
Abstract
Limited evidence has shown that practising musical activities in aging, such as choral singing, could lessen age-related speech perception in noise (SPiN) difficulties. However, the robustness and underlying mechanism of action of this phenomenon remain unclear. In this study, we used surface-based morphometry combined with a moderated mediation analytic approach to examine whether singing-related plasticity in auditory and dorsal speech stream regions is associated with better SPiN capabilities. 36 choral singers and 36 non-singers aged 20–87 years underwent cognitive, auditory, and SPiN assessments. Our results provide important new insights into experience-dependent plasticity by revealing that, under certain conditions of practice, amateur choral singing is associated with age-dependent structural plasticity within auditory and dorsal speech regions, which is associated with better SPiN performance in aging. Specifically, the conditions of practice that were associated with benefits on SPiN included frequent weekly practice at home, several hours of weekly group singing practice, singing in multiple languages, and having received formal singing training. These results suggest that amateur choral singing is associated with improved SPiN through a dual mechanism involving auditory processing and auditory–motor integration and may be dose dependent, with more intense singing associated with greater benefit. Our results, thus, reveal that the relationship between singing practice and SPiN is complex, and underscore the importance of considering singing practice behaviours in understanding the effects of musical activities on the brain–behaviour relationship.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Speech perception in noise (SPiN) difficulties are common among elderly adults. The causes of these difficulties remain unclear, but a growing body of evidence suggests that brain aging within different functional networks may be the main source of these difficulties. Though brain senescence is unavoidable, the aging brain retains the ability to reorganise itself via a variety of physical, intellectual, artistic, and social activities (for a review, see, e.g. Kramer et al. 2004; Scarmeas and Stern 2003), a phenomenon known as experience-dependent plasticity, which could play a part in mitigating the effects of brain senescence on human cognition and behaviour, including SPiN.
Musical activities are one kind of plasticity-inducing activities that have been studied relatively extensively over the past decade using brain imaging methods. Studies have shown structural differences in several brain networks related to music, cognitive and sensory processing in young musical instrument players (professional and amateur) compared to non-musicians (e.g. Gaser and Schlaug 2003; Bermudez et al. 2009; Schlaug et al. 1995), and in young opera singers compared to non-singers (Kleber et al. 2016). Importantly, a study revealed that structural brain aging, evaluated using the BrainAge framework (Franke et al. 2010), was decelerated for both amateur and professional musical instrument players compared to non-musicians (Rogenmoser et al. 2017). Although the effects of musical activities on SPiN are heterogeneous (for a review, see Coffey et al. 2017), a number of studies have shown that playing a musical instrument (Bidelman and Alain 2015; Fleming et al. 2019; Fostick 2019; Parbery-Clark et al. 2011; Zendel et al. 2019; Zendel and Alain 2012; White-Schwoch et al. 2013; Alain et al. 2014) or singing in a choir (Dubinsky et al. 2019), can mitigate age-related SPiN difficulties. However, the nature of the underlying mechanisms, as well as the type and amount of practice required to positively influence SPiN capabilities, remain unknown.
Though there is no consensus on the mechanisms of action, one hypothesis is that musical activities reduce the impact of aging on the function and structure of auditory and dorsal speech stream regions, which are important networks involved in auditory processing and auditory–motor integration. The dorsal speech stream connects the superior temporal gyrus (STG) directly to the precentral gyrus (PrG) and inferior frontal gyrus (IFG), and indirectly through the inferior parietal lobule. Several neuroimaging studies have linked functional and structural aging within auditory and dorsal speech stream regions to SPiN difficulties (e.g. Du et al. 2016; Wong et al. 2009, 2010; Hwang et al. 2007; Erb and Obleser 2013; Tremblay et al. 2019, 2021; Bilodeau-Mercure et al. 2015; Salvi et al. 2002; Manan et al. 2015, 2017; Sheppard et al. 2011; Perron et al. 2021). For instance, the volume of the left IFG has been found to predict SPiN performance in older but not in younger adults (Wong et al. 2010). Moreover, a decreased activity in the left STG during SPiN has been observed in older compared to younger adults (Wong et al. 2009; Hwang et al. 2007), as well as an increased activity associated with better performance for older adults in the right (Wong et al. 2009) and left PrG (Du et al. 2016). The dorsal speech stream is believed to be involved in the mapping of auditory speech sounds into auditory–motor speech representations (Hickok and Poeppel 2007; Rauschecker and Scott 2009). This process could serve to facilitate disambiguation of incoming speech sounds during SPiN through top-down motor information (for a review, see McGettigan and Tremblay 2018).
Playing a musical instrument and singing also require a functional auditory–motor integration mechanism in which motor commands and auditory feedback are continuously compared to monitor performance and make precise sensorimotor adjustments. Auditory–motor integration involved in musical performance has been shown to recruit a brain network similar to the dorsal speech stream (Zatorre et al. 2007). One study showed that this network is similar for different kinds of musical performance, such as singing and playing a musical instrument (Segado et al. 2018). The auditory–motor integration in the dorsal speech stream is believed to be a shared mechanism between speech, language, singing and music (Loui 2015). The practice of musical activities in general could, therefore, refine SPiN processing through enhanced auditory–motor integration. Consistent with this idea, a recent study has shown that a SPiN advantage for young professional instrumentalists compared to young non-musicians was associated with slightly increased activity in right auditory associative and frontal motor cortices (Du and Zatorre 2017), supporting the notion that musical activities may influence SPiN through their effect on auditory and dorsal speech stream regions. Recently, using the same sample of instrumentalists and non-musicians, Li et al. (2021) showed that young professional instrumentalists exhibited higher microstructural properties in the structure of the bilateral arcuate fasciculus (i.e. higher fractional anisotropy in the right direct branch and lower radial diffusivity in the left anterior branch)—which forms the framework of the dorsal speech stream—that correlated with SPiN performance. Relatedly, a recent study by our group found anatomical differences in the structure of the bilateral arcuate fasciculus, primarily in terms of intermispheric differences, between young and older non-singers and amateur choral singers (Perron et al. 2021). However, these differences were not associated with better SPiN performance for amateur singers. This result was surprising given that we expected singing to have a greater impact on the dorsal speech stream than playing a musical instrument. Indeed, studies have shown that singing has a greater effect on the structure of the arcuate fasciculus than playing an instrument (Halwani et al. 2011), and that speech is more alike to singing than playing an instrument, at the behavioural and neural levels. In particular, Christiner and Reiterer (2013) studied singing ability, musicality, and musical instrument experience in singers with different levels of expertise and showed that singing ability is a better predictor of speech-related skills (i.e. imitation of speech) than musicality and musical instrument experience. The authors proposed that the ability to sing has a greater relationship with speech because these two skills share common basis in terms of vocal and motor production, development, neural orchestration, and auditory cognition, in contrast to playing a musical instrument, which is only close to speech in terms of auditory processing.
While it is possible that singing does not influence SPiN to the same extent as musical instrument playing, it is also possible that the relationship between musical activities and SPiN follows a dose-dependent pattern, with more intense and/or more frequent practice being associated with increased benefits. This is consistent with the finding that professional instrumentalists and amateur singers have a different dorsal speech stream compared to those not practising musical activities, but these differences are associated with SPiN benefits only in professionals, not in amateurs (Du and Zatorre 2017; Perron et al. 2021; Li et al. 2021). Although our participants had, on average, slightly more years of continuous musical experience (M = 17.68 ± 14.14 years, range = [2, 62]) than those in Du's study (M = 16.27 ± 3.77 years, range = [11, 24]), the differences in results could be explained by the difference in musical expertise (amateurs vs. professionals), but also by the fact that our participants were more heterogeneous, i.e. they started singing at different ages and practised for different numbers of hours and at different frequencies per week. In contrast, the professional instrumentalists in Du’s study had a more homogeneous number of years of practice, in addition to having started practising before the age of 7 and reporting practising at least three times a week. The notion of a dose-dependent effect would also be consistent with the literature on music-induced plasticity, which shows that differences in brain anatomy between instrumentalists and non-musicians are related to several practice-related behaviours, such that increased plasticity is related to lower age of onset of musical practice, a higher number of years of practice, a higher number of weekly hours of practice, etc. (Amunts et al. 1997; Schlaug et al. 1995; Steele et al. 2013; for a review, see Merrett et al. 2013). This notion is in line with the Overlap, Precision, Emotion, Repetition, Attention (OPERA) hypothesis (Patel 2011, 2012, 2014), which suggests that musical activities affect speech skills by driving plasticity within speech networks when musical practice is frequent. In our previous study (Perron et al. 2021), although we found that the number of years of continuous singing was associated to a small extent with the structure of the arcuate fasciculus, we did not identify a relationship between the number of years of continuous singing and SPiN. However, several other singing practice behaviours, such as frequency of singing, duration of choral practice, age at onset of singing, etc., could drive plasticity within auditory and dorsal speech stream regions and, in turn, lead to better SPiN performance. Additional studies are needed to understand whether the effect of singing practice on SPiN is dose dependent, and which singing practice behaviours are more important. This information is critical for the development of music-based rehabilitation interventions to maintain SPiN throughout aging.
The overall goal of this study was, therefore, to identify whether specific amateur singing behaviours (e.g. singing frequency) are associated with benefits on SPiN performance, and whether these benefits are related to the structure of auditory and dorsal speech stream regions. The specific aims were threefold: (1) to compare age-related SPiN performance between amateur singers and non-singers, (2) to determine what aspects of a persons’ singing experience is related to SPiN performance, and (3) to determine if the relationship between choral singing and SPiN performance is associated with the cortical thickness, surface, and volume of auditory and dorsal speech stream regions. We expected that, by decomposing the singing practice into different singing behaviours, behavioural differences would be found for singers with a higher level of engagement in singing practice compared to those less engaged. Specifically, we expected frequent singing to be associated with better SPiN performance, which would be related to a less negative age effect on the structure of both auditory and dorsal speech stream regions.
Materials and methods
Participants
Seventy-two healthy native French-speaking adults aged 20–87 years old [M = 54.10 ± 18.42, 41 females (F)] were recruited. All participants were recruited from a companion study (#192–2018) aiming to investigate the impact of choral singing on speech production, articulation, and cognition in aging. Participants were recruited through recruitment advertisements distributed in the community and in ~ 175 choirs in the Quebec City area.
All participants were right-handed according to the Edinburgh Handedness Inventory (Oldfield 1971). Participants had no self-reported history of hearing, language, speech, psychological, neurological, or neurodegenerative disorder at the time of the MRI. Participants were also screened for cognition using the Montreal Cognitive Assessment (MoCA) (Nasreddine et al. 2005). For the MoCA, a less strict cut-off (20/30) than the original cut-off score (26/30) was used to avoid false positives (Waldron-Perrine and Axelrod 2012). Participants were divided into two groups: 36 choral singers (M = 53.19 ± 18.78, range = [20,86], 19F) and 36 non-singers (M = 55.00 ± 18.27, range = [22,87], 22F).
Choral singers were defined as individuals singing in a choir for at least 2 years and with a weekly choral practice of at least 60 consecutive minutes. A choir was defined as any organised group of amateur singers. Nine participants reported that their choir's primary singing style was classical, 9 reported that theirs was popular, 2 reported that theirs was the gospel, and 16 reported that theirs was mixed (i.e. combining several singing styles). Non-singers were defined as individuals not involved in any form of group singing, and not performing professionally or solo singing regularly. None of the participants were professional musicians or regular musical instrument players. Singers and non-singers did not differ in age, education, handedness, number of spoken languages, cognition, and hearing (all p ≥ 0.30). A summary of participants’ information is provided in Table 1. The study was approved by the Comité d’éthique de la recherche sectoriel en neurosciences et santé mentale, Institut Universitaire en Santé Mentale de Québec (#192-2017; #1495-2018). All participants provided informed consent and received a small monetary compensation for their participation. Behavioural and diffusion MRI data collected as part of this project have been published elsewhere (Perron et al. 2021).
Musical activity questionnaire
All participants answered a questionnaire on past and present singing and musical practice behaviours including singing context, formal singing training (yes, no), number of singing languages, age at onset of choral singing, number of years of singing experience, frequency per week of group singing, number of hours of weekly group singing, frequency per week of practice at home and number of hours of weekly practice at home. A singing ratio was calculated (years of singing experience/age). Given the relatively small sample size (N = 36), to examine the effect of singing practice behaviours, most variables were dichotomized (e.g. the practice frequency at home was dichotomized as once a week and more than once a week). A summary of the singing behaviours and distribution of singers in each dichotomous category is provided in Table 2. The most frequently reported singing language was French (94%), followed by English (69%), German (28%), Latin (25%), Spanish (8%), and other languages (Russian, Portuguese, Hebrew and Afrikaans). Those who reported singing in only one language sang either in French or English. Importantly, all participants had little to no musical instrument experience and the two groups were matched in terms of no, present and past musical instrument experience. Participants’ musical instrument experience is detailed elsewhere (Perron et al. 2021).
Experimental design
The experiment consisted of three visits on three separate days. The first and third visit took place at the Speech and Hearing Neuroscience Laboratory at the CERVO Research Centre. All procedures took place in a double-walled soundproof room. Participants completed questionnaires, underwent audiometric evaluations, completed a SPiN task and voice and speech production tasks. These two visits had the duration of 3 h and included several breaks. The second visit was a 1 h MRI session at the IRM Québec-Mailloux Clinic in Quebec City.
Audiometric evaluation
Pure-tone thresholds in dB HL were measured with a calibrated clinical audiometer (AC40, Interacoustic, Danemark). The following frequencies: 0.25, 0.5, 1, 2, 3, 4, 6, 8 kHz were assessed in each ear separately. None of the participants wore hearing aids or cochlear implants and none had been diagnosed with any type of hearing loss. Peripheral hearing was operationalized as an extended pure tone average threshold (PTA) at 0.5, 1, 2, 4 and 6 kHz of the best ear (best ear PTA). According to this measure, nine participants showed signs of mild hearing loss (PTA between 26 and 40 dB) and 1 participant showed signs of moderate hearing loss (PTA between 41 and 60 dB). Among these participants, 6 were non-singers aged between 62 and 86 years and 4 were singers aged between 73 and 80 years. No difference for the extended PTA of each ear and the extended best ear PTA was found between singers and non-singers (Table 1). The extended best ear PTA was included in all statistical analyses as covariate to control for the impact of hearing on SPiN performance.
SPiN task
Participants completed a classic AX discrimination in noise task in a double-walled soundproof room. The standardisation and creation of the stimuli and task are detailed in our previous work (Perron et al. 2021). The task consisted in the discrimination of 300 minimal pairs (150 identical, 150 different) of monosyllabic Quebec-French Consonant–Vowel–Consonant (CVC) syllables created using the lab’s Quebec-French oral language database SyllabO + (Bédard et al. 2017). The syllables were presented using Presentation Software (Neurobehavioural System, USA) through high quality headphones (DT 770 Pro, Beyer Dynamic Inc., Germany). Participants were given a maximum of 3 s to determine if the second syllable was identical or different to the first using a response box (RB-840, Cedrus Corporation, USA). The syllables were presented in the absence of noise or simultaneously with a multi-talker babble noise created by Perrin and Grimault (2005) under two signal-to-noise ratios (SNR; Pressuresignal/Pressurenoise): + 3 dB and − 3 dB. All stimuli and experiment files are available on the Scholar Portal Dataverse: https://doi.org/10.5683/SP2/8IX6QZ.
MRI data acquisition
The data were acquired on an Achieva TX Philips 3.0 Tesla Scanner at the IRM Québec-Mailloux Clinic in Quebec City. Structural MR images were acquired with a T1-weighted 3D-MPRAGE sequence (TR = 8.3 ms, TE = 4.0 ms, FOV = 240 mm, flip angle = 8°, 240 × 240 acquisition matrix, 180 slices/ volume, no gap, voxel size = 1 mm3). Throughout the procedure, the head of each participant was immobilised using a set of cushions. The complete image acquisition protocol had a 45-min duration and also included two BOLD fMRI sequences (speech production task and resting state) and a diffusion MRI sequence. Some of the diffusion MRI data (focusing on the bilateral arcuate fasciculus) has been analysed and published elsewhere (Perron et al. 2021).
MRI data processing
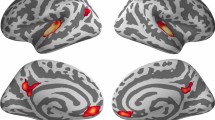
Structural MRI data processing was performed using FreeSurfer software version 6.0.0 (http://freesurfer.net) (Dale et al. 1999; Fischl and Dale 2000; Fischl et al. 1999), including motion correction and conformation, intensity normalisation, non-brain tissue removal using skull stripping algorithms, grey and white matter segmentation and tessellations with automated topology correction. The output of each step was inspected by two of the authors (MP and PT) and manual interventions were performed when required. The native-space surface representations were then parcelled into 74 anatomical regions per hemisphere using the Destrieux 2009 atlas (Destrieux et al. 2010). To address our goal of examining plasticity within auditory and dorsal speech areas, the following ROIs were selected a priori: (1) the pars opercularis of the inferior frontal gyrus (pIFG), (2) the inferior frontal sulcus (IFS), (3) the ventral precentral sulcus (vPrS), (4) the dorsal precentral sulcus (dPrS), (5) the precentral gyrus (PrG), (6) the central sulcus, (7) the supramarginal gyrus (SMG), 8) the angular gyrus (ANG), (9) the planum temporale (PT), (10) the superior temporal sulcus (STS), (11) the transverse temporal gyrus (TTG), 12) the transverse temporal sulcus (TTS), (13) the lateral aspect of the superior temporal gyrus (lSTG) and (14) the planum polare (PP). The structural or functional aging of these regions have been previously associated with aging SPiN (e.g. Du et al. 2016; Wong et al. 2009, 2010; Hwang et al. 2007; Erb and Obleser 2013; Bilodeau-Mercure et al. 2015; Salvi et al. 2002; Manan et al. 2015, 2017; Sheppard et al. 2011; Tremblay et al. 2021). The mean anatomical location of the ROIs is represented in Fig. 1. Because structural and functional differences related to singing and playing an instrument have been observed bilaterally in the dorsal speech stream (Du and Zatorre 2017; Li et al. 2021; Perron et al. 2021), we decided to investigate each of our 14 ROIs bilaterally, resulting in 28 ROIs. Given this large number of ROIs and the fact that we did not expect all ROIs to be associated with singing-related SPiN benefits, we did not include additional, control ROIs.

Location of the regions of interest (ROIs) represented on the group average inflated left surface: (1) the pars opercularis of the inferior frontal gyrus (pIFG), (2) the inferior frontal sulcus (IFS), (3) the ventral precentral sulcus (vPrS), (4) the dorsal precentral sulcus (dPrS), (5) the precentral gyrus (PrG), 6) the central sulcus, (7) the supramarginal gyrus (SMG), (8) the angular gyrus (ANG), (9) the planum temporale (PT), (10) the superior temporal sulcus (STS), (11) the transverse temporal gyrus (TTG), (12) the transverse temporal sulcus (TTS), (13) the lateral aspect of the superior temporal gyrus (lSTG) and (14) the planum polare (PP)
In addition, given that little is known about amateur singing-related structural brain plasticity in aging, we decided to investigate three structural metrics that provide distinct information: ROI cortical thickness (distance between the boundary of grey/white matter division and grey matter/pial surface), surface area (total area of the surface occupied by one brain region) and volume. Thickness and surface measures are influenced by different genetic sources (Panizzon et al. 2009), and they have distinct trajectories of anatomical changes that are influenced by several factors (Raznahan et al. 2011).
Statistical analyses
All data were analysed using SPSS 27 for Mac OS (IBM, New York, USA, www.ibm.com). The percentage of correct responses (accuracy) and reaction times (RT) for correct responses were analysed. For RT, the individual data of each participant were inspected and any trial that deviated by more than two standard deviations from the mean of a condition was removed. The average RT was then calculated with the remaining trials. In addition to accuracy, we also analysed performance within the signal detection theory framework (Macmillan and Creelman 1991). That is, sensitivity (d´) and response bias (c) were calculated for each condition. D´ measures the capacity to correctly recognise whether pairs are the same or different while c reflects the internal decision-making strategy as well as biases. A high value of d’ indicates a good capacity for discrimination. A negative value of c indicates a bias toward responding “identical”, whereas a positive value of c represents a bias towards responding “different”. A value of zero indicates the absence of bias.
Since we previously reported that there was no difference in the effect of age in the two noise conditions (i.e. no interaction between age and condition) as well as no difference in SPiN performance between singers and non-singers in the two noise conditions for d' (Perron et al. 2021), here we averaged the two conditions of noise for each metric (mean accuracy, d´, c, RT). For each dependent variable (mean accuracy, d´, c, RT), normality was assessed via visual inspection of histograms and Q-Q plots and the homogeneity of variance was assessed using Levene’s tests (p ≥ 0.05). Any data points that were more than 1.5 box lengths from the edge of the boxplots (outliers) was removed. No more than 2 participants were excluded per metric (See Supplementary Information 1). The quiet condition was removed because of a strong ceiling effect. For RT, a square root transformation was applied to normalise the distribution.
To address the first aim of this study—to compare age-related SPiN performance between amateur singers and non-singers—we conducted simple moderation analyses using the macro PROCESS (v.3.5) (model #1) for SPSS (Hayes 2017) (Fig. 2a). Moderation analyses were conducted separately for each dependent (Y) variable (mean accuracy, d´, c, RT), with age (mean-centered) as the continuous independent (X) variable and group (non-singers, singers) as the categorical moderator. Peripheral hearing (best ear PTA) was included as covariate.

Conceptual and statistical analytical models for the a moderation and b moderated mediation analyses. The moderation and moderated mediation analyses were conducted separately for each dependent variable (mean accuracy, d´, c, RT) and singing practice behaviours. Different moderated mediation analyses were also conducted for each ROI metric (surface, volume, thickness)
To address the second aim of this study—to determine if different aspects of a persons’ singing experience is related to SPiN performance—we first identified the behavioural measures that were affected by age in the singers by conducting multiple linear regression analyses separately for each dependent variable (mean accuracy, d´, c and RT) with age (mean-centered) and hearing as independent variables. These analyses were performed to select behavioural measures that showed an age effect specifically in singers for inclusion in the moderation analyses. Next, for each dependent (Y) variable that showed an age effect, a series of moderation analyses (model #1) (Fig. 2a) were conducted with age (mean-centered) as the independent (X) variable and each different singing parameter as moderator (M) in separate models. Peripheral hearing (best ear PTA) was included as covariate.
To address the third aim of this study—to determine if the relationship between choral singing and SPiN performance is associated with the cortical thickness, surface, and volume of auditory and dorsal speech stream regions—a moderated mediation framework (model #58) was developed (Fig. 2b), which allows one to establish causality in a statistical sense (e.g. Baron and Kenny 1986). Each behavioural metric was analysed separately. For each behavioural metric, the models included age (mean-centered) as the independent variable (X), singing (group or singing practice behaviours) as the moderator (M) and one ROI metric (surface, volume, or thickness) as mediator (W) per analysis. Peripheral hearing (best ear PTA) was included as a covariate. This moderated mediation model evaluated the direct effect of age on SPiN as well as conditional indirect age effects (ab) (i.e. indirect age effects for each level of a moderator) on SPiN through individual brain structures. The indirect effect of age on SPiN through brain anatomy represents the mechanisms by which age affects SPiN; the analysis determines whether this mechanism varies as function of the moderator (e.g. singing frequency). For accuracy and d’, a positive indirect effect indicates that cases higher on age (i.e. older) are estimated to be better on SPiN through brain anatomy whereas a negative indirect effect indicates that cases higher on age are estimated to be worse on SPiN through brain anatomy (Hayes 2017). For RT, it is the opposite. For c, a significant indirect effect indicates a change in response strategy with age through brain anatomy. A significant moderated mediation is confirmed if at least one pairwise comparison between conditional indirect effects was statistically significant (Hayes 2015). For continuous variables, the levels of moderator were the 16th, 50th and 84th percentiles of the distribution (Hayes 2017). For all analyses, we report the partially standardised regression coefficients (β) (age mean-centered). For all moderated mediations, a bootstrapping approach was used to test for the significance of the effect (p < 0.05), using a bias-corrected bootstrapping with 20 000 samples.
Results
SPiN in aging singers and non-singers
Simple moderation analyses were conducted to compare SPiN performance in singers and non-singers. The descriptive statistics for SPiN performance are provided in Supplementary information 1. Holding hearing constant, the analyses revealed significant age effects on accuracy (β = – 0.154, t = – 3.564, p < 0.001), d´ (β = – 0.014, t = – 3.821, p < 0.001) and RT (β = 0.272, t = 2.470, p = 0.016) (Fig. 3), suggesting a decrease in SPiN performance with age. In addition, hearing was negatively related to accuracy (β = – 0.252, t = – 3.637, p < 0.001) and d´ (β = – 0.018, t = – 3.278, p = 0.002), suggesting lower SPiN performance with higher PTA. There was no group effect and no interactions with group, suggesting no overall behavioural differences between singers and non-singers.

Relationships between age and SPiN performance. The scatter plots show: a accuracy, b sensitivity (d´), and c reaction time. In each graph, the y-axis represents one of the dependent measures of performance with the effect of hearing removed (unstandardized residuals), and the x-axis shows the variable age. Dotted lines represented 95% confidence intervals of the regression line
SPiN in aging singers
Age effects on SPiN for singers were further investigated by conducting multiple linear regressions to select behavioural measures for inclusion in subsequent moderation analyses. The multiple regression models for accuracy (F(2,34) = 53.199, adj. R2 = 0.754, p < 0.001) and d´ (F(2,35) = 53.941, adj. R2 = 0.752, p < 0.001) were statistically significant with a significant contribution of age and hearing. The multiple regression models for c (F(2,33) = 0.098, adj. R2 = – 0.058, p = 0.907) and RT (F(2,33) = 3.280, adj. R2 = 0.121, p = 0.051) were not statistically significant. Based on these results, simple moderation analyses were conducted to investigate whether singing practice behaviours are associated with less negative age effects on accuracy and d’. The results are reported below separately for the age-independent and age-dependent statistical effects.
Age-Independent statistical effects
Holding age and hearing constant, the moderation analyses revealed main effects of the number of singing languages on accuracy (β = 1.032, = 3.337 p = 0.002) and d´ (β = 0.088, t = 3.045, p = 0.005), with increased d’ and accuracy (i.e. better SPiN performance) for singers who sang in different languages (Fig. 4a). The results also showed a main effect of practice frequency at home on accuracy (β = 2.748 t = 2.348, p = 0.026) and d´ (β = 0.228, t = 2.331, p = 0.027) (Fig. 4b), suggesting better accuracy for singers who practised at least once a week at home compared to those who practised less than once a week.

Relationships between age, singing practice behaviours and SPiN performance. The regression plots show measures of sensitivity (d´) or accuracy on the y-axis and age on the x-axis. The graphs were produced using the command line from the output of PROCESS. a Main effect and moderating effect of the number of singing languages on the relationship between age and SPiN performance. b Main effect of practice frequency at home on SPiN performance. c Moderating effect of the number of hours of group singing on the relationship between age and SPiN. d Moderating effect of formal singing training on the relationship between age and SPiN performance. Asterisks indicate significant age effect on SPiN performance at p ≤ 0.05, and ns indicate non-significant age effect on SPiN performance
Age-dependent statistical effects
Holding hearing constant, the number of singing languages positively moderated the relation between age and accuracy (β = 0.065, t = 3.469, p = 0.002) and d´ (β = 0.004, t = 2.173, p = 0.038), revealing a decrease in the negative age effect on SPiN performance with an increase in the number of singing languages (Fig. 4a). Conditional effects revealed a significant age effect on accuracy and d' for singers who sang in one (accuracy: β = – 0.204, t = – 3.351, p = 0.002; d´: β = – 0.018, t = – 3.486, p = 0.002) or two languages (accuracy: β = – 140, t = – 2.412, p = 0.022; d´: β = – 0.015, t = – 2.933, p = 0.006), but no significant age effect for those who sang in four languages (accuracy: β = – 0.010, t = – 0.147, p = 0.885; d´: β = – 0.007, t = – 1.166, p = 0.252). A follow-up analysis revealed no correlation between the number of singing and spoken languages (Pearson correlation: r = 0.063, p = 0.715). The number of hours of group singing also positively moderated the relationship between age and accuracy (β = 0.142, t = 2.042 p = 0.050), revealing no significant negative age effect on accuracy for singers who sang in a choir for at least 3 hours a week (β = – 0.144, t = – 2.012 p = 0.053) compared to those who sang in a choir for less than 3 hours a week (β = – 0.286, t = – 3.225 p = 0.003) (Fig. 4c). Finally, formal singing training positively moderated the relation between age and d’ (β = 0.013, t = 2.139, p = 0.040), revealing no significant negative age effect on d’ for singers with formal singing training (β = – 0.004, t = – 0.554 p = 0.584) compared to those with no formal singing training (β = – 0.017, t = – 3.364 p = 0.002) (Fig. 4d).
SPiN in aging singers and non-singers: practice frequency
As one of our hypotheses was that frequent singing would be associated with mitigation of age-related decline in SPiN performance, and because we observed that higher frequency of practice at home was associated with better SPiN performance, we compared non-singers, singers who frequently practised and singers who practised infrequently using an Analysis of Covariance (ANCOVA). Controlling for hearing and age, significant moderate-sized group effects were found for accuracy (F(2,70) = 3.360, p = 0.041, \(\eta_{{\text{P}}}^{2}\) = 0.094) (Fig. 5a) and d´ (F(2,71) = 3.577, p = 0.034, \(\eta_{{\text{P}}}^{2}\) = 0.098) (Fig. 5b). Pairwise comparisons for accuracy showed that singers who practised frequently performed significantly better than non-singers (mean difference = 2.3%, p = 0.020) and almost significantly better than singers who practised infrequently (mean difference = 2.5%, p = 0.054). Non-singers and singers who practised infrequently did not differ on accuracy (mean difference = 0.2%, p = 0.865). Pairwise comparisons for d’ showed that singers who practised frequently performed significantly better than non-singers (mean difference = 0.188, p = 0.019) and singers who practised infrequently (mean difference = 0.217, p = 0.036). Non-singers and singers who practised infrequently did not differ on d’ (mean difference = 0.029, p = 0.767).

The box plots illustrate the categorical effect of singing on a accuracy and b sensitivity (d´) with the effects of hearing and age removed (residuals), separately for non-singers, singers with infrequent practice at home (< 1/week), and singers with frequent practice at home (≥ 1/week). Each dot represents one participant. Asterisks indicate significance at p ≤ 0.05, and ns indicate non-significance. Error bars represent represented the 95% confidence intervals of the mean
Brain structure and SPiN performance as a function of singing practice behaviours
Given the lack of behavioural differences between the non-singers and singers, we did not investigate the neuroanatomical foundation of singing-related benefits on SPiN. Instead, we focused on the relationship between brain structure and SPiN performance (accuracy, d´) as a function of the singing practice behaviours identified in the previous analysis as being associated with better SPiN performance (i.e. number of singing languages, practice frequency at home, number of hours of group singing, and formal singing training). Moderated mediations were based on behavioural results, meaning that only combinations of singing practice behaviours and behavioural measures (accuracy, d´) that were identified as significant in Sect. 3.2 were included in the analyses. All significant moderated mediations are illustrated in Fig. 6 and the results of all other pathways (including age effects of ROIs and brain differences related to each singing practice behaviour) are summarised in Supplementary Information 2–4.

Illustration of the ROIs showing a significant moderated mediation, presented on group average inflated brain surfaces. a ROIs showing an indirect effect of age on SPiN performance through cortical thickness positively moderated by the number of singing languages. b ROIs showing an indirect effect of age on SPiN performance through cortical thickness positively moderated by the number of hours of group singing. c ROIs showing an indirect effect of age on SPiN performance through cortical thickness positively moderated by formal singing training. In the figure, regions identified in white show a moderated indirect effect of age on accuracy through ROI structure, regions identified in yellow showed a moderated indirect effect of age on sensitivity through ROI structure (d´), and regions identified in blue identifies showed a moderated indirect effect of age for both accuracy and d´
For accuracy, the analyses revealed that the indirect statistical effect of age on SPiN through the thickness of the bilateral lSTG, bilateral PrG, right pIFG, right TTG, right TTS and right PP was positively moderated by the number of singing languages. The analyses also revealed that the indirect effect of age on SPiN through the thickness of the right dPrS was positively moderated by the number of hours of group singing. As can be seen in Supplementary Material 2, the thickness of these ROIs did not differ as a function of the number of singing languages or the number of hours of group singing. In all models, decomposing the main indirect pathway revealed that age had a significant negative effect on the thickness of these ROIs, and that the thickness of these ROIs was significantly (except for the thickness of the right dPrS) and positively associated with accuracy. The positive and significant moderated mediations suggest that the indirect effect was a positive function of the number of singing languages (Table 3a) and the number of hours of group singing (Table 4a), meaning that as the number of singing languages and the number of hours of group singing increased, the age effect on accuracy through the thickness of these ROIs was less negative. No moderated mediation effect was found for practice frequency at home.
For d’, the analyses revealed that the indirect statistical effect of age on SPiN through thickness of the bilateral PrG and lSTG was positively moderated by the number of singing languages. The analyses also revealed that the indirect effect of age on SPiN through thickness of the right pIFG, IFS, PrG and vPrS was positively moderated by formal singing training. As can be seen in Supplementary Material 3, the thickness of these ROIs did not differ as a function of the number of singing languages or formal singing training. In all models, decomposing the main indirect pathway revealed that age had a significant negative effect on the thickness of these ROIs, and that the thickness of these ROIs was positively, but not always significantly, associated with d’. The positive and significant moderated mediations suggest that the indirect effect was a positive function of the number of singing languages (Table 3b) and formal singing training (Table 4b), meaning that as the number of singing language increases and formal singing training is completed, the age effect on d’ through the thickness of these ROIs was less negative. No moderated mediation effect was found for practice frequency at home.
Additional analyses were performed to examine the neurobiological correlates of the age-independent effects associated with practice frequency at home, as these effects could not be captured by our moderated mediation models. Simple mediation analyses (model #4) were performed for each behavioural metric (accuracy, d´), with group (non-singers, singers who frequently practised and singers who practised infrequently) as a multicategorial dependent variable (X), and one ROI metric (surface, volume, or thickness) as mediator (W) per analysis. Peripheral hearing (best ear PTA) and age were included as covariates. The analysis shows that higher practice frequency at home was associated with better SPiN performance, as shown in Sect. 3.3, but these behavioural differences were not mediated by the structure of any ROI.
Discussion
This study is the first to use surface-based morphometry to investigate the relationship between amateur singing, SPiN, and brain structure in aging. Our main hypothesis was that singing would be associated with better SPiN in aging through singing-induced plasticity within auditory and dorsal speech stream regions, but only under certain conditions of practice. Consistent with this hypothesis, we found no overall beneficial relationship between choral singing and SPiN. Yet, exploration of several singing practice behaviours revealed better SPiN performance for singers who frequently practised at home, who received formal singing training, who sang in a choir for at least 3 h per week and for those who sang in multiple languages, which support the hypothesis of a dose-dependent relationship between amateur choral singing and SPiN. These specific singing profiles were associated with a reduced association between age and SPiN through the structure of auditory and dorsal speech stream regions. The behavioural and neurobiological results are discussed in the next sections.
Aging SPiN and singing practice
One of the main findings of this study is that there was no significant overall behavioural difference between singers and non-singers. This finding is at odds with longitudinal and cross-sectional studies showing that playing a musical instrument as an amateur (Fostick 2019; Zendel and Alain 2012; Zendel et al. 2019) or a professional (Parbery-Clark et al. 2011; Bidelman and Alain 2015) have positive effects on SPiN performance in groups of middle-aged to older adults. It is also at odds with one training study that showed that attending 10 weeks of choral singing positively affected SPiN in aging (Dubinsky et al. 2019). It is, however, consistent with a recent study showing no benefits of 12-week choral singing training in middle-aged adults (Hennessy et al. 2021), and with cross-sectional studies observing no difference in SPiN performance between young and old instrumentalists and non-musicians (e.g. Madsen et al. 2019; Boebinger et al. 2015; Fleming et al. 2019). Such heterogeneity could be related to several factors, including personality factors (Corrigall et al. 2013), auditory or genetic predispositions (e.g. Mankel and Bidelman 2018) or methodological differences. Another potentially important explanatory factor is that different kinds of musical activities (e.g. amateur singing, opera singing, playing the violin), or different practice behaviours, may differently affect SPiN performance. The present study supports this notion since benefits in SPiN performance were found only for singers with specific singing profiles. More specifically, our results suggest that singing is associated with better SPiN performance in aging under specific conditions, i.e. singing in a choir for more than 3 h per week, practising frequently at home, obtaining formal singing training, and singing in several languages, which supports the notion of a dose–response relationship.
Furthermore, we found that the different singing practice behaviours did not have the same effects on accuracy and d'. While we found that practice frequency at home and number of singing languages were positively associated with both measures, the number of hours of group singing and formal singing training only had a positive association with accuracy and d’, respectively. Whereas accuracy is a global measure of performance that reflects both discrimination capacities and decision-making, d’ is a more specific measure describing the ability to discriminate between identical and different syllable pairs. One hypothesis for this difference is that formal singing training has a stronger and more durable impact on the speech motor system than increasing the duration of choral singing sessions, and could, therefore, further reinforce speech motor representations, which would in turn improve the ability to discriminate speech. These results suggest that practice frequency is a more important factor than practice duration, which is consistent with prior studies. Notably, Molloy et al. (2012) showed that long training sessions in auditory frequency discrimination reduced learning speed and long-term improvements compared to short or repeated training sessions. Similarly, Wright and Sabin (2007) examined how changing the amount of daily training influenced learning over multiple days in two auditory discrimination tasks. They showed that improvement in auditory discrimination over time may require a specific amount of daily training, and that training above this threshold has no effect on learning. The frequency of practice therefore appears to be an important factor to improve SPiN, which is consistent with the OPERA hypothesis which stipulates that repetition is essential to trigger lasting neuroplasticity (Patel 2011, 2012, 2014).
Although our results support the notion of a dose-dependent relationship between singing and SPiN performance, we cannot totally discard a potential effect of motivation. That is, singers with frequent practice may have better SPiN performance not because they frequently sing, but because they are more motivated to sing than singers with infrequent practice, and thus more engaged in their practice, leading to SPiN benefits (McAuley et al. 2011). Further studies are needed to investigate the role of motivation on music-related benefits.
Two previous (independent) studies examined the effect of singing training on SPiN. Dubinsky et al. (2019) showed SPiN benefits after 10 weeks of choral training including 2 h of choral-group session and up to 1 h of individual musical and vocal exercises per week in middle-aged to older adults. More recently, using a similar choral training design, but over a 12-week period, Hennessy et al. (2021) showed no SPiN benefits related to training in middle-aged adults with subjective hearing loss. The authors suggested that perhaps their SPiN task was too easy and not sensitive enough to detect group differences. Other explanations could be the difference in participants’ age in the two studies (i.e. middle-aged versus middle-aged to older adults) and the presence of self-reported hearing impairment in Hennessy et al. (2021). The present study, singers who practised regularly are comparable to the participants in these two studies (i.e. at least 2 h of choir per week and a weekly practice at home), and those had better SPiN performance than non-singers and singers with infrequent practice. Another study has shown that the number of hours of musical instrument practice per week positively correlates with SPiN performance in older amateur instrumentalists (Zendel and Alain 2012). Future studies are needed to determine under what conditions singing can improve SPiN, and whether the effects of intensive singing training or amateur singing experience are maintained over time even when practice is stopped, or whether continued practice is necessary to maintain benefits on SPiN.
In addition to the effect of practice frequency, we also found a SPiN advantage for aging singers who sang in multiple languages. The number of singing languages was not correlated with the number of spoken languages, suggesting that singers sang without necessarily understanding the words they produced. This suggests that the SPiN advantage of singing in various languages is associated with auditory–motor learning related to bilingualism. This hypothesis is discussed in the following section.
Brain structure and SPiN performance as a function of singing practice behaviours
Our results show that specific singing practice behaviours were associated with a less negative age effect on SPiN through the structure of auditory cortical and dorsal speech stream regions of both hemispheres, but especially the right. Specifically, we found that singing in multiple languages was associated with a less negative age effect on SPiN through the structure of the right TTS/TTG (i.e. primary auditory area), right PP and bilateral lSTG. These mostly right lateralized auditory regions have been shown to participate in the spectro-temporal analysis of all sounds with a specialisation for music features (i.e. pitch, melody, rhythm) (e.g. Liégeois-Chauvel et al. 1998; Penhune et al. 1999; Samson and Zatorre 1994). Bilingual experience has been linked to differences in the structure of brain regions supporting acoustic and auditory processing (Ressel et al. 2012; Martensson et al. 2012; Wong et al. 2008; Golestani et al. 2007). For instance, Ressel et al. (2012) found larger volume in the bilateral TTG/TTS in bilingual compared to monolingual speakers and Martensson et al. (2012) found an increase in thickness of the left STG after three months of intensive foreign language learning compared to a control group. Singing, especially in multiple languages, is a linguistic experience relying heavily on auditory processing, which could lead to a more efficient or more resilient auditory network through structural plasticity.
A few prior studies on the musician advantage on SPiN in aging have also shown that musical training is associated with an improvement in auditory processing for SPiN. For singers, Dubinsky et al. (2019) observed an increase in frequency following response (FFR) in the auditory brainstem in older adults after 10 weeks of choral training compared to a control group, which predicted improvements in pitch discrimination and, in turn, SPiN advantage. For instrumentalists, it has been shown that older adults with past musical training do not exhibit neural timing delays in the auditory brainstem compared to older adults with no past musical training in response to consonant–vowel transitions (i.e. between /d/ and /a/ in /da/) in noise and in quiet (White-Schwoch et al. 2013). Bidelman and Alain (2015) have shown that older instrumentalists are faster at categorising vowels, which was associated with higher neural encoding of speech at auditory cortical level compared to non-musicians. In sum, singing and musical instrument practice could be associated with improved auditory processing related to structural and functional plasticity in primary and secondary auditory areas, which in turn would be associated with SPiN benefits. However, further studies are needed to compare the relationship between different musical activities and auditory processing during SPiN to determine if different activities (singing vs. playing a musical instrument) show some specificity.
In this study, we also found that three factors (singing in multiple languages; having received formal singing training and singing in a choir for at least 3 h weekly) were associated with a less negative relationship between age and SPiN performance through the structure of the bilateral dorsal speech stream regions. The dorsal speech stream is thought to play a role in auditory–motor integration by connecting temporal and frontal cortices (Hickok and Poeppel 2007; Rauschecker and Scott 2009). Our results are consistent with the finding that young adults with a professional musical training perform better during a SPiN task than non-musicians, and that this advantage was associated with upregulation in the right dorsal speech stream (Du and Zatorre 2017) and higher microstructural quality of the bilateral arcuate fasciculus (Li et al. 2021).
Singing is a musical activity characterised by precise sensory–motor adjustments to produce specific sounds, which requires auditory–motor integration. The maintenance of auditory–motor integration could represent the mechanism through which singing is associated with mitigated age-related SPiN decline. The notion of a relationship between speech perception and speech production is long-standing (for a review, see McGettigan and Tremblay 2018). For example, it has been shown that speech sounds classification can be modulated by speech motor learning (Lametti et al. 2014) and speech adaptation (Grabski et al. 2013). Consistent with the notion of a benefit through auditory–motor integration, we found that singing in multiple languages was associated with better SPiN performance in aging through the structure of the left PrG (corresponding to the primary and premotor cortices). According to the Directions Into Velocities of Articulators (DIVA) model of speech production (Guenther 1994, 1995; Guenther et al. 2006), the left PrG contains speech motor representations. During SPiN, disambiguation of incoming speech sounds could be facilitated through top-down motor information (for a review, see McGettigan and Tremblay 2018). In this study, our results suggest that singing in multiple languages, which involves auditory–motor learning, could strengthen auditory–motor integration, and therefore allow more efficient top-down information during SPiN. In addition to a relationship with the structure of the left PrG, we also found that singing in different languages and attending formal singing training is associated with better SPiN performance in aging through the structure of the right PrG. A role for the right PrG in SPiN via intracortical interactions with the left motor cortex has been shown using dual-coil transcranial magnetic stimulation (Nuttall et al. 2018). In addition, increased activation in the right PrG during SPiN has also been associated with better performance in aging (Wong et al. 2009), suggesting a compensation mechanism. In summary, our results suggest that specific singing practice behaviours are associated with the maintenance of auditory–motor integration through structural plasticity in bilateral auditory and dorsal speech stream regions. Specifically, singing in multiple languages was associated with SPiN benefits through the structure of bilateral auditory and dorsal speech stream regions, particularly bilateral lSTG and bilateral PrG for both accuracy and d'. This suggests that singing in multiple languages may affect an auditory–motor integration mechanism that occurs bilaterally. In contrast, the other singing behaviours were associated with age-related SPiN benefits only through the right dorsal speech stream. The role of the right hemisphere in SPiN is discussed in the following section.
Relationship between singing and SPiN through the right hemisphere
An interesting finding of the present study is that the relationship between singing practice behaviours and SPiN appears to be primarily associated with auditory and dorsal speech stream regions in the right hemisphere. For auditory regions, this result is not surprising. Indeed, studies have shown that the processing of continuous and complex auditory signals—whether speech or non-speech—involves bilateral supratemporal plane regions (Deschamps et al. 2016; Tremblay et al. 2013). Here, the greater proportion of right vs. left hemisphere auditory regions could be explained by the presence of noise. Indeed, several studies have shown that right auditory regions are important for processing noise. Notably, Shtyrov et al. (1998) found that, in a noisy environment, the mismatch negativity elicited by auditory consonant–vowel syllables in the left auditory cortex decreases while that of the right hemisphere increases, suggesting that noisy signals may be processed in the right hemisphere. Relatedly, Santosa et al. (2014) used functional near-infrared spectroscopy (fNIRS) to study hemodynamic response in the bilateral auditory cortex in response to music, noise and a mixture of noise and music. They found that music and noise processing were associated with bilateral activation in most participants. In contrast, the processing of music and noise simultaneously was right lateralized. Taken together, these results highlight the importance of the right hemisphere in processing noise, including speech in noise.
For the dorsal speech stream, the finding that the relationship between singing practice behaviours and SPiN is primarily associated with right hemisphere regions was not expected, given that the dorsal speech stream is thought to be left lateralized (Rauschecker and Scott 2009; Hickok and Poeppel 2007). Yet, most aging studies have shown that age-related structural and functional decline of the bilateral dorsal speech stream is associated with SPiN difficulties (e.g. Du et al. 2016; Wong et al. 2010, 2009; Hwang et al. 2007; Erb and Obleser 2013; Tremblay et al. 2019, 2021; Bilodeau-Mercure et al. 2015; Salvi et al. 2002; Manan et al. 2015, 2017; Sheppard et al. 2011; Perron et al. 2021). The role of the right dorsal speech stream in SPiN is largely unknown. Interestingly, in explaining their findings of a beneficial effect of professional instrument playing on SPiN through the right dorsal speech stream, Li et al. (2021) suggested that the right dorsal stream could be a functional extension of the left dorsal speech stream for segregating speech sounds from noisy background. Overall, the literature suggests that both hemispheres are involved in speech processing, but the right hemisphere may be particularly important for processing noisy speech signals. Finally, it is well known that music processing is relatively right lateralized (e.g. Tervaniemi and Hugdahl 2003). It is, therefore, likely that engaging in a musical activity promotes brain plasticity in the right hemisphere.
One idea to explore in future studies is that, because singing is more closely related to speech than playing a musical instrument (e.g. Christiner and Reiterer 2013), singing could result in greater plasticity in the left speech network compared to playing a musical instrument, and could be associated with higher gains in SPiN through its effects on speech processing. This would be consistent with our results showing that most aspects of singing practice are positively associated with SPiN performance via the right hemisphere, but that aspects of singing that require language (i.e. singing in multiple languages) are positively associated with SPiN performance via both hemispheres.
Relationship between singing and SPiN through cortical thickness
Our analyses revealed that that SPiN benefits in aging were associated with cortical thickness, but not with cortical volume or surface. Cortical volume is usually interpreted as the product of cortical thickness and surface. According to the radial unit hypothesis (Rakic 1988), cortical surface and thickness represent distinct morphometric characteristics. While surface is determined by the number of vertical ontogenetic columns, thickness is determined by the number and size of cells in a column. Animal studies have shown that it is possible to modify the development of cortical thickness with no change to cortical surface by genetic mutations affecting the intermediate progenitor cells involved in neurogenesis (for a review, see Pontious et al. 2008), suggesting that these two cortical measures are regulated by distinct mechanisms. In line with these previous results, Panizzon et al. (2009) have found that cortical thickness and surface are influenced by different genetic sources.
The current results suggest that singing-related benefits are associated with a specific mechanism primarily related to cortical thickness. One hypothesis is that these results could reflect a mechanism of genesis of neurons within the ontogenetic columns (Rakic 1988). This would be consistent with evidence showing that neurogenesis is possible in few regions of the adult human brain, including the hippocampus (e.g. Eriksson et al. 1998). Further, animal studies have shown that neurogenesis can occur in the grey matter of the cerebral cortex (Dayer et al. 2005; Gould et al. 1999). Notably, Dayer et al. (2005) found oligodendrocyte precursor cells in the neocortex of adult rats that seemed to generate new neurons. Although we identified some ROI structural differences related to singing practice behaviours (see Supplementary Materials 2 and 3), age-related SPiN benefits were not associated with a significantly larger or smaller ROI thickness. Instead, age-related benefits were associated with a reduced impact of age on the relationship between SPiN performance and cortical thickness, which may reflect a mechanism of preservation or modification of neuronal morphology in aging.
Limitations
The main limitation of this study is the relatively small sample sizes (N = 36 per group). A consequence of the small sample is that we had to dichotomize variables to describe singing practice behaviours (e.g. formal training vs. no training, as opposed to using a variable measuring more precisely the amount of training).
Conclusion
Choral singing is a musical activity that is demanding on multiple levels: sensorimotor, cognitive, and emotional. Because of this, choral singing has the potential to promote brain plasticity. Here, we investigated whether choral singing is associated with better SPiN performance in aging through experience-dependent structural plasticity within auditory and dorsal speech stream regions. Our results suggest that choral singing is associated with SPiN benefits in aging, that this relationship is a dose dependent, and may reflect differences in auditory processing and auditory–motor integration. These findings highlight the importance of considering the characteristics of singers when investigating singing-induced plasticity. Understanding the singing behaviours that most influence neuroplasticity and SPiN is necessary to develop singing-based prevention strategies for communication-mediated activities throughout the entire lifespan.
Availability of data and material
All stimuli and experiment files are publicly available on the Scholar portal Dataverse: https://doi.org/10.5683/SP2/8IX6QZ. Individual data are not available because informed consent obtained from participants did not include consent to public data sharing, and we were not granted the right to request it a posteriori by our local research ethics committee.
Code availability
The software used in this study is freely available online: http://freesurfer.net.
References
Alain C, Zendel BR, Hutka S, Bidelman GM (2014) Turning down the noise: the benefit of musical training on the aging auditory brain. Hear Res 308:162–173. https://doi.org/10.1016/j.heares.2013.06.008
Amunts K, Schlaug G, Jancke L, Steinmetz H, Schleicher A, Dabringhaus A, Zilles K (1997) Motor cortex and hand motor skills: structural compliance in the human brain. Hum Brain Mapp 5(3):206–215. https://doi.org/10.1002/(SICI)1097-0193(1997)5:3%3c206::AID-HBM5%3e3.0.CO;2-7
Baron RM, Kenny DA (1986) The moderator-mediator variable distinction in social psychological research: conceptual, strategic, and statistical considerations. J Pers Soc Psychol 51(6):1173–1182
Bédard P, Audet AM, Drouin P, Roy J-P, Julie R, Tremblay P (2017) SyllabO+: a new tool to study sublexical phenomena in spoken Quebec French. Behav Res 49:1852–1863. https://doi.org/10.3758/s13428-016-0829-7
Bermudez P, Lerch JP, Evans AC, Zatorre RJ (2009) Neuroanatomical correlates of musicianship as revealed by cortical thickness and voxel-based morphometry. Cereb Cortex 19(7):1583–1596. https://doi.org/10.1093/cercor/bhn196
Bidelman GM, Alain C (2015) Musical training orchestrates coordinated neuroplasticity in auditory brainstem and cortex to counteract age-related declines in categorical vowel perception. J Neurosci 35(3):1240–1249. https://doi.org/10.1523/JNEUROSCI.3292-14.2015
Bilodeau-Mercure M, Lortie CL, Sato M, Guitton MJ, Tremblay P (2015) The neurobiology of speech perception decline in aging. Brain Struct Funct 220(2):979–997. https://doi.org/10.1007/s00429-013-0695-3
Boebinger D, Evans S, Rosen S, Lima CF, Manly T, Scott SK (2015) Musicians and non-musicians are equally adept at perceiving masked speech. J Acoust Soc Am 137(1):378–387. https://doi.org/10.1121/1.4904537
Christiner M, Reiterer S (2013) Song and speech: examining the link between singing talent and speech imitation ability. Front Psychol. https://doi.org/10.3389/fpsyg.2013.00874
Coffey EBJ, Mogilever NB, Zatorre RJ (2017) Speech-in-noise perception in musicians: a review. Hear Res 352:49–69. https://doi.org/10.1016/j.heares.2017.02.006
Corrigall KA, Schellenberg EG, Misura NM (2013) Music training, cognition, and personality. Front Psychol 4:222. https://doi.org/10.3389/fpsyg.2013.00222
Dale AM, Fischl B, Sereno MI (1999) Cortical surface-based analysis I. Segmentation and Surface Reconstruction. Neuroimage 9(2):179–194. https://doi.org/10.1006/nimg.1998.0395
Dayer AG, Cleaver KM, Abouantoun T, Cameron HA (2005) New GABAergic interneurons in the adult neocortex and striatum are generated from different precursors. J Cell Biol 168(3):415–427. https://doi.org/10.1083/jcb.200407053
Deschamps I, Hasson U, Tremblay P (2016) The structural correlates of statistical information processing during speech perception. PLoS ONE 11(2):e0149375–e0149375. https://doi.org/10.1371/journal.pone.0149375
Destrieux C, Fischl B, Dale A, Halgren E (2010) Automatic parcellation of human cortical gyri and sulci using standard anatomical nomenclature. Neuroimage 53(1):1–15. https://doi.org/10.1016/j.neuroimage.2010.06.010
Du Y, Zatorre RJ (2017) Musical training sharpens and bonds ears and tongue to hear speech better. Proc Natl Acad Sci USA 114(51):13579–13584. https://doi.org/10.1073/pnas.1712223114
Du Y, Buchsbaum BR, Grady CL, Alain C (2016) Increased activity in frontal motor cortex compensates impaired speech perception in older adults. Nat Commun 7:12241. https://doi.org/10.1038/ncomms12241
Dubinsky E, Wood EA, Nespoli G, Russo FA (2019) Short-term choir singing supports speech-in-noise perception and neural pitch strength in older adults with age-related hearing loss. Front Neurosci 13:1153. https://doi.org/10.3389/fnins.2019.01153
Erb J, Obleser J (2013) Upregulation of cognitive control networks in older adults’ speech comprehension. Front Syst Neurosci 7:116. https://doi.org/10.3389/fnsys.2013.00116
Eriksson PS, Perfilieva E, Björk-Eriksson T, Alborn AM, Nordborg C, Peterson DA, Gage FH (1998) Neurogenesis in the adult human hippocampus. Nat Med 4(11):1313–1317. https://doi.org/10.1038/3305
Fischl B, Dale AM (2000) Measuring the thickness of the human cerebral cortex from magnetic resonance images. Proc Natl Acad Sci USA 97(20):11050–11055. https://doi.org/10.1073/pnas.200033797
Fischl B, Sereno MI, Dale AM (1999) Cortical surface-based analysis. II: inflation, flattening, and a surface-based coordinate system. Neuroimage 9(2):195–207. https://doi.org/10.1006/nimg.1998.0396
Fleming D, Belleville S, Peretz I, West G, Zendel BR (2019) The effects of short-term musical training on the neural processing of speech-in-noise in older adults. Brain Cogn 136:103592. https://doi.org/10.1016/j.bandc.2019.103592
Fostick L (2019) Card playing enhances speech perception among aging adults: comparison with aging musicians. Eur J Ageing 16(4):481–489. https://doi.org/10.1007/s10433-019-00512-2
Franke K, Ziegler G, Kloppel S, Gaser C, Alzheimer’s Disease Neuroimaging I (2010) Estimating the age of healthy subjects from T1-weighted MRI scans using kernel methods: exploring the influence of various parameters. Neuroimage 50(3):883–892. https://doi.org/10.1016/j.neuroimage.2010.01.005
Gaser C, Schlaug G (2003) Brain structures differ between musicians and non-musicians. J Neurosci 23(27):9240–9245
Golestani N, Molko N, Dehaene S, LeBihan D, Pallier C (2007) Brain structure predicts the learning of foreign speech sounds. Cereb Cortex 17(3):575–582. https://doi.org/10.1093/cercor/bhk001
Gould E, Reeves AJ, Graziano MS, Gross CG (1999) Neurogenesis in the neocortex of adult primates. Science 286(5439):548–552. https://doi.org/10.1126/science.286.5439.548
Grabski K, Tremblay P, Gracco VL, Girin L, Sato M (2013) A mediating role of the auditory dorsal pathway in selective adaptation to speech: a state-dependent transcranial magnetic stimulation study. Brain Res 1515:55–65. https://doi.org/10.1016/j.brainres.2013.03.024
Guenther FH (1994) A neural network model of speech acquisition and motor equivalent speech production. Biol Cybern 72(1):43–53
Guenther FH (1995) Speech sound acquisition, coarticulation, and rate effects in a neural network model of speech production. Psychol Rev 102(3):594–621. https://doi.org/10.1037/0033-295x.102.3.594
Guenther FH, Ghosh SS, Tourville JA (2006) Neural modeling and imaging of the cortical interactions underlying syllable production. Brain Lang 96(3):280–301. https://doi.org/10.1016/j.bandl.2005.06.001
Halwani GF, Loui P, Ruber T, Schlaug G (2011) Effects of practice and experience on the arcuate fasciculus: comparing singers, instrumentalists, and non-musicians. Front Psychol 2:156. https://doi.org/10.3389/fpsyg.2011.00156
Hayes AF (2015) An index and test of linear moderated mediation. Multivari Behav Res 50(1):1–22. https://doi.org/10.1080/00273171.2014.962683
Hayes AF (2017) Introduction to Mediation, Moderation, and Conditional Process Analysis, 2 A Regression-Based Approach. Guilford Publications
Hennessy S, Wood A, Wilcox R, Habibi A (2021) Neurophysiological improvements in speech-in-noise task after short-term choir training in older adults. Aging (albany NY) 13(7):9468–9495. https://doi.org/10.18632/aging.202931
Hickok G, Poeppel D (2007) The cortical organization of speech processing. Nat Rev Neurosci 8(5):393–402. https://doi.org/10.1038/nrn2113
Hwang JH, Li CW, Wu CW, Chen JH, Liu TC (2007) Aging effects on the activation of the auditory cortex during binaural speech listening in white noise: an fMRI study. Audiol Neurootol 12(5):285–294. https://doi.org/10.1159/000103209
Kleber B, Veit R, Moll CV, Gaser C, Birbaumer N, Lotze M (2016) Voxel-based morphometry in opera singers: Increased gray-matter volume in right somatosensory and auditory cortices. Neuroimage 133:477–483. https://doi.org/10.1016/j.neuroimage.2016.03.045
Kramer AF, Bherer L, Colcombe SJ, Dong W, Greenough WT (2004) Environmental influences on cognitive and brain plasticity during aging. J Gerontol A Biol Sci Med Sci 59(9):M940-957. https://doi.org/10.1093/gerona/59.9.m940
Lametti DR, Rochet-Capellan A, Neufeld E, Shiller DM, Ostry DJ (2014) Plasticity in the human speech motor system drives changes in speech perception. J Neurosci 34(31):10339–10346. https://doi.org/10.1523/JNEUROSCI.0108-14.2014
Li X, Zatorre R, Du Y (2021) The microstructural plasticity of the arcuate fasciculus undergirds improved speech in noise perception in musicians. Cereb Cortex. https://doi.org/10.1093/cercor/bhab063
Liégeois-Chauvel C, Peretz I, Babaï M, Laguitton V, Chauvel P (1998) Contribution of different cortical areas in the temporal lobes to music processing. Brain 121(Pt 10):1853–1867. https://doi.org/10.1093/brain/121.10.1853
Loui P (2015) A dual-stream neuroanatomy of singing. Music Percept 32(3):232–241. https://doi.org/10.1525/mp.2015.32.3.232
Macmillan NA, Creelman CD (1991) Detection theory: a user’s guide. Cambridge University Press, New York
Madsen SMK, Marschall M, Dau T, Oxenham AJ (2019) Speech perception is similar for musicians and non-musicians across a wide range of conditions. Sci Rep 9(1):10404. https://doi.org/10.1038/s41598-019-46728-1
Manan HA, Franz EA, Yusoff AN, Mukari SZ (2015) The effects of aging on the brain activation pattern during a speech perception task: an fMRI study. Aging Clin Exp Res 27(1):27–36. https://doi.org/10.1007/s40520-014-0240-0
Manan HA, Yusoff AN, Franz EA, Mukari SZMS (2017) Effects of aging and background babble noise on speech perception processing: an fMRI Study. Neurophysiology 49(6):441–452. https://doi.org/10.1007/s11062-018-9707-5
Mankel K, Bidelman GM (2018) Inherent auditory skills rather than formal music training shape the neural encoding of speech. Proc Natl Acad Sci USA 115(51):13129–13134. https://doi.org/10.1073/pnas.1811793115
Martensson J, Eriksson J, Bodammer NC, Lindgren M, Johansson M, Nyberg L, Lovden M (2012) Growth of language-related brain areas after foreign language learning. Neuroimage 63(1):240–244. https://doi.org/10.1016/j.neuroimage.2012.06.043
McAuley J, Henry M, Tuft S (2011) Musician advantages in music perception: an issue of motivation, not just ability. Music Percept 28:505–518. https://doi.org/10.1525/mp.2011.28.5.505
McGettigan C, Tremblay P (2018) Links Between Perception and ProductionExamining the roles of motor and premotor cortices in understanding speech. In: Rueschemeyer S-A, Gaskell MG (eds) Oxford University Press. https://doi.org/10.1093/oxfordhb/9780198786825.013.14
Merrett DL, Peretz I, Wilson SJ (2013) Moderating variables of music training-induced neuroplasticity: a review and discussion. Front Psychol 4:606. https://doi.org/10.3389/fpsyg.2013.00606
Molloy K, Moore DR, Sohoglu E, Amitay S (2012) Less is more: latent learning is maximized by shorter training sessions in auditory perceptual learning. PLoS ONE 7(5):e36929–e36929. https://doi.org/10.1371/journal.pone.0036929
Nasreddine ZS, Phillips NA, Bedirian V, Charbonneau S, Whitehead V, Collin I, Cummings JL, Chertkow H (2005) The Montreal Cognitive Assessment, MoCA: a brief screening tool for mild cognitive impairment. J Am Geriatr Soc 53(4):695–699. https://doi.org/10.1111/j.1532-5415.2005.53221.x
Nuttall HE, Kennedy-Higgins D, Devlin JT, Adank P (2018) Modulation of intra- and inter-hemispheric connectivity between primary and premotor cortex during speech perception. Brain Lang 187:74–82. https://doi.org/10.1016/j.bandl.2017.12.002
Oldfield RC (1971) The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia 9(1):97–113. https://doi.org/10.1016/0028-3932(71)90067-4
Panizzon MS, Fennema-Notestine C, Eyler LT, Jernigan TL, Prom-Wormley E, Neale M, Jacobson K, Lyons MJ, Grant MD, Franz CE, Xian H, Tsuang M, Fischl B, Seidman L, Dale A, Kremen WS (2009) Distinct genetic influences on cortical surface area and cortical thickness. Cereb Cortex 19(11):2728–2735. https://doi.org/10.1093/cercor/bhp026
Parbery-Clark A, Strait DL, Anderson S, Hittner E, Kraus N (2011) Musical experience and the aging auditory system: implications for cognitive abilities and hearing speech in noise. PLoS ONE 6(5):e18082. https://doi.org/10.1371/journal.pone.0018082
Patel AD (2011) Why would Musical Training Benefit the Neural Encoding of Speech? The OPERA Hypothesis Front Psychol 2:142. https://doi.org/10.3389/fpsyg.2011.00142
Patel AD (2012) The OPERA hypothesis: assumptions and clarifications. Ann N Y Acad Sci 1252:124–128. https://doi.org/10.1111/j.1749-6632.2011.06426.x
Patel AD (2014) Can nonlinguistic musical training change the way the brain processes speech? The expanded OPERA hypothesis. Hear Res 308:98–108. https://doi.org/10.1016/j.heares.2013.08.011
Penhune VB, Zatorre RJ, Feindel WH (1999) The role of auditory cortex in retention of rhythmic patterns as studied in patients with temporal lobe removals including Heschl’s gyrus. Neuropsychologia 37(3):315–331
Perrin F, Grimault N (2005) Fonds sonores. Laboratoire Unités Mixtes de Recherche, Centre National de la Recherche Scientifique 5020, Lyon, France
Perron M, Theaud G, Descoteaux M, Tremblay P (2021) The frontotemporal organization of the arcuate fasciculus and its relationship with speech perception in young and older amateur singers and non-singers. Hum Brain Mapp. https://doi.org/10.1002/hbm.25416
Pontious A, Kowalczyk T, Englund C, Hevner RF (2008) Role of intermediate progenitor cells in cerebral cortex development. Dev Neurosci 30(1–3):24–32. https://doi.org/10.1159/000109848
Rakic P (1988) Specification of cerebral cortical areas. Science 241(4862):170–176
Rauschecker JP, Scott SK (2009) Maps and streams in the auditory cortex: nonhuman primates illuminate human speech processing. Nat Neurosci 12(6):718–724. https://doi.org/10.1038/nn.2331
Raznahan A, Shaw P, Lalonde F, Stockman M, Wallace GL, Greenstein D, Clasen L, Gogtay N, Giedd JN (2011) How does your cortex grow? J Neurosci 31(19):7174–7177. https://doi.org/10.1523/JNEUROSCI.0054-11.2011
Ressel V, Pallier C, Ventura-Campos N, Diaz B, Roessler A, Avila C, Sebastian-Galles N (2012) An effect of bilingualism on the auditory cortex. J Neurosci 32(47):16597–16601. https://doi.org/10.1523/JNEUROSCI.1996-12.2012
Rogenmoser L, Kernbach J, Schlaug G, Gaser C (2017) Keeping brains young with making music. Brain Struct Funct. https://doi.org/10.1007/s00429-017-1491-2
Salvi RJ, Lockwood AH, Frisina RD, Coad ML, Wack DS, Frisina DR (2002) PET imaging of the normal human auditory system: responses to speech in quiet and in background noise. Heart Res 170(1–2):96–106
Samson S, Zatorre RJ (1994) Contribution of the right temporal lobe to musical timbre discrimination. Neuropsychologia 32(2):231–240. https://doi.org/10.1016/0028-3932(94)90008-6
Santosa H, Hong MJ, Hong KS (2014) Lateralization of music processing with noises in the auditory cortex: an fNIRS study. Front Behav Neurosci 8:418. https://doi.org/10.3389/fnbeh.2014.00418
Scarmeas N, Stern Y (2003) Cognitive reserve and lifestyle. J Clin Exp Neuropsychol 25(5):625–633. https://doi.org/10.1076/jcen.25.5.625.14576
Schlaug G, Jancke L, Huang Y, Staiger JF, Steinmetz H (1995) Increased corpus callosum size in musicians. Neuropsychologia 33(8):1047–1055. https://doi.org/10.1016/0028-3932(95)00045-5
Segado M, Hollinger A, Thibodeau J, Penhune V, Zatorre RJ (2018) Partially overlapping brain networks for singing and cello playing. Front Neurosci 12:351. https://doi.org/10.3389/fnins.2018.00351
Sheppard JP, Wang JP, Wong PC (2011) Large-scale cortical functional organization and speech perception across the lifespan. PLoS ONE 6(1):e16510. https://doi.org/10.1371/journal.pone.0016510
Shtyrov Y, Kujala T, Ahveninen J, Tervaniemi M, Alku P, Ilmoniemi RJ, Näätänen R (1998) Background acoustic noise and the hemispheric lateralization of speech processing in the human brain: magnetic mismatch negativity study. Neurosci Lett 251(2):141–144. https://doi.org/10.1016/s0304-3940(98)00529-1
Steele CJ, Bailey JA, Zatorre RJ, Penhune VB (2013) Early musical training and white-matter plasticity in the corpus callosum: evidence for a sensitive period. J Neurosci 33(3):1282–1290. https://doi.org/10.1523/JNEUROSCI.3578-12.2013
Tervaniemi M, Hugdahl K (2003) Lateralization of auditory-cortex functions. Brain Res Brain Res Rev 43(3):231–246. https://doi.org/10.1016/j.brainresrev.2003.08.004
Tremblay P, Baroni M, Hasson U (2013) Processing of speech and non-speech sounds in the supratemporal plane: auditory input preference does not predict sensitivity to statistical structure. Neuroimage 66:318–332. https://doi.org/10.1016/j.neuroimage.2012.10.055
Tremblay P, Perron M, Deschamps I, Kennedy-Higgins D, Houde JC, Dick AS, Descoteaux M (2019) The role of the arcuate and middle longitudinal fasciculi in speech perception in noise in adulthood. Hum Brain Mapp 40(1):226–241. https://doi.org/10.1002/hbm.24367
Tremblay P, Brisson V, Deschamps I (2021) Brain aging and speech perception: effects of background noise and talker variability. Neuroimage 227:117675. https://doi.org/10.1016/j.neuroimage.2020.117675
Waldron-Perrine B, Axelrod BN (2012) Determining an appropriate cutting score for indication of impairment on the Montreal Cognitive Assessment. Int J Geriatr Psychiatry 27(11):1189–1194. https://doi.org/10.1002/gps.3768
White-Schwoch T, Woodruff Carr K, Anderson S, Strait DL, Kraus N (2013) Older adults benefit from music training early in life: biological evidence for long-term training-driven plasticity. J Neurosci 33(45):17667–17674. https://doi.org/10.1523/JNEUROSCI.2560-13.2013
Wong PC, Warrier CM, Penhune VB, Roy AK, Sadehh A, Parrish TB, Zatorre RJ (2008) Volume of left Heschl’s Gyrus and linguistic pitch learning. Cereb Cortex 18(4):828–836. https://doi.org/10.1093/cercor/bhm115
Wong PC, Jin JX, Gunasekera GM, Abel R, Lee ER, Dhar S (2009) Aging and cortical mechanisms of speech perception in noise. Neuropsychologia 47(3):693–703. https://doi.org/10.1016/j.neuropsychologia.2008.11.032
Wong PC, Ettlinger M, Sheppard JP, Gunasekera GM, Dhar S (2010) Neuroanatomical characteristics and speech perception in noise in older adults. Ear Hear 31(4):471–479. https://doi.org/10.1097/AUD.0b013e3181d709c2
Wright BA, Sabin AT (2007) Perceptual learning: how much daily training is enough? Exp Brain Res 180(4):727–736. https://doi.org/10.1007/s00221-007-0898-z
Zatorre RJ, Chen JL, Penhune VB (2007) When the brain plays music: auditory–motor interactions in music perception and production. Nat Rev Neurosci 8(7):547–558. https://doi.org/10.1038/nrn2152
Zendel BR, Alain C (2012) Musicians experience less age-related decline in central auditory processing. Psychol Aging 27(2):410–417. https://doi.org/10.1037/a0024816
Zendel BR, West GL, Belleville S, Peretz I (2019) Musical training improves the ability to understand speech-in-noise in older adults. Neurobiol Aging 81:102–115. https://doi.org/10.1016/j.neurobiolaging.2019.05.015
Acknowledgements
We thank Valérie Brisson for her valuable help in the development of the task and for her contribution to the recruitment and testing of participants. We also thank Julie Poulin, Emilie Belley, Lisa-Marie Deschênes, Anne-Christine Bricaud, Elena Vaccaro, and Antoine Halbaut for their contributions to the recruitment and testing of participants, as well as all others research assistants and research interns who contributed to the project. Technical support for protocol development and data acquisition was provided by the “Centre intégré en neuroimagerie et neurostimulation de Québec” (CINQ).
Funding
This study was supported by grants to PT from the Drummond Foundation (2016RFA proposal #27) and the Natural Sciences and Engineering Research Council of Canada (NSERC grant # RGPIN-2019-06534), and two Globalink research internships from MITACS. PT holds a Career award from the Fonds de la Recherche en Santé du Québec (FRQ-S, #35016). MP was funded by graduate scholarships from the Fonds de la Recherche en Santé du Québec (FRQS) and the Canadian Institutes of Health Research (CIHR).
Author information
Authors and Affiliations
Contributions
MP: Conceptualisation, Methodology, Investigation, Project administration, Formal analysis, Visualisation, Writing—Original draft preparation, Data Curation. JV: Conceptualisation, Writing—Reviewing and Editing. PT: Conceptualisation, Funding acquisition, Methodology, Investigation, Supervision, Resources, Project administration, Writing—Reviewing and Editing, Data Curation.
Corresponding author
Ethics declarations
Conflicts of interest/competing interests
The authors have no relevant financial or non-financial interests to disclose.
Ethics approval
Ethic approval was obtained from the Comité d’éthique de la recherche sectoriel en neurosciences et santé mentale, Institut Universitaire en Santé Mentale de Québec (#192–2017; #1495–2018). The procedures used in this study adhere to the tenets of the Declaration of Helsinki.
Consent to participate
Informed consent was obtained from all individual participants included in the study.
Consent for publication
Not applicable.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Perron, M., Vaillancourt, J. & Tremblay, P. Amateur singing benefits speech perception in aging under certain conditions of practice: behavioural and neurobiological mechanisms. Brain Struct Funct 227, 943–962 (2022). https://doi.org/10.1007/s00429-021-02433-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00429-021-02433-2