
Abstract
Toxoplasma gondii can infect a wide range of warm-blooded animals, causing a global toxoplasmosis zoonotic epidemic. Surface antigen 1 (SAG1) protein is expressed at the proliferative tachyzoite stage, whereas matrix antigen 1 (MAG1) is expressed at the bradyzoite and tachyzoite stages. These two proteins were found to perform protective roles in previous studies; however, their synergetic protective efficacy as a DNA vaccine against toxoplasmosis has not been clarified. In this study, we constructed recombinant pcDNA3.1( +)-TgMAG1 (pMAG1), pcDNA3.1( +)-TgSAG1 (pSAG1), and pcDNA3.1( +)-TgMAG1-TgSAG1 (pMAG1-SAG1) plasmids and administered them intramuscularly to immunize mice. The levels of anti-T. gondii IgG in serum and cytokines, such as Interleukin (IL)-4, IL-10, and Interferon (IFN)-γ, in splenocytes were measured using ELISA and the respective culture supernatants. Lethal doses of T. gondii (type I) RH strain tachyzoites were administered to immunized mice, and mortality was assessed. Conversely, mice infected with low doses of tachyzoites were monitored to determine their survival rates, and parasite burden analyses of the brains and livers were conducted. The bivalent TgMAG1 and TgSAG1 DNA vaccines exhibited excellent protective immunity against toxoplasmosis in mice, with higher serum IgG and splenocyte IFN-γ release levels, longer survival days, and reduced parasite burden in the brain and liver tissues (p < 0.05). These findings provide a new perspective for the development of T. gondii vaccines.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Toxoplasma gondii (T. gondii), a member of the phylum, Apicomplexa, which comprises intracellular protozoan parasite, can infect humans, especially immunocompromised individuals, and other warm-blood animals, leading to zoonotic toxoplasmosis (Dubey 2008). As an opportunistic pathogen, the human infection rate of T. gondii ranges from 10 to 60%, and in some high-virulence areas, this rate approaches 95% (Pappas et al. 2009; Almería et al. 2018; Li and Zhou 2018). In general, at least one-third of people worldwide have a history of T. gondii infection, making it a public health threat (Saadatnia and Golkar 2012). When pregnant individuals are infected with T. gondii, this can result in harm to the fetus, and may even lead to abortion (Milne et al. 2020). Chronic and long-term T. gondii infections may also trigger severe symptoms, such as encephalitis or neurological symptoms in immune systems and young individuals with congenital infection (Rosenberg et al. 2009; Dubey 2010; Ahmadpour et al. 2017). However, due to their toxic effects, currently available toxoplasmosis medication options are not recommended as treatments. Therefore, effective immunizations are far more crucial for toxoplasmosis prevention. There are three developmental stages in the life cycle of T. gondii that are infective: the tachyzoite stage, bradyzoite stage, and sporozoite stage. The tachyzoite stage occurs in acute infection cases, whereas the bradyzoite stage occurs in chronic infection cases. The sporozoite stage only occurs in a definitive host under sexual reproduction conditions. Sporozoites infect humans and animals and are transformed into tachyzoites upon infection, followed by a transformation into bradyzoites (Attias et al. 2020). These three stages are important for the development of antigen targets against toxoplasmosis.
Surface antigen 1 (SAG1) has garnered increasing attention as it can interact with host cells and participate in the process of adhesion, invasion, and regulation of host immune responses. In a previous study, subunit vaccines based on the SAG1 antigen were developed and found to exhibit good performance in the protection of limiting maternofetal transmission (Letscher-Bru et al. 2003). However, the selection of adjuvants to boost immunogenicity for the induction of long-lasting protective immunity proved challenging (Chuang et al. 2013). SAG1-based DNA vaccines were verified to effectively prevent lethal T. gondii infection (Nielsen et al. 1999; Ivory and Chadee 2004). Moreover, to obtain enhanced protection, SAG1 was combined with rhoptry antigen 2 (ROP2) to generate a multivalent vaccine that significantly blocked T. gondii infection (Fachado et al. 2003). Matrix antigen 1 (MAG1) is a type of dense granular protein that forms the parasitophorous vacuolar matrix in tachyzoite vacuoles. MAG1 is also abundant in the cyst wall and matrix in bradyzoite vacuoles (Ferguson and Parmley 2002; Parmley et al. 2002). Previously, MAG1 was found to induce cell responses throughout the T. gondii infection periods (Cristina et al. 2004). Immunization with MAG1 induced immune protective effects in a murine model (Parmley et al. 2002). In infected humans and pigs, high levels of anti-MAG1 IgG antibody generation were detected (Gamble et al. 2000). MAG1-deletion T. gondii infection had obviously reduced virulence in the acute stage and led to almost no brain cysts in the chronic stage. The immunomodulatory effect of MAG1 favored the survival of T. gondii and the establishment of latent infection occurred via the suppression of inflammasome activation (Tomita et al. 2021). Based on these findings, MAG1 is a suitable target for toxoplasmosis and possesses good immunogenic properties. However, the effect of the bivalent DNA vaccine of T. gondii SAG1 and MAG1 has not been determined.
In this study, pMAG1, pSAG1, and pMAG1-SAG1 plasmids were constructed and used as DNA vaccines to immunize mice. The specific antibody generation levels were determined in immunized mice by measuring serum anti-T. gondii IgG levels and splenocyte-secreted cytokine levels. The protective efficacy was determined by analyzing the survival rate and parasite burden in T. gondii-challenged mice after immunization. To our knowledge, this is the first study to assess the protective ability of TgMAG1 and TgSAG1 bivalent DNA vaccines in mice to lay a foundation for the development of protective strategies against toxoplasmosis.
Materials and methods
Ethics statements
The T. gondii passage assays were performed using female Kuming mice (8–12 weeks old, SPF grade).
For the vaccination experiments, 6–8-week-old BALB/c mice (SPF grade) were used. All mice were administered an adequate diet and handled in strict accordance with the guidelines of the Experimental Animals Center, Jilin University, Changchun, China. All mouse experiments strictly complied with the animal ethics enforcement of the Animal Welfare and Research Ethics Committee of Jilin University (Permit Number: pzpx20190929065).
Cells, parasites, and STAg preparation
HEK-293 cells were conserved in our laboratory. Cells were cultured using complete DMEM (Invitrogen, California) containing 10% FBS and used for plasmid transfection. The highly virulent T. gondii (type I) RH strain was maintained in our lab. T. gondii tachyzoites were used for plasmid construction and mice challenge. Tachyzoites were enriched in infected Kunming mice and the peritoneal fluid was harvested (Chen et al. 2013; Zheng et al. 2013). Cell debris was removed via low-speed centrifugation, and tachyzoites were collected from the supernatant via high-speed centrifugation. The tachyzoites pellets were resuspended in PBS, frozen at − 80 ℃, and melted at 4 ℃ in three cycles, subjected to thorough ultrasonic fragmentation, and centrifuged at 10,000 × g for 30 min. Finally, STAg in the supernatants was collected after filtration with a 0.22-µm pore-size filter membrane.
Preparation of recombinant plasmids of pMAG1, pSAG1, or pMAG1-SAG1
To construct the eukaryotic expression plasmids of pMAG1, pSAG1, and pMAG1-SAG1, gene-specific primers were designed to target the sequences (Genbank Nos. AF251813 and S76248). The primer sets are presented in Table 1 and were synthesized by Sangon Biotech (Changchun). T. gondii tachyzoites RNA was obtained using Trizol (TaKaRa, Dalian). cDNA was prepared using M-MLV reverse transcriptase (TaKaRa). The TgMAG1 and TgSAG1 DNA fragments were cloned via PCR using T. gondii cDNA and then purified using the TIANgel Midi Purification Kit (TIANGEN, Beijing). The purified DNA fragments were inserted into the pMD-18-T vector (TaKaRa). The PCR identified positive plasmids of pMD-MAG1 and pMD-SAG1 were further verified via sequencing. The plasmids of pMD-MAG1 and pcDNA3.1 (Invitrogen) were separately linearized using EcoR I and Hind III restriction enzymes, dephosphorylated using Fast AP (Invitrogen), and linked with T4 DNA ligase (TaKaRa) according to the manufacturer’s instructions. The positive recombinant expression plasmid of pcDNA3.1( +)-TgMAG1 (pMAG1) was identified via PCR (516 bp size) using the primer sets of MAG1-F and MAG1-R. Similar procedures were carried out to clone the pSAG1 fragment into the pcDNA3.1( +) vector to generate the pcDNA3.1( +)-TgSAG1 (pSAG1) recombinant plasmid, with Hind III/Xho I as the restriction enzyme set. The positive pSAG1 plasmid was identified via PCR (450 bp size) using the primer sets of SAG1-F and SAG1-R. Fragments of MAG1 and SAG1 were cloned into the EcoR I/Xho I linearized pcDNA3.1( +) vector to generate pcDNA3.1( +)-TgMAG1-TgSAG1 (pMAG1-SAG1), with EcoR I/Hind III and EcoR I/Xho I as the respective restriction enzyme sets. The positive pMAG1-SAG1 plasmid was identified via PCR (966 bp size) using the primer sets of MAG1-F and SAG1-R. The endotoxin-free plasmid for the transfection assays was prepared using the EasyPure HiPure Plasmid MiniPrep Kit (EM111, TIANGEN).
Expression of MAG1, SAG1, MAG1, and SAG1 in HEK-293 cells
The endotoxin-free plasmids of pMAG1, pSAG1, and pMAG1-SAG1 and the empty vector were expressed in HEK-293 cells through transfection assays with lipofectamine 2000 (Invitrogen). The protein expression levels of MAG, SAG1, MAG, and SAG1 were detected via western blot. Cells were collected at 48 h and fully lysed using RIPA buffer (Beyotime) for 30 min on ice. Thereafter, the mixture was centrifuged at 12,000 × g for 3 min and the supernatants were mixed with protein loading buffer. After boiling at 100 ℃ for 10 min, the prepared protein samples were separated via SDS-PAGE and transferred onto the NC membrane under the conditions of 200 mA/70 min. The membranes were then blocked in 5% skim milk at room temperature for 2 h and incubated with the primary antibody (T. gondii monoclonal antibody; 1:100 dilution; J26A; Invitrogen) at 37 ℃ for 1 h. After three washes with TBST buffer, the membranes were incubated with the secondary detection antibody (HRP-labeled goat anti-mouse IgG (H + L) secondary antibody; 1:500 dilution; 31,430; Invitrogen) at 37 ℃ for 1 h. After three washes with TBST buffer, the membranes were developed using ECL reagent (Vazyme).
Mice immunization
The endotoxin-free plasmids of pMAG1, pSAG1, and pMAG1-SAG1 and the empty vector of pcDNA3.1 were separately administered via intramuscular injection (anterior tibial muscle) into BALB/c mice (100 μL/mice). Mice were randomly selected and divided into the pMAG1 immunity group (100 μg/mice, i.m.), pSAG1 immunity group (100 μg/mice, i.m.), pMAG1-SAG1 immunity group (100 μg/mice, i.m.), pcDNA3.1 treatment group (100 μg/mice, i.m.), and PBS treatment group (100 µL/mice), with 21 mice per group. Mice in each group were administered three injections on the last day of weeks 0, 2, and 4, respectively. Prior to immunization, blood samples were collected, and serum samples were separated for further use.
Measurement of the specific anti-T. gondii IgG antibody
To determine the pMAG1, pSAG1, and pMAG1-SAG1-triggered anti-T. gondii IgG antibody levels, ELISA was conducted. STAg with a concentration of 10 µg/mL was added to the ELISA plates and incubated overnight at 4 °C. The ELISA plates were removed on alternate days and coated with 5% bovine serum albumin (BSA) for 2 h at room temperature. The plates were then incubated with mouse tail vein serum (1:500 dilution) for 1 h at 37 °C, followed by HRP-labeled goat anti-mouse IgG (1:2000 dilution, Sigma-Aldrich) for 1 h. A substrate solution containing 15 μL H2O2, 10 mL citrate phosphate, and 4 mg o-phenylenediamine was dispensed into the wells of the plate for 30 min at room temperature; the reaction was then terminated using 2 M H2SO4. The absorptance at 490 nm was measured and all samples were measured in triplicate. When the OD490 nm value of each group was lower than twice the average of the PBS group, it was judged as negative; however, when the OD490 nm value was higher than twice the average of the PBS group, it was judged as positive.
Measurement of cytokine release from splenocyte supernatants
To evaluate cytokine production, splenocytes were collected 2 weeks after the last immunization (before the challenge) from three mice in each group. Briefly, fresh spleen tissues were isolated and prepared into a suspension using a wire mesh. The red blood cells in the mixture were removed using the RBC lysis solution (11,814,389,001, Sigma-Aldrich). The purified splenocyte suspensions were prepared with DMEM containing 10% FBS and seeded at 1 × 105 cells/well in 96-well tissue culture plates (Costar, Henan). After STAg (5 mg/mL) stimuli treatment, the cell supernatants were collected at 1 day, 3 days, and 4 days, and analyzed to determine the IL-4, IL-10, and IFN-γ contents using the appropriate commercial ELISA kits (M4000B, M1000B, MIF00, R&D systems, Minnesota).
Monitoring of immunized mice survival time and parasite burden after challenge with T. gondii
Fifteen mice were challenged with 1 × 104 tachyzoites (type I) RH strain in each group at week 6. Mice that could not consume food and water or had a body weight loss of more than 20% were killed. Finally, twelve mice were randomly selected to monitor daily mortality, record the number of days survived, and compare the experimental groups with the control groups. The brain tissues and liver tissues were isolated from the remaining three mice on day 3 after the challenge among the experimental groups and control groups. The TIANamp Genomic DNA Kit (TIANGEN) was used to extract genomic DNA from the brain tissue and liver tissue according to the instructions. Parasite burden was quantified using a real-time fluorescence quantitative PCR (qPCR) method described previously (Contini et al. 2005) and SYBR Green I qPCR mix (Monad). The qPCR primer sets for the T. gondii B1 gene were 5′-AACGGGCGAGTAGCACCTGAGGAGA-3′ (forward primer) and 5′-TGGGTCTACGTCGATGGCATGACAAC-3′ (reverse primer). The amplification system contained 10 μL of SYBR Green qPCR Mix, 200 ng of template DNA, 0.8 μL of primer sets (10 μM), and ddH2O up to 20 μL. The following amplification program was employed: 95 ℃ for 30 s followed by 30 cycles of 95 ℃ for 15 s and 60 ℃ for 30 s. The melt curve was set as the default on the light cycler 480 machine (Roche). The parasite burden per mg of tissue was calculated to determine the final rate of parasite reduction relative to that of the control group.
Statistical analysis
All data were analyzed using SPSS 14.0 and one-way analysis of variance (ANOVA). P values less than 0.05 were considered to indicate a significant difference. The same lowercase letters indicate no significant difference, whereas different lowercase letters indicate a significant difference. Graphs or survival charts were generated using GraphPad Prism 7.00 (GraphPad Software Inc., La Jolla, CA, USA).
Results
Identification of the protein expression of MAG1, SAG1, MAG1, and SAG1
The detailed inserted sequences and translated proteins are listed in supplementary file 1. The pMAG1, pSAG1, and pMAG1-SAG1 plasmids were transformed into HEK-293 cells. After 48 h of incubation, the cell lysates were quantified using the BCA method. Equal amounts of cell lysates (30 μg) were used for SDS-PAGE. The proteins were transferred to NC membranes and analyzed for protein expression based on a reaction with the anti-T. gondii antibody. The results are shown in Fig. 1; the pMAG1, pSAG1, or pMAG1-SAG1 transfection cell lysates exhibited well reactogenicity with the anti-T. gondii antibody and had the expected protein sizes of 18 kDa, 16 kDa, and 33 kDa, respectively. No specific reaction bands were observed for the HEK-293 and pcDNA3.1( +) cell lysates. These data indicate that pMAG1, pSAG1, and pMAG1-SAG1 were successfully expressed in vitro.

pMAG1, pSAG1, and pMAG1-SAG1 protein expression analysis in vitro
HEK-293 cells were transfected with pcDNA3.1( +) or the pMAG1, pSAG1, and pMAG1-SAG1 plasmids for 48 h. The cell lysates were electrophoresed on 12% SDS-PAGE gels, transferred onto NC membranes, and reacted with an anti-T. gondii antibody. Each transfection plasmid is indicated on its corresponding lane. M represents pre-stained protein Marker III (Blue, MP121221, TIANGEN).
Specific IgG antibody response
Three mice were selected from every group and serum was collected on the last day of weeks 0, 2, 4, and 6, and used to determine the levels of anti-T. gondii IgG via ELISA. The results are shown in Fig. 2. Mice treated with PBS or pcDNA3.1 ( +) control could generate significantly lower levels of IgG antibodies than those administered pMAG1, pSAG1, or pMAG1-SAG1 (p < 0.01). The levels of IgG in the single antigenic groups (pMAG1 or pSAG1) were lower than those in the bivalent antigenic groups (pMAG1-SAG1, p < 0.05); however, no significant difference was found between the pMAG1 group and pSAG1 group (p > 0.05).

Measurement of the specific anti-T. gondii IgG level in the serum of inoculated mice using ELISA
Immune sera were collected from 3 mice on the last day of weeks 0, 2, 4, and 6 after the first immunization. Results are presented as the means of OD 490 ± SD. Statistical differences among the five groups at each timepoint were determined using one-way ANOVA and indicated by lowercase letters. P values less than 0.05 were considered to indicate a significant difference. The same lowercase letters indicate no significant difference, whereas different lowercase letters indicate a significant difference.
Cytokine production
Splenocytes were prepared 2 weeks after the last immunization of mice and the culture supernatants were collected 24 h, 72 h, and 96 h after STAg stimulation. The IFN-gamma, IL-4, and IL-10 levels were determined and the results are displayed in Fig. 3. The IFN-γ release levels generated by splenocytes were higher in the pMAG1, pSAG1, or pMAG1-SAG1 immunized groups than the pcDNA3.1 and PBS control groups (p < 0.01). Moreover, the IFN-γ release levels in pMAG1-SAG1 immunized mice were obviously higher than those in pMAG1 or pSAG1 immunized mice (p < 0.05). However, the IL-4 and IL-10 levels did not significantly differ among the experimental groups.

Production of cytokines in the splenocytes of immunized mice (n = 3)
Fresh spleen tissue was removed from mice that had been immunized for 2 weeks, prepared into splenocyte suspensions, and stimulated to measure IFN-γ (at 96 h), IL-4 (at 24 h), and IL-10 (at 72 h). Statistical differences among the five groups at each timepoint were determined using one-way ANOVA. There was no significant difference (P < 0.05) among the five groups for the IL-4 and IL-10 values, and no signs were labeled. For the IFN-gamma values, statistical analysis was performed using the five groups. The same lowercase letters indicate no significant difference, whereas different lowercase letters indicate significant difference.
Protective efficacy of pMAG1, pSAG1, or pMAG1-SAG1
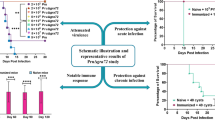
To determine the protective efficacy of the DNA vaccines, mice (n = 10) in all groups were challenged with lethal doses of T. gondii tachyzoites (1 × 104/mice) at week 2 after the final immunization. Thereafter, the survival days and percentages in different groups were monitored. As shown in Fig. 4, pMAG1, pSAG1, and pMAG1-SAG1 immunization significantly prolonged the survival days of mice compared to the pcDNA3.1 and PBS control (p < 0.05). Death of both pcDNA3.1 and PBS control mice occurred within 4–7 days. In contrast, the survival days of mice immunized with pMAG1-SAG1 increased to 16–32 days while that of mice immunized with pMAG1 or pSAG1 extended to 7–16 days. The differences among the immunization groups were analyzed and the results revealed that pMAG1-SAG1 immunization had a better performance in prolonging the survival time than pMAG1 or pSAG1 immunization. All mice were dead within 32 days post-infection.

Survival curves of immunized mice after challenge with 1 × 104 tachyzoites (lethal dose) at week 2 after the last immunization. Each group comprised ten mice. Mice were monitored daily to determine their survival time over 32 days after the challenge
To determine the number of parasite burden in mice brains and livers, 1 × 104 T. gondii tachyzoites were administered to all groups of mice (n = 3) at week 2 after the final immunization. Real-time PCR was used to quantify the number of parasites in the brain and liver of three mice in every group. As shown in Table 2, mice-administered pMAG1, pSAG1, and pMAG1-SAG1 had a significant reduction in the parasite in all tissues compared to mice administered the control (pcDNA3.1 and PBS). The parasite number in the pMAG1-SAG1 group was significantly reduced compared to that in the pMAG1 and pSAG1 groups. No significant differences were found between the pMAG1 and pSAG1 groups in terms of parasite number.
Discussion
DNA vaccines were first developed in the 1990s, serving as the third generation of vaccine after pathogen vaccines and subunit vaccines. The nucleic acid vaccine can express the target molecules in the eukaryotic system and simulate their natural counterparts to effectively induce immune responses in hosts. The nucleic acid vaccine has the excellent advantages of simple preparation, low cost, and friendliness to operators, and is suitable for large-scale production. DNA vaccination has been verified to share protective roles against pathogenic microorganisms, especially the intracellular parasite T. gondii, in animals and humans (Warner et al. 2021). T. gondii undergoes several stages and develops diverse morphologies during its life cycle. Previous studies revealed that hosts vaccinated with different stage-specific targets would cause limited growth of T. gondii and protect hosts from infection (Mévélec et al. 2005; Liu et al. 2009). DNA vaccines with multi-antigens have been found to exhibit better immune effects than vaccines comprising a single antigen for the protection against toxoplasmosis (Fachado et al. 2003; Beghetto et al. 2005). Thus, the development of a DNA vaccine with multi-antigens that are involved in the growth stage and invasion stage will compensate for the use of a single antigen as a DNA vaccine. SAG1 located on the surface of T. gondii tachyzoites was reported to induce effective and durable humoral and cellular immune responses in immunized mice (Nielsen et al. 1999; Liu et al. 2006; Siachoque et al. 2006). In contrast, MAG1 was found to localize in the matrix and the wall of tissue cysts and is synthetized in tachyzoites (Ferguson and Parmley 2002; Parmley et al. 2002). By analyzing the antibody response against BAG1 and MAG1 antigen, the specific response against bradyzoites was found to be triggered early at the time of infection in humans (Cristina et al. 2004). Based on research on SAG1 wrapped by virus-like particle antigen for delivery, this system could trigger anti-T. gondii antibody generation in mouse, thereby preventing toxoplasmosis (Choi and Park 2020). In this study, we prepared a T. gondii bivalent DNA vaccine comprising MAG1 and SAG1 which are expressed in tachyzoites growth stage and tissue cysts infectious stages and determined whether this vaccine could serve as a potent vaccine against acute and chronic T. gondii infection in mice. Single SAG1 or MAG1 immunization was also employed as controls to compare the bivalent DNA vaccine effects with those of the single DNA vaccine. Mice immunized with the pSAG1, pMAG1, or pMAG1-SAG1 plasmids produced specific antibodies against T. gondii, which induced protection against T. gondii challenge. Moreover, immunization with the multi-antigen plasmid, pMAG1-SAG1, induced a stronger humoral and cellular immune response, increased the survival days, and decreased parasite burden in the brain and liver tissues of T. gondii tachyzoites-challenged mice compared with mice administered a single antigen.
The level of anti-T. gondii IgG antibody was determined in this study. Significant increases in IgG levels were observed in the pSAG1-MAG1 group at week 4 after immunization, and reached the highest level at week 6. The three DNA vaccines were suggested to produce strong parasite-specific humoral immunity against T. gondii, especially the multi-antigen DNA vaccine. Cytokines play important roles in regulating T helper cell activities. According to their function difference, cytokines are mainly classified into Th1 type and Th2 type cytokines. IFN-γ is known as a dominant product of Th1-type cytokines that suppress Th2-type immune responses, while the presence of IL-4 in the early stages of infection will inhibit the differentiation of protective Th1 cells (Nan et al. 2007; Nakagome et al. 2009). IFN-γ and IL-4 act as regulatory factors and mediate the dynamic balance between Th1 and Th2 immune responses (Torres et al. 2004). In this study, immunization with the pSAG1-MAG1 vaccine led to high levels of IFN-γ, which significantly enhanced Th1 cell-mediated immunity compared with immunization with the other vaccines. However, there was no significant difference in the production of IL-4 and IL-10 between the groups.
Survival days and parasite burden are the most important parameters during the assessment of the protective effect of pSAG1, pMAG1, and pSAG1-MAG1 vaccines against T. gondii challenge. Several studies revealed that vaccination with recombinant proteins encoding T. gondii antigens displayed certain protection, increased the survival time of animals, and reduced the parasite burden. These vaccines comprised the DNA vaccines encoding TgGRA14 (Ahmadpour et al. 2017), TgROP5 and TgGRA15 (Chen et al. 2015), TgROP5 and TgROP18 (Grzybowski et al. 2015), and TgGRA1 and TgMIC3 (Gong et al. 2016). In the present study, BALB/c mice were challenged with a lethal dose of 1 × 104 tachyzoites of T. gondii RH strain to assess the survival rate and parasite reduction. Mice immunized with the pSAG1-MAG1 DNA vaccine had prolonged survival time and caused fewer parasites burden in the brain and liver tissues compared with the other vaccines. In terms of protection against the challenge of T. gondii, the multi-antigen DNA vaccine was found to perform better and more effectively than the single-agent vaccine in this study.
Conclusion
Taken together, the production of specific cellular and humoral immune responses in BALB/c mice can be induced by pSAG1, pMAG1, and pSAG1-MAG1 DNA vaccine immunization. The T. gondii bivalent MAG1 and SAG1 DNA vaccine provided more enhanced immunity against T. gondii infection in BALB/c mice, which suggested its potential as an anti-T. gondii vaccine. Other potent effective antigens, suitable adjuvants, and other animal models can also be investigated in future research.
Data availability
The data generated or analyzed in this study are included in this article. If further clarification is needed, requests may be directed to the corresponding author.
References
Ahmadpour E, Sarvi S, Soteh MBH, Sharif M, Rahimi MT, Valadan R, Tehrani M, Khalilian A, Montazeri M, Daryani A (2017) Evaluation of the immune response in balb/c mice induced by a novel DNA vaccine expressing gra14 against Toxoplasma gondii. Parasite Immunol 39
Almería S, Cabezón O, Paniagua J, Cano-Terriza D, Jiménez-Ruiz S, Arenas-Montes A, Dubey JP, García-Bocanegra I (2018) Toxoplasma gondii in sympatric domestic and wild ungulates in the mediterranean ecosystem. Parasitol Res 117:665–671
Attias M, Teixeira DE, Benchimol M, Vommaro RC, Crepaldi PH, Souza WD (2020) The life-cycle of Toxoplasma gondii reviewed using animations. Parasit Vectors 13:588
Beghetto E, Nielsen HV, Porto PD, Buffolano W, Guglietta S, Felici F, Petersen E, Gargano N (2005) A combination of antigenic regions of Toxoplasma gondii microneme proteins induces protective immunity against oral infection with parasite cysts. J Infect Dis 191:637–645
Chen J, Huang SY, Li ZY, Yuan ZG, Zhou DH, Petersen E, Zhang NZ, Zhu XQ (2013) Protective immunity induced by a DNA vaccine expressing eIF4A of Toxoplasma gondii against acute toxoplasmosis in mice. Vaccine 31:1734–1739
Chen J, Li ZY, Petersen E, Huang SY, Zhou DH, Zhu XQ (2015) DNA vaccination with genes encoding Toxoplasma gondii antigens rop5 and gra15 induces protective immunity against toxoplasmosis in kunming mice. Expert Rev Vaccines 14:617–624
Choi WH, Park JS (2020) Immunogenicity and protective effect of a virus-like particle containing the SAG1 antigen of Toxoplasma gondii as a potential vaccine candidate for toxoplasmosis. Biomedicines 8:91
Chuang SC, Ko JC, Chen CP, Du JT, Yang CD (2013) Induction of long-lasting protective immunity against Toxoplasma gondii in balb/c mice by recombinant surface antigen 1 protein encapsulated in poly (lactide-co-glycolide) microparticles. Parasit Vectors 6:34
Contini C, Seraceni S, Cultrera R, Incorvaia C, Sebastian A, Picot S (2005) Evaluation of a real-time pcr-based assay using the lightcycler system for detection of Toxoplasma gondii bradyzoite genes in blood specimens from patients with toxoplasmic retinochoroiditis. Int J Parasitol 35:275–283
Cristina MD, Porto PD, Buffolano W, Beghetto E, Spadoni A, Guglietta S, Piccolella E, Felici F, Gargano N (2004) The Toxoplasma gondii bradyzoite antigens bag1 and mag1 induce early humoral and cell-mediated immune responses upon human infection. Microbes Infect 6:164–171
Dubey JP (2008) The history of Toxoplasma gondii–the first 100 years. J Eukaryot Microbiol 55:467–475
Dubey JP (2010) Toxoplasma gondii infections in chickens (gallus domesticus): prevalence, clinical disease, diagnosis and public health significance. Zoonoses Public Health 57:60–73
Fachado A, Rodriguez A, Angel SO, Pinto DC, Vila I, Acosta A, Amendoeira RR, Lannes-Vieira J (2003) Protective effect of a naked DNA vaccine cocktail against lethal toxoplasmosis in mice. Vaccine 21:1327–1335
Ferguson DJP, Parmley SF (2002) Toxoplasma gondii mag1 protein expression. Trends Parasitol 18:482
Gamble HR, Andrews CD, Dubey JP, Webert DW, Parmley SF (2000) Use of recombinant antigens for detection of Toxoplasma gondii infection in swine. J Parasitol 86:459–462
Gong PT, Cao L, Guo YB, Dong H, Yuan SX, Yao XH, Ren WZ, Yao L, Xu ZL, Sun Q, Zhang XC (2016) Toxoplasma gondii: protective immunity induced by a DNA vaccine expressing gra1 and mic3 against toxoplasmosis in balb/c mice. Exp Parasitol 166:131–136
Grzybowski MM, Dziadek B, Gatkowska JM, Dzitko K, Długońska H (2015) Towards vaccine against toxoplasmosis: evaluation of the immunogenic and protective activity of recombinant rop5 and rop18 Toxoplasma gondii proteins. Parasitol Res 114:4553–4563
Ivory C, Chadee K (2004) DNA vaccines: designing strategies against parasitic infections. Genet Vaccines Ther 2:17
Letscher-Bru V, Pfaff AW, Abou-Bacar A, Filisetti D, Antoni E, Villard O, Klein J-P, Candolfi E (2003) Vaccination with Toxoplasma gondii sag-1 protein is protective against congenital toxoplasmosis in balb/c mice but not in cba/j mice. Infect Immun 71:6615–6619
Li YW, Zhou HY (2018) Moving towards improved vaccines for Toxoplasma gondii. Expert Opin Biol Ther 18:273–280
Liu KY, Zhang DB, Wei QK, Li J, Li GP, Yu JZ (2006) Biological role of surface Toxoplasma gondii antigen in development of vaccine. World J Gastroenterol 12:2363–2368
Liu S, Shi L, Cheng YB, Fan GX, Ren HX, Yuan YK (2009) Evaluation of protective effect of multi-epitope DNA vaccine encoding six antigen segments of Toxoplasma gondii in mice. Parasitol Res 105:267–274
Mévélec M, Bout D, Desolme B, Marchand H, Magné R, Bruneel O, Buzoni-Gatel D (2005) Evaluation of protective effect of DNA vaccination with genes encoding antigens gra4 and sag1 associated with gm-csf plasmid, against acute, chronical and congenital toxoplasmosis in mice. Vaccine 23:4489–4499
Milne G, Webster JP, Walker M (2020) Toxoplasma gondii: an underestimated threat? Trends Parasitol 36:959–969
Nakagome K, Okunishi K, Imamura M, Harada H, Matsumoto T, Tanaka R, Miyazaki J-I, Yamamoto K, Dohi M (2009) Ifn-gamma attenuates antigen-induced overall immune response in the airway as a th1-type immune regulatory cytokine. J Immunol 183:209–220
Nan CL, Lei ZL, Zhao ZJ, Shi LH, Ouyang YC, Song XF, Sun QY, Chen DY (2007) Increased th1/th2 (ifn-gamma/il-4) cytokine mrna ratio of rat embryos in the pregnant mouse uterus. J Reprod Dev 53:219–228
Nielsen HV, Lauemøller SL, Christiansen L, Buus S, Foomsgaard A, Petersen E (1999) Complete protection against lethal Toxoplasma gondii infection in mice immunized with a plasmid encoding the sag1 gene. Infect Immun 67:6358–6363
Pappas G, Roussos N, Falagas ME (2009) Toxoplasmosis snapshots: global status of Toxoplasma gondii seroprevalence and implications for pregnancy and congenital toxoplasmosis. Int J Parasitol 39:1385–1394
Parmley S, Slifer T, Araujo F (2002) Protective effects of immunization with a recombinant cyst antigen in mouse models of infection with Toxoplasma gondii tissue cysts. J Infect Dis 185(Suppl 1):S90-95
Rosenberg C, Craeye SD, Jongert E, Gargano N, Beghetto E, Porto PD, Vorup-Jensen T, Petersen E (2009) Induction of partial protection against infection with Toxoplasma gondii genotype ii by DNA vaccination with recombinant chimeric tachyzoite antigens. Vaccine 27:2489–2498
Saadatnia G, Golkar M (2012) A review on human toxoplasmosis. Scand J Infect Dis 44:805–814
Siachoque H, Guzman F, Burgos J, Patarroyo ME, Marin JEG (2006) Toxoplasma gondii: immunogenicity and protection by p30 peptides in a murine model. Exp Parasitol 114:62–65
Tomita T, Mukhopadhyay D, Han B, Yakubu R, Tu V, Mayoral J, Sugi T, Ma Y, Saeij JPJ, Weiss LM (2021) Toxoplasma gondii matrix antigen 1 is a secreted immunomodulatory effector. mBio 12:e00603–e00621
Torres KCL, Dutra WO, Gollob KJ (2004) Endogenous il-4 and ifn-gamma are essential for expression of th2, but not th1 cytokine message during the early differentiation of human cd4+ t helper cells. Hum Immunol 65:1328–1335
Zheng B, Lu S, Tong Q, Kong Q, Lou D (2013) The virulence-related rhoptry protein 5 (ROP5) of Toxoplasma Gondii is a novel vaccine candidate against toxoplasmosis in mice. Vaccine 31:4578–4584
Warner RC, Chapman RC, Davis BN, Davis PH (2021) Review of DNA vaccine approaches against the parasite Toxoplasma gondii. J Parasitol 107:882–903
Funding
This work was supported by grants from the scientific and technological planning project of Jilin province (No. 20200403041SF), National Natural Science Foundation of China (No. 31302076), National Key R&D Program of China (2021YFF0702900), and the Jilin Scientific and Technological Development Program (20210506017ZP).
Author information
Authors and Affiliations
Contributions
LLC designed the research. PTG and XHL acquired the budget. SGC, PPZ, NZ, and XHL conducted the experiments. XZS, NZ, YBG, and NW performed the statistical analysis. LLC, PTG, and SGC drafted the manuscript. BKB and YL revised the English language. PTG revised the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval
All mice were administered an adequate diet and handled in strict accordance with the guidelines of Experimental Animals Center, Jilin University, Changchun, China. All mouse experiments strictly complied with the animal ethics enforcement of the Animal Welfare and Research Ethics Committee of Jilin University (Permit Number: pzpx20190929065).
Consent to participate
This study did not involve human participants; therefore, volunteer consent to participate in this study was not required.
Consent for publication
All authors agreed to the publication of this paper in parasitology research.
Conflict of interests
The authors declare no competing interests.
Additional information
Handling Editor: Xing-Quan Zhu
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Cao, L., Liu, J., Cao, S. et al. Protective efficacy of Toxoplasma gondii bivalent MAG1 and SAG1 DNA vaccine against acute toxoplasmosis in BALB/c mice. Parasitol Res 122, 739–747 (2023). https://doi.org/10.1007/s00436-022-07745-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-022-07745-8