
Abstract
Species composition and productivity are influenced by water and N availability in semi-arid grasslands. To assess the effects of increased N deposition and water supply on plant species composition and productivity, two field experiments with four N addition treatments, and three N and water combination treatments were conducted in alpine grassland in the mid Tianshan mountains, northwest China. When considering N addition alone, aboveground biomass (AGB) of forbs (FAGB) responded less to N addition than AGB of grasses (GAGB). GAGB increased as an effect of N combined with water addition but FAGB did not show such an effect, reflecting a stronger response of grasses to the interaction of water availability and N than forbs. Under all treatments, N allocation to the aboveground tissue did not change for either forbs or grasses. N deposition and water addition did not alter species richness in the present study. These results suggest that N addition generally promoted AGB but had little effect on species richness in wet years. Snowfall in winter combined with rainfall in the early growing season likely plays a critical role in regulating plant growth of the subsequent year in the alpine grassland.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Anthropogenic activity is altering the global climate and atmosphere, including precipitation patterns and N deposition (IPCC 2007; Galloway et al. 2008). Grasslands, covering approximately 25 % of the land surface of the Earth, are limited usually by the amount of precipitation and available N (Huxman et al. 2004; Brookshire et al. 2012). Climate change (in terms of precipitation and temperature) is the predominant driver at the large scale, whereas N deposition drives grassland composition at the local scale (Gaudnik et al. 2011). Understanding how grassland plant communities respond to concurrent alterations in N and water availability is critical for projections of vegetation dynamics under future global change (Yang et al. 2011).
N deposition can stimulate plant growth and enhance the soil N pool, and long-term continuous N inputs may influence plant community composition in temperate grassland (Clark and Tilman 2008; Song et al. 2011). In general, increased N availability due to elevated N deposition will enhance aboveground biomass (AGB) and reduce species richness (DeFalco et al. 2003; Li et al. 2011). Relationships between N addition rate and species loss responses have been found in different grasslands. For example, long-term low chronic N deposition (10 kg N ha−1 year−1) can lead to significant species loss in a semi-arid temperate grassland in the US (Clark and Tilman 2008). Grass species richness decreased linearly with increasing N deposition of 5–35 kg N ha−1 year−1 in acidic grasslands of the UK (Stevens et al. 2004). The threshold for N-induced species loss to mature Eurasian grasslands was reported to be < 17.5 kg N ha−1 year−1 (Bai et al. 2010) or 30 kg N ha−1 year−1 (Song et al. 2011). However, the response of plant growth and species richness to N addition in alpine grasslands is still unclear.
Although N impacts many ecosystem processes, the extent of its influence is ultimately determined by the availability of water. Precipitation has been identified as a primary factor limiting vegetation growth in semi-arid grassland (Reichmann et al. 2013). It is considered to be a primary factor causing fluctuations in the plant community biomass in an Inner Mongolia grassland (Bai et al. 2004). Rain-event size (small/large) during the growing season affected the AGB in a semi-arid grassland of North America (Cherwin and Knapp 2012). Changes in rainfall and snow regimes may be of great consequence for C and N cycling because precipitation has a major effect on AGB and N cycling is strongly linked to drying-wetting cycles in an alpine grassland (Li et al. 2012a). Fisk et al. (1998) showed that water availability resulting from snowpack was relatively high, and it influenced plant growth and N cycling as snowpack accumulation dominated the growing-season length, water availability and plant distribution in an alpine meadow. Changes in the amount of winter precipitation may be just as important as summer precipitation for grassland processes because snowmelt may percolate to depth, affecting spring-time plant growth and summer soil water contents (Chimner and Welker 2005; Duparc et al. 2013).
Interaction between water and N is an increasingly more important issue in grassland ecosystems. Water availability affects N use efficiency in arid and semi-arid regions more than in wet regions (Akmal et al. 2010). Changes in AGB and species richness occurred at an N addition rate of 105 kg N ha−1 in a wet year and no effect was found in the dry years in the mature community of a semi-arid grassland of Inner Mongolia (Bai et al. 2010). The AGB and species richness, however, were not affected at an N addition rate of 100 kg N ha−1 in a desert grassland in spite of variability in seasonal precipitation (Ladwig et al. 2012). Species richness was reduced under N addition, whereas it did not vary with the interactive effect of precipitation and N addition in a temperate steppe (Yang et al. 2011).
These results show that the interactive effect of N and precipitation on the AGB and species richness was not consistent in grassland ecosystems. Furthermore, N addition and increased precipitation experiments have been conducted in both arid and semi-arid grasslands, whereas little is known about the interactions of precipitation and N deposition on the AGB and species richness in alpine grasslands.
Bayinbuluk, the alpine grassland site of the central Tianshan mountains in northwest China, is far from urban areas. This grassland receives a relatively low ambient N deposition—only about 8 kg N ha−1 year−1 (with 50 % dry and 50 % wet deposition, according to our monitoring data)—slightly below normal critical load (10–20 kg N ha−1 year−1) of N deposition for grassland worldwide (Bobbink et al. 2010). Together with rapid economic growth, N deposition has increased significantly at an average rate of increase of about 0.4 kg N ha−1 year−1 from 1980 to 2010 in China (Liu et al. 2013). N-fertilizer application has also increased steadily since the 1980s due to the expansion of cotton and cereal/fruit/vegetable production on some southern and northern Tianshan mountains in northwest China. Therefore, we expect further increased atmospheric N deposition in our study area. The elevated N deposition may threaten the function of the alpine grassland ecosystem and affect greenhouse gas emissions (Li et al. 2012b). In addition, anthropogenic forcing may contribute significantly to observed increases in precipitation in the mid-latitudes of the Northern Hemisphere, including the Tianshan mountains (Zhang et al. 2007). For the 2050s, the projections of the general circulation models show that this region will have wetter winters and that extreme precipitation events will be more intense in the spring than in the summer (Basher et al. 2010). Seasonality change and the type of precipitation may be more important for modulation of the N addition effect than the overall amount of precipitation for the alpine grassland. We are especially interested in the effect of the increase in both water supply (due to winter snow or early spring precipitation) and N deposition on AGB and plant species abundance in alpine grasslands. We therefore conducted two 4-year field experiments with N addition and increased N plus water supply in order to better understand how the alpine grassland responds to the integrative effects of increasing N and precipitation.
Materials and methods
Study area
The study was conducted at the Bayinbuluk Grassland Ecosystem Research Station, Chinese Academy of Sciences (42°53.1′N, 83°42.5′E). Bayinbuluk alpine grassland is located in the southern Tianshan mountains, Xinjiang Uygur Autonomous Region, China, and covers a total area of approximately 23,000 km2. The grassland is in the Tianshan mountains basin at a mean altitude of 2,500 m a.s.l. Calculated from local meteorological data (1981–2012), the mean annual precipitation (MAP) is 282.3 mm and the mean precipitation between late October and mid July of the subsequent year is 171.8 mm. The mean annual temperature is −4.8 °C.
Experimental design
There were four N addition treatments in experiment I: 0 kg N ha−1 year−1 (N0), 10 kg N ha−1 year−1 (N10), 30 kg N ha−1 year−1 (N30) and 90 kg N ha−1 year−1 (N90). Three combined N and water addition treatments were employed in experiment II: no N and no water addition (N0W0), no N and 7.5-mm water supply, accounting for roughly 1/3 of the total amount of snow from January to April (N0W1), and 30 kg N ha−1 year−1 and 7.5-mm water supply (N30W1). Each treatment included four blocks (each 4 × 8 m with a 1-m—wide buffer zone). Two microplots (0.5 × 0.5 m, with a 1-m—wide buffer zone) were established in the eastern part of each block. Metal squares 0.4 m tall were driven 0.3 m deep into the soil to prevent surface runoff and lateral contamination. 15NH4NO3 (10.2 atom %) and NH 154 NO3 (10.12 atom %; Shanghai Research Institute of Chemical Industry) with the same N addition rate was applied to the two 15N microplots separately within one plot to identify the fate of different N forms (NH +4 -N and NO3 −-N).
N fertilizer as NH4NO3, and water, were added to all plots in late May and June each year from 2009 to 2012. During each application, NH4NO3 was weighed, dissolved in 8 L water, and applied to each block using a sprayer to evenly distribute the fertilizer. At the onset of the study in May 2009, the vegetation of experiment I (with only N treatments) was dominated by perennial grasses (Stipa purpurea, Festuca ovina, Agropyron cristatum and Koeleria cristata) and perennial forbs (Oxytropis glabra, Potentilla multifida and Potentilla bifurca). The experiment II area (combined N and water treatments) was dominated by perennial grass (Elymus nutans and K. cristata) and perennial forbs (Oxytropis glabra, P. multifida and P. bifurca). None of the plots in the two experiments had been grazed since 2005.
Sampling and analysis
Plant AGB was harvested at the soil surface using a 1 × 1-m quadrat (no spatial overlap) in each block at the end of July each year. The plant samples, harvested at peak biomass time, were classed into forbs and grasses. Soil samples were carefully collected at each microplot using a soil corer of 8-cm diameter to a depth of 0.5 m and sieved to remove roots (mesh size 0.2 mm). Soil samples from microplots were air-dried and were ground to pass a 74-µm screen for total N, 15NH4–N, 15NO3–N and isotope analysis. The clean root and aboveground plant material were dried for 48 h at 65 °C before weighing. Species richness is the average number of plant species in the four replicates of each treatment using a 1 × 1-m quadrat.
The percentage of fertilizer N recovery in shoot, root and soil was determined from all microplots using Eqs. 1 and 2 (Liu et al. 2005), where all 15N was expressed as the atom % excess corrected for background abundance (0.3682 % at the alpine grassland site). Shoot, root and soil samples (0–50 cm for plant root and soil) were analyzed for total N and 15N abundance by an elemental analyzer coupled to a mass spectrometer (Delta Plus, Finnigan, Pittsburg, PA).
where NRFP = N recovery from 15N-labeled fertilizer of the plant, NRFS = N recovery from 15N-labeled fertilizer of the soil, atom % 15N excessplant = atom % 15N excess from the plant—atom % 15N excess from background level, atom % 15N excesssoil = atom % 15N excess from the soil—atom % 15N excess from background level, atom % 15N excess fertilizer = atom % 15N excess from the labeled fertilizer N—atom % 15N excess from background level, Nplant = total N in plant (kg ha−1) = plant biomass × total N (%) of plant sample, Nsoil = total N in soil (kg ha−1) = soil bulk density × soil depth × total N amount (%) of soil sample, and Nfertilizer = fertilizer N applied (kg ha−1).
ANOVAs were used to test for main and interactive effects of year, N addition and water supply treatments on AGB and species richness. Analysis of linear regression was performed to test the long-term change (1981–2012) in precipitation. All statistical analyses were performed separately for each experiment with SPSS software 13.0 (SPSS, Chicago, IL). All figures were drawn using Origin software 7.5 (OriginLab, Northampton, MA).
Results
Precipitation variability
MAP, total precipitation (TP; snow and rain between late October and mid July of the subsequent year) and precipitation between late October and April of the subsequent year (SNOW) averaged 282.3, 171.8 and 34.4 mm from 1981 to 2012, respectively (Fig. 1). Rain in the growing season from May to July accounted for 56 % of MAP. MAP was 16.5, 38.1 and 19.5 % above average (282.3 mm) in 2009, 2010 and 2011, but was 0.9 % lower than average in 2012. TP was 13.2, 24.7 and 34.1 % above the average (171.8 mm) in 2009, 2010 and 2011, while it was 9.1 % below the average in 2012 (156.2 mm). SNOW was 109.9 % above the average (34.4 mm) in 2011, but it was 43.9 % below average in 2012 (19.3 mm). MAP significantly increased during the previous 32 years, but TP and SNOW did not (Fig. 1). TP between snow in winter and rain in the growing season has an equal or even more important effect on plant growth in alpine grassland. We therefore define the year 2011 as the “wet year”.

Precipitation change from 1981 to 2012 in alpine grassland of the Tianshan mountains. MAP Mean annual precipitation, TP total precipitation (snow in winter and rain in the growing season between late October and mid July), SNOW snow between late October and April
Aboveground biomass
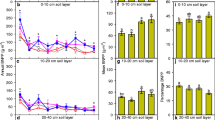
AGB of grasses (GAGB) and forbs (FAGB), respectively, ranged from 27.2 to 199.4 g m−2 and 8.9 to 115.4 g m−2 during the course of the 4-year experiment (Fig. 2). In general, N deposition increased GAGB across N addition treatments at all times and GAGB at the rate of 90 kg N ha−1 year−1 was significantly higher than that of the no N addition treatment in the first 2 years. FAGB was suppressed and significantly lower than GAGB in the lowest snowfall year and at a rate of 30 and 90 kg N ha−1 year−1 (all p ≤ 0.001) in the wettest year. GAGB, FAGB and total GAGB and FAGB was significantly different across the 4 years. No significant interactive effect of N vs. year on AGB was found (Table 1).

Effect of N addition on the aboveground biomass from 2009 to 2012 in alpine grassland of the Tianshan mountains. Significant differences of aboveground biomass between treatments are indicated by different letters. Grasses Perennial grasses, Forbs perennial forbs, Total total perennial grasses and perennial forbs, N 0 0 kg N ha−1 year−1, N 10 10 kg N ha−1 year−1, N 30 30 kg N ha−1 year−1, N 90 90 kg N ha−1 year−1
GAGB and FAGB ranged from 47.2 to 280.5 g m−2 and 5.5 to 61.2 g m−2 across all N and water addition treatments. GAGB significantly increased as an effect of N combined with water addition at all times and was significantly higher than FAGB (Fig. 3). FAGB was significantly decreased only in the wettest year (2011). GAGB was significantly higher than FAGB in the wettest year and lowest snowfall year (all p < 0.001). We found that water supply interacted with N addition on GAGB and total GAGB and FAGB (Table 1).

Effect of increased water and N addition on the aboveground biomass from 2009 to 2012 in alpine grassland of the Tianshan mountains. Significant differences of aboveground biomass between treatments are indicated by different letters. N 0 W 0 0 kg N ha−1 year−1 and 0-mm water supply, N 0 W 1 0 kg N ha−1 year−1 and 7.5-mm water supply, N 30 W 1 30 kg N ha−1 year−1 and 7.5-mm water supply; for other abbreviations, see Fig. 2
Species richness responses
In experiment I, the average species richness was 8.7 and no significant species loss was found across all N addition gradients (Fig. 4). Although species richness was different across the 4 years (Table 1), no significant interactive effect of N vs. year on AGB was found. The average species richness was 7.6 in experiment II and no significant interactive effect of N addition and water supply vs. years on species richness was found (Table 1; Fig. 5).

Effect of N addition on the species richness from 2009 to 2012 in alpine grassland of the Tianshan mountains. For abbreviations, see Fig. 2

Effect of increased water and N addition on species richness from 2009 to 2012 in alpine grassland of the Tianshan mountains. For abbreviations, see Fig. 3
Fate of labeled fertilizer N
The fate of fertilizer 15N is illustrated in Table 2. 15NH4-N recovered by shoot, root and soil ranged from 0.31 to 2.85, 0.79–11.34 and 1.45–9.17 kg ha−1 in N10, N30 and N90, respectively. 15NO3-N recovered by shoot, root and soil ranged from 0.69 to 3.37, 1.0910.67 and 1.20–5.52 kg N ha−1 for the three N addition rates, respectively. Shoot biomass absorbed more 15NO3-N than 15NH4-N. No significant effect was found for the N recovery rate of shoot and root across N deposition gradients. Mean N retained in shoot, root and soil was 8.5, 22.5 and 19.5 %, respectively, and N loss was 49.5 % on average (calculated according to Table 2).
Discussion
N deposition resulted in increased AGB in two semi-arid grasslands in Inner Mongolia (Bai et al. 2010; Song et al. 2012), indicating potential N limitation in these grasslands. In the present study, N addition (N30 and N90) increased GAGB compared with the control (N0), but little additional response was observed for N10. In contrast, N30 and N90 decreased FAGB by 32.0 and 53.0 % in the wettest year and 46.0 and 47.0 % in the lowest snowfall year compared with N0, respectively. These results suggest that: (1) the low-level N addition rate (10 kg N ha−1 year−1) could have had a small effect on the AGB of grasses/forbs in the alpine grassland, (2) GAGB/FAGB increased/decreased at either the mid- or high-level N addition rate in the alpine grassland.
The AGB of semi-arid grassland ecosystems is typically affected by seasonal precipitation (Bai et al. 2004; Guo et al. 2012). For example, AGB increased significantly due to a higher amount of growing season rainfall (Heisler-White et al. 2008). Based on long-term (32-year) observational precipitation, we found 20.0, 19.8 and 15.1 % more precipitation than the average (137.4 mm) during the growing season in 2009, 2010 and 2011, respectively. In 2012 the growing season precipitation (136.9 mm) was close to the average long-term precipitation. Therefore, precipitation during the growing season did not largely contribute to the variations of AGB in the alpine grassland. Snow (from late October to April of the subsequent year) was 44.5 and 109.9 % above the average (34.4 mm) in 2010 and 2011, but 43.9 % below the average in 2012. Changes in winter precipitation have the potential to influence grassland structure and function, especially plant AGB (Chimner et al. 2010). Earlier melt-out dates will cause a considerable shift in species composition favoring higher and faster growing plants in alpine ecosystems (Jonas et al. 2008). Changes in snow depth can influence vegetation growth of the subsequent year through increasing the amount of water and changing the soil thermal regime, microclimate and available N (Peng et al. 2010). Snow addition (+150 %) in winter increased plant biomass by 50 % compared to ambient snow conditions in a prairie of the USA, but plant biomass increased by 44 % with increasing (+50 %) precipitation in summer (Chimner et al. 2010). Therefore, annual snowfall may help explain the AGB variations because snow melt in spring could accelerate plant growth of the subsequent year in the alpine grassland. Precipitation during the growing season may also influence plant AGB production. Precipitation during winter and growing season (January–July) controlled the variations of biomass accumulation in the Inner Mongolia grassland (Bai et al. 2004). In the present study, the mean TP between late October and mid July of the subsequent year was 13.2, 24.7 and 34.1 % higher in 2009, 2010 and 2011, but 9.1 % lower in 2012 than the long-term mean value (171.8 mm), respectively. By contrast to our study, the AGB was positively related to seasonal precipitation in semi-arid grassland with a higher annual temperature because of almost no snow cover in the winter (Cherwin and Knapp 2012). Therefore, TP (precipitation between late October and April of the subsequent year) had a crucial influence on the GAGB/FAGB in the alpine grassland.
Grassland ecosystems are primarily water limited and secondarily N limited (Ladwig et al. 2012). N fertilizer application under wet conditions increased AGB in a semi-arid grassland (Gao et al. 2011). In our study, a positive effect of N addition on GAGB occurred in the wettest year, which was 106.6 and 74.5 % higher at N30 and N90 than at N0, respectively. However, FAGB was decreased by 46 and 47 % in the lowest snowfall year when N was applied at a rate of 30 and 90 kg N ha−1 year−1, respectively. Similar to the above results, GAGB at N30W1 (combination of N with water application) was significantly higher than that at N0W0 (no N and no water addition). Furthermore, GAGB was significantly higher than FAGB at N30W1 at all times (all p < 0.001; Table 1). These results suggest the change of GAGB/FAGB response to N and water addition in alpine grassland is primarily limited by TP (snow in winter and rain in the growing season) and secondarily limited by N. GAGB could have been enhanced by N addition and/or combined N and water supply only in the wet year.
N addition led to a large reduction in species richness in the mature community owing to the increased AGB (Bai et al. 2010). In addition, the increased precipitation notably increased the species richness under N addition due to the re-emergence of annual grass species in the temperate grassland (Yang et al. 2011). However, we found no significant loss of species richness under elevated N deposition and water supply in the alpine grassland. Species competition has usually been used to explain the loss in species richness under increased N deposition (Bessler et al. 2009; Hautier et al. 2009), but low plant cover may result in weak inter-specific competition (Kinugasa et al. 2012). In our study area, the vegetation type is dominated by perennial grasses with almost no annual grass. The species richness was low and the mean species number ranged from six to 10.8 in each quadrat (1 × 1 m) across all plots.
No significant difference was found for the N recovery rate of shoots and roots across N deposition treatments. This indicates that almost half (49.5 %) of the fertilizer N was lost to the environment either through N gases or leaching loss pathways (Song and Farwell 2011; Bergstermann et al. 2011; Butterbach-Bahl et al. 2013; Zhang et al. 2013). Furthermore, coexisting plants might be able to absorb different N forms to avoid competition for resources (Harrison et al. 2007). Recous et al. (1988) reported plant uptake of NO3 − being more efficient than that of NH4 +, and a similar result was found in the present study. We found much more fertilizer N remained in the root and the soil with an increasing N addition rate in the alpine grassland (Table 2). Currie and Nadelhoffer (1999) reported similar results in two temperate forests using the same 15NH4 and 15NO3 labeling isotope technique.
In summary, N addition alone and/or together with early spring water supply significantly increased the AGB of dominant perennial grasses rather than forbs, particularly in the wet year. In contrast, no significant change was found for plant species richness in the Bayinbuluk alpine grassland of the Tianshan mountains. A higher loss of fertilizer N may be a crucial factor explaining the effect of N addition on the AGB and species richness. Precipitation has significantly increased from 1900 to 2005 in central Asia (IPCC 2007), and similar results were also found in the past 32 years in our study area (Fig. 1). The greater GAGB occurred under wet conditions in our study area, i.e., higher TP between late October and mid July of the subsequent year, including snow in winter and rainfall in the growing season. The increase in snowfall in winter and rainfall in the growing season would have increased snow depth, soil water content, and impacted the thermal regime, N availability and microbial activity as a consequence. Therefore, N deposition could increase AGB when the water status is improved, but not species richness, in the alpine grassland.
References
Akmal M, Schellberg J, Khattak RA (2010) Biomass allocation and nitrogen distribution in ryegrass under water and nitrogen supplies. J Plant Nutr 33:1777–1788
Bai YF, Han XG, Wu JG, Chen ZZ, Li LH (2004) Ecosystem stability and compensatory effects in the inner Mongolia grassland. Nature 431:181–184
Bai YF, Wu JG, Clark CM, Naeem S, Pan QM, Huang JH, Zhang LX, Han XG (2010) Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: evidence from inner Mongolia Grasslands. Glob Change Biol 16:358–372
Basher MA, Liu T, Kabir MA, Ntegeka V, Willems P (2010) Climate change impact on the hydrological extremes in the Kaidu river basin, China. 30th Annual USSD Conference, California, pp 637–651
Bergstermann A, Cardenas L, Bol R, Gilliam L, Goulding K, Meijide A, Scholefield D, Vallejo A, Well R (2011) Effect of antecedent soil moisture conditions on emissions and isotopologue distribution of N2O during denitrification. Soil Biol Biochem 43:240–250
Bessler H, Roscher C, Temperton VM, Buchmann N, Schmid B, Schulze ED, Weisser WW, Engels C (2009) Aboveground overyielding in grassland mixtures is associated with reduced biomass partitioning to belowground organs. Ecology 90:1520–1530
Bobbink R, Hicks K, Galloway J, Spranger T, Alkemade R, Ashmore M, Bustamante M, Cinderby S, Davidson E, Dentener F, Emmett B, Erisman JW, Fenn M, Gilliam F, Nordin A, Pardo L, De Vries W (2010) Global assessment of nitrogen deposition effects on terrestrial plant diversity: a synthesis. Ecol Appl 20:30–59
Brookshire ENJ, Gerber S, Menge DN, Hedin LO (2012) Large losses of inorganic nitrogen from tropical rainforests suggest a lack of nitrogen limitation. Ecol Lett 15:9–16
Butterbach-Bahl K, Baggs EM, Dannenmann M, Kiese R, Scophie Zechmeister-Boltenstern (2013) Nitrous oxide emissions from soils: how well do we understand the process and their controls? Philos Trans R Soc B 368:20130122
Cherwin K, Knapp A (2012) Unexpected patterns of sensitivity to drought in three semi-arid grasslands. Oecologia 169:845–852
Chimner RA, Welker JM (2005) Ecosystem respiration responses to experimental manipulations of winter and summer precipitation in a mixed grass prairie, WY, USA. Biogeochemistry 73:257–270
Chimner RA, Welker JM, Morgan J, LeCain D, Reeder J (2010) Experimental manipulations of winter snow and summer rain influence ecosystem carbon cycling in a mixed-grass prairie, Wyoming, USA. Ecohydrology 3:284–293
Clark CM, Tilman D (2008) Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands. Nature 451:712–715
Currie WS, Nadelhoffer KJ (1999) Dynamic redistribution of isotopically labeled cohorts of nitrogen inputs in two temperate forests. Ecosystems 2:4–18
DeFalco LA, Bryla DR, Smith-Longozo V, Nowak RS (2003) Are Mojave desert annual species equal? Resource acquisition and allocation for the invasive grass Bromus madritensis subsp. rubens (Poaceae) and two native species. Am J Bot 90:1045–1053
Duparc A, Redjadj C, Viard-Cretat F, Lavorel S, Austrheim G, Loison A (2013) Co-variation between plant above-ground biomass and phenology in sub-alpine grasslands. Appl Veg Sci 16:305–316
Fisk MC, Schmidt SK, Seastedt TR (1998) Topographic patterns of above- and belowground production and nitrogen cycling in alpine tendra. Ecology 79:2253–2266
Galloway JN, Townsend AR, Erisman JW, Bekunda M, Cai ZC, Freney JR, Martinelli LA, Seitzinger SP, Sutton MA (2008) Transformation of the nitrogen cycle: recent trends, questions, and potential solutions. Science 320:889–892
Gao YZ, Chen Q, Lin S, Giese M, Brueck H (2011) Resource manipulation effects on net primary production, biomass allocation and rain-use efficiency of two semiarid grassland sites in inner Mongolia, China. Oecologia 165:855–864
Gaudnik C, Corcket E, Clment B, Delmas CE, Gombert Courvoisier S, Muller S, Alard D (2011) Detecting the footprint of changing atmospheric nitrogen deposition loads on acid grasslands in the context of climate change. Glob Change Biol 17:3351–3365
Guo Q, Hu ZM, Li SG, Sun XM, Yu GR (2012) Spatial variations in aboveground net primary productivity along a climate gradient in Eurasian temperate grassland: effects of mean annual precipitation and its seasonal distribution. Glob Change Biol 18:3624–3631
Harrison KA, Bol R, Bardgett RD (2007) Preferences for different nitrogen forms by coexisting plant species and soil microbes. Ecology 88:989–999
Hautier Y, Niklaus PA, Hector A (2009) Competition for light causes plant biodiversity loss after eutrophication. Science 324:636–638
Heisler-White JL, Knapp AK, Kelly EF (2008) Increasing precipitation event size increases aboveground net primary productivity in a semi-arid grassland. Oecologia 158:129–140
Huxman TE, Smith MD, Fay PA, Knapp AK, Shaw MR, Loik ME, Williams DG (2004) Convergence across biomes to a common rain-use efficiency. Nature 429:651–654
IPCC (2007) Fourth assessment report, climate change 2007: synthesis report. Cambridge University Press, Cambridge
Jonas T, Rixen C, Sturm M, Stoeckli V (2008) How alpine plant growth is linked to snow cover and climate variability. J Geophys Res 113:G03013
Kinugasa T, Tsunekawa A, Shinoda M (2012) Increasing nitrogen deposition enhances post-drought recovery of grassland productivity in the Mongolian steppe. Oecologia 170:857–865
Ladwig LM, Collins SL, Swann AL, Xia Y, Allen MF, Allen EB (2012) Above-and belowground responses to nitrogen addition in a Chihuahuan desert grassland. Oecologia 169:177–185
Li JZ, Lin S, Taube FH, Pan QM, Dittert K (2011) Above and belowground net primary productivity of grassland influenced by supplemental water and nitrogen in inner Mongolia. Plant Soil 340:253–264
Li KH, Gong YM, Song W, Lv JL, Chang YH, Hu YK, Tian CY, Christie P, Liu XJ (2012a) No significant nitrous oxide emissions during spring thaw under grazing and nitrogen fertilization in an alpine grassland. Glob Change Biol 18:2546–2554
Li KH, Gong YM, Song W, He GX, Hu YK, Tian CY, Liu XJ (2012b) Responses of CH4, CO2 and N2O fluxes to increasing nitrogen deposition in alpine grassland of the Tianshan mountains. Chemosphere 88:140–143
Liu XJ, Ai YW, Zhang FS, Lu SH, Zeng XZ, Fan MS (2005) Crop production, nitrogen recovery and water use efficiency in rice-wheat rotation as affected by non-flooded mulching cultivation (NFMC). Nutr Cycl Agroecosyst 71:289–299
Liu XJ, Zhang Y, Han WX, Tang AH, Shen JL, Cui ZL, Vitousek P, Erisman JW, Goulding K, Christie P, Fangmeier A, Zhang FS (2013) Enhanced nitrogen deposition over China. Nature 494:459–462
Peng SS, Piao SL, Ciais P, Fang JY, Wang XH (2010) Change in winter snow depth and its impacts on vegetation in China. Glob Change Biol 16:3004–3013
Recous S, Fresneau C, Faurie G, Mary B (1988) The fate of labeled 15N urea and ammonium nitrate applied to a winter wheat crop. 1. Nitrogen transformations in the soil. Plant Soil 112:205–214
Reichmann LG, Sala OE, Peters DPC (2013) Precipitation legacies in desert grassland primary production occur through previous-year tiller density. Ecology 94:435–443
Song XZ, Farwell SO (2011) Application of pyrolysis cryogenic trap gas chromatography/atomic emission detection (Py-CryT-GC/AED) for the study of nitrogen forms and dynamics in a grassland soil. Anal Lett 44:2746–2759
Song L, Bao X, Liu X, Zhang Y, Christie P, Fangmeier A, Zhang F (2011) Nitrogen enrichment enhances the dominance of grasses over forbs in a temperate steppe ecosystem. Biogeosciences 8:2341–2350
Song L, Bao XM, Liu XJ, Zhang FS (2012) Impact of N addition on plant community in a semi-arid temperate in China. J Arid Land 4:3–10
Stevens CJ, Dise NB, Mountford JO, Gowing DJ (2004) Impact of nitrogen deposition on the species richness of grasslands. Science 303:1876–1879
Yang HJ, Li Y, Wu MY, Zhang Z, Li LH, Wan SQ (2011) Plant community responses to nitrogen addition and increased precipitation: the importance of water availability and species traits. Glob Change Biol 17:2936–2944
Zhang XB, Zwiers FW, Hegerl GC, Lambert FH, Gillett NP, Solomon S, Stott PA, Nozawa T (2007) Detection of human influence on twentieth-century precipitation trends. Nature 448:461–465
Zhang YH, He NP, Zhang GM, Huang JH, Wang QB, Pan QM, Han XG (2013) Ammonia emissions from soil under sheep grazing in inner Mongolian grasslands of China. J Arid Land 5:155–165
Acknowledgments
This work was financially supported by the Chinese National Basic Research Program (2014CB954202), the National Natural Science Foundation of China (41340041), the Distinguished Talent Program of NSFC, the One-hundred Talents Program of the Chinese Academy of Sciences, and an Innovative Group Grant (31121062). We gratefully acknowledge Dr Arvin Mosier and Dr Daniela Famulari for linguistic corrections and two anonymous reviewers for their constructive comments on earlier versions of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Hormoz BassiriRad.
Rights and permissions
About this article

Cite this article
Li, K., Liu, X., Song, L. et al. Response of alpine grassland to elevated nitrogen deposition and water supply in China. Oecologia 177, 65–72 (2015). https://doi.org/10.1007/s00442-014-3122-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-014-3122-4