
Abstract
Plant size is a major predictor of ecological functioning. We tested the hypothesis that feeding damage to plants increases with plant size, as the conspicuousness of large plants makes resource finding and colonisation easier. Further, large plants can be attractive to herbivores, as they offer greater amounts and ranges of resources and niches, but direct evidence from experiments testing size effects on feeding damage and consequently on plant fitness is so far missing. We established a common garden experiment with a plant size gradient (10–130 cm height) using 21 annual Brassicaceae species, and quantified plant size, biomass and number of all aboveground components (flowers, fruits, leaves, stems) and their proportional feeding damage. Plant reproductive fitness was measured using seed number, 1000 seed weight and total seed weight. Feeding damage to the different plant components increased with plant size or component biomass, with mean damage levels being approximately 30 % for flowers, 5 % for fruits and 1 % for leaves and stems. Feeding damage affected plant reproductive fitness depending on feeding damage type, with flower damage having the strongest effect, shown by greatly reduced seed number, 1000 seed weight and total seed weight. Finally, we found an overall negative effect of plant size on 1000 seed weight, but not on seed number and total seed weight. In conclusion, being conspicuous and attractive to herbivores causes greater flower damage leading to higher fitness costs for large plants, which might be partly counterbalanced by benefits such as enhanced competitive/compensatory abilities or more mutualistic pollinator visits.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Body size is of great importance in most organisms at physiological, genetic and ecological levels (Peters 1983). Large body size has various inherent benefits such as high dispersal ability (Hemptinne et al. 2012) and high competitiveness (Brown and Maurer 1986), but also has costs such as higher energy requirements or high conspicuousness to enemies (Blanckenhorn 2000; Remmel and Tammaru 2009). Large plants are very apparent and offer a wide range of resources and niches to associated animals, and are consequently easy to locate and attractive for herbivores (Feeny 1976; Lawton 1983). This can lead to increasing herbivore species richness with increasing plant size, as shown by several studies for naturally grown plants, where focus was on single groups of associated animals and mainly on an intraspecific plant size gradient (Lawton and Price 1979; Neuvonen and Niemelä 1981; Haysom and Coulson 1998).
To analyse the effects of plant conspicuousness and attractiveness to herbivores on the plant, it is useful to consider not only herbivore presence but also feeding damage. Feeding damage refers directly to the process of herbivory including feeding intensity, since herbivores that are present on plants do not necessarily feed on them. The extent of feeding damage may increase with plant size due to the greater apparency of large plants. Furthermore, high abilities to compensate for feeding damage of fast-growing plant species can contrast with high investments in defensive compounds of slow-growing plant species (growth–defence trade-off; Herms and Mattson 1992; Endara and Coley 2011). If this growth–defence trade-off manifests as a more effective defence against herbivores of small compared to large plant species, it may consequently result in overall higher herbivory levels of large compared to small plant species. Several studies have explored the effects of variation in intraspecific plant size on herbivory and found increased feeding damage with an increase in plant size when focusing on single plant components, such as flowers or leaves (Williams and Free 1979; Hainsworth et al. 1984; Tenow and Larsson 1987). However, so far, no studies have simultaneously analysed the effect of interspecific differences in plant size on feeding damage to different plant components.
Studying effects of plant size along an interspecific gradient has several advantages over intraspecific studies. Interspecific gradients can have a much broader range of plant size irrespective of environmental parameters such as nutrient availability, competition or the influence of other organisms in comparison to intraspecific plant size gradients (e.g. Buchanan and Underwood 2013). Interspecific experiments may also lead to more general conclusions than intraspecific studies as they offer the possibility to detect patterns across species. On the other hand, they have to cope with phylogenetic influences, which may be minimised by choosing a high number of closely related species. Using a large interspecific plant size gradient and a plant component-based approach, the effects of plant size on feeding damage can be analysed in great detail and will allow more comprehensive conclusions about potential effects of plant size on feeding damage to different components.
Not only plant size but also different component characteristics, such as size, biomass and number, may be of importance for herbivores specialised on a specific component (Espírito-Santo et al. 2007). Plants with many large components, particularly plants with high component biomass, should be highly attractive because of high food availability for herbivores. For this reason, these parameters should be considered in studies focusing on plant size effects.
Different plant characteristics can influence feeding damage, while feeding damage can oppositely influence the plants’ reproductive fitness. The effect of feeding damage on plant reproductive fitness may depend on the damaged plant component. While a negative impact of feeding damage by florivores on reproductive components and by seedeaters on seed number is not surprising (Moyes and Raybould 1997; Williams 2010), the effect of vegetative damage on reproductive fitness may be negative, neutral or even positive via (over)compensation or induced resistance to other herbivores (Strauss and Agrawal 1999; Puentes and Ågren 2012; McArt et al. 2013). Given a negative effect of feeding damage on plant reproductive fitness, plant size may consequently have an overall negative effect on plant reproductive fitness due to increased feeding damage with increasing plant size.
Up to now, complex interactions between feeding damage, plant size and reproductive fitness have never been investigated and may be particularly important for annual plants which do not have the opportunity for a delayed compensatory response across years. In this study, we analysed the effects of plant size on proportional feeding damage for different plant components along an interspecific plant size gradient. Most importantly, we analysed the effect of size-related feeding damage on plant reproductive fitness. Plant size was measured as plant height of 21 closely related annual brassicaceous plant species. We thereby determined proportional feeding damage to every aboveground plant component (i.e. to flowers, fruits, leaves and stems) and simultaneously involved component characteristics, such as size, biomass and number. This not only disentangles effects of plant size and component characteristics such as leaf number but also accounts for potential species-specific differences. Regarding other plant characteristics, such as flower structure or secondary plant substances, we chose species that were similar to each other to minimise plant size confounded differences among used species. As our study is conducted as a common garden experiment, plants were grown in a standardised way and effects of habitat and landscape are avoided. Thus, with our study, we are able to draw a comprehensive picture of the effects of plant size on feeding damage and plant fitness under standardised conditions, and come to more general conclusions using a broader plant size gradient than it would be possible using only a single plant species. The following hypotheses are tested:
-
1.
Proportional feeding damage to the different plant components increases with increasing plant size, as well as with increasing resource availability, i.e. component number and biomass.
-
2.
Proportional feeding damage to the different plant components, particularly to reproductive components (flowers and fruits), negatively affects plant reproductive fitness.
-
3.
Plant reproductive fitness decreases with increasing plant size.
Materials and methods
Study site and sampling design
The common garden experiment (Online Resource 1) was established on a grassland area dominated by grasses and herbs, including brassicaceous herbs, in Göttingen (Lower Saxony, Germany; 51.5°N, 9.9°E) in summer 2010. A total of 25 species from the family Brassicaceae that differed in size were chosen. Plant species that did not flower between mid-June and mid-July 2010 were excluded from the dataset to avoid phenological dissimilarity in the local insect community of the study area. The remaining 21 plant species covered a gradient in plant size from 12.65 cm ± 1.05 cm [Diplotaxis muralis (L.) DC.] to 120.50 cm ± 2.95 cm height (Raphanus sativus L. oleiformis) (Fig. 1f). Many plant characteristics were similar among the tested plant species, such as flower structure, the presence of glucosinolates as secondary plant substances and their pollination ecology in that insect pollination increases seed set (Klotz et al. 2002). The plant species we used include neophytes, cultivated plants and indigenous weeds and are all common German annuals, allowing us to directly assess their response to herbivory in contrast to perennial species, which may respond across years (e.g. Buchanan and Underwood 2013). Four plots per plant species, in total 100 plots (16 later excluded), were arranged in monoculture in a completely randomised design. Plots measured 1 m2 and were separated by 30 cm. Plants were managed in their plant density to cover approximately 100 % of the plot until they were fully blooming and were prevented from exceeding plot borders. Potential effects of plant height were thereby disentangled from the area covered by the plants, implying a negative relationship between plant size and plant density per area, which commonly occurs under natural conditions. Nevertheless, we accounted for the differences in plant density by involving the number and biomass per plot of components of interest, such as the number of leaves per plot in terms of feeding damage to leaves, in the analyses (described below). All plots were fertilised once equally (with NPK fertiliser 15:6:12) and regularly watered and weeded. We did not apply any pesticide during the course of the experiment.

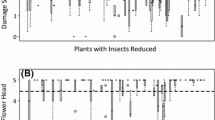
Effects of plant size, component biomass and number on feeding damage to a flowers, b fruits, c, d leaves and e stems. Axes of variables were transformed corresponding to analyses (feeding damage to the different components: asin-sqrt-transformation; biomass leaves per plot: sqrt-transformation; number leaves per plot: log-transformation). Predictions derive from the model with the lowest AICc including all explanatory variables with a parameter weight ≥0.6. f Plant sizes of the smallest, an intermediate and the largest species of the study are shown relative to one another (Diplotaxis muralis (L.) DC. 12.65 cm ± 1.05 cm, Sisymbrium officinale L. 69.80 cm ± 3.10 cm and Raphanus sativus L. oleiformis 120.50 cm ± 2.95 cm height). Single pictures are taken from Schlinkert (2014)
Plant appearance and chemistry traits
Plant size (height from the ground to the top of the plant) and the number of flowers, leaves and stems of first and second order were recorded per plant individual for five randomly selected plant individuals per plot at the time of full blossom. Number of fruits was counted at the time of full ripeness. Flower size was measured as petal length, leaf size in cm2 (we always measured the lowest living leaf as leaf size depends on its position) and stem diameter in mm (measured at ground level). Size of fruits was measured when fully ripe as length × width in mm. Mean plant size and mean size of the different plant components (flowers, fruits, leaves and stems) were calculated for each plot. The number of flowers, fruits, leaves and stems of five plant individuals were extrapolated to plot level (mean per plant individual × number of plants in the plot). Flower colour depended on the plant species (yellow or white). To measure component biomass, we harvested fruits, leaves and stems from plants in one quarter of each plot. To assess flower biomass, we harvested flowers from plants in one quarter of each of two randomly selected plots per plant species. Flowers were picked at the time of full blossom, stems and leaves were harvested at the time of early ripening, and fruits were harvested at the time of full ripeness of each plot. Harvested plant individuals were counted and components were oven-dried for 48 h at 60 °C to get dry biomass weights. Biomass of the different components per plot was extrapolated by multiplying the mean biomass per plant individual of the relevant plot with its plant individual number. Since only two randomly selected plots per plant species were sampled to assess flower biomass, we extrapolated flower biomass of the remaining two plots per plant species by multiplying their plant individual number with the averaged flower biomass per plant individual of the two sampled plots. Additionally, we assessed chemical traits of leaves, namely nitrogen, carbon and glucosinolate content (Online Resource 2).
Only leaves and stems for biomass assessment were taken from identical plant individuals, as the point in time of harvest was identical, while we excluded destructively treated plants from further examinations (i.e. leaves and stems, flowers and fruits were each harvested from different individuals). When harvesting the different plant components, we left five randomly selected and individually marked plant individuals per plot intended to develop pods for the measurement of fitness parameters.
Plant reproductive fitness
Plant reproductive fitness can be measured as the number of seeds a plant individual produces, referring to the number of its potential descendants. 1000 seed weight, i.e. the weight of a single seed × 1000, is often used as an indicator for the fitness of the produced seeds, while the product of seed number and single seed weight refers to the overall seed output, the yield in terms of crops. To assess plant reproductive fitness, a subset of 20 randomly selected fruits from the five randomly selected plant individuals per plot was opened. Seeds of ripe and closed fruits were counted and oven-dried for 48 h at 60 °C to measure their 1000 seed weight. The number of seeds per plant individual was estimated by multiplying the mean seed number per pod with the total number of pods of the plant individual. Total seed weight per individual was assessed by multiplying the seed number per individual with the 1000 seed weight divided by 1000 (the single seed weight) for the plant individual. Further, mean values per plot for seed number, 1000 seed weight and total seed weight per individual were calculated.
The natural capabilities of the different plant species regarding seed number, 1000 seed weight and total seed weight differ. These differences were accounted for by using relative rather than total values for the plant reproductive fitness parameters. Seed number (%), 1000 seed weight (%) and total seed weight per individual (%) refer to the percentage of the maximal capability for each plant species. The maximal capability of a species was determined as the mean of the ten maximal values observed in our study, using a total of 40 randomly chosen plant individuals per species. Thereby, these values refer to the natural capabilities of the specific breeding lines and varieties of the plants used in our experiment under the specific natural conditions of our experimental site. They originate from plant individuals with access by pollinating insects and below-average levels of feeding damage (63.65 ± 26.04, 64.24 ± 23.86 and 94.21 ± 34.37 % of the mean proportional feeding damage per plant species regarding seed number, total seed weight and 1000 seed weight).
Feeding damage to the different plant components
Feeding damage to plant components was recorded from five randomly selected plant individuals per plot. Feeding damage to flowers, fruits and leaves was recorded from the same individuals as plant reproductive fitness parameters, while feeding damage to stems was assessed from different individuals as we dissected the first and second order stems at the time of full blossom. To assess flower feeding damage, the numbers of podless stalks and developed fruits per plant individual were counted at the time of full ripeness. Podless stalks occur when buds and flowers are fed on (Williams 2010). Therefore, feeding damage to flowers was defined as percentage of podless stalks based on the number of potential fruits (developed fruits plus podless stalks). Additionally, a subset of 20 ripe fruits per sampled plant individual was opened and categorised as damaged by herbivores if we found loopholes, galleries, pitted seeds or an animal inside the fruit. Fruit feeding damage was defined as the percentage of damaged fruits. Leaf feeding damage (percentage of the damaged leaf surface) was estimated during the time of full blossom using a transparent grid template and involving all leaves from the sampled individuals. Feeding damage to stems was defined as percentage of the stem sections with mines or loopholes. Means of feeding damage for the different plant components were calculated for each plot.
Statistics
The effect of plant size on feeding damage (%) to flowers, fruits, leaves and stems was analysed using linear mixed effects models (nlme R package, Pinheiro et al. 2015; R v.2.12.2, R Development Core Team 2015). As covariates, the number and biomass of the relevant plant component (flowers, fruits, leaves or stems), flower colour regarding the analysis of feeding damage to flowers, and all two-way interactions were included. Size of the plant components was not part of the models due to high correlation with other explanatory variables (Online Resource 3: table ESM3a). Plant species was used as a random effect in models to avoid pseudoreplication (four plots per plant species).
The effect of feeding damage on plant fitness was likewise analysed using linear mixed effects models with plant species as a random effect. Seed number (%), 1000 seed weight (%) and total seed weight (%) were used as response variables and feeding damage to flowers, fruits, leaves and stems (%, the latter as binomial variable), including all two-way interactions, as explanatory variables. The net effect of plant size on plant fitness was analogously analysed, using linear mixed effects models and plant size as an explanatory variable.
Correlations between explanatory variables were tested for each model and we found significant correlations (p value <0.05) between several variables (Online Resource 3). Multicollinearity of explanatory variables was controlled and did not exceed the value of three for the variance inflation factor in any model (single exception: 3.17 for the model testing the effect of plant size, stem number and biomass on feeding damage to stems), allowing their parallel use in models (Zuur et al. 2010; HH-package, Heiberger 2015). Diagnostic plots were examined and variables transformed (log-, square-root- or arcsine-square-root-transformations were used) whenever necessary to avoid heteroscedasticity or non-normal distribution of errors.
Models were simplified by calculating AICc values for all full model subsets using the dredge function in the MuMIn package (Barton 2015). With respect to the non-independence between some explanatory variables (Online Resource 3), parameter weights were used for the identification of explanatory variables and interactions that consistently contributed to the models’ explanatory power. Parameter weights were computed by averaging models with ΔAICc <2 (Burnham and Anderson 2002; Grueber et al. 2011). We defined parameters with a parameter weight ≥0.6 as important for the explication of the response variable’s variance. Summary output of the model with the lowest AICc including all explanatory variables with a parameter weight ≥0.6 led to given estimates with standard errors (Tables 1, 2) that are centred and standardised to improve their interpretability (Schielzeth 2010).
Results
Effects of plant size and component characteristics on feeding damage to the different plant components
We recorded overall feeding damage to flowers of 29.82 ± 2.83 % (arithmetic mean and standard error), feeding damage to fruits of 5.10 ± 0.91 %, feeding damage to leaves of 1.13 ± 0.17 % and feeding damage to stems of 0.88 ± 0.22 % (see Online Resource 4 for all). Based on our observations, feeding damage to flowers was attributed to pollen beetles and their larvae (Meligethes aeneus Fabricius) and feeding damage to fruits mainly to weevils (cabbage seedpod weevil Ceutorhynchus obstrictus Marsham and C. floralis Paykull) and to a lesser extend to the brassica pod midge (Dasineura brassicae Winnertz). Feeding damage by leaf chewers was composed of a mainly point-wise feeding pattern, but we also observed fenestration feeding and leaf mining patterns. Flea beetles (Phyllotreta nemorum L. and P. nigripes (Fabricius) caused the most feeding damage to leaves, while damage to stems was done by weevils (C. pallidactylus Marsham and C. napi Gyllenhaal). In general, most observed herbivorous species were specialised on the family of Brassicaceae, but had no strong specialisation on any single plant species of the experiment.
Plants along the plant size gradient were similar in characteristics such as their defensive compounds or nitrogen content of leaves (Online Resource 2). Some characteristics of single plant components were not independent from plant size and were consequently involved in the analyses as covariables whenever possible to disentangle their effects from plant size effects. Thereby, we found that proportional flower feeding damage was positively influenced by plant size, while number, biomass and colour of flowers had no effect (Table 1; Fig. 1a). Proportional fruit feeding damage was also (even though weakly) positively affected by plant size, while the number and biomass of fruits had no influence (Table 1; Fig. 1b). In contrast, proportional leaf and stem feeding damage were positively affected by component biomass, but plant size had no influence (Table 1; Fig. 1c, e). Only feeding damage to leaves was affected by component number, as feeding damage tended to decrease with increasing leaf number (Table 1; Fig. 1d).
Effects of feeding damage to the different plant components on plant reproductive fitness
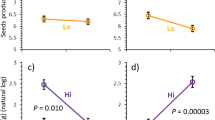
Plant reproductive fitness parameters refer to the degree (%) to which a plant individual met the maximal capability of its plant species. We recorded mean seed number as 57.43 ± 2.82 %, mean 1000 seed weight as 62.14 ± 1.81 % and mean total seed weight as 51.85 ± 2.91 % (see Online Resource 4 for all). Seed number per individual (%), 1000 seed weight (%) and total seed weight per individual (%) decreased with increasing flower feeding damage (Table 2; Fig. 2a, b, d). Interestingly, we found interaction effects between flower and stem feeding damage: the negative effect of flower feeding damage on seed number (%) and on total seed weight (%) was stronger on plots with stem feeding damage than on plots without stem feeding damage (Table 2; Fig. 2a, d). 1000 seed weight (%) was not influenced by stem feeding damage (Table 2). Leaf feeding damage had no influence on seed number (%) and total seed weight (%) but a slight positive influence on 1000 seed weight (%) (Table 2; Fig. 2c). Fruit feeding damage had no effect on any plant reproductive fitness parameter (Table 2).

Effects of feeding damage to the different plant components on a seed number (% of the species’ maximum), b, c 1000 seed weight (%) and d total seed weight (%). Axes of variables were transformed corresponding to analyses (feeding damage to the different components, seed number (%) and total seed weight (%): asin-sqrt-transformation). Predictions derive from the model with the lowest AICc including all explanatory variables with a parameter weight ≥0.6. a, d Solid line plants with stem damage, dashed line plants without stem damage
Net effect of plant size on plant reproductive fitness
To assess the overall fitness consequences of being large or small, we analysed the effect of plant size on plant reproductive fitness, namely on seed number per individual (%), 1000 seed weight (%) and total seed weight per individual (%). We thereby found a negative net effect of plant size on 1000 seed weight (%) (parameter weight = 1; estimate = 0.127, standard error = 0.043), but not on seed number (%) (parameter weight = 0) or total seed weight (%) (parameter weight = 0).
Discussion
Proportional feeding damage to the different plant components was hypothesised to increase with increasing plant size. However, we found different effects for the specific plant components. Proportional feeding damage to flowers and fruits increased with increasing plant size and was not influenced by resource availability, i.e. component number and biomass. In contrast, feeding damage to leaves and stems only increased with increasing biomass of leaves and stems but not with plant size. The influence of feeding damage on plant reproductive fitness depended on the type of feeding damage. Flower feeding damage had the strongest effect on reproductive fitness as it reduced seed number (%), 1000 seed weight (%) and total seed weight (%). Feeding damage to leaves and stems played a minor role and feeding damage to fruits had no influence on any fitness parameter. Regarding the overall effects of plant size on reproductive fitness, we found a negative net effect of plant size only on 1000 seed weight (%), while seed number (%) and total seed weight (%) remained unaffected. These findings generally support our hypotheses but also indicate specific effects of plant size on the proportional feeding damage of individual plant components and also specific consequences for plant reproductive fitness.
Effects of plant size and component characteristics on feeding damage to the different plant components
Large plants are highly apparent and attractive to associated animals due to their wide range of resources and niches, and are thus expected to be easily found and colonised by numerous herbivores (Feeny 1976; Lawton 1983). Resulting from these direct effects of plant size and potential effects of other size-related plant traits, a positive effect of plant size on herbivore species richness has been shown several times (Lawton and Price 1979; Neuvonen and Niemelä 1981; Haysom and Coulson 1998), but studies testing its effect on feeding damage are rare and less general due to their focus on a single type of feeding damage and single plant species (e.g. Tenow and Larsson 1987; Alonso and Herrera 1996; Castagneyrol et al. 2013). Feeding damage to both flowers and fruits was strongly affected by plant size across the 21 tested plant species, which has previously only been shown for an intraspecific plant size gradient in species such as oilseed rape (Brassica napus L.) (Williams and Free 1979) and scarlet gilia (Ipomopsis aggregata (Pursh) V. E. Grant) (Hainsworth et al. 1984).
In contrast, feeding damage to both leaves and stems was correlated to respective component biomass, but not to plant size. Component biomass is composed of the number, size and mass density and is a measurement of the quantity of resources that can be expected to attract herbivores (Lawton 1983; Araujo et al. 2006). Studies that tested for an effect of total plant biomass on leaf and stem herbivores such as galling insects, which are specialised on a specific plant resource like young tissue of meristems, failed to detect any relationship (Espírito-Santo et al. 2007). However, biomass of the respective component instead of total plant biomass showed a positive effect on these herbivores (Araujo et al. 2006). This emphasises the importance of plant component biomass for feeding damage to a particular plant component. As biomass of both leaves and stems was positively correlated with plant size, we should keep in mind that we cannot completely disentangle these effects (Lawton 1983), even though these correlations were not too strong for a combined analysis.
Nevertheless, we can state that plant size was a more important driver for feeding damage to flowers and fruits than for feeding damage to leaves and stems. This is not surprising, as large, highly attractive leaves and stems, the type most often damaged, are often situated on the lower part of the plant (personal observation; Collinge and Louda 1988; Dechert and Ulber 2004). Therefore, under natural conditions, they are often hidden from herbivores by neighbouring vegetation, independent of plant size. In contrast, flowers and fruits of the tested species are situated at the exposed top of the plant, where plant size is synonymous with inflorescence height. Flowers and fruits of large plants under natural conditions often overtop those of smaller plants if their flowers and fruits are situated at the top of the plant, making smaller plants more hidden, resulting in greater feeding damage for larger plants. The role of plant size regarding the search strategy of herbivores for food plants may therefore depend on the component position, on whether or not its visibility depends on plant size. The difference in search strategies, manifested under natural conditions, probably led to a high attractiveness of plots with large plants for herbivores searching for flowers and fruits and a disinterest in plant size of herbivores searching for big leaves and stems.
It is interesting that number and biomass of flowers and fruits per plot did not influence feeding damage to these components, while results of increasing feeding damage to leaves and stems with increasing component biomass per plot supported the expected positive relationship to resource availability. Adult pollen beetles and their larvae feed mainly on pollen and pod miners feed on seeds, while flea beetles feed on the green tissue of the leaf blade and weevils feed on the inner parts of the stems and may even hollow them out completely (Williams 2010; Juran et al. 2011). Thus, a minor part of flower and fruit biomass is edible, while the biomass of leaves and stems better reflects the amount of edible components. Component number had no effect on the feeding damage to flowers, fruits and stems. The mere number of entities within components may be of little value for many herbivores if component biomass is simultaneously considered, which comprises component size and mass density, besides number. Size and mass density of components may also be of importance regarding shelter from enemies, as for example stem mining larvae take refuge from parasitoids in stems with a large diameter and high amount of biomass, being beyond the reach of parasitoid ovipositors (Ulber 2003). We detected an influence of the entity number of components only for feeding damage to leaves. As leaf feeding damage was attributed mainly to mobile herbivores like flea beetles (personal observations), leaf number may represent resource availability, as suggested in previous studies by increasing clutch size of different butterflies with increasing number of food plant leaves (Vasconcellos-Neto and Ferreira Monteiro 1993; Reudler Talsma et al. 2008). Surprisingly, the feeding damage to leaves in our study was not positively but negatively affected by leaf number. Leaf herbivory can affect plants early in the season as leaves of seedlings and young plants are already attacked by cabbage flea beetles (personal observations; Alford et al. 2003). Plant species may cope with early herbivory by investment in defence mechanisms (resistance) or in regrowth (tolerance), while plants with a high regrowth capacity are known for overcompensation (Strauss and Agrawal 1999). The negative relationship between feeding damage to leaves and leaf number was possibly caused by overcompensatory creation of leaves (i.e. assimilation capacity) as a response to leaf damage (glucosinolate composition of leaves was not correlated with leaf damage; Online Resource 2: table ESM2).
Feeding damage to flowers was not significantly influenced by flower colour, although pollen beetles, the major florivores in our study, are known to favour yellow flowers due to visual or physiological colour-related aspects of flowers such as production of volatiles (Giamoustaris and Mithen 1996). Abundance of larvae and adult pollen beetles was observed to be higher on yellow flowers than on white flowers but did not translate to increased flower feeding damage. However, adult pollen beetles have been shown to avoid buds for feeding that have the preferred size for oviposition (Ekbom and Borg 1996) and may avoid feeding on buds already occupied by their larvae. Since their home range for feeding is wider than for oviposition (Ekbom and Borg 1996), adults may have laid eggs mainly in yellow buds and may have switched regularly to white flowers for feeding, leading to similar damage levels to yellow and white flowers.
Effects of feeding damage to the different plant components on plant reproductive fitness
Our finding that feeding damage to flowers negatively affected plant reproductive fitness agrees with results from other studies (McCall and Irwin 2006 and studies cited therein). Feeding on pollen and flower components by pollen beetles and their larvae is known to lead to podless stalks and weakened pods (Free and Williams 1979; Williams 2010). Consequently, feeding damage to flowers often leads to a reduction of the total seed number (Steffan-Dewenter and Tscharntke 1999; Williams 2010). Florivory may additionally have an indirect negative effect on seed number in reducing the attractiveness of flowers to pollinators and the availability of pollen, followed by decreased effectiveness of pollination by insects and wind (McCall and Irwin 2006). Plants with a high amount of damaged flowers, for example due to pollen beetles, may compensate by producing new flowers but at the expense of the remaining flowers’ 1000 seed weight (Trumble et al. 1993). The negative effects of feeding damage to flowers on seed number (%) and total seed weight (%) were reinforced by feeding damage to stems. Tunnelling of stem mining flea beetle larvae may cause distortion of tissue and consequent loss of plant vigour (Juran et al. 2011). Additionally, holes caused by the female’s ovipositor and by larvae emerging from stems often provide gateways for fungal infestations (Juran et al. 2011). Both plant vigour loss and fungal infestations as a consequence of stem mining may weaken a plant and reduce its ability to compensate for flower damage.
A slight stimulating effect on the 1000 seed weight (%) could be ascribed to feeding damage to leaves. Overcompensation has been described for many brassicaceous species and genera used in our study, such as Raphanus raphanistrum L., B. napus, Sinapis alba L. and several more (Gavloski and Lamb 2000; Agrawal 2001, and others) in that feeding damage to leaves can increase plant reproductive fitness, including via increased seed weight (e.g. Agrawal 2001). Another explanation might be that the 1000 seed weight (%) could have been positively influenced by high leaf biomass rather than by high feeding damage to leaves, as feeding damage to leaves was positively related to leaf biomass (Table 1). High leaf biomass implies high photosynthetic potential and a larger supply of carbon for reproduction. Plant reproductive effort (the ratio of reproductive biomass to total biomass) is in general higher in monocarpic than polycarpic species since the former allocate their energy mainly to reproduction and the latter mainly to competition and predator avoidance (Kawano and Nagai 1975). Hence, high leaf biomass may lead to high seed weight particularly in monocarpic plants, which were used in our study.
Feeding damage to fruits has a negative effect on the seed number of several species like I. aggregata (Hainsworth et al. 1984), Oenothera biennis L. (McArt et al. 2013) and others. Cabbage seedpod weevils often cause severe seed losses through feeding on seeds of Brassicaceae, particularly as forerunners of the brassica pod midge or secondary infestations by fungal pathogens (Moyes and Raybould 1997; Alford et al. 2003; Williams 2010). In contrast, many studies analysing brassicaceous species have found little effect of pre-dispersal seed predation on seed number (Williams and Free 1979; Free et al. 1983; Duggan 1985; but see Williams and Free 1978), consistent with results from our study. We observed cabbage seed weevil larvae causing the majority of feeding damage to fruits but infrequent secondary infestations of pods by brassica pod midges. We further observed relatively high parasitism rates of cabbage seedpod weevil larvae (on average 78.03 ± 4.44 %) by chalcid wasps, which may reduce the consumption by host larvae (Moyes and Raybould 1997). Due to infrequent secondary infestations by brassica pod midges and potentially low consumption rates by parasitised weevil larvae, the expected negative effect of feeding damage to fruits on plant reproductive fitness may have been weak and consequently compensated for by the plants (Williams and Free 1979).
Net effect of plant size on plant reproductive fitness
We hypothesised a negative net effect of plant size on plant reproductive fitness as a consequence of increased feeding damage (hypothesis 1) and consequently reduced plant fitness (hypothesis 2). Although size-related feeding damage to flowers resulted in reduced seed number (% of the plant species’ maximum), 1000 seed weight (%) and total seed weight (%), we could only demonstrate a negative net effect of plant size on 1000 seed weight (%). The other two fitness parameters, seed number (%) and total seed weight (%), which was probably mainly influenced by seed number, remained unaffected by plant size. Bearing in mind that unconsidered size-related plant traits also potentially contributed to these patterns, we propose that fitness costs for large plants in terms of seed loss due to herbivores may be counterbalanced by benefits of large plants, including greater abilities for compensation (Strauss and Agrawal 1999; Williams and Free 1979), higher competitiveness for light and other resources (Weiner 1985) or high conspicuousness to pollinators (Donnelly et al. 1998).
Conclusions
Detailed analyses of feeding damage to different plant components in relation to plant size and the linkage of size-related feeding damage to fitness consequences led to comprehensive conclusions. We showed a component-specific effect of plant size on feeding damage. Herbivore damage to components at the top of the plant, i.e. to flowers and fruits, was sensitive to plant size. Contrastingly, component biomass played the most important role, independently of plant size, for herbivores of the more hidden leaves and stems. We suggest that the search strategy of herbivores for food plants depends on the component of interest, particularly on its position on the plant.
The effect of feeding damage to the different plant components on plant reproductive fitness was likewise not uniform. Feeding damage to flowers played by far the most important role in reducing plant reproductive fitness and negatively affected seed number, 1000 seed weight and total seed weight. Due to the increase in feeding damage to flowers with increasing plant size and its strong negative effect on plant fitness, we could demonstrate a negative net effect of plant size on 1000 seed weight. Thus, being large and thereby highly conspicuous and attractive to herbivores caused a disadvantage regarding 1000 seed weight. Regarding seed number and total seed weight, we propose that fitness costs for large plants caused by herbivore damage were counterbalanced by benefits of large plants, which can assumed to be high competitiveness for light and other resources (Weiner 1985), high conspicuousness to pollinators (Donnelly et al. 1998) or a greater ability to compensate for feeding damage, indicating a growth–defence trade-off through the production of more numerous seeds. These conclusions are of great relevance as our findings are drawn from patterns across 21 plant species within a highly standardised experiment.
Author contribution statement
TT, CW and HS conceived and designed the experiments; HS, ML and PK performed the experiments. HS and YC performed statistical analyses. HS and TT wrote the manuscript; all authors provided editorial advice.
References
Agrawal AA (2001) Transgenerational consequences of plant responses to herbivory: an adaptive maternal effect? Am Nat 157:555–569. doi:10.1086/319932
Alford DV, Nilsson C, Ulber B (2003) Insect pests of oilseed rape crops. In: Alford DV (ed) Biocontrol oilseed rape pests. Blackwell, Oxford, pp 9–31
Alonso C, Herrera CM (1996) Variation in herbivory within and among plants of Daphne laureola (Thymelaeaceae): correlation with plant size and architecture. J Ecol 84:495–502
Araujo APA, de D’arc Paula J, Carneiro MAA, Schoereder JH (2006) Effects of host plant architecture on colonization by galling insects. Austral Ecol 31:343–348. doi:10.1111/j.1442-9993.2006.01563.x
Barton K (2015) MuMIn: multi-model inference. R package version 1.13.4. http://CRAN.R-project.org/package=MuMIn
Blanckenhorn WU (2000) The evolution of body size: what keeps organisms small? Q Rev Biol 75:385–407. doi:10.1086/393620
Brown JH, Maurer BA (1986) Body size, ecological dominance and Cope’s rule. Nature 324:248–250
Buchanan AL, Underwood N (2013) Attracting pollinators and avoiding herbivores: insects influence plant traits within and across years. Oecologia 173:473–482. doi:10.1007/s00442-013-2629-4
Burnham KP, Anderson DR (2002) Model selection and multi-model inference: a practical information-theoretic approach, 2nd edn. Springer, New York
Castagneyrol B, Giffard B, Péré C, Jactel H (2013) Plant apparency, an overlooked driver of associational resistance to insect herbivory. J Ecol 101:418–429. doi:10.1111/1365-2745.12055
Collinge SK, Louda SM (1988) Patterns of resource use by a drosophilid (Diptera) leaf miner on a native crucifer. Ann Entomol Soc Am 81:733–741
Dechert G, Ulber B (2004) Interactions between the stem-mining weevils Ceutorhynchus napi Gyll. and Ceutorhynchus pallidactylus (Marsh.) (Coleoptera: Curculionidae) in oilseed rape. Agric For Entomol 6:193–198. doi:10.1111/j.1461-9555.2004.00220.x
Donnelly SE, Lortie CJ, Aarssen LW (1998) Pollination in Verbascum thapsus (Scrophulariaceae): the advantage of being tall. Am J Bot 85:1618–1625
Duggan AE (1985) Pre-dispersal seed predation by Anthocharis cardamines (Pieridae) in the population dynamics of the perennial Cardamine pratensis (Brassicaceae). Oikos 44:99–106
Ekbom B, Borg A (1996) Pollen beetle (Meligethes aeneus) oviposition and feeding preference on different host plant species. Entomol Exp Appl 78:291–299. doi:10.1111/j.1570-7458.1996.tb00793.x
Endara M-J, Coley PD (2011) The resource availability hypothesis revisited: a meta-analysis. Funct Ecol 25:389–398. doi:10.1111/j.1365-2435.2010.01803.x
Espírito-Santo MM, Neves FDS, Andrade-Neto FR, Fernandes GW (2007) Plant architecture and meristem dynamics as the mechanisms determining the diversity of gall-inducing insects. Oecologia 153:353–364. doi:10.1007/s00442-007-0737-8
Feeny P (1976) Plant apparency and chemical defense. In: Wallace JW, Mansell RL (eds) Biochem Interact between plants insects. Springer, New York, pp 1–40
Free JB, Williams IH (1979) The distribution of insect pests on crops of oil-seed rape (Brassica napus L.) and the damage they cause. J Agric Sci 92:139–149
Free JB, Ferguson AW, Winfield S (1983) Effect of various levels of infestation by the seed weevil (Ceutorhynchus assimilis Payk.) on the seed yield of oil-seed rape (Brassica napus L.). J Agric Sci 101:589–596
Gavloski J, Lamb R (2000) Compensation for herbivory in cruciferous plants: specific responses to three defoliating insects. Environ Entomol 29:1258–1267. doi:10.1603/0046-225X-29.6.1258
Giamoustaris A, Mithen R (1996) The effect of flower colour and glucosinolates on the interaction between oilseed rape and pollen beetles. Entomol Exp Appl 80:206–208. doi:10.1111/j.1570-7458.1996.tb00919.x
Grueber CE, Nakagawa S, Laws RJ, Jamieson IG (2011) Multimodel inference in ecology and evolution: challenges and solutions. J Evol Biol 24:699–711. doi:10.1111/j.1420-9101.2010.02210.x
Hainsworth FR, Wolf LL, Mercier T (1984) Pollination and pre-dispersal seed predation: net effects on reproduction and inflorescence characteristics in Ipomopsis aggregata. Oecologia 63:405–409
Haysom KA, Coulson JC (1998) The Lepidoptera fauna associated with Calluna vulgaris: effects of plant architecture on abundance and diversity. Ecol Entomol 23:377–385. doi:10.1046/j.1365-2311.1998.00152.x
Heiberger RM (2015) HH: statistical analysis and data display: Heiberger and Holland. R package version 3.1-15. http://CRAN.R-project.org/package=HH
Hemptinne J-L, Magro A, Evans EW, Dixon AFG (2012) Body size and the rate of spread of invasive ladybird beetles in North America. Biol Invasions 14:595–605. doi:10.1007/s10530-011-0101-0
Herms D, Mattson W (1992) The dilemma of plants: to grow or defend. Q Rev Biol 67:283–335. doi:10.1086/417659
Juran I, Gothlin Čuljak T, Grubišić D (2011) Rape stem weevil (Ceutorhynchus napi Gyll. 1837) and cabbage stem weevil (Ceutorhynchus pallidactylus Marsh. 1802) (Coleoptera: Curculionidae)—important oilseed rape pests. Agric Conspec Sci 76:93–100
Kawano S, Nagai Y (1975) The productive and reproductive biology of flowering plants, 1: life history strategies of three Allium species in Japan. Bot Mag 88:281–318
Klotz S, Kühn I, Durka W (2002) BIOLFLOR—Eine Datenbank zu biologisch-ökologischen Merkmalen der Gefäßpflanzen in Deutschland (BIOLFLOR database—search and information system on vascular plants in Germany). Schriftenr für Veg 38
Lawton JH (1983) Plant architecture and the diversity of phytophagous insects. Annu Rev Entomol 28:23–39
Lawton JH, Price PW (1979) Species richness of parasites on hosts: agromyzid flies on the British Umbelliferae. J Anim Ecol 48:619–637
McArt SH, Halitschke R, Salminen JP, Thaler JS (2013) Leaf herbivory increases plant fitness via induced resistance to seed predators. Ecology 94:966–975. doi:10.1890/12-1664.1
McCall AC, Irwin RE (2006) Florivory: the intersection of pollination and herbivory. Ecol Lett 9:1351–1365. doi:10.1111/j.1461-0248.2006.00975.x
Moyes CL, Raybould AF (1997) Herbivory by the cabbage seed weevil (Ceutorhynchus assimilis) in natural populations of Brassica oleracea. Int Symp Brassica 97 Xth Crucif Genet Work 459:315–322
Neuvonen S, Niemelä P (1981) Species richness of Macrolepidoptera on Finnish deciduous trees and shrubs. Oecologia 51:364–370
Peters RH (1983) The ecological implications of body size. doi: 10.1017/CBO9780511608551
Pinheiro J, Bates D, DebRoy S et al (2015) nlme: linear and nonlinear mixed effects models. R package version 3.1-120. http://CRAN.R-project.org/package=nlme%3E
Puentes A, Ågren J (2012) Additive and non-additive effects of simulated leaf and inflorescence damage on survival, growth and reproduction of the perennial herb Arabidopsis lyrata. Oecologia 169:1033–1042. doi:10.1007/s00442-012-2276-1
R Development Core Team (2015) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL http://www.R-project.org/
Remmel T, Tammaru T (2009) Size-dependent predation risk in tree-feeding insects with different colouration strategies: a field experiment. J Anim Ecol 78:973–980. doi:10.1111/j.1365-2656.2009.01566.x
Reudler Talsma JH, Biere A, Harvey JA, van Nouhuys S (2008) Oviposition cues for a specialist butterfly-plant chemistry and size. J Chem Ecol 34:1202–1212. doi:10.1007/s10886-008-9519-y
Schielzeth H (2010) Simple means to improve the interpretability of regression coefficients. Methods Ecol Evol 1:103–113. doi:10.1111/j.2041-210X.2010.00012.x
Schlinkert H (2014) Plant size gradient in Brassicaceae. doi: 10.6084/m9.figshare.1246843
Steffan-Dewenter I, Tscharntke T (1999) Effects of habitat isolation on pollinator communities and seed set. Oecologia 121:432–440. doi:10.1007/s004420050949
Strauss SY, Agrawal AA (1999) The ecology and evolution of plant tolerance to herbivory. Trends Ecol Evol 14:179–185. doi:10.1016/S0169-5347(98)01576-6
Tenow O, Larsson S (1987) Consumption by needle-eating insects on Scots pine in relation to season and stand age. Ecography (Cop) 10:249–260. doi:10.1111/j.1600-0587.1987.tb00766.x
Trumble JT, Kolodny-Hirsch DM, Ting IP (1993) Plant compensation for arthropod herbivory. Annu Rev Entomol 38:93–119
Ulber B (2003) Parasitoids of Ceutorhynchid stem weevils. In: Alford DV (ed) Biocontrol oilseed rape pests. Blackwell, Oxford, pp 87–96
Vasconcellos-Neto J, Ferreira Monteiro R (1993) Inspection and evaluation of host plant by the butterfly Mechanitis lysimnia (Nymph., Ithomiinae) before laying eggs: a mechanism to reduce intraspecific competition. Oecologia 95:431–438. doi:10.1007/BF00320999
Weiner J (1985) Size hierarchies in experimental populations of annual plants. Ecology 66:743–752. doi:10.2307/1940535
Williams IH (2010) The major insect pests of oilseed rape in Europe and their management: an overview. In: Williams IH (ed) Biocontrol-based Integr Manag oilseed rape pests. Springer, Dordrecht, pp 1–44
Williams IH, Free JB (1978) The feeding and mating behaviour of pollen beetles (Meligethes aeneus Fab.) and seed weevils (Ceutorhynchus assimilis Payk.) on oil-seed rape (Brassica napus L.). J Agric Sci 91:453–459. doi:10.1017/S0021859600046554
Williams IH, Free JB (1979) Compensation of oil-seed rape (Brassica napus L.) plants after damage to their buds and pods. J Agric Sci 92:53–59. doi:10.1017/S0021859600060494
Zuur AF, Ieno EN, Elphick CS (2010) A protocol for data exploration to avoid common statistical problems. Methods Ecol Evol 1:3–14. doi:10.1111/j.2041-210X.2009.00001.x
Acknowledgments
Special thanks for help in the field go to Susanne Jahn and Brigitte Jünemann, as well as to Christiane Schlinkert, Eugen Schlinkert, Barbara Scheid, Christina Fischer, Verena Rösch, Georg Everwand, Jenny Osterburg, Annabelle Rohlfing, Anna-Katharina Franke, Petra Kubisch, Alexandra Arndt, Carina Burmeister and Felix Steinmeyer. KWS SAAT and the botanical gardens of the universities of Bayreuth and Göttingen provided part of the seeds. We thank Carsten Thies for advice and Kristy Udy for linguistic improvements of the manuscript and two anonymous reviewers for their comments and suggestions, which improved the manuscript. H.S. was supported by the Ministry of Science and Culture of Lower Saxony.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Andreas Prinzing.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Schlinkert, H., Westphal, C., Clough, Y. et al. Feeding damage to plants increases with plant size across 21 Brassicaceae species. Oecologia 179, 455–466 (2015). https://doi.org/10.1007/s00442-015-3353-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-015-3353-z