the hybrid is the most abundant on dunes dominated by A. breviligulata, where A. arenaria is sparse and patchy, the hybrid could competitively exclude both its parents in years to come, with implications for dune ecosystem function and services as discussed below.
Intrinsic differences in growth form and morphology
Our monoculture experiments revealed that there are intrinsic differences in growth and morphology among the three beachgrass taxa (Figs. 1, 2, 3, Table 1) that strongly suggest the hybrid will spread quickly and likely capture more sand than either parent species. We found that the hybrid had an additive pattern of seasonal shoot production that could explain the nearly two-fold difference in shoot numbers compared to its parent species (Fig. 1). In particular, A. arenaria produced most of its shoots in the late summer to fall (August to November) while A. breviligulata did so in the winter to early summer (January to June). However, the hybrid displayed both patterns of shoot production with breaks in production in mid-summer and mid-winter. Thus, the hybrid appears to grow nearly year-round, a phenological trait that could allow it to spread much faster than either parent species.
We also found that the hybrid has morphological and growth form characteristics that may allow it to capture as much or more sand than either parent species. For example, A. arenaria plants grew in a clumped fashion with multiple thin shoots and leaves produced from each rhizome (Figs. 3, 6, Table 1). A wind tunnel experiment showed that even though the shoots and leaves of A. arenaria are thin, and thus have a lower sand capture efficiency than A. breviligulata, the higher shoot densities of A. arenaria result in greater sand deposition depth (Zarnetske et al. 2012). Likewise, in response to sand burial, our experiment shows that A. arenaria produces longer vertical rhizomes that allow its shoots to remain at the sand surface (Figs. 3, 6, Table 1), thus contributing to continual sand accretion and plant growth (Baye 1990; Zarnetske et al. 2013). In contrast, A. breviligulata produced fewer but thicker shoots and wider leaves attached to shorter more horizontally oriented rhizomes, creating a more creeping growth form that captures less sand than A. arenaria per given area (Figs. 3, 6, Table 1). The differential growth patterns result in taller and narrower dunes for A. arenaria and shorter and wider dunes for A. breviligulata (Hacker et al. 2012; Biel et al. 2019). Interestingly, the hybrid displayed a mixture of traits compared to its parents, including more and taller shoots of intermediate weight, leaves of intermediate width and area, and shoots attached to rhizomes of intermediate length (Figs. 3, 6, Table 1). These traits suggest a tighter and more robust growth form, which we hypothesize could lead to similar or even greater vertical sand deposition than A. arenaria (Fig. 6) and ultimately taller dunes as a byproduct. The hybrid also produces greater belowground biomass than its parent species, a trait that has been known to increase dune stability in the face of erosion (Bryant et al. 2019; De Battisti and Griffin 2020).

Illustration of the hypothesized growth form and morphological differences of the three beachgrass taxa over time. Hypothesized taxon-specific sand capture and deposition patterns, assuming similar sand supply, are delimited by the solid and dashed lines. The stippled line and value represent average plant height. Taxa are Ammophila arenaria (AMAR), A. breviligulata (AMBR), and hybrid (HYBR). Adapted from
Possible mechanisms of competition among the hybrid and its parent species
Growth patterns in clonal plants have been described as either “guerilla,” characterized by long rhizomes between shoots to allow the plant to spread widely, or “phalanx,” characterized by densely clumped shoots with shorter rhizomes to allow plants to grow in place (Doust 1981). The more clumped growth form of A. arenaria has been described as phalanx type while the lateral spread of A. breviligulata as guerilla-like (Baye 1990). Our experiments suggest that A. breviligulata employs a more guerrilla-like growth form when in competition with A. arenaria, increasing its lateral growth to avoid competition. This is supported by an increase in standardized rhizome length for A. breviligulata when competing with A. arenaria but also with the hybrid (Figs. 5, 6). Likewise, A. arenaria caused a significant decline in A. breviligulata shoot density likely given its phalanx-style of growth. In contrast, we suggest that the hybrid, with its intermediate rhizome length but exceptional shoot density, is using a mixed phalanx–guerilla growth form that allows it to spread laterally into open space and then grow to high shoot densities (Fig. 6), both of which gives it a competitive advantage over its parent species. For example, when grown with the hybrid, the two congener parents, but particularly A. breviligulata, had significantly lower shoot densities, shoot weights, and shoot heights (Fig. 5).
Our Ricker estimates of competition between A. arenaria and A. breviligulata followed similar patterns to two other common garden experiments, with A. breviligulata competitively dominating A. arenaria (Zarnetske et al. 2013; Biel and Hacker 2021). However, all the studies generally show that, despite the reciprocal but unequal competitive effects between A. breviligulata and A. arenaria, competitive exclusion is less common than coexistence. For example, Zarnetske et al. (2013) found that sand deposition rates mediated the strength and direction of interspecific interactions, allowing species to coexist at higher sand deposition rates. Moreover, Biel and Hacker (2021) found competitive exclusion, but the probability of coexistence increased with increasing sand burial and warming treatments. We did not manipulate sand deposition in our experiment but used a constant rate that was less than half that of the highest rates of the other two studies (17 cm per year compared to ~ 45 cm per year for Zarnetske et al. 2013 and Biel and Hacker 2021). The studies also differed in initial planting densities, which could affect the comparisons among the studies. For example, Biel and Hacker (2021) initiated their experiment in smaller plots with four times higher starting densities than the other two studies, likely increasing the intensity of competition between A. breviligulata and A. arenaria. Additionally, seasonal differences in the timing of the installation and termination of our experiment compared to the other studies could have created some differences in the outcome of the interactions similar to what has been shown in Sinclair et al. (2020).
Shifts in beachgrass dominance and changes in dune ecosystem functions and services
Our study suggests that the Ammophila hybrid could spread quickly given its nearly year-round growth phenology and its potential to outcompete its parent species. Currently, the hybrid mostly occurs across a 250 km region in southern Washington and northern Oregon where dunes are dominated by A. breviligulata with sparser and patchy A. arenaria (Mostow et al. 2021; Askerooth et al. 2024). Based on Ricker model results, we hypothesize that even though A. breviligulata is most abundant in the region now, it might ultimately be competitively displaced by the hybrid in the future. Ammophila arenaria could experience the same fate, especially if its relative abundance is smaller than that of the hybrid over time. Such a landscape-scale change in dominant beachgrass species is plausible given the historical pattern of beachgrass invasions in the Pacific Northwest (Seabloom and Wiedemann 1994; Hacker et al. 2012). Ammophila arenaria was the dominant beachgrass after its introduction and establishment in the mid-1900s until the introduction of A. breviligulata near the Columbia River in the 1930s. In areas where the two beachgrasses co-occurred, A. breviligulata steadily outcompeted and replaced A. arenaria as the dominant beachgrass (Seabloom and Wiedemann 1994; Hacker et al. 2012; David et al. 2015; Zarnetske et al. 2015). Taken together, the results of this experiment and the historical precedent point towards the hybrid replacing A. breviligulata as the dominant beachgrass in northern Oregon and southern Washington, albeit over decades of time.
In addition, the spread of the hybrid may be facilitated or hindered by the hybrid swarm currently developing (Mostow 2022). Later generation and backcross hybrids may not have the same interactions with their parents compared to the F1 individuals used in this experiment. Further studies of variation in morphology and growth form of F1, F2, and backcross individuals will be necessary to understand the effect of the hybrid swarm on competitive displacement. Additionally, the presence of backcrossed individuals suggests the future possibility of adaptive introgression between these species. This could be especially important if gene flow from A. breviligulata increases the competitive ability or fitness of A. arenaria (the only parent with a documented backcross), changing the well-established pattern of interspecific competition.
In this system, a series of sequential introductions of closely related non-native Ammophila species resulted in rapid invasion, landscape level changes, and now hybridization. If the hybrid does come to dominate dunes in the US Pacific Northwest, this ecological change could have a significant effect on dune morphology, creating both taller and wider dunes than those built by its parents. In addition, the Ammophila beachgrasses not only compete with one another but also with native plants, including the native perennial dune grass Leymus (Elymus) mollis (Hacker et al. 2012; Zarnetske et al. 2013). The displacement of A. arenaria by A. breviligulata has been associated with declines in native plant diversity and cover (Seabloom and Wiedemann 1994; Hacker et al. 2012; David et al. 2015). Although we do not know how the hybrid beachgrass interacts with native plant species, changes in beachgrass dominance are likely to have cascading effects on native biodiversity. Moreover, if the hybrid does come to dominate areas of Pacific Northwest dunes, it could affect dune carbon storage. Dune grasses play an important role in moderating the rate and total amount of carbon stored in dunes by accreting sand, which carries its own carbon stock, and storing carbon in belowground growth (Jay 2021; Stepanek 2023).
In conclusion, the results of this study add to the growing consensus that hybridization is a viable evolutionary mechanism promoting invasion success (Ellstrand and Schierenbeck 2000; Hovick and Whitney 2014). Our study is unique in that it compared functional traits of the hybrid with both parent species, included intraspecific and interspecific interactions, used first-generation individuals, and was conducted in a common garden setting under realistic environmental conditions. Given our results, we conclude that the hybrid beachgrass is likely a better invader than its parent species, with the potential to spread widely and fundamentally change Pacific Northwest dune ecosystems in the future.
Data availability
Datasets used in this study are available from the corresponding author on reasonable request.
Code availability
Code developed in this study is available from the corresponding author on reasonable request.
References
Abbott RJ (1992) Plant invasions, interspecific hybridization and the evolution of new plant taxa. Trends Ecol Evol 7:401–405
Ainouche M, Baumel A, Salmon A (2004) Spartina anglica C. E. Hubbard: a natural model system for analysing early evolutionary changes that affect allopolyploid genomes. Biol J Linnean Soc 82:475–484
Anderson E, Stebbins GL (1954) Hybridization as an evolutionary stimulus. Evolution 8:378–388
Askerooth R, Mostow R, Ruggiero P, Barreto F, Hacker SD (2024) The distribution, spread, and ecological effects of a novel, invasive beachgrass hybrid, Ammophila arenaria × A. breviligulata, U.S. Ecosphere 15:e4830
Baye P (1990) Comparative growth responses and population ecology of European and American beachgrasses (Ammophila spp.) in relation to sand accretion and salinity. The University of Western Ontario, London
Biel RG, Hacker SD (2021) Warming alters the interaction of two invasive beachgrasses with implications for range shifts and coastal dune functions. Oecologia 197:757–770
Biel RG, Hacker SD, Ruggiero P, Cohn N, Seabloom EW (2017) Coastal protection and conservation on sandy beaches and dunes: context-dependent tradeoffs in ecosystem service supply. Ecosphere 8:e01791
Biel RG, Hacker SD, Ruggiero P (2019) Elucidating coastal foredune ecomorphodynamics in the US Pacific Northwest via Bayesian networks. J Geophys Res Earth Surf 124:1919–1938
Bryant DB, Anderson Bryant M, Sharp JA, Bell GL, Moore C (2019) The response of vegetated dunes to wave attack. Coast Eng 152:103506
Comai L (2005) The advantages and disadvantages of being polyploid. Nat Rev Genet 6:836–846
David AS, Zarnetske PL, Hacker SD, Ruggiero P, Biel RG, Seabloom EW (2015) Invasive congeners differ in successional impacts across space and time. PLoS ONE 10:e0117283
De Battisti D, Griffin JN (2020) Below-ground biomass of plants, with a key contribution of buried shoots, increases foredune resistance to wave swash. Ann Bot 125:325–334
Doust LL (1981) Population dynamics and local specialization in a clonal perennial (Ranunculus repens): I. the dynamics of ramets in contrasting habitats. J Ecol 69:743–755
Ellstrand NC, Schierenbeck KA (2000) Hybridization as a stimulus for the evolution of invasiveness in plants? Proc Natl Acad Sci 97:7043–7050
Hacker SD, Zarnetske P, Seabloom E, Ruggiero P, Mull J, Gerrity S, Jones C (2012) Subtle differences in two non-native congeneric beach grasses significantly affect their colonization, spread, and impact. Oikos 121:138–148
Hacker SD, Jay KR, Cohn N, Goldstein EB, Hovenga PA, Itzkin M, Moore LJ, Mostow RS, Mullins EV, Ruggiero P (2019) Species-specific functional morphology of four US Atlantic coast dune grasses: biogeographic implications for dune shape and coastal protection. Diversity 11:82. https://doi.org/10.3390/d11050082
Hovick SM, Whitney KD (2014) Hybridisation is associated with increased fecundity and size in invasive taxa: meta-analytic support for the hybridisation-invasion hypothesis. Ecol Lett 17:1464–1477
Hovick SM, Campbell LG, Snow AA, Whitney KD (2012) Hybridization alters early life-history traits and increases plant colonization success in a novel region. Am Nat 179:192–203
Inouye BD (2001) Response surface experimental designs for investigating interspecific competition. Ecology 82:2696–2706
Jay KR (2021) Investigating the role of dune grasses, carbon storage, and marine nutrient subsidies to the functions and services of U.S. Central Atlantic coastal dune ecosystems. Oregon State University Corvallis, Oregon, USA
Kuester A, Conner JK, Culley T, Baucom RS (2014) How weeds emerge: a taxonomic and trait-based examination using United States data. New Phyto 202:1055–1068
Lee CE (2002) Evolutionary genetics of invasive species. Trends Ecol Evol 17:386–391
Mostow R (2022) Hybridization of non-native dune-building beachgrasses on the US Pacific Northwest coast: characterization of functional morphology, hybrid swarm composition, and ecological consequences of Ammophila arenaria x A. breviligulata. Oregon State University, Corvallis
Mostow RS, Barreto F, Biel R, Meyer E, Hacker SD (2021) Discovery of a dune-building hybrid beachgrass (Ammophila arenaria × A. breviligulata) in the U.S. Pacific Northwest. Ecosphere 12:e03501
Parepa M, Fischer M, Krebs C, Bossdorf O (2014) Hybridization increases invasive knotweed success. Evol Appl 7:413–420
Pyšek P, Richardson DM (2007) Traits associated with invasiveness in alien plants: where do we stand? In: Nentwig N (ed) Biological invasions. Springer, Berlin, pp 97–125
R Development Core Team (2019) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Rieseberg LH, Archer MA, Wayne RK (1999) Transgressive segregation, adaptation and speciation. Heredity 83:363–372
Schierenbeck KA, Ellstrand NC (2009) Hybridization and the evolution of invasiveness in plants and other organisms. Biol Invasions 11:1093–1105
Seabloom EW, Wiedemann AM (1994) Distribution and effects of Ammophila breviligulata Fern. (American Beachgrass) on the foredunes of the Washington coast. J Coast Res 10:178–188
Seabloom EW, Ruggiero P, Hacker SD, Mull J, Zarnetske P (2013) Invasive grasses, climate change, and exposure to storm-wave overtopping in coastal dune ecosystems. Glob Change Biol 19:824–832
Sinclair MN, Woods NN, Zinnert JC (2020) Seasonal facilitative and competitive trade-offs between shrub seedlings and coastal grasses. Ecosphere 11:e02995
Stebbins GL (1959) The role of hybridization in evolution. P Am Philos Soc 103:231–251
Stebbins GL (1985) Polyploidy, hybridization, and the invasion of new habitats. Ann Mo Bot Gard 72:824–832
Stepanek J (2023) Carbon storage in US Pacific Northwest coastal dunes: the role of invasive beachgrasses and sand supply. Oregon State University, Corvallis
Theoharides KA, Dukes JS (2007) Plant invasion across space and time: factors affecting nonindigenous species success during four stages of invasion. New Phytol 176:256–273
Underwood AJ (1997) Experiments in ecology: their logical design and interpretation using analysis of variance. Cambridge University Press, Cambridge
Vilà M, Weber E, Antonio CMD (2000) Conservation implications of invasion by plant hybridization. Biol Invasion 2:207–217
Wiedemann AM, Pickart A (1996) The Ammophila problem on the Northwest coast of North America. Landsc Urban Plan 34:287–299
Zarnetske PL, Seabloom EW, Hacker SD (2010) Non-target effects of invasive species management: beachgrass, birds, and bulldozers in coastal dunes. Ecosphere 1:1–20
Zarnetske PL, Hacker SD, Seabloom EW, Ruggiero P, Killian JR, Maddux TB, Cox D (2012) Biophysical feedback mediates effects of invasive grasses on coastal dune shape. Ecology 93:1439–1450
Zarnetske PL, Gouhier TC, Hacker SD, Seabloom EW, Bokil VA (2013) Indirect effects and facilitation among native and non-native species promote invasion success along an environmental stress gradient. J Ecol 101:905–915
Zarnetske PL, Ruggiero P, Seabloom EW, Hacker SD (2015) Coastal foredune evolution: the relative influence of vegetation and sand supply in the US Pacific Northwest. J R Soc Interface 12:20150017
Acknowledgements
Thanks to the Hatfield Marine Science Center staff, especially C. Moffett, B. Butler, and J. Lewis for help with installation and maintenance of the experiment, to R. Antaki, R. Askerooth, M. Barker, D. Fee, H. Fulton-Bennett, H. Ly, B. Lynn, C. Magel, Z. Meunier, C. Nye, V. Quennessen, C. Ringer, and J. Stepanek for their invaluable help in the field, and to OSU undergraduate students H. Ly and H. Le Boeuf who processed plant samples in the lab.
Funding
This work was supported by a NOAA Oregon Sea Grant (NA18OAR4170072) grant to SDH and FSB and a National Science Foundation Graduate Research Fellowship and Mamie Markham Research Award (Hatfield Marine Science Center, Newport, OR) to RSM.
Author information
Authors and Affiliations
Contributions
RSM and SDH conceived and designed the experiment. RSM performed the experiments and analyzed the data, with assistance from SDH and FSB. RSM and SDH wrote the manuscript, and all authors reviewed and edited the manuscript.
Corresponding author
Ethics declarations
Conflicts of interest
The authors declare no conflicts of interest.
Ethics approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Additional information
Communicated by Jennifer Funk.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Mostow, R.S., Barreto, F.S. & Hacker, S.D. A hybrid beachgrass (Ammophila arenaria × A. breviligulata) is more productive and outcompetes its non-native parent species. Oecologia 205, 81–94 (2024). https://doi.org/10.1007/s00442-024-05548-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-024-05548-5