
Abstract
The aim of this study was to determine whether exogenous administration of C-type natriuretic peptide (CNP) induces functional and morphological vascular changes in spontaneously hypertensive rats (SHR) compared with normotensive rats. Male 12-week-old normotensive Wistar and SHR were administered with saline (NaCl 0.9%) or CNP (0.75 μg/h/100 g) for 14 days (subcutaneous micro-osmotic pumps). Systolic blood pressure (SBP) was measured in awake animals and renal parameters were evaluated. After decapitation, the aorta was removed, and vascular morphology, profibrotic markers, and vascular reactivity were measured. In addition, nitric oxide (NO) system and oxidative stress were evaluated. After 14-days of treatment, CNP effectively reduced SBP in SHR without changes in renal function. CNP attenuated vascular remodeling in hypertensive rats, diminishing both profibrotic and pro-inflammatory cytokines. Also, CNP activated the vascular NO system and exerted an antioxidant effect in aortic tissue of both groups, diminishing superoxide production and thiobarbituric acid-reactive substances, and increasing glutathione content. These results show that chronic treatment with CNP attenuates the vascular damage development in a model of essential hypertension, inducing changes in fibrotic, inflammatory, oxidative, and NO pathways that could contribute to beneficial long-term effects on vascular morphology, extracellular matrix composition, and function. The knowledge of these effects of CNP could lead to improved therapeutic strategies to not only control BP but also reduce vascular damage, primarily responsible for the risk of cardiovascular events.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
C-type natriuretic peptide (CNP) is extensively distributed in the cardiovascular system, particularly in endothelial cells [10, 17, 37], placing it strategically among the mechanisms involved in vascular integrity and tone regulation. The composition of the vessel wall, primarily the proportion of collagen and elastin fibers in the extracellular matrix, determines the mechanical properties of vessels [41]. Alterations in media thickness and extracellular matrix composition are characteristic of sustained elevation of blood pressure (BP) [18].
The spontaneously hypertensive rats (SHR) constitutes a model of hypertension that develops high systolic BP (SBP) from 4 to 6 weeks of age, reaching 180–200 mmHg from 12 weeks of age [35], and exhibits abnormal growth, apoptosis, inflammation, and fibrosis; all mechanisms that have been implicated in hypertensive arterial remodeling [14]. As in essential hypertension, SHR also present endothelial dysfunction, increased levels of reactive oxygen species (ROS) and vasoconstrictor factors, and decreased bioavailability and effectiveness of nitric oxide (NO) [7, 27].
One of the main signaling pathways involved in arterial remodeling is transforming growth factor beta 1 (TGF-β1), a pleiotropic factor expressed within vascular cells. Through the activation of Smad2/3 and 4 proteins, TGF-β1 can increase the synthesis of collagen in vascular smooth muscle cells [2]. In addition, TGF-β1 stimulates the synthesis of its own positive modulators by Smad pathway [8] and the production of ROS [16]. On the other hand, higher ROS production increases the expression of pro-inflammatory cytokines which, in turn, increases ROS production at sites of inflammation and injury [36]. mRNA of interleukin 6 (IL-6), IL-1β and tumor necrosis factor-alpha (TNF-α) are increased in the aorta of SHR compared with normotensive ones [13, 34].
In previous studies, we demonstrated that acute CNP administration decreases BP and increases endothelial NO synthase (eNOS) activity through natriuretic peptide receptor type-C (NPR-C) coupled Gi protein activation in the aorta of SHR [3, 4, 6], mediating conductance and peripheral vascular relaxation [3]. However, the acute response of NO system to CNP is impaired in vascular tissue of SHR [3, 4, 6]. Also, NPR-C activation induces a drop in ROS production in the vascular smooth muscle of SHR [33], showing the potential antioxidant effect of CNP.
Recently, new therapeutic strategies increase the bioavailability of natriuretic peptides (NP) for the treatment of heart failure by inhibiting their degradation [22, 39]. However, the effect of CNP on vascular morphology and tone when it is administered chronically in hypertensive states is still unknown. We hypothesized that CNP has beneficial effects on the characteristic vascular remodeling associated with high BP and can also improve vascular function by decreasing ROS and inducing NO system activation. Therefore, the aim of the present study was to evaluate and to compare the effect of chronic administration of CNP on BP and vascular remodeling and function in hypertensive and normotensive rats.
Materials and methods
Animals
Twelve-week-old male Wistar and SHR were purchased from Instituto de Investigaciones Médicas A. Lanari, Facultad de Medicina (Universidad de Buenos Aires, Argentina). Rats were housed in a humidity and temperature-controlled environment with an automatic 12-h light-dark cycle. They were fed standard rat chow from Nutrimentos Purina (Buenos Aires, Argentina) and tap water ad libitum up to the day of the experiments.
All experimental protocols were performed in accordance with the Guide for the Care and Use of Laboratory Animals (National Institutes of Health, Publication No. 85-23, Revised 1996) and with regulation 6344/96 of Administración Nacional de Medicamentos, Alimentos y Tecnología Médica (A.N.M.A.T.). Experimental procedures were approved by the Comité de Ética de la Facultad de Farmacia y Bioquímica (C.E.F.F.B.), Universidad de Buenos Aires, Buenos Aires, Argentina.
Materials
Antibodies against eNOS, TGF-β1, Smad proteins, and IL-6 were purchased from Santa Cruz Biotechnology (Santa Cruz, CA); antibody against phospho-eNOS (Ser1177) was purchased from Cell Signaling Technology Inc., (Danvers, MA); antibody against TNF-α was purchased from R&D Systems, (Minneapolis, MN); human CNP-22, antibody against actin, and all other chemicals used in these experiments were purchased from Sigma-Aldrich Chemical Co (St Louis, MO).
Experimental design
Wistar and SHR were randomly assigned to the CNP-treated group (n = 14): chronic infusion with CNP (0.75 μg/h/100 g) [15], or the control group (n = 14): chronic infusion with NaCl 0.9% w/v (saline), for 14 days. Chronic infusion in both groups was performed using Alzet micro-osmotic pumps (Model 1002) as we previously described [31]. SBP was measured in awake animals (tail-cuff method) with a MP100 Pulse Transducer, PanLab (Quad Bridge Amp, ADInstruments), and recorded with a polygraph (Quad Bridge Amp, ADInstruments) and a data acquisition software (PowerLab 8/30 and LabChart, Australia) at 0, 7, and 14 days of treatment. Renal parameters were evaluated to determine if changes in BP could be due to a renal effect of chronic CNP administration. Animals were weighted and placed in metabolic cages at the beginning and at the end of the experimental period. 24-h urine and plasma were collected to determine diuresis (gravimetrically), urinary sodium concentration (ion analyzer, Tecnolab, Model T-412), and creatinine clearance to estimate the glomerular filtration rate (GFR, Wiener lab, Argentine). The concentration of end products derived from NO metabolism (nitrites and nitrates, NOx) was determined according to the procedure we previously described [4]. Also, plasma samples were obtained for systemic oxidative stress evaluation at the end of the experimental period.
Vascular reactivity
The aorta from Wistar and SHR was removed immediately after decapitation and placed in cold oxygenated Krebs buffer. Isometric tension (g) was measured as we previously described [3] in the presence of phenylephrine (PE, 0.1 nmol/L–100 μmol/L), or acetylcholine (ACh, 0.01 nmol/L–100 μmol/L) and sodium nitroprusside (SNP, 0.1 pmol/L–10 μmol/L) in PE-preconstricted rings at EC80 of the maximum response to avoid differences between groups. Vascular reactivity was expressed as percentage of contraction with PE to KCl (90 mmol/L) maximum response ratio, or percentage of relaxation after PE precontraction.
Histological evaluation and immunolabeling
Hematoxylin-eosin staining allowed us to obtain media cross-sectional area (CSA) and media thickness (M) to lumen diameter (L) ratio in × 40 tissue sections [5]. Picrosirius red staining was performed to quantify the content of collagen expressing the results as percentage of staining area (% staining area/total area) [31]. Elastin fibers from tunica media were assessed from aorta slices stained with Verhoeff-Van Gieson staining [28]. The number of elastin layers was normalized to media thickness.
Immunohistochemistry for TGF-β1 and Smad proteins was performed on aorta sections using primary polyclonal anti-Smad protein antibody (1:100 dilution) or anti-TGF-β1 antibody (1:50 dilution). Sections were developed with 3,39-diaminobenzidine solution (DAB) as chromogen (Dako EnVision H + system-HRP) as we described previously [31].
TNF-α and IL-6 immunohistochemistry was developed using a commercial kit (Vectastain ABC, Universal Elite, Vector Laboratories, CA, USA) and primary polyclonal anti-IL-6 antibody (1:100 dilution) or anti-TNF-α antibody (1:50 dilution). Sections were developed with DAB solution as chromogen for 15 min, counterstained with hematoxylin, dehydrated, cleared, and mounted [12]. Negative controls were performed by omitting the primary antibodies. Results were expressed as the percentage of area with positive staining/mm2. Morphological analyses were performed at a magnification of × 400 or × 200, respectively. In all cases, the mean percentage values were then calculated for each rat.
Histological and immunohistological assays were analyzed using an Olympus BX51 light microscope equipped with a digital camera (Qcolor 3, Olympus America, Inc., Richmond Hill, Ontario, Canada). To perform an adequate images analysis, × 200 or × 400 magnification was used. Measurements were carried out with Image-Pro Plus 4.5.1.29 software (Media Cybernetics, LP, Silver Spring, MD, USA) or Image J software (NIH - National Institute of Health, USA). The measurements were performed blindly and under similar light, gain, offset, and magnification conditions.
IL-1β was not detected by immunohistochemistry in our conditions. Cytokine levels were determined in aorta homogenates of SHR by enzyme-linked immunosorbent assay (ELISA). The OptEIA system (BD, Franklin Lakes, NJ, US) was used for IL-1 according to manufacturer’s instructions. Results were expressed as pg/mg of protein.
Determination of NOS activity and Western blot analysis
Aortic NOS activity was measured using [14C] L-arginine as substrate as described previously [4]. Results were expressed as picomoles of [14C] L-citrulline per gram wet weight per minute. Western blotting for eNOS (1/500 dilution), phospho-eNOS (Ser1177, 1/500 dilution), and β-actin (1/5000 dilution) were performed in aortic tissue as we previously described [3].
Oxidative stress evaluation
Lipid oxidative damage was assessed in plasma and aorta slices by measuring the extent of formation of 2-thiobarbituric acid-reactive substances (TBARS) using a fluorometer (Hitachi High-Technologies Corporation, F-3010) (excitation/emission wavelength 515 nm/555 nm) as we previously described [31]. A calibration curve was prepared using 1,1,3,3-tetramethoxypropane as standard [21]. Lipid peroxidation products were expressed as nmol of malondialdehyde (MDA) per milliliter of plasma or milligram of protein.
Glutathione concentration was measured in plasma and aorta, using a solution of 1 mol/L perchloric acid and 2 mmol/L EDTA (1:1 or 1:3, respectively). Samples were homogenized and centrifuged (20,000g, 20 min, 4 °C). The supernatant was filtrated through 0.22 μm cellulose acetate membranes (Corning Inc., NY, US). HPLC analysis was performed in a Perkin Elmer LC 250 liquid chromatography (Perkin Elmer ISS 250, Waltham, MA, US), equipped with a Perkin Elmer LC ISS 200 advanced sample processor and IV-visible detector (Waters 486 Tunable Absorbance Detector). A Supelcosil LC-18 (250 × 4.6 mm ID, 5 μm particle size) column was used for sample separation. Reduced (GSH) and oxidized glutathione (GSSG) was eluted at a flow rate of 1.2 mL/min with 20 mmol/L sodium phosphate (pH 2.7). GSH was spectrophotometrically detected [23]. However, GSSG was not detected under our conditions. Results were expressed as microgram of GSH per gram wet weight of the aorta and micromole per liter of plasma.
Basal superoxide anion (O2•-) production was measured in aortic rings of 1–2 mm of width using the lucigenin-enhanced chemiluminescence method with a low concentration of lucigenin [20]. Briefly, aorta-emitted luminescence was measured with a liquid scintillation counter (Labsystems Luminoskan RS Microplate Reader, Labsystem, Ramsey, Minnesota) using lucigenin solution (50 mmol/L Na3PO4, 1 mmol/L EGTA, 150 mmol/L sucrose, 20 μmol/L lucigenin, pH 7,4, 37 °C). Then, 250 μmol/L-reduced nicotinamide adenine dinucleotide phosphate was added and the emitted luminescence was measured. The luminescence value was expressed as arbitrary units (AU) per dry weight. To assess the specificity of the method, the measurement was performed in the presence of superoxide dismutase (200 U/mL) or 4-Hydroxy-2,2,6,6-tetramethylpiperidine 1-oxyl (20 mmol/L) as O2•- scavengers.
Statistical analysis
All values are expressed as mean ± S.E.M and n in each figure or table indicates the number of rats used. The Prism program (Graph Pad Software, Inc., San Diego, CA, USA) was used for statistical analysis.
The values of vascular reactivity responses to ACh and SNP are expressed as a percentage of the preceding contraction induced by phenylephrine. Sigmoidal dose-response model (four-parameter logistic equation with variable slope) was used as non-linear regression. The concentration of the agonist producing a half-maximal response (EC50) was determined after logit transformation of the normalized concentration-response curves, and it is reported as the negative logarithm (pEC50) of the mean of individual values for each tissue. The maximal relaxant effect (Emax) was considered to be the maximal amplitude response reached in concentration-effect curves to PE, ACh, or SNP.
Comparisons between two groups were analyzed by unpaired Student t test. Comparisons between more groups were performed by 2-way ANOVA followed by Bonferroni multiple comparisons test. Results were considered significant at p < 0.05.
Results
Systolic blood pressure in response to exogenous administration of CNP
SHR exhibited higher SBP levels than Wistar rats. After 7 days of treatment, CNP decreased SBP in SHR, showing a decrease of 36 mmHg at day 14 (18.5% of change). However, in SHR treated with CNP, SBP remained over the values found in control and CNP-treated normotensive rats. In contrast, there were no differences in SBP of Wistar groups at the end of the experimental protocol (Fig. 1). In addition, chronic treatment with CNP did not affect either body weight gain or renal function in both normotensive and hypertensive rats (Table 1).

Effects of chronic CNP treatment on systolic blood pressure. Control: saline administration for 14 days; CNP: CNP administration (0.75 μg/h/100 g) for 14 days; Data are expressed as mean ± S.E.M. (n = 12) and were analyzed using two-way ANOVA. The factors tested were treatment vs. strain: interaction significant (p < 0.001) and treatment vs. age: interaction significant (p < 0.001); simple effects were analyzed by a Bonferroni post hoc test for multiple comparisons between groups of interest †††p < 0.001 vs. Wistar control same age; ‡‡‡p < 0.001 vs. SHR control same age; *p < 0.05 vs. SHR 12-week-old; ***p < 0.001 vs. SHR 12-week-old
Evaluation of vascular structure and composition of the vascular wall in response to exogenous administration of CNP
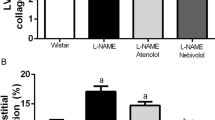
Hypertensive rats showed arterial wall hypertrophy, with increased CSA and M/L ratio compared with normotensive rats (Table 2). Also, aortic collagen content was higher in SHR than in Wistar rats (Fig. 2a, Table 2), but no significant differences were found in the number of elastic lamellae between both groups (Wistar = 8.07 ± 0.38; SHR = 8.14 ± 0.16). The number of elastic fibers to media thickness ratio was decreased in SHR compared with Wistar rats (Table 2).

Effects of chronic CNP treatment on vascular morphology. Control: 14-days saline administration; CNP: 14-days CNP administration (0.75 μg/h/100 g). a The red color of Sirius red staining under the light microscope indicates total collagen deposits, representative images are from the aorta of each group. b Representative micrographs of Verhoeff-van Gieson staining, black fiber indicates the elastin layer. All images are at the same magnification (× 400); scale bar = 50 μm
The morphological analysis showed that the parameters evaluated were not modified in Wistar rats by chronic treatment with CNP. Conversely, CNP decreased 27% of CSA and 34% of M/L of the aorta in SHR (Table 2). In addition, sections stained with collagen-specific Picrosirius red showed that CNP administration induced a drop of 43% of change in collagen content in SHR (Fig. 2a). Also, the administration of CNP for 14 days enhanced 71% of the elastin to media thickness ratio in SHR (Fig. 2b).
Immunohistochemistry for TGF-β1 and Smad proteins as factors responsible for vascular fibrosis in hypertension were also evaluated: SHR showed a higher percentage of staining for these factors compared with Wistar rats. CNP treatment reduced 27% of staining for TGF-β1 and decreased 54% of Smad immunostaining in SHR (Fig. 3a, b, Table 2). In addition, SHR showed higher immunostaining for both pro-inflammatory cytokines IL-6 and TNF-α in the aorta compared with Wistar control rats. Also, after 14 days of treatment with CNP, the staining for both pro-inflammatory cytokines decreased in SHR (% of changes in IL-6 and TNF-α levels induced by CNP treatment in SHR: 82% vs. 76%, respectively) (Fig. 4a, b). The measurement of IL-1β in aortas of SHR showed a decrease of 27% of the cytokine level in CNP-treated group (SHR: control = 1.96 ± 0.09; CNP = 1.42 ± 0.06*; *p < 0.05; n = 5 rats/group).

Effects of CNP chronic treatment on TGF-β1 and Smad immunostaining. Control: saline administration for 14 days; CNP: CNP administration (0.75 μg/h/100 g) for 14 days; C-: negative control. a Representative micrographs of TGF-β1 immunohistochemistry. b Representative micrographs of Smad proteins immunohistochemistry. All images are at the same magnification (× 400); scale bar = 50 μm

Effects of CNP chronic treatment on inflammatory markers. Control: saline administration for 14 days; CNP: CNP administration (0.75 μg/h/100 g) for 14 days. a Representative micrographs of IL-6 immunohistochemistry. b Representative micrographs of TNF-α immunohistochemistry. All micrographs are at the same magnification (× 200); scale bar = 100 μm
Study of vascular reactivity in response to chronic treatment with CNP
Phenylephrine-induced vasoconstriction was greater in SHR than in Wistar rats. Emax was increased in normotensive rats treated with CNP but it was decreased in CNP-treated SHR (Table 3). Endothelium-dependent vasorelaxation was also evaluated. The lowest concentrations of ACh increased the vascular tone of aortas in control SHR. Maximal ACh-induced vasodilatation was lower in SHR than in Wistar rats, and treatment with CNP for 14 days improved impaired relaxation, increasing the maximal ACh-induced vasodilation and pEC50 in SHR. However, since the pEC50 was calculated not having reached the plateau at low concentrations of acetylcholine, the accuracy of this value could be limited. CNP did not modify endothelium-dependent vasorelaxation in Wistar rats (Fig. 5b). SNP-induced relaxation was assessed in aortic rings to examine endothelium-independent relaxation in response to CNP chronic administration. CNP attenuated the potency to SNP only in Wistar rats (Fig. 5c, Table 3).

Evaluation of vascular reactivity in response to chronic CNP treatment in normotensive and hypertensive rats. Control: saline administration for 14 days; CNP: CNP administration (0.75 μg/h/100 g) for 14 days; concentration-response curves using a PE phenylephrine; b ACh acetylcholine; and c SNP sodium nitroprusside. Data are expressed as mean ± S.E.M. (n = 8). Data were analyzed using non-linear regression and a PE: two-way ANOVA, strain × treatment interaction: significant (p < 0.001); simple effects were analyzed by a Bonferroni post hoc test for multiple comparisons between groups of interest †††p < 0.001 vs. Wistar control; ‡‡‡p < 0.001 vs. SHR control. b ACh: two-way ANOVA, strain × treatment interaction: significant (p < 0.01); simple effects were analyzed by a Bonferroni post hoc test for multiple comparisons between groups of interest †††p < 0.001 vs. Wistar control; ‡p < 0.05 vs. SHR control; ‡‡‡p < 0.001 vs. SHR control. c SNP: two-way ANOVA, Bonferroni post hoc test; strain **p < 0.01 vs. Wistar control; ***p < 0.001 vs. Wistar control; treatment: no significant effect; strain × treatment interaction: no significant effect
Evaluation of NO system response to chronic CNP treatment
NOx excretion was evaluated as an indicator of NO systemic production. SHR presented higher NOx excretion in 24-h urine than Wistar rats, and an increase of 48% of NOx excretion was observed in normotensive rats after treatment with CNP (Fig. 6a). In order to verify whether the increase in NOx was due to an increase in activity and/or expression of NOS, NOS activity and expression in the aorta were studied. The aortic activity of the enzyme was higher in hypertensive than in normotensive rats and chronic treatment with CNP increased NOS activity in the aorta in both groups (Fig. 6b). The increase in vascular NOS activity in response to CNP treatment was more marked in Wistar than in SHR (changes in NOS activity induced by CNP treatment (%): Wistar 122% vs. SHR 66%). Also, aortic eNOS protein content was higher in SHR than in Wistar rats. Chronic treatment with CNP increased 21% of eNOS total expression in normotensive rats and eNOS phosphorylation was linked to increased expression of eNOS in this group. However, CNP did not modify eNOS total expression in SHR but increased 27% of eNOS phosphorylation levels (Fig. 6c, d).

NO system response to chronic treatment with CNP in normotensive and hypertensive rats. Control: 14 days saline administration; CNP: 14 days CNP administration (0.75 μg/h/100 g). a NOx, nitrites and nitrates (end products derived from NO metabolism) renal excretion. b NOS, aortic NO synthase activity. c Aortic eNOS protein content. d Aortic phospho-eNOS (Ser1177) protein expression. Data are mean ± S.E.M. (n = 6 to 8 rats per group). Statistical analysis was performed using a NOx: Two-way ANOVA, strain × treatment interaction: significant (p < 0.05); simple effects were analyzed by a Bonferroni post hoc test for multiple comparisons between groups of interest ††p < 0.01 vs. Wistar control; b NOS: two-way ANOVA, strain × treatment interaction: significant (p < 0.05); simple effects were analyzed by a Bonferroni post hoc test for multiple comparisons between groups of interest †††p < 0.001 vs. Wistar control; ‡‡‡p < 0.001 vs. SHR control; c eNOS: two-way ANOVA, strain × treatment interaction: significant (p < 0.05); simple effects were analyzed by a Bonferroni post hoc test for multiple comparisons between groups of interest ††p < 0.01 vs. Wistar control; and d phospho-eNOS (Ser1177): two-way ANOVA, strain × treatment interaction: significant (p < 0.05); simple effects were analyzed by a Bonferroni post hoc test for multiple comparisons between groups of interest ‡‡p < 0.01 vs. SHR control
Evaluation of oxidative stress in response to chronic CNP treatment
SHR showed more ROS production than normotensive rats and displayed lower levels of GSH; SHR also showed higher content of TBARS in both the aorta and plasma when compared with the Wistar group (Fig. 7). In order to verify if CNP induces an antioxidant effect in normotensive and hypertensive rats due to the regulation of ROS, we evaluated the production of O2•- in the aorta. Our results showed that SHR exhibited higher levels of O2•- compared with Wistar rats and CNP treatment induced a drop in O2•- production in both groups (Fig. 7a), but it was more marked in SHR (changes in O2•- levels induced by CNP treatment: Wistar 21% vs. SHR 57%). Chronic treatment with CNP increased the levels of GSH in the aorta and plasma of both Wistar and SHR. The effect of CNP on vascular tissue was more marked in Wistar rats, but in plasma, it was similar between both groups of animals (changes in GSH levels induced by CNP treatment in the aorta: Wistar 387% vs. SHR 117%; in plasma: Wistar: 98% vs. SHR: 95%). CNP also decreased 25% of aortic TBARS levels in SHR (Fig. 7c, e).

Effects of chronic CNP treatment on vascular and systemic oxidative stress. Control: saline administration for 14 days; CNP: CNP administration (0.75 μg/h/100 g) for 14 days. a O2•-: superoxide anion production; b GSH content; and c TBARS, thiobarbituric acid-reactive species concentration in the aorta. d GSH content and e TBARS concentration in plasma. Data are expressed as mean ± S.E.M. (n = 6–8 rats/group). Statistical analysis was performed using a O2•- and (b) GSHaorta: two-way ANOVA, strain × treatment interaction: significant (p < 0.001); (c) TBARSaorta: two-way ANOVA, strain × treatment interaction: significant (p < 0.05); simple effects were analyzed by a Bonferroni post hoc test for multiple comparisons between groups of interest ††p < 0.01 vs. Wistar control; †††p < 0.001 vs. Wistar control; ‡p < 0.05 vs. SHR control; ‡‡‡p < 0.01 vs. SHR control. d GSHplasma: two-way ANOVA, Bonferroni post hoc test; strain: no significant effect; treatment *p < 0.05 vs. control; **p < 0.01 vs. control; strain × treatment interaction: no significant effect. e TBARSplasma: two-way ANOVA, Bonferroni post hoc test; strain ###p < 0.001 vs. Wistar; treatment: no significant effect; strain × treatment interaction: no significant effect
Discussion
The novelty of this work lies in demonstrating that exogenous administration of CNP attenuates vascular changes associated with essential hypertension. Although SBP remains higher in CNP-treated SHR than in normotensive rats after 14 days of treatment, CNP exerts beneficial effects on both vascular dysfunction and remodeling in SHR. In addition, chronic administration of CNP does not induce either alteration in vascular wall integrity or a hypotensive effect but enhances NO system and improves oxidative state in normotensive animals.
Our study showed an imbalance in the response to vasoconstrictor and vasodilator factors in hypertensive rats. Moreover, we showed that the addition of lower concentrations of ACh increased vascular tone in SHR. In accordance with this result, it was recently described that ACh induces endothelium-dependent contractions in mouse carotid arteries through transient receptor potential channel C5 (TRPC5)-mediated Ca2+ entry and by stimulating cyclooxygenase (COX)-2-linked production of prostanoids in endothelial cells [20]. Furthermore, we demonstrated that the impact of chronic treatment with CNP on vascular reactivity differs between hypertensive and normotensive rats. In normotensive rats, the response to PE was increased and the vasodilator potency of NO donor was diminished. Although the aorta is a conduit vessel, these could be compensatory mechanisms that would contribute to maintain BP values in normotensive rats chronically treated with CNP in contrast to our previous findings with acute administration of the peptide [6].
On the other hand, response to PE was decreased and endothelium-dependent vasorelaxation was increased in SHR after chronic CNP treatment, suggesting that CNP could be enhancing the relaxant tone in hypertensive states, acting as vasodilator as well as modulating other vasoactive factors. A previous study supports this hypothesis showing that administration of a NPR-C selective agonist, cANP(4-23), induces an increase in the vasodilator response to carbachol in SHR, whereas no differences were observed in Wistar Kyoto rats [19]. However, the fact that the effect of NPR-C selective agonist on the relaxation induced by carbachol could be due to increased plasma levels of other NP, rather than exclusively to activation of the NPR-C receptor, cannot be overlooked because the high-affinity binding of cANP(4-23) to its receptor could decrease the clearance of circulating NP.
Chronic treatment with CNP enhances NOS activity in the aorta of both groups. However, the percentage of increase induced by CNP was lower in SHR and no significant increase was observed in NOx excretion compared with Wistar rats, suggesting an alteration in long-term NO system response to CNP which could contribute, at least in part, to the maintenance of high levels of BP in SHR. These results agree with our previous findings showing lower vascular NO system response to in vitro administration of CNP [4]. It is interesting to note that protein levels of the NPR-C receptor are decreased in the aorta of SHR compared with normotensive rats [19], and NO decreases NPR-C expression [1] which is involved in the activation of eNOS induced by CNP [3, 4, 6]. Previous data on eNOS protein expression in SHR are controversial. Studies performed on vascular smooth muscle showed lower eNOS expression in hypertensive animals [7, 11, 19]. In contrast, other authors found an increase in eNOS expression in SHR at 8, 12, 16, and 18 weeks of age compared with their normotensive controls [3, 4, 27, 38, 40]. The present study showed that not only the activity of the enzyme and urinary excretion of NO metabolites are higher in control SHR compared with Wistar rats, but also eNOS protein expression in the aorta.
In addition, analysis of eNOS protein levels after the 14-day treatment with CNP showed another difference between normotensive and hypertensive rats. In the aorta of Wistar rats, CNP treatment induced an increase in eNOS expression and a proportional increase in its phosphorylation at Ser1177. However, in SHR, CNP increased phosphorylation of the enzyme but total protein content was unchanged. Although the enhanced NOS activity induced by chronic CNP administration could be responsible for the lower contractile response to PE in SHR, this was not evaluated in the present study. Therefore, lower expression of the NPR-C receptor [19] that mediates activation of the NO system [4], higher basal activity of NOS, and no effect of CNP on eNOS protein level in SHR could be some of the mechanisms involved in altered vascular homeostasis observed in this model of hypertension.
Although the NO system appears to be exacerbated in SHR, this model has higher values of BP and decreased response to ACh or SNP. These functional alterations could be associated with the higher oxidative state found in SHR. CNP administration diminished vascular and plasmatic oxidative damage in SHR. Moreover, in normotensive rats, we observed an improvement in antioxidant species content even without changes in SBP. In agreement with our results, other study showed that activation of NPR-C receptor induces a drop in NADPH oxidase activity by decreasing p47phox and Nox4 subunits expression and the enzyme activity in vascular smooth muscle cells [19]. Therefore, CNP would enhance the bioavailability of vascular NO not only by inducing eNOS activity but also by decreasing oxidative state. Consequently, NO system enhancement could be involved in the effects of chronic CNP treatment on ACh-induced relaxation and BP drop in SHR.
It is well established that NADPH oxidase activation, and the consequent increase in the production of O2•-, also participates in the profibrotic effect of TGF-β1 [32]. Moreover, increased ROS can, in turn, stimulate the TGF-β1/Smad pathway [16, 32]. In the present study, we have demonstrated, for the first time, the in vivo effects of CNP administration on characteristic vascular changes of essential hypertension. Chronic treatment with CNP decreased both the media to lumen ratio and the early signs of fibrosis in the aorta. In addition, we observed that CNP decreased both TGF-β1 and Smad content in aortas from SHR. In vitro studies have suggested that CNP is able to modulate extracellular matrix deposits and profibrotic effects of several cytokines, since an increase in any of them induces the expression of CNP in cell culture [25] and overexpression of CNP reduces neointima formation after external injury in carotid arteries [30].
Also, vascular morphological changes observed in control SHR could be related to TGF-β1 that can exert pro-apoptotic or anti-apoptotic effects depending on the cell type, the conditions to which cells are exposed, or the combination of both [24, 29]. Also, this growth factor could be involved in the remodeling induced by CNP in aortas from hypertensive rats, but the underlying mechanisms need to be further examined in future studies.
We also found higher levels of pro-inflammatory cytokines in SHR than in Wistar rats. In fact, increased mRNA levels of IL-6, IL-1β, and TNF-α were previously found in the aorta of SHR [34]. In the present study, we showed that 14-day treatment with CNP decreased aortic IL-6, IL-1β, and TNF-α content in SHR, with no changes in cytokines measured in normotensive rats. It has been previously described that pro-inflammatory cytokines stimulate the release of CNP in endothelial cells [26], providing evidence of a possible role of CNP as a modulator of the inflammatory process. In addition, NO may also limit the activation of endothelial cells induced by pro-inflammatory cytokines, such as IL-6 [9]. Consequently, one of the possible mechanisms that might participate in the anti-inflammatory effect of CNP observed in the aorta of SHR is by stimulating NO system through NPR-C activation. However, further studies are needed to clarify the role of this peptide in this complex process.
Therefore, this study shows for the first time that exogenous CNP administration decreases pro-inflammatory markers in the aorta, in addition to the drop in early signs of hypertension-related fibrosis. The decreased BP may have partially contributed to the observed vascular changes in SHR. However, the changes observed on NO system and oxidative stress in Wistar rats suggests a direct effect of CNP on the regulation of vascular homeostasis. Moreover, these effects would have a beneficial impact on endothelial dysfunction and vascular remodeling associated with hypertension. Also, it is important to note that chronic administration of CNP does not alter integrity of the vascular wall in normotensive animals, but it improves vascular function and morphology in hypertensive rats, suggesting that the effect of this peptide could be dependent on the presence of hemodynamic and/or structural alterations.
Conclusion
Consequently, CNP not only acts as a key factor in the regulation of vascular homeostasis but also this peptide is able to diminish the alterations associated with increased risk of cardiovascular events in hypertensive states. The current study provides valuable evidence on the effect of chronic administration of CNP that could be useful in the improvement of therapeutic strategies that may represent a breakthrough in the treatment of hypertension not only by decreasing BP but also by protecting against associated target organ damage.
References
Arejian M, Li Y, Anand-Srivastava MB (2009) Nitric oxide attenuates the expression of natriuretic peptide receptor C and associated adenylyl cyclase signaling in aortic vascular smooth muscle cells: role of MAPK. Am J Physiol Heart Circ Physiol 296(6):H1859–H1867. https://doi.org/10.1152/ajpheart.01108.2008
Barcellos-Hoff MH, Dix TA (1996) Redox-mediated activation of latent transforming growth factor-beta1. Mol Endocrinol 10:1077–1083. https://doi.org/10.1210/mend.10.9.8885242
Caniffi C, Cerniello FM, Gobetto MN, Sueiro ML, Costa MA, Arranz C (2016) Vascular tone regulation induced by C-type natriuretic peptide: differences in endothelium-dependent and -independent mechanisms involved in normotensive and spontaneously hypertensive rats. PLoS One 11(12):e0167817. https://doi.org/10.1371/journal.pone.0167817
Caniffi C, Elesgaray R, Gironacci M, Arranz CT, Costa MA (2010) C-type natriuretic peptide effects on cardiovascular nitric oxide system in spontaneously hypertensive rats. Peptides 31(7):1309–1318. https://doi.org/10.1016/j.peptides.2010.03.030
Ceron CS, Rizzi E, Guimaraes DA, Martins-Oliveira A, Cau SB, Ramos J, Gerlach RF, Tanus-Santos JE (2012) Time course involvement of matrix metalloproteinases in the vascular alterations of renovascular hypertension. Matrix Biol 31(4):261–270. https://doi.org/10.1016/j.matbio.2012.01.009
Costa MA, Elesgaray R, Caniffi C, Fellet A, Arranz C (2007) Role of cardiovascular nitric oxide system in C-type natriuretic peptide effects. Biochem Biophys Res Commun 359(1):180–186. https://doi.org/10.1016/j.bbrc.2007.05.095
Crabos M, Coste P, Paccalin M, Tariosse L, Daret D, Besse P, Bonoron-Adèle S (1997) Reduce basal NO-mediated dilation and decreased NO-synthase expression in coronary vessels of spontaneously hypertensive rats. J Mol Cell Cardiol 29:55–65. https://doi.org/10.1006/jmcc.1996.0251
Cucoranu I, Clempus R, Dikalova A, Phelan PJ, Ariyan S, Dikalov S, Sorescu D (2009) NAD(P) H oxidase 4 mediates transforming growth factor-beta1-induced differentiation of cardiac fibroblasts into myofibroblasts. Circ Res 97:900–907. https://doi.org/10.1161/01.RES.0000187457.24338.3D
De Caterina R, Libby P, Peng HB, Thannickal VJ, Rajavashisth TB, Gimbrone MA Jr et al (1995) Nitric oxide decreases cytokine-induced endothelial activation. Nitric oxide selectively reduces endothelial expression of adhesion molecules and proinflammatory cytokines. J Clin Invest 96(1):60–68. https://doi.org/10.1172/JCI118074
D'Souza SP, Davis M, Baxter GF (2004) Autocrine and paracrine actions of natriuretic peptides in the heart. Pharmacol Ther 101(2):113–129. https://doi.org/10.1016/j.pharmthera.2003.11.001
Dubois G (1996) Decreased L-arginine-nitric oxide pathway in cultured myoblasts from spontaneously hypertensive versus normotensive Wistar-Kyoto rats. FEBS Lett 392(3):242–244
Giani JF, Miquet JG, Muñoz MC, Burghi V, Toblli JE, Masternak MM, Kopchick JJ, Bartke A, Turyn D, Dominici FP (2012) Upregulation of the angiotensin-converting enzyme 2/angiotensin-(1-7)/Mas receptor axis in the heart and the kidney of growth hormone receptor knock-out mice. Growth Hormon IGF Res 22(6):224–233. https://doi.org/10.1016/j.ghir.2012.08.003
Gonzalez W, Fontaine V, Pueyo ME, Laquay N, Messika-Zeitoun D, Philippe M, Arnal JF̧, Jacob MP, Michel JB (2000) Molecular plasticity of vascular wall during N(G)-nitro-L-arginine methyl ester-induced hypertension: modulation of proinflammatory signals. Hypertension 36(1):103–109
Intengan HD, Schiffrin EL (2001) Vascular remodeling in hypertension: roles of apoptosis, inflammation, and fibrosis. Hypertension 38(3 Pt 2):581–587
Itoh T, Nagaya N, Murakami S, Fujii T, Iwase T, Ishibashi-Ueda H, Yutani C, Yamagishi M, Kimura H, Kangawa K (2004) C-type natriuretic peptide ameliorates monocrotaline-induced pulmonary hypertension in rats. Am J Respir Crit Care Med 170(11):1204–1211. https://doi.org/10.1164/rccm.200404-455OC
Jiang F, Liu GS, Dusting GJ, Chan EC (2014) NADPH oxidase-dependent redox signaling in TGF-β-mediated fibrotic responses. Redox Biol 2:267–272. https://doi.org/10.1016/j.redox.2014.01.012
Kelsall CJ, Chester AH, Sarathchandra P, Singer DRJ (2006) Expression and localization of C-type natriuretic peptide in human vascular smooth muscle cells. Vasc Pharmacol 45:368–373. https://doi.org/10.1016/j.vph.2006.06.011
Leong XF, Ng CY, Jaarin K (2015) Animal models in cardiovascular research: hypertension and atherosclerosis. Biomed Res Int 2015:528757. https://doi.org/10.1155/2015/528757
Li Y, Sarkar O, Brochu M, Anand-Srivastava MB (2014) Natriuretic peptide receptor-C attenuates hypertension in spontaneously hypertensive rats: role of nitroxidative stress and Gi proteins. Hypertension 63(4):846–855. https://doi.org/10.1161/HYPERTENSIONAHA.113.01772
Liang C, Zhang Y, Zhuo D, Lo CY, Yu L, Lau CW, Kwan YW, Tse G, Huang Y, Yao X (2019) Endothelial cell transient receptor potential channel C5 (TRPC5) is essential for endothelium-dependent contraction in mouse carotid arteries. Biochem Pharmacol 159:11–24. https://doi.org/10.1016/j.bcp.2018.11.002
Magnani ND, Marchini T, Vanasco V, Tasat DR, Alvarez S, Evelson P (2013) Reactive oxygen species produced by NADPH oxidase and mitochondrial dysfunction in lung after an acute exposure to residual oil fly ashes. Toxicol Appl Pharmacol 270(1):31–38. https://doi.org/10.1016/j.taap.2013.04.002
Mangiafico S, Costello-Boerrigter LC, Andersen IA, Cataliotti A, Burnett JC Jr (2013) Neutral endopeptidase inhibition and the natriuretic peptide system: an evolving strategy in cardiovascular therapeutics. Eur Heart J 34(12):886–893c. https://doi.org/10.1093/eurheartj/ehs262
Marchini T, Magnani ND, Paz ML, Vanasco V, Tasat D, González Maglio DH, Alvarez S, Evelson PA (2014) Time course of systemic oxidative stress and inflammatory response induced by an acute exposure to Residual Oil Fly Ash. Toxicol Appl Pharmacol 274(2):274–282. https://doi.org/10.1016/j.taap.2013.11.013
McCaffrey TA (2000) TGF-betas and TGF-beta receptors in atherosclerosis. Cytokine Growth Factor Rev 11(1–2):103–114
Moyes AJ, Khambata RS, Villar I, Bubb KJ, Baliga RS, Lumsden NG, Xiao F, Gane PJ, Rebstock AS, Worthington RJ, Simone MI, Mota F, Rivilla F, Vallejo S, Peiró C, Sánchez Ferrer CF, Djordjevic S, Caulfield MJ, MacAllister RJ, Selwood DL, Ahluwalia A, Hobbs AJ (2014) Endothelial C-type natriuretic peptide maintains vascular homeostasis. J Clin Invest 124(9):4039–4051. https://doi.org/10.1172/JCI74281
Osterbur K, Yu DH, DeClue AE (2013) Interleukin-1β, tumor necrosis factor-α and lipopolysaccharide induce C-type natriuretic peptide from canine aortic endothelial cells. Res Vet Sci 94(3):478–483. https://doi.org/10.1016/j.rvsc.2012.10.002
Piech A, Dessy C, Havaux X, Feron O, Balligand JL (2003) Differential regulation of nitric oxide synthases and their allosteric regulators in heart and vessels of hypertensive rats. Cardiovasc Res 57(2):456–467
Pieraggi M, Nejjar I, Julian M, Bouissou H (1986) Staining of elastic tissue by Verhoeff’s iron hematoxylin. Ann Pathol 6(1):74–77
Pollman MJ, Naumovski L, Gibbons GH (1999) Vascular cell apoptosis: cell type-specific modulation by transforming growth factor-beta1 in endothelial cells versus smooth muscle cells. Circulation 99:2019–2026
Qian JY, Haruno A, Asada Y, Nishida T, Saito Y, Matsuda T, Ueno H (2002) Local expression of C-type natriuretic peptide suppresses inflammation, eliminates shear stress-induced thrombosis, and prevents neointima formation through enhanced nitric oxide production in rabbit injured carotid arteries. Circ Res 91(11):1063–1069
Romero M, Caniffi C, Bouchet G, Elesgaray R, McLaughlin M, Tomat A et al (2013) Sex differences in the beneficial cardiac effects of chronic treatment with atrial natriuretic peptide in spontaneously hypertensive rats. PLoS One 8(8):e71992. https://doi.org/10.1371/journal.pone.0071992
Ruiz-Ortega M, Rodríguez-Vita J, Sanchez-Lopez E, Carvajal G, Egido J (2007) TGF-beta signaling in vascular fibrosis. Cardiovasc Res 74(2):196–206. https://doi.org/10.1016/j.cardiores.2007.02.008
Saha S, Li Y, Lappas G, Anand-Srivastava MB (2008) Activation of natriuretic peptide receptor-C attenuates the enhanced oxidative stress in vascular smooth muscle cells from spontaneously hypertensive rats: implication of Gi alpha protein. J Mol Cell Cardiol 44:336–344. https://doi.org/10.1016/j.yjmcc.2007.11.003
Sanz-Rosa D, Oubiña MP, Cediel E, de las Heras N, Vegazo O, Jiménez J, Lahera V, Cachofeiro V (2005) Effect of AT1 receptor antagonism on vascular and circulating inflammatory mediators in SHR: role of NF-κB/IκB system. Am J Physiol Heart Circ Physiol 288:H111–H115. https://doi.org/10.1152/ajpheart.01061.2003
Schiffrin EL (2012) Vascular remodeling in hypertension: Mechanisms and Treatment. Hypertension 59:367–374. https://doi.org/10.1161/HYPERTENSIONAHA.111.187021
Sprague AH, Khalil RA (2009) Inflammatory cytokines in vascular dysfunction and vascular disease. Biochem Pharmacol 78:539–552. https://doi.org/10.1016/j.bcp.2009.04.029
Stingo AJ, Clavell AL, Heublein DM, Wei CM, Pittelkow MR, Burnett JCJ (1992) Presence of C-type natriuretic peptide in cultured human endothelial cells and plasma. Am J Phys 263:H1318–H1321. https://doi.org/10.1152/ajpheart.1992.263.4.H1318
Vapaatalo H, Mervaala E, Nurminen ML (2000) Role of endothelium and nitric oxide in experimental hypertension. Physiol Res 49(1):1–10
Vardeny O, Miller R, Solomon SD (2014) Combined neprilysin and renin-angiotensin system inhibition for the treatment of heart failure. J Am Coll Cardiol HF (14)00338–2. https://doi.org/10.1016/j.jchf.2014.09.001
Vaziri ND, Ni Z, Oveisi F (1998) Upregulation of renal and vascular nitric oxide synthase in young spontaneously hypertensive rats. Hypertension 31(6):1248–1254
Wagenseil JE, Mecham RP (2009) Vascular extracellular matrix and arterial mechanics. Physiol Rev 89(3):957–989. https://doi.org/10.1152/physrev.00041.2008
Acknowledgments
The authors thank Sandra Landín for secretarial work, Gabriela Noceti for technical assistance, and Ana Borthwick for proofreading and language assistance.
Funding
This study was funded by Universidad de Buenos Aires (UBA) [UBACYT: 200-201001-00060]; Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET) [PIP-112-201101-00581]; and Instituto de Química y Metabolismo del Fármaco (IQUIMEFA), UBA-CONICET, Argentina.
Author information
Authors and Affiliations
Contributions
•Conceived of or designed study: Carolina Caniffi, Cristina Arranz
•Performed research: Carolina Caniffi, Flavia M. Cerniello, Gonzalo Bouchet, María L. Sueiro, Analía Tomat, Daniel González Maglio, Jorge E. Toblli
•Analyzed data: Carolina Caniffi, Flavia M. Cerniello, Daniel González Maglio, Jorge E. Toblli
•Contributed new methods or models: Carolina Caniffi, Analía Tomat, Jorge E. Toblli, Cristina Arranz
•Wrote the paper: Carolina Caniffi, Analía Tomat, Cristina Arranz
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
All procedures performed in studies involving animals were in accordance with the ethical standards of the institution or practice at which the studies were conducted.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Dr. Jorge E. Toblli passed away in 2018
Rights and permissions
About this article

Cite this article
Caniffi, C., Cerniello, F.M., Bouchet, G. et al. Chronic treatment with C-type natriuretic peptide impacts differently in the aorta of normotensive and hypertensive rats. Pflugers Arch - Eur J Physiol 471, 1103–1115 (2019). https://doi.org/10.1007/s00424-019-02287-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00424-019-02287-y