
Abstract
Resource selection is a dynamic process driven by habitat valuation and risk avoidance in heterogeneous landscapes. Resource selection and movement decisions of individuals may be sensitive to intrinsic factors, such as body condition, and variation in these choices may have consequences on subsequent survival. We evaluated northern bobwhite (Colinus virginianus) brood resource selection patterns to quantify utility of different cover types during the development period using integrated step-selection analysis in a Bayesian hierarchical modeling framework with three brood stages: flightless broods ≤ 14 days old, dependent broods 15–35 days old, and independent broods over 35 days old. Broods showed strongest selection for native grasslands that were burned and grazed at least once in the previous two years, and agricultural fields. Brood mobility improved with age; broods > 35 days old travelled farther on average and took daily steps > 200 m more frequently than younger broods. Young broods ≤ 14 days old did not select for idle native grasslands, while broods > 35 days old did select for that cover type. Young broods also selected areas farther from trees compared to older broods. We evaluated the survival consequences of resource selection by comparing patterns in choices of broods that succeeded to choices of broods that failed to survive to 35 days. Successful broods chose habitats with more shrub cover and areas farther from trees compared to failed broods. Our results suggest that conservation planning should consider age-specific patterns in habitat use and demographic consequences of habitat choice for greatest effectiveness.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Movement and resource selection patterns of individuals determine population-level distributions across heterogeneous landscapes (Lima and Zollner 1996). Individuals make choices perceived to maximize net quality of available food, cover, and other resources contributing to survival and reproductive success (i.e., fitness; Cooper and Millspaugh 1999). Patterns in choice reflect resource requirements and the relative value of available habitats. Resource selection is a spatial process describing animal movements through a variable landscape (Thurfjell et al. 2014; Avgar et al. 2016). Individuals tend to stay in or move toward environments perceived to maximize resource quality and minimize predation risk (Dinkins et al. 2012; De Cesare et al. 2014; Atuo and O’Connell 2018).
Population distributions and individual home ranges incorporate patterns of daily movement and resource valuation (Schooley 1994; Mayor et al. 2009; Dzialak et al. 2012). Selection also varies by individual and can be influenced by intrinsic factors, such as life stage, breeding status, and body condition (Paasivaara and Pöysä 2008; Brooke et al. 2015; Kane et al. 2017). For young animals, habitat choices and movement patterns may be influenced by age and development (Wiltermuth et al. 2015; Jenkins et al. 2017). Patterns of resource selection and movement can cascade to influence survival of young (Rotella and Ratti 1992; Mainguy et al. 2006; Goddard and Dawson 1999). The contribution of variation in individual behavior to population-level processes, such as resource selection and survival, has not been thoroughly addressed in applied ecological research (Merrick and Koprowski 2017).
Northern bobwhite (Colinus virginianus; hereafter bobwhite) is an early successional, disturbance-dependent species experiencing range-wide declines on rapidly changing landscapes in North America (Hernández et al. 2013). Productivity and recruitment contribute significantly to bobwhite population growth (McConnell et al. 2018). Nest-site characteristics for bobwhite have been well studied; vegetation must provide adequate cover to conceal incubating adults and nests from predators and protect them from heavy rainfall or temperature extremes (Klimstra and Roseberry 1975; Collins et al. 2009; Carroll et al. 2015). During the brood phase, vegetation must additionally facilitate movement and foraging activities of young. Many factors, such as dispersal from nesting habitat, nutritional requirements, predation pressure, weather, and mobility, can explain brood resource selection and movement decisions (Sonerud 1985; Rotella and Ratti 1992; Tanner et al. 2017).
We quantified resource selection and movement patterns of bobwhite broods from hatch through their development period. We evaluated brood choices across a mixed landscape of grasslands managed for wildlife, agricultural croplands, cool-season pastures, and hayfields. We estimated population-level selection patterns within a Bayesian hierarchical integrated step-selection analysis (iSSA; Online Resource 1; Avgar et al. 2016). We evaluated both movement and resource utility by incorporating daily step length patterns into our evaluation of resource selection (Fortin et al. 2005; Forester et al. 2009; Thurfjell et al. 2014; Avgar et al. 2016). We also evaluated changes in selection patterns during the development period and the influence of variation in this behavior on the fate of young broods < 35 days old. Within our full resource selection models, we tested age- and fate-specific hypotheses. We had a priori expectations of differential brood responses to the following vegetation types:
-
(a)
Agriculture. Young broods would avoid agricultural croplands due to their high protein diet requirements and a lack of adequate invertebrate prey in cropland monocultures (Palmer et al. 2001). Accordingly, we expected successful young broods would avoid agriculture, while failed broods may show stronger selection for agriculture (Taylor and Burger 2000).
-
(b)
Idle native grasslands. Young broods would avoid native grasslands that were not burned or grazed within the previous two years due to juveniles’ small body size and limited mobility through thick, undisturbed vegetation (Doxon and Carroll 2010). We expected successful young broods to avoid idle native grasslands, while failed broods may show stronger selection for those grasslands.
-
(c)
Shrub cover. Broods would select for shrub cover throughout their development period as shrubs provide thermal protection and overhead cover from predators (Perkins et al. 2014; Carroll et al. 2015; Unger et al. 2015; Brooke et al. 2017). Successful young broods would show stronger selection for protective shrub cover than young broods that failed.
-
(d)
Tree composition. Broods would avoid tree cover and areas close to trees, as they provide perches for aerial predators and habitat for mesocarnivores, such as Red-tailed hawks (Buteo jamaicensis) and raccoons (Procyon lotor; Dinkins et al. 2012; Dzialak et al. 2012). Young flightless broods would select areas farther from trees compared to older broods who could flush from predators. Successful young broods would select areas with less tree cover and areas farther from trees compared failed broods (Lohr et al. 2011).
-
(e)
Daily movement. Daily distance travelled would be greater with age and successful broods would move shorter distances on average compared to failed broods (Lohr et al. 2011).
Methods
Study area
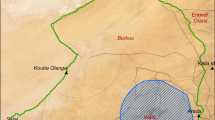
We tracked movement and resource use of bobwhite broods on five state conservation areas and neighboring private lands in southwest Missouri from 01 June to as late as 13 December 2016–2018 (Fig. 1a). Wah’Kon-Tah Prairie (1226 ha), Stony Point Prairie Conservation Area (388 ha), and Wade and June Shelton Memorial Conservation Area (129 ha) were extensive native grassland sites managed with fire, grazing, and mowing (Fig. 1b). Shawnee Trail (1471 ha) and Robert E. Talbot Memorial Conservation Areas (1764 ha) were intensively managed sites that incorporated narrow rows of strip crops and woody field borders among larger grassland units, providing food and cover for wildlife in smaller management units (2–24 ha; Fig. 1c). Native grasslands on conservation areas included remnant and reconstructed prairies composed of warm season grasses (e.g. Andropogon gerardii, Schizachyrium scoparium, and Sorghastrum nutans) and diverse, abundant forbs (e.g. Liatris pycnostachya, Echinacea pallida, and Solidago spp.). Mixed grasslands on conservation areas and private lands were composed of a mix of native warm season grasses, forbs (Ambrosia spp.), and non-native grasses (Festuca arundinacea, Dactylis glomerata, Phleum pratense). Private lands surrounding these conservation areas were primarily cool-season grazing pastures and hayfields (Festuca arundinacea, Dactylis glomerata, Phleum pratense), closed canopy woodland, with agricultural fields of corn (Zea mays), soybean (Glycine max), and winter wheat (Triticum spp.). Grassland management on conservation areas included prescribed burning, conservation grazing, disking, and woody and invasive species control (herbicide and mowing).

Study site locations on five conservation areas in southwest Missouri (left). Extensive native prairies (center) are larger continuous tracts of native grassland (dark grey) interspersed with shrubs and trees (black) and bordered by non-native grasslands (light grey). Intensively managed conservation areas (right) incorporate agricultural strips (white), woodland units and woody fencerows (black) within a matrix of restored native prairies (dark grey) and non-native mixed or cool-season grasslands (light grey)
Brood tracking and captures
We monitored resource selection and movement patterns of bobwhite broods daily using radio-telemetry. Most bobwhite broods were tracked from hatch. Some broods were opportunistically tracked post hatch when radio-tagged adults who did not participate in nest incubation were discovered attending broods. For broods ≤ 3 weeks old, habitat choices were determined by tracking radio-tagged attending adults. For broods > 3 weeks old, we used locations of radio-tagged juveniles to evaluate brood resource selection patterns if attending adults were depredated or abandoned their independent young. We found young brood mates tended to move together as a unit through the summer and into fall even without an attending adult present.
We captured broods 2–4 weeks old using the corral technique and a thermal imaging camera before first light (Smith et al. 2003). Our corrals were 1 m tall with a PVC pipe frame and panel walls made from extra-strength window screen. We also used garden netting as a cover for the corral to prevent individuals from flushing and escaping on capture. We sutured radio-tags on one to six juveniles per brood weighing > 20 g with 0.6 to 0.8 g transmitters (American Wildlife Enterprises, Monticello, FL, USA), which had 45–60 days of expected battery life (Terhune et al. 2020). Transmitters never exceeded 4% of a juvenile’s body mass. We attached transmitters low on the back, below the scapula along the thoracic vertebrae, and clipped antennae to 7 cm to avoid entanglement in grasses (Terhune et al. 2020).
Broods were tracked on average 6 days per week and observations with at least two consecutive daily locations were included in our resource selection analysis as a used step. We rotated the order of broods tracked and collected at least one roost location before first light each week. Broods were tracked to within 10 m of their location each day unless they were inaccessible, such as on restricted private lands, in which case locations were triangulated. Brood locations were projected from observer UTM coordinates and the distance and azimuth to the brood. We collected data on cover type, management, and woody vegetation at each location. All guidelines for the care and use of animals were followed and animal procedures were approved under University of Missouri Animal Care and Use Committee protocol #8766.
Cover type covariates
We included covariates representing the structural and compositional diversity of vegetation on conservation areas and their surrounding private lands. Classification of herbaceous cover types, management practices, and woody vegetation structure was informed by bobwhite ecological literature and applied management objectives. Herbaceous cover types included agricultural row crop (Ag), idle agricultural fields (Agid), native grass (N), mixed grass (M), and cool-season grass (C) and were characterized by percent cover within a 50 m radius for each used and available location (Online Resource Table S1). We further characterized grassland cover types by management practices using a 2-year burning (pb) and grazing (gr) history or a within-year mowing history (mw); we classified grasslands as idle (id) if no management had occurred over a two-year period (Online Resource Table S1). Burns occurred September–March prior to each breeding season, while grazing occurred April–September for 90- or 120-day periods during the brood season. For example, the category Npbgr classified native grasslands that were burned and grazed at least once either in the same year or two separate years within a two-year management history. Ngr classified native grasslands that were grazed at least once in the previous two years, but not burned. These included patches within patch-burn grazing systems that had not been burned in two years and units within conservation grazing-only systems. We hypothesized grasslands managed with fire, grazing or both may have greater utility than idle grasslands for bobwhite broods, as these disturbances increase bare ground, plant diversity and arthropod abundance (Taylor et al. 1999; Engle et al. 2008). Croplands were available to broods within intensively managed sites and on private lands surrounding all conservation areas except Wah’Kon-Tah. Intensively managed sites maintained small units of crops and food plots for wildlife use. Much of the private lands surrounding conservation areas in southwest Missouri, USA have been converted to pasture, hayfields, and cultivated crop. These cover types were largely classified from field observations of plant community composition and maps from area wildlife managers. Where field crew access was limited, some private lands were classified using 2018 CropScape and 2016 National Agriculture Imagery Program datasets (USDA 2016, 2018). We characterized woody vegetation composition and configuration as either percent shrub (Sh) or tree cover (Tr) within 50 m and by distance to nearest tree (TrD; m) from each used and available location. Shrub and tree cover were mapped using Light Detection and Ranging data. We created a digital surface model and a digital elevation model at 3.6 m resolution and calculated their difference to estimate vegetation height in ArcGIS. We classified elevations of 0–0.7 m as ground cover, 0.7–3.5 m as shrub cover, and 3.5–40 m as tree cover. We validated woody cover classification by comparing results with ground-truthed GPS points and aerial imagery (USDA 2016) across the study extent.
Used-available sampling design
We simultaneously evaluated effects of bobwhite brood age on habitat selection and daily movement decisions in a Bayesian, hierarchical conditional logistic regression that included both a resource utilization kernel and a movement kernel (Avgar et al. 2016). We incorporated spatial processes into a biologically meaningful sample of availability by drawing control points from an empirical distribution of observed movements (Fortin et al. 2005; Rhodes et al. 2005; Forester et al. 2009; Thurfjell et al. 2014). We sampled ten random, available step lengths from a log normal distribution based on observed daily movement distributions of each brood (Online Resource 2; Eq. S1, Eq. S2). We also sampled ten random, available step headings from a uniform distribution of all possible headings (0–2π radians). Vegetation types at end points of each used step were then compared to ten projected end points of each available step.
We included all daily brood step observations for our population-level, age-specific, step-selection analysis. To compare selection patterns between broods that failed and broods that succeeded, we only used step observations for broods ≤ 35 days old and we excluded broods whose fate could not be determined at 35 days old.
Hierarchical Bayesian selection model
We estimated the probability (P) brood (n) in age or fate class (k) would use location (j) with habitat features (m) at time (t) given a set (J) of available steps and locations(i) where i ≠ j using a conditional logistic regression (\(P\left( {x_{njktm} } \right)\); Eq. 1). Resource selection was attributed to multiple cover type and management predictors m observed at the end point of each daily used step H(\(x_{njtm}\)) and each daily available step H(\(x^{\prime}_{nitm}\)). Resource selection was quantified as a vector of cover type coefficients describing selection for each brood and each age or fate class (\(\beta_{nkm}\); Eq. 1). Movement patterns were estimated from observations of daily distance travelled (\(L_{njt}\)) and quantified in the movement coefficient vector \(\alpha_{nk}\) for each age and fate class. Variation in selection and movement among age classes and between broods that survived and broods that failed at 35 days were evaluated via an interaction with a class dummy variable, \(S_{nkt}\) (Eq. 1; Cooper and Millspaugh 1999; Bloom et al. 2013). The linear function representing the utility (\(U\)) of location \(j\) at time \(t\) to brood \(n\) for class \(k\), (\(U_{njtkm}\)), provided by a set of m habitat resources is \(U_{njktm} = \beta_{nkm} \cdot x_{njtm} \cdot S_{nkt}\).
We implemented our iSSA in a hierarchical Bayesian framework to account for individual- and population-level variations through random sampling (Gillies et al. 2006). Brood-level random regression coefficient vectors for resource selection, \(\beta_{nkm}\) and movement, \(\alpha_{nk}\), represented samples of population-level selection patterns, \(\beta_{km}^{*}\) and \(\alpha_{k}^{*}\) (Gillies et al. 2006; Thomas et al. 2006). This model structure included vague prior distributions for brood-level parameters (Eq. 2), and hyperparameters (Eq. 3; Thomas et al. 2006). Hyperparameter vectors, \(\beta_{km}^{*}\) and \(\alpha_{k}^{*}\), described mean population-level selection behavior and their precision, \(\sigma_{\beta km}^{*}\) and \(\sigma_{\alpha k}^{*}\) (Thomas et al. 2006). Random regression coefficients in the conditional logistic likelihood function address problems of non-independence among consecutive observations of individuals, differences in availability and preferences among individuals, and unbalanced data across individuals sampled (Gilles et al. 2006; Duchesne et al. 2010). This hierarchical structure allows inference on population-level patterns, by accounting for bias associated with unbalanced, correlated observations of individual animals, dependence of local availability on choice, and animal movement patterns (Online Resource 3; Gillies et al. 2006; Thomas et al. 2006; Duchesne et al. 2010).
Candidate model development and evaluation
We evaluated a set of cover type and management parameters representative of vegetation available on public and private lands. This set included agricultural row crop (Ag; corn, soybean, sunflower (Helianthus annuus), winter wheat), idle agriculture (Agid), native grasslands that were idle (Nid), grazed (Ngr), burned (Npb), or mowed/hayed (Nmw), and mixed grasslands that were idle (Mid), grazed (Mgr), burned (Mpb), or mowed (Mmw). We examined multicollinearity among habitat covariates within models and avoided combinations of covariates that resulted in a variable inflation factor (VIF) > 2.5. We excluded cool-season grasslands to avoid multicollinearity (Online Resource 4).
We evaluated age- and fate-specific resource selection behaviors for agriculture, idle native grasslands, shrub (Sh) and tree cover (Tr), distance to trees (TrD), and daily step length distributions (Step; Table S1). This set of available cover types was evaluated as the most parsimonious variables addressing (1) the value of agricultural units available to broods on intensively managed sites and surrounding private lands, (2) the importance of disturbance on native grassland utility, (3) the influence of woody composition on brood choice, and (4) changes in habitat availability with growth and development. To evaluate age-specific variation in resource selection, brood stages were classified based on the ages at which young can fly short distances (≤ 14 days) and females may abandon a brood to attempt a second nest (35 days). A three-stage model included broods that were flightless ≤ 14 days old, dependent 14–35 days old, and independent > 35 days old. A two-stage model compared selection behavior of young broods ≤ 35 days old and old broods > 35 days old. A single-stage model included no age class information. To evaluate the influence of choice on survival of young, we subset our data to include only observations of broods whose fate at 35 days was known. We then compared selection behavior of successful and failed broods ≤ 35 days old.
We fit resource selection models in a Bayesian framework in Program R version 3.6.1 (R Core Development Team 2017) using JAGS (Plummer 2003) via the JagsUI package (Kellner 2019). All models evaluating age- and fate-dependent patterns in resource selection were run with 3 chains of 30,000 iterations, a burn-in of 10,000 iterations, and a thinning rate of 4, which resulted in 15,000 total posterior samples for each parameter. We inspected trace plots for mixing across Markov chains, checked that R-hat values were < 1.1 (Brooks and Gelman 1998). We evaluated model performance by ranking models based on Widely Applicable Information Criterion and interpreting the top model (WAIC, Watanabe 2010). The proportions of posterior samples that do not span zero (f value) indicate the strength of habitat selection (positive) or avoidance (negative). The magnitude and uncertainty of age- and fate-dependent patterns in resource selection were quantified as the proportion of posterior distributions non-overlapping between age or fate classes.
Results
Population-level brood resource selection and movement
We monitored resource selection patterns of 101 bobwhite broods for a total of 2790 step choice sets from 2016 to 2018. These data contained 627 steps from 80 broods ≤ 14 days old, 1092 steps from 91 broods 15 to 35 days old, and 1071 steps from 45 broods 36 to 114 days old. Brood step length was not correlated with variation in hours between daily tracking locations; therefore, we did not include time within resource selection models (Online Resource Figure S1). Ranges of habitat and step length data for used and available locations are summarized in Online Resource Table S1.
All resource selection models performed better than the null model (Online Resource Table S2). The model containing 3 stage-specific estimates for resource selection and step length distribution performed better than the 2-stage and the null model that did not consider effects of age class (∆WAIC = 35.8, 105.2, respectively; Online Resource Table S2). Therefore, we focused interpretation on the top-ranked model that included parameters for resource selection and step length distribution for three stages: flightless broods ≤ 14 days old, dependent broods 15 to 35 days old, and independent broods > 35 days old.
Broods showed the strongest selection among native grasslands for areas that were burned and grazed within the past two years (Npbgr = 0.41, 95% CRI 0.24, 0.57, f = 1.00), followed by areas burned only (Npb = 0.31, 95% CRI 0.05, 0.54 f = 0.99), and mowed (Nmw = 0.21, 95% CRI 0.05, 0.35, f = 0.99; Fig. 2, Fig. 3a, b, Online Resource Table S3). Broods showed weak selection for native grasslands that were grazed only (Ngr = 0.12, 95% CRI = − 0.11, 0.30, f = 0.88; Fig. 2, Online Resource Table S3). Broods > 35 days old selected for idle native grasslands (Nid[2] = 0.36, 95% CRI = 0.05, 0.63, f = 0.99), whereas broods ≤ 14 days old did not select for idle native grasslands (Nid[0] = − 0.16, 95% CRI = − 0.73, 0.22, f = 0.74; Fig. 2, Fig. 3c, Online Resource Table S3). Broods > 35 days showed stronger selection for idle native grasslands than broods ≤ 14 days old (proportion non-overlapping posteriors 0.843; Table S4). Among mixed grasslands, bobwhite selected for areas mowed in that year (Mmw = 0.29, 95% CRI = 0.15, 0.42, f = 1.00). All age classes were selected for agricultural row crop and idle agricultural fields (Ag[0] = 0.39, 95% CRI = 0.12, 0.66; Ag[1] = 0.36, 95% CRI = 0.12, 0.58; Ag[2] = 0.52, 95% CRI = 0.33, 0.69, f = 1.00; Fig. 2, Fig. 3d, Online Resource Table S3, Table S4). Among woody habitat components, broods were selected for shrub cover and avoided locations further from trees (Figs. 2, 3e, f, Online Resource Table S3). Selection for shrub cover and avoidance of areas farther from trees was stronger for older broods than younger broods (0.968 and 0.871 non-overlapping posteriors, respectively; Figs. 2, 3e, f, Online Resource Table S4). Broods ≤ 14 days old took shorter steps (mean = 129 m) and travelled < 200 m in a day more frequently than older age classes, while broods > 35 days old took longer steps (mean = 167) and travelled > 200 m more frequently than younger age classes (1.00 non-overlapping posteriors; Fig. 4, Online Resource Table S3, Table S4). The 95th percentile of our step length distribution curves for young flightless broods ≤ 14 days old, dependent broods 15–35 days old, and independent broods > 35 days were 309 m, 334 m, and 404 m, respectively.

Posterior distributions of population-level habitat selection parameters estimating utility of agricultural fields (Ag), native grasslands (N), and mixed grasslands (M) that were idle (id), grazed (gr), burned (pb), grazed and burned (pbgr), or mowed (mw), as well as brood selection for shrub (Sh) and tree cover (Tr), and selection for distance from tree (TrD) for northern bobwhite broods in southwest Missouri 2016–2018. The solid line at y = 0 represents the threshold between support for selection (+) or avoidance (−) of each habitat parameters. Stage-specific effects S[−] are presented with black mean point estimates for broods ≤ 14 days old [0] (black circle), 14 to 35 days old [1] (black triangle), and > 35 days old [2] (black square) and 95% CRI (bar). Habitat selection parameters that are stage-independent display mean point estimates in white (white square) and 95% CRI (bar)

Predicted relative probability of use of habitats by northern bobwhite broods in southwest Missouri 2016–2018 as a function of: a percent cover within 50 m of native grassland that was burned and grazed, or b only burned and not grazed within the past two years; and stage-dependent predicted relative probability of use as a function of c percent cover within 50 m of idle native grassland, d agricultural crop, e shrub cover, and f predicted relative probability of use as a function of distance to nearest tree for young flightless broods ≤ 14 days old (S[0]; dotted line), dependent broods 14 to 35 days old (S[1]; dashed line), and independent broods > 35 days old (S[2]; solid line)

Distributions of daily distanced travelled by northern bobwhite broods in southwest Missouri 2016–2018 derived from the mean, 2.5, and 97.5% credible intervals of posterior samples of the step length parameter. Stage-specific step length distributions for broods ≤ 14 days old (S[0], dotted line, light grey), 14–35 days old (S[1], dashed line, medium grey), and > 35 days old (S[2], solid line, dark grey). Mean predicted daily distance travelled for each stage are plotted with point estimates for broods ≤ 14 days old (black circle), 15–35 days old (black triangle), and > 35 days old (black square)
Resource selection and movement patterns of successful and failed broods
We evaluated resource selection and movement of 80 broods ≤ 35 days old to compare patterns between broods that did and did not survive to 35 days old. These data contained 273 steps from 29 failed broods, F[0], and 1187 steps from 51 successful broods, F[1]. We removed 21 broods from this analysis because brood fate was unknown due to attending adult mortality, dropped radio-tags, or because we were unable to track broods on restricted private lands.
Our top model suggested brood resource selection differed between broods that failed and broods that succeeded in surviving to 35 days because it performed better than our null model without fate classes (∆WAIC = 2.3; Online Resource Table S2). We did not detect a difference in selection for agricultural cover, idle native grassland cover, tree cover, or daily step lengths between broods that failed and broods that survived to 35 days; however, there were differences in selection for shrub cover and distance to trees (Fig. 5, Online Resource Table S3, Table S5). Broods that failed weakly avoided shrub cover (Sh[0] = − 0.21, 95% CRI = − 0.59, 0.129, f = 0.88) and were less likely to choose habitats with greater percent cover compared to successful broods (0.921% posterior support; Online Resource Table S5), while broods that succeeded selected for shrub cover (Sh[1] = 0.19, 95% CRI = 0.039, 0.32, f = 0.99) and were more likely to use habitat with greater percent cover within a 50 m radius (80.7% posterior support; Fig. 6, Online Resource Table S3, Table S5). Both failed and successful broods avoided areas further from trees than randomly available (Fig. 5, Online Resource Table S3). However, broods that failed were more likely to select areas closer to trees than broods that succeeded (76.4% posterior support; Figs. 5, 6, Online Resource Table S3, Table S5).

Split violin plots showing distributions of habitat parameters evaluating selection patterns of northern bobwhite broods in southwest Missouri that failed and broods that survived to 35 days old. Cover type effects for agriculture (Ag), idle native grasslands (Nid), shrub (Sh) and tree (Tr) cover, and distance to nearest tree (TrD) are presented as split violins for failed (left, grey) and successful (right, white) broods [0|1]. Means for failed broods (grey triangle) and successful broods (white triangle) and 95% CRI (bar) are shown in violin plots

Fate-dependent predicted relative probability of use for northern bobwhite broods in southwest Missouri 2016–2018 as a function of a percent shrub cover within 50 m and b distance to nearest tree for successful broods that survived to 35 days old (F[1]; solid line) and broods that failed before 35 days old (F[0]; dot-dash line)
Discussion
Bobwhite broods showed the strongest selection for native grasslands that were both burned and grazed at least once within the previous two years. We also found resource selection patterns changed with age. Young broods avoided idle native grasslands, while older broods selected for those cover types. Selection for shrubs increased with age and older broods selected areas closer to trees compared to young broods. We found variation in these choices had consequences on early-life survival. Selection for woody cover differed between young broods that survived to 35 days and young broods that failed. Broods that survived were more likely to choose areas with greater percent shrub cover and areas farther from trees than broods that failed. We evaluated movement as a behavioral component of resource selection and found daily distance travelled increased with age, but movement patterns did not differ between broods that succeeded and broods that failed to 35 days.
Population-level habitat selection
Coupling grazing and fire on tallgrass prairies and grasslands promote heterogeneity in vegetation structure and composition (Fuhlendorf and Engle 2004; Veen et al. 2008). In Missouri, this practice of patch-burn grazing is implemented with 1/3 of a unit burned (Oct–Mar) and then grazed (Apr–Aug) for 90–120 days. In subsequent years, another third is burned until a given rest year or until the cycle is repeated. This promotes a shifting mosaic that facilitates brood use since at least one-third to one-half of an area has been disturbed in the last 2 years. Broods likely selected for burned and grazed native grasslands because those diverse, disturbed prairie plant communities provided adequate cover, open bare ground, and high invertebrate abundance for improved juvenile foraging, growth, mobility, and survival (Hurst 1972; Taylor et al. 1999; Engle et al. 2008; Doxon and Carroll 2010; Kamps et al. 2017).
We also found broods selected for native grasslands that were burned, but not grazed in the previous two years. Small, patchy prescribed fires remove accumulated litter to create open foraging habitat in close proximity to thicker roosting habitat (Taylor et al. 1999; Kamps et al. 2017). This can reduce brood home range size, which is indicative of improved habitat quality (De Vos and Mueller 1993). We also found weaker selection for native grasslands that were grazed, but not burned in the previous two years. Light to moderate grazing that leaves adequate vegetation height for cover can benefit broods, but fire is an important component of grassland disturbance regimes (Sutter and Ritchison 2005; Harper et al. 2015).
Contrary to our predictions, we found broods selected for both native and mixed grasslands that were mowed or hayed within a breeding season. We expected broods would avoid cut grasses because these practices remove cover, have greater litter if not hayed, and may not improve plant diversity or habitat structure for bobwhite broods (McCoy et al. 2001; Taylor et al. 1999; Gruchy and Harper 2014). Mowing/haying may negatively influence survival through direct mortality of broods or indirectly through increased rates of depredation of young in vegetation that reduces cover (Bollinger et al. 1990). Timing of the management practice may explain differences between predicted and observed effects in our study. Mowing and haying of native grasses typically happened in late summer after July 1 and after peak nesting season, meaning units were not cut for much of the brood season (Harper et al. 2007; Birckhead et al. 2014). Mowing also only changes vegetation structure temporarily. Warm-season grass canopy can grow to heights that provide adequate cover for broods within a few weeks of harvest (Birckhead et al. 2014).
Age-dependent mobility and habitat selection
Non-environmental factors can influence resource selection patterns (Jones 2001). For precocial young that leaves the nest soon after hatching, age strongly influences development and mobility. Individual physical condition can affect resource selection by influencing perceived habitat quality and how an individual moves across a landscape (Merrick and Koprowski 2017). The first 35 days of life are a critical period for success of bobwhite broods and during this time broods may exhibit resource requirements unique to later stages of development (Lusk et al. 2005; Hannon and Martin 2006). We found that bobwhite broods exhibited age-specific differences in movement and habitat selection.
Age influenced daily distances travelled by broods. We found the radius of habitat available to broods older than 35 days increased by just over 30% within a 114-day development period. Taylor and Guthery (1994) reported daily home range size and minimum daily distances traveled nearly doubled from pre-fledging to post-fledging brood periods (0.7 ha and 277 m to 1.4 ha and 589 m, respectively). Post-fledging broods may travel greater distances and may be better able to explore within their daily home ranges than pre-fledging broods. Age can also influence the relative quality of different habitats as resource needs and risk perception change with development. Small flightless young survive to stages where they can thermoregulate, flush from predators, modify foraging behavior and diet, and potentially better use new habitats. Age and mobility thereby interact with resource selection to influence selective pressures, such as body condition, predation, and exposure (Merrick and Koprowski 2017).
All age classes of bobwhite broods in our study were selected for row crop and idle agricultural fields. We expected young broods to avoid row crops due to poor insect prey abundance or availability (Taylor and Burger 2000). Foraging rates and growth of juvenile bobwhite are low within soybean, corn, and wheat fields compared to field borders and native grasslands, suggesting cropland provides suboptimal foraging habitat (Puckett et al. 1995; Palmer et al. 2001; Doxon and Carroll 2010; Lohr et al. 2011). However, grassland vegetation strips within strip crop units or along field borders may provide adjacent foraging habitat and pesticide restrictions on public lands resulting in weedy fields may provide sufficient insect prey (Doxon and Carroll 2007). Selection for agriculture may also be related to risk avoidance (Dzialak et al. 2012). In crop fields, broods can move easily along the ground, sight approaching ground predators, and have adequate canopy height for cover from aerial predators. While agricultural areas may provide marginal foraging habitat, predator avoidance and improved juvenile survival may be a fitness benefit explaining selection patterns (Palmer et al. 2001; Sinnott 2020).
Bobwhite broods are less likely to occupy fields with dense grass and litter cover (Taylor et al.1999; Osborne et al. 2012). We hypothesized thick grass and litter cover would be more limiting to small, flightless juveniles compared to larger, more mobile broods. We found young broods did not select for idle native grasslands, while older broods did select for those grasslands. Size is an important factor mediating age-dependent habitat utility. With growth, older bobwhite broods were better able to move, forage, and utilize thick idle native grasslands (Doxon and Carroll 2010).
Woody vegetation composition and structure are important components of brood habitat for this shrub-obligate species (Martin et al. 2009; Unger et al. 2015). We found young broods did not select for shrub cover, but young broods that survived to 35 days old and broods > 35 days old did select for shrub cover. Shrubs provide protection from warm daytime temperatures and escape cover from predators (Carroll et al. 2015). We also found both younger broods and older broods selected areas closer to trees than randomly available, but, broods > 35 days old selected areas closer to trees compared to broods ≤ 14 days old. Young broods may avoid areas closer to trees than older broods because they cannot yet flush from predators; predator avoidance strategies may depend on age-specific behavioral responses to threats (Dinkins et al. 2012; De Cesare et al. 2014; Perkins et al. 2014).
Fitness consequences of resource selection behavior
While preferred habitats are often assumed to provide fitness benefits, ecological or perceptual traps may result in selection patterns of individuals or populations that do not optimize important demographic rates (Patten and Kelly 2010; Dzialak et al. 2011; De Cesare et al. 2014; Merrick and Koprowski 2017). Few studies link resource selection behaviors with demographic responses by examining survival consequences of variation in choices among individuals (Jones 2001; Bloom et al. 2013; De Cesare et al. 2014; Merrick and Koprowski 2017). Recent studies have found vegetation composition, management, and habitat suitability can influence survival of bobwhite young (Kamps et al. 2017; Tanner et al. 2019; Sinnott 2020). However, these studies have not addressed demographic consequences of individual-level variation in habitat selection behavior. We found daily step lengths of bobwhite broods that failed were similar to movement choices made by broods that survived to 35 days. Other studies have reported larger movements early in the brood period and greater dispersal distances in fragmented landscapes reduce brood survival (Rotella and Rati 1992; Hannon and Martin 2006). Alternatively, larger brood movements from nest sites may result in better habitat for greater brood survival (Paasivaara and Pöysä 2008). While daily movements were not significantly different among successful and failed broods in our study, more work is needed linking habitat-specific movement and dispersal patterns to bobwhite brood survival.
Selection for agricultural row crop, idle native grassland, and tree cover was similar for broods that succeeded and broods that failed to 35 days. However, selection for woody cover differed. Successful broods chose habitats with more shrub cover, while failed broods weakly avoided those areas. Attending adults choosing areas with more shrub cover are providing broods taller vegetation for greater visual obstruction from aerial predators (Hannon and Martin 2006; Collins et al. 2009; Perkins et al. 2014). Taller vegetation can also create fine-scale thermal refuges during summer daytime peak temperatures; failure to choose more moderate microclimates under shrubby vegetation may lead to thermal stress for young (Carroll et al. 2015). While all young broods selected areas closer to trees than randomly available, successful broods selected areas farther from trees than failed broods, suggesting bobwhite mortality risk is greater for those using mature wooded habitat (Lohr et al. 2011).
Conclusion
Three questions related to patterns of resource selection and movement decisions can help us more robustly understand animal distributions: (1) which habitats are animals choosing? (2) what influences choice? (3) what are the fitness consequences of variation in choice? Patterns of resource selection and movement contain important information about habitat valuation and risk aversion, and the extent to which these are driven by age, season and landscape composition to explain fitness outcomes. We found shrub cover and native grasslands managed with grazing and fire were especially valuable for success of young broods. We encourage researchers to examine hypothesized drivers of successful and failed strategies in animals for deeper learning about how fitness cascades to demographic outcomes.
References
Atuo FA, O’Connell TJ (2018) Superpredator proximity and landscape characteristics alters nest site selection and breeding success of a subordinate predator. Oecologia 186:817–829
Avgar T, Potts JR, Lewis MA, Boyce MS (2016) Integrated step selection analysis: bridging the gap between resource selection and animal movement. Methods Ecol Evol 7:619–630
Birckhead JL, Harper CA, Keyser PD, McIntosh D, Holcomb ED, Bates GE, Waller JC (2014) Structure of avian habitat following hay and biofuels production in native warm-season grass stands in the Mid-South. J Southeas Assoc Fish Wildl Agencies 1:115–121
Bloom PM, Clark RG, Howerter DW, Armstrong LM (2013) Multi-scale habitat selection affects offspring survival in a precocial species. Oecologia 173:1249–1259
Bollinger EK, Bollinger PB, Gavin TA (1990) Effects of hay-cropping on eastern populations of the Bobolink. Wildl Soc Bull 18:142–150
Brooke JM, Peters DC, Unger AM, Tanner EP, Harper CA, Keyser PD, Clark JD, Morgan JJ (2015) Habitat manipulation influences northern bobwhite resource selection on a reclaimed surface mine. J Wildl Manag 79:1264–1276
Brooke JM, Tanner EP, Peters DC, Tanner AM, Harper CA, Keyser PD, Clark JD, Morgan JJ (2017) Northern bobwhite breeding season ecology on a reclaimed surface mine. The J Wildl Manag 81:73–85
Brooks SP, Gelman A (1998) General methods for monitoring convergence of iterative simulations. J Comput Graph Stat 7:434–455
Carroll JM, Davis CA, Elmore RD, Fuhlendorf SD, Thacker ET (2015) Thermal patterns constrain diurnal behavior of a ground-dwelling bird. Ecosphere 7:e01403
Collins BM, Williams CK, Castelli PK (2009) Reproduction and microhabitat selection in a sharply declining northern bobwhite population. Wilson J Ornithol 121:688–695
Cooper AB, Millspaugh JJ (1999) The application of discrete choice models to wildlife resource selection studies. Ecology 80:566–575
De Vos T, Mueller BS (1993) Reproductive ecology of northern bobwhite in North Florida. In: Church KE, Dailey TV (eds) Proc Natl Quail Symp III. Kansas Department of Wildlife and Parks, Pratt, pp 83–90
De Cesare NJ, Hebblewhite M, Bradley M, Hervieux D, Neufeld L, Musiani M (2014) Linking habitat selection and predation risk to spatial variation in survival. J Anim Ecol 83:343–352
Dinkins JB, Conover MR, Kirol CP, Beck JL (2012) Greater Sage-Grouse (centrocercus urophasianus) select nest sites and brood sites away from avian predators. Auk 129:600–610
Doxon ED, Carroll JP (2007) Vegetative and invertebrate community characteristics of Conservation Reserve Program fields relative to gamebirds in western Kansas. Am Midl Nat 158:243–259
Doxon ED, Carroll JP (2010) Feeding Ecology of ring-necked pheasant and northern bobwhite chicks in Conservation Reserve Program fields. J Wildl Manag 74:249–256
Duchesne T, Fortin D, Courbin N (2010) Mixed conditional logistic regression for habitat selection studies. J Anim Ecol 79:548–555
Dzialak MR, Olson CV, Harju SM, Webb SL, Mudd JP, Winstead JB, Hayden-Wing LD (2011) Identifying and prioritizing greater sage-grouse nesting and brood-rearing habitat for conservation in human-modified landscapes. PLoS ONE 6(10):e26273. https://doi.org/10.1371/journal.pone.0026273
Dzialak MR, Olson CV, Harju SM, Webb SL, Winstead JB (2012) Temporal and hierarchical spatial components of animal occurrence: conserving seasonal habitat for greater sage-grouse. Ecosphere 3:30. https://doi.org/10.1890/ES11-00315.1
Engle DM, Fuhlendorf SD, Roper A, Leslie DM Jr (2008) Invertebrate community response to a shifting mosaic of habitat. Rangel Ecol Manag 61:55–62
Forester JD, Im HK, Rathouz PJ (2009) Accounting for animal movement in estimation of resource selection functions: sampling and data analysis. Ecology 90:3554–3565
Fortin D, Beyer HL, Boyce MS, Smith DW, Duchesne T, Mao JS (2005) Wolves influence elk movements: behavior shapes a trophic cascade in Yellowstone National Park. Ecology 86:1320–1330
Fuhlendorf SD, Engle DM (2004) Application of the fire-grazing interaction to restore a shifting mosaic on tallgrass prairie. J Appl Ecol 41:604–614
Gillies CS, Hebblewhite M, Nielsen SE, Krawchuk MA, Aldridge CL, Frair JL, Saher DJ, Stevens CE, Jerde CL (2006) Application of random effects to the study of resource selection by animals. J Anim Ecol 75:887–898
Goddard AD, Dawson RD (1999) Factors influencing the survival of neonate sharp-tailed grouse Tympanuchus phasianellus. Wildl Biol 15:60–67
Gruchy JP, Harper CA (2014) Effects of field management practices on northern bobwhite habitat. J Southeast Assoc Fish Wildl Agenc 1:133–141
Hannon SJ, Martin K (2006) Ecology of juvenile grouse during the transition to adulthood. J Zool 269:422–433
Harper CA, Bates GE, Hansbrough MP, Gudlin MJ, Gruchy JP, Keyeser PD (2007) Native warm-season grasses identification, establishment, and management for wildlife and forage production in the Mid-South. University of Tennessee Extension, Knoxville
Harper CA, Birckhead JL, Keyser PD, Waller JC, Backus MM, Bates GE, Holcomb ED, Brooke JM (2015) Avian habitat following grazing native warm-season grass forages in the Mid-South United States. Rangel Ecol Manag 68:166–172
Hernández F, Brennan LA, DeMaso SJ, Sands JP, Wester DB (2013) On reversing the Northern bobwhite population decline: 20 years later. Wildl Soc Bull 37:177–188
Hurst GA (1972) Insects and bobwhite quail brood habitat management. Proc Natl Quail Symp I 1:65–82
Jenkins JMA, Thompson FR III, Faaborg J (2017) Behavioral development and habitat structure affect postfledging movements of songbirds. J Wildl Manag 81:144–153
Jones J (2001) Habitat selection studies in avian ecology: a critical review. Auk 118:557–562
Kamps JT, Palmer WE, Terhune TM, Hagan G, Martin JA (2017) Effects of fire management on northern bobwhite brood ecology. Europ J Wildl Res 63:27
Kane K, Sedinger JS, Gibson D, Blomberg E, Atamian M (2017) Fitness landscapes and life-table response experiments predict the importance of local areas to population dynamics. Ecosphere 8:e01869. https://doi.org/10.1002/ecs2.1869
Kellner K (2019) jagsUI: a wrapper around 'rjags' to streamline 'JAGS' analyses. R package version 1.5.1. https://CRAN.R-project.org/package=jagsUI
Kentie R, Hooijmeijer JCEW, Trimbos KB, Groen NM, Piersma T (2013) Intensified agricultural use of grasslands reduces growth and survival of precocial shorebird chicks. J Appl Ecol 50:243–251
Klimstra WD, Roseberry JL (1975) Nesting ecology of the bobwhite in Southern Illinois. Wildl Monogr 41:3–37
Lima SL, Zollner PA (1996) Towards a behavioral ecology of ecological landscapes. Trends Ecol Evol 11:131–135
Lohr M, Collins BM, Castelli PM, Williams CK (2011) Life on the edge: northern bobwhite ecology at the northern periphery of their range. J Wildl Manag 75:52–60
Lusk JJ, Guthery FS, Cox SA, Demaso SJ, Peoples AD (2005) Survival and growth of northern bobwhite chicks in western Oklahoma. Am Midl Nat 153:389–395
Mainguy J, Gauthier G, Giroux JF, Duclos I (2006) Habitat use and behaviour of Greater Snow Geese during movements from nesting to brood-rearing areas. Can J Zool 84:1096–1103
Martin NC, Martin JA, Carroll JP (2009) Northern bobwhite brood habitat selection in South Florida. In: Cederbaum SB, Faircloth BC, Terhune TM, Thompson JJ, Carroll JP (eds) Proc Natl Quail Symp VI, pp 88–97
Mayor SJ, Schneider DC, Schaefer JA, Mahoney SP (2009) Habitat selection at multiple scales. Ecoscience 16:238–247
McConnell MD, Monroe AP, Chandler R, Palmer WE, Wellendorf SD, Burger LW Jr, Martin JA (2018) Factors influencing Northern bobwhite recruitment, with implications for population growth. Auk 135:1087–1099
McCoy TD, Kurzejeski EW, Burger LW Jr, Ryan MR (2001) Effects of conservation practice, mowing, and temporal changes on vegetation structure on CRP fields in northern Missouri. Wildl Soc Bull 29:979–987
Merrick MJ, Koprowski JL (2017) Should we consider individual behavior differences in applied wildlife conservation studies? Biol Conserv 209:34–44
Osborne DC, Sparling DW, Hopkins RL II (2012) Influence of Conservation Reserve Program mid-contract management and landscape composition on northern bobwhite in tall fescue monocultures. J Wildl Manag 76:556–574
Paasivaara A, Pöysä H (2008) Habitat-patch occupancy in the common goldeneye (Bucephalaclangula) at different stages of the breeding cycle: implications to ecological processes in patchy environments. Can J Zool 86:744–755
Palmer WE, Lane MW II, Bromley PT (2001) Human-imprinted northern bobwhite chicks and indexing arthropod foods in habitat patches. J Wildl Manag 65:861–870
Patten MA, Kelly JF (2010) Habitat selection and the perceptual trap. Ecol Appl 20:2148–2156
Perkins R, Boal C, Rollins D, Perez RM (2014) Northern bobwhite predator avoidance behavior in response to varying types of threat. J Wildl Manag 78:1272–1281
Plummer M (2003) JAGS: a program for analysis of Bayesian graphical models using Gibbs sampling. In: 3rd International workshop on distributed statistical computing (DSC 2003); Vienna, Austria, p 124
Puckett KM, Palmer WE, Bromley PT, Anderson JR Jr, Sharpe TL (1995) Bobwhite nesting ecology and modern agriculture: a management experiment. Proc Annu Conf Southeast Assoc Fish Wildl Agencies 49:505–515
R Core Team (2017) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Rhodes JR, McAlpine CA, Lunney D, Possingham HP (2005) A spatially explicit habitat selection model incorporating home range behavior. Ecology 86:1119–1205
Rotella JJ, Ratti JT (1992) Mallard brood movements and wetland selection in southwestern Manitoba. J Wildl Manag 56:508–515
Sandercock BK, Alfaro-Barrios M, Casey AE, Johnson TN, Mong TW, Odom KJ, Strum KM, Winder VL (2014) Effects of grazing and prescribed fire on resource selection and nest survival of upland sandpipers in an experimental landscape. Landsc Ecol 30:325–337
Schooley RL (1994) Annual variation in habitat selection: patterns concealed by pooled data. J Wildl Manag 58:367–374
Sinnott EA (2020) PhD dissertation, School of Natural Resources, University of Missouri, Columbia, Missouri, USA
Smith MD, Hammond AD, Burger LW, Palmer WE, Carver AV, Wellendorf SD (2003) A technique for capturing northern bobwhite chicks. Wildl Soc Bull 31:1054–1060
Sonerud GA (1985) Brood movements in grouse and waders as defence against winstay search in their predators. Oikos 44:287–300
Sutter B, Ritchison G (2005) Effects of grazing on vegetation structure, prey availability, and reproductive success of Grasshopper Sparrows. J Field Ornithol 76:345–351
Tanner EP, Elmore RD, Fuhlendorf SD, Davis CA, Dahlgren DK, Orange JP (2017) Extreme climatic events constrain space use and survival of a ground-nesting bird. Glob Change Biol 23:1832–1846
Tanner EP, Orange JP, Davis CA, Elmore RD, Fuhlendorf SD (2019) Behavioral modifications lead to disparate demographic consequences in two sympatric species. Ecol Evol 9:9273–9289
Taylor JD II, Burger LW Jr (2000) Habitat use by breeding northern bobwhites in managed old-field habitats in Mississippi. In: Brennan LA, Palmer WE, BurgerPruden LWTL (eds) Proc Natl Quail Symp IV. Tall Timbers Research Station, Tallahassee, Florida, pp 7–15
Taylor JS, Guthery FS (1994) Daily movements of northern bobwhite broods in southern Texas. Wilson Bull 106:148–150
Taylor JS, Church KE, Rusch DH (1999) Microhabitat selection by nesting and brood-rearing northern bobwhite in Kansas. J Wildl Manag 63:686–694
Terhune TM II, Caudill D, Terhune VH, Martin JA (2020) A modified suture technique for attaching radiotransmitters to Northern bobwhite. Wildl Soc Bull 44:1–10
Thomas DL, Johnson D, Griffith B (2006) A Bayesian random effects discrete-choice model for resource selection: population-level selection inference. J Wildl Manag 70:404–413
Thurfjell H, Ciuti S, Boyce MS (2014) Applications of step-selection functions in ecology and conservation. Mov Ecol 2:4
Unger AM, Tanner EP, Harper CA, Keyser PD, Van Manen FT, Morgan JJ, Baxley DL (2015) Northern bobwhite seasonal habitat selection on a reclaimed surface coal mine in Kentucky. J Southeast Assoc Fish Wildl Agencies 2:235–246
USDA-FSA-APFO Aerial Photography Field Office (2016) National Geospatial Data Asset (NGDA) NAIP Imagery. http://gis.apfo.usda.gov/arcgis/rest/services/NAIP
USDA National Agricultural Statistics Service Cropland Data Layer (2018) Published crop-specific data layer [Online]. https://nassgeodata.gmu.edu/CropScape/. verified 11/01/2018). USDA-NASS, Washington, DC. Accessed 11 Jan 2018
Veen GF, Blair JM, Smith MD, Collins SL (2008) Influence of grazing and fire frequency on small-scale plant community structure and resource variability in native tallgrass prairie. Oikos 117:859–866
Watanabe S (2010) Asymptotic equivalence of Bayes cross validation and widely applicable information criterion in singular learning theory. J Mach Learn Res 11:3571–3594
Wiltermuth MT, Anteau MJ, Sherfy MH, Pearse AT (2015) Habitat selection and movements of Piping Plover broods suggest a tradeoff between breeding stages. J Ornithol 156:999–1013
Acknowledgements
This project was funded by Missouri Department of Conservation with additional support from the University of Missouri and USDA Forest Service Northern Research Station. KJ Botzet, JE Guenther, JH Heuschkel, J Huang, JR Jones, LL McElroy, WI Payette, JA Swift, TB Thorne, MA Wheeler, NC Yerden, AL Zak contributed to field data collection. NC Burrell, RK Hedges, MA Hill, FL Loncarich, JR Steele, and SD Whitaker provided maps of cover type and management histories. JS Fraser and WD Dijak helped derive habitat metrics. JM Jenkins, TW Bonnot, and T Avgar assisted with model development.
Author information
Authors and Affiliations
Contributions
FRT, TRT originally formulated the idea, EAS, MDW, TRT, FRT developed methodology, EAS, TRT conducted fieldwork, EAS, FRT developed mathematical models, and EAS, FRT, MDW, TRT wrote the manuscript.
Corresponding author
Additional information
Communicated by Markku Orell.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Sinnott, E.A., Weegman, M.D., Thompson, T.R. et al. Resource selection and movement by northern bobwhite broods varies with age and explains survival. Oecologia 195, 937–948 (2021). https://doi.org/10.1007/s00442-021-04893-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-021-04893-z