
Abstract
Large dense core vesicles and chromaffin granules accumulate solutes at large concentrations (for instance, catecholamines, 0.5–1 M; ATP, 120–300 mM; or Ca2+, 40 mM (12)). Solutes seem to aggregate to a condensed protein matrix, which is mainly composed of chromogranins, to elude osmotic lysis. This association is also responsible for the delayed release of catecholamines during exocytosis. Here, we compile experimental evidence, obtained since the inception of single-cell amperometry, demonstrating how the alteration of intravesicular composition promotes changes in the quantum characteristics of exocytosis. As chromaffin cells are large and their vesicles contain a high concentration of electrochemically detectable species, most experimental data comes from this cell model.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Successful neurotransmission is based on the capability of neurons to release enough chemicals to be detected by a target cell. Because neurotransmitters are diluted following secretion, it is crucial to package them in a very high concentration in the secretory organelle.
Exocytosis is the most important mechanism for secretion for a large number of substances including transmitters, hormones, most autacoids, or antibodies. Regulated exocytosis is a calcium-dependent mechanism that entails the fusion of a secretory organelle (synaptic vesicle, large dense core vesicle, chromaffin granule) with plasma membrane followed by the total or partial release of its content.
Chromaffin granules are a particular kind of large dense core vesicles present in chromaffin cells. Because of their high abundance in the adrenal medulla and their relatively easy culture for decades, chromaffin cells have been the most popular model to study the mechanisms involved in the stimulus-secretion coupling, including the exocytotic process itself.
The vesicular cocktail of chromaffin granules
The large concentration of solutes that chromaffin granules can accumulate in their interior has intrigued scientists for over half a century [64]. The theoretical osmolarity calculated by the sum of the concentration of vesicular solutes can be estimated to be about 1500 mOsm. This large osmotic gradient towards the cytosol (≈320 mOsm) can only be reduced by the aggregation of solutes thus forming a condensed matrix at acidic pH (≈5.5) to elude osmotic lysis. By altering the osmolarity of extracellular media, from 200 to 750 mOsm, of cultured chromaffin cells, we have observed profound and reversible changes in the kinetics of exocytosis [11, 58, 59].
We could classify intravesicular compounds into two major groups depending on their capacity to move across the vesicle’s membrane. Hence, amines, ascorbate, H+, Ca2+, and ATP are “mobile components” as they are moving in and out of the vesicle, whereas chromogranins, peptides, and other proteins are “immobile components” as they cannot easily leave the vesicles [64]. All mobile compounds of the vesicular cocktail are in equilibrium with the cytosol and the matrix, and as such, they are main candidates to be involved in the regulation of exocytosis since changes in any one of these species will affect the others.
Secretory vesicles in chromaffin cells are able to package very high concentrations of solutes maintaining a physiological osmolarity, thanks to proteins (such as chromogranins) or molecules (such as ATP) stored inside the granules.
Chromogranins
Chromogranins are the most abundant intragranular proteins in chromaffin cells. Chromogranins (Chromogranins A and B and secretogranin II) are acidic soluble glycoproteins responsible in part for the characteristic dense core of chromaffin vesicles seen in electron microscopy images. They are the major constituents of large dense core vesicles (including chromaffin granules), accounting for more than 80% of soluble intravesicular proteins. All granins consist of single-polypeptide chains. Chromogranins bind calcium, catecholamines, and ATP with low affinity but high capacity. For instance, a single chromogranin A molecule is able to bind 32–93 Ca2+ ions [65].
The manipulation of the chromogranin content by chga/chgb gene ablation reduces the amine content in a 50%, confirming the importance of the intravesicular matrix in the regulation of catecholamine cargo and exocytosis [18, 19, 42]. However, the calculated interaction between chromogranin A and catecholamines only could account for less than 20% of the total [55]. This discrepancy could be explained if we consider the presence of chromogranin B and other soluble components in millimolar concentrations, which will interact with chromogranins and catecholamines. This inner cocktail as a functional mixture also involves ATP and Ca2+ [36].
ATP
In terms of abundance, ATP is after catecholamines, the most important intravesicular component. Chromaffin granules behave as “ATP sequestrants” as vesicular ATP accounts for ≈75% of all cellular content [6, 16]. It should be noticed that almost all secretory vesicles from all known animal species contain ATP [7]. ATP is transported inside granule by a vesicular nucleotide carrier—the VNUT—(or Slc17a9) [53]. Decreased transporter expression results in a dramatic reduction in the exocytosis of ATP and catecholamines, both in the firing frequency and in the quantum size [22].
Three major roles have been proposed for vesicular ATP (1) to act as a neurotransmitter/mediator, (2) as ATP is a necessary actor in the granule motion and in the latest steps of exocytosis, it could be a functional reservoir including the participation in the own exocytosis of the vesicle, and (3) to contribute to the accumulation of vesicular components [7]. The colligative properties of ATP with catecholamines have been largely studied in vitro but only recently the information available about its role in living cells and in the dynamic of exocytosis has become available [22]. Although unproved with physiological stimuli, the presence of large concentrations just “across the granule membrane” of the two essential factors to trigger exocytosis (Ca2+ and ATP) opens the exciting question of the possible participation of vesicular components in its secretion [13, 43].
The gradient of H+ across granule membrane
The vesicular pH is ≈5.5, two orders of magnitude more acidic than cytosol. This is a crucial factor in understanding the equilibrium of its inner components as this pH coincides with the maximum stability of chromogranins and their optimal capacity to bind soluble species [28]. This is a result of their high content of glutamic and aspartic acid residues that produces a pI of 4.4–5.4 [23]. The H+ gradient across the granule membrane is also responsible for maintaining the high concentrations of amines, Ca2+, and ATP inside the vesicles as H+ are the counter-ions used by the carriers of these species [30]. The H+ gradient depends on the activity of a vesicular proton pump the V-ATPase, which is continuously pumping H+ to acidify the vesicle [45]. Chloride anions enter in vesicles to bring negative charges allowing the acidification by reducing the voltage gradient.
The effects of vesicle alkalinization prove the role of the pH gradient. For instance, the V-ATPase blocker bafilomycin reduces the released quantum size and slows down the process of exocytosis [13].
The regulation of the pH gradient across the granule membrane is probably a physiological target of several second messengers, which modify the kinetics of exocytosis and the granule cargo [12, 13]. We explored two of these cell-signaling pathways [10]. For example, the activation of the classical cGMP/PKG pathway by nitric oxide (NO) and other agents slows down catecholamine release in single exocytotic events, without changing the quantum size, an effect that can be rapidly reversed using NO scavengers [40]. Similar results were found after activation of the cAMP/PKA pathway, although strong stimulation of this kinase also causes a notable increase in the quantal release of catecholamines [9, 39]. Other drugs like estrogens also slow down exocytosis through a non-genomic mechanism that involves cAMP [35]. Activation of these two pathways causes a rapid alkalinization of granules [13].
Drugs as false neurotransmitters
In addition to the naturally occurring compounds, several drugs such as tyramine are accumulated inside secretory vesicles and displace the endogenous amines and promote leakage of them [17, 50]. Also, lipid soluble weak bases such as amphetamines have received attention since they are concentrated in a pH-dependent manner inside vesicles, reducing the quantum size (the amount of catecholamines released per single exocytotic event) by moving catecholamines towards the cytosol [25, 43, 56]. However, many other drugs share these chemical features of being permeable weak bases. It has been described how antihypertensive drugs like hydralazine [37] and β-adrenergic blockers [41] accumulate in the chromaffin granules and reduce the quantum size of catecholamine exocytosis. This accumulation of drugs inside the vesicles can also produce marked changes in the composition of the “mobile components,” like amines, Ca2+, and perhaps ATP, although it has received little attention by the scientific community. For example, vesicle content is depleted by incubation with reserpine or tetrabenazine, drugs that deplete vesicle contents [44]. Similarly, some drugs like CCCP used as “selective tools” for the study of mitochondrial calcium are potent drugs that displace mobile components from chromaffin granules. False neurotransmitters are co-released with catecholamines promoting pharmacological effects. Some of them like hydralazine [37] are rapidly stored in secretory vesicles whereas the others like β-blockers are slowly but progressively accumulated in vesicles, which could explain the delayed onset of some antihypertensive drugs [41].
Vesicles behave like a bi-compartmental storage site where the free portion accounts for only ≈10% of the total catecholamines [54]. This portion is probably associated with the clear halo observed in electron microscopy [15]. Changes in pH will rapidly affect this free fraction, which will initially modify the kinetics of exocytosis without altering the quantum size. However, strong or long-lasting inhibition of the V-ATPase also causes the leakage of amines and other soluble components like Ca2+ and ATP, which despite the decrease in the quantum content also promotes granule movement and exocytosis [12, 13].
Amperometry: a direct view of exocytosis
Although several methods for measuring the release of amines, such as catechols, indoles, or histamine have been implemented for over a century, only amperometry possesses the sensitivity, time course, and spatial resolution. In these experiments, a microelectrode is positioned onto a chromaffin cell surface to directly detect the secreted catecholamines. The water layer between electrode surface and cell membrane is thin enough that the electrode senses exocytosis similar to a postsynaptic cell.
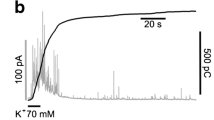
In amperometry, exocytosis is viewed as an amperometric spike. From this trace, several parameters can be extracted that yield important information about the amines released and their kinetics of release during an exocytotic event. Figure 1 shows a typical amperometric spike and the parameters that can be extracted from it.

Amperometry reveals that exocytosis occurs far more slowly than predicted by diffusion. a A typical amperometric trace obtained by a 5-s application of 5 mM BaCl2. Each secretory spike normally comes from a single fusion event. Note the large differences observed in spike sizes. When exocytosis events occur at high frequency, elevations of the basal level “envelops” are observed (arrow heads). b Kinetics parameters extracted from a secretory spike. The Imax is the maximum oxidation current of the spike which roughly corresponds to the maximal concentration reaching electrode surface, expressed in picoAmperes. The ascending slope (m) is taken by linearization of a segment between 25 and 75% of Imax. It is expressed in pascal per millisecond. As spike duration cannot be reliably set, the duration of spike at half height (t 1/2) is used instead as the measurement for spike duration: this parameter is expressed in millisecond. The integral of the current or area under the curve (Q) reflects the amount of catecholamines released, the apparent quantum size. It is expressed in picoCoulombs. Note the presence of a pre-spike feature or “foot” that precedes full exocytosis (dashed area), which corresponds to catecholamines diffusing from the fusion pore [14, 38]. c When spike simulation is carried out either by random walk or by equations that use the measured diffusional constant for catecholamines [27], the predicted speed of release is about three order of magnitude faster and the spike amplitude is predicted to be much greater
Amperometry evidenced the nature of catecholamine release during exocytosis
Two unexpected discoveries were noticed once chromaffin granules exocytosis could be observed:
-
(1)
The release of single quanta is quite prolonged. Single events of catecholamine release showed a much slower kinetics than predicted by a simple diffusion of catecholamines in aqueous solution, which should last few microseconds. Conversely, amperometric spikes have a duration of several tens of milliseconds; this indicates that free diffusion is impeded [14, 54] (Fig. 1).
-
(2)
Partial vs. full quantal release. Most of the granule content seems to be retained and only a partial release occurs.
Electrolysis of the secreted contents of single vesicles from bovine chromaffin cells typically yield a net charge (Q) that averaged ≈1 pC (5 attomoles or 3 × 106 molecules). As indicated in Fig. 1, the average value is distributed over a broad range. This has been attributed to the wide range of vesicle size [63].
However, patch-amperometry, a technique that can force the release towards the full collapse of vesicles, found Q values of ≈3 pC, thus, indicating that under normal stimulation most granules incomplete fusion or partial release [41, 42]. This interpretation received a further support from the so-called in vivo cytometry, a technique that placed a microelectrode in a flow stream of isolated vesicles that directly collide with the electrode surface causing their contents to be expelled [48]. This higher catecholamine content of secretory vesicles was also confirmed using a modification of this technique called “intracellular cytometry” by placing a very sharp electrode inside a cell monitoring directly the rupture vesicles [24]. These observations received support from data on chromaffin cells obtained by combining whole-cell with cell-attached capacitance. These authors showed that weak stimuli evoked release by transitory fusion whereas strong stimuli switched the release to the full-fusion mode [21].
Two main mechanisms have been claimed to explain why the granules empty slowly and in an incomplete fashion. Probably both are involved. The first mechanism concerns the physical restriction to the free diffusion of catecholamines caused by the dilatation/closing of fusion pore, an aqueous channel that makes the initial connection of vesicles with extracellular media. This means that most exocytotic events should be partial because the fusion pore only lasts long enough to permit flowing out only a fraction of the material to be secreted—partial exocytosis—.
The second mechanism arises from the retention of solutes in the granule matrix, which associates catecholamines, vesicular proteins (chromogranins), ATP, and Ca2+. This association is also shared by other cells like mastocytes which behave similar to chromaffin cells to changes in temperature and ionic composition in spite of their different matrix composition [51]. Exocytosis is also largely delayed in the presence of cross-linking agents like glutaraldehyde or formaldehyde that should freeze the dissociation of catecholamines from chromogranins [8]. Moreover, in experiments on chromaffin cells cultured in astrocyte-conditioned media, the phenotype of the chromaffin cells switches to a neuronal-like form. These chromaffin cells had reduced their granule matrix, and their secretory spikes resulted drastically accelerated [3]. However, the role of vesicular matrix could only be studied in detail when animals lacking chromogranins became available (see next paragraph).
The controversy between the supporters of full-fusion and partial fusion has lasted for decades. The presence of partial fusion, where all or part of the content is released during a transitory fusion event, has been reported in chromaffin and other cell types [1, 2, 29, 34].
Vesicular proteins can be retained in secretory vesicles depending on their size, the size of the fusion pore and the latency of the opening state. Live cell imaging of fluorescently labeled cargo and granule coat proteins showed that the granule can indeed remain seemingly intact despite an open pore to the outside [20, 31, 46, 49, 52, 57]. The size of this dilated pore has been estimated as 7–12 nm by probing the accessibility to the lumen for molecules up to several hundred kilodaltons [26, 33]. Larger peptides can be released through this pore, albeit slowly [49], or with a delay [4], which requires a pore size that is larger than the threshold for irreversible fusion. It has therefore been proposed that the initially narrow pore detected by electrical measurements can expand slowly, thereby acting as a filter for the released cargo based on its molecular size [4, 31]. However, even after release of peptides as large as tissue plasminogen activator (tPA-EGFP, ~100 kDa), the granule often remains intact for a prolonged period (minutes). Two observations indicate that even this expanded pore can close again: the first, by exploiting the pH dependence of a luminal EGFP-label, the resealing of granules was directly demonstrated by periodic acidification of the external medium [49] and the second, when cells are bathed in fluid phase markers (e.g., rhodamine, horseradish peroxidase) during stimulation, the dye becomes trapped in a fraction of the granules [5, 32, 46, 61, 62].
Data on chromaffin cells obtained by combining whole-cell with cell-attached capacitance showed that weak stimuli evoked release by transitory fusion whereas strong stimuli switched the release to the full-fusion mode [21].
The reuse of secretory vesicles means that every run causes the progressive loss of proteins of the matrix. This fact can offer an explanation for the heterogeneous size and density of vesicles observed in electron microscopy, which range from black to light gray. Thus, the relatively slow kinetics of exocytosis are a result of the bi-phasic distribution of soluble components inside vesicles (free and matrix-associated forms) as well as the relative size of species and the fusion pore dynamics.
Conclusions
It is now well established that the contents of secretory vesicles are closely associated. This association, common to a wide range of vesicles, was originally proposed to play a role in exocytosis of mucin from goblet cells [60]. Experimental results from amperometry and patch clamp experiments have established that the rate of extrusion from the vesicle is much slower than predicted for a diffusion-controlled process in free solution, consistent with the lowered extrusion rate predicted for a disassociation-controlled process. However, theoretical arguments suggest that the rate of expansion of the fusion pore and the association of intravesicular components are the rate-limiting steps in small molecule extrusion [47]. Clearly, more research is required to resolve these alternatives.
References
Ales E, Tabares L, Poyato JM, Valero V, Lindau M, and Alvarez de Toledo G. High calcium concentrations shift the mode of exocytosis to the kiss-and-run mechanism Nat Cell Biol 1: 40–44, 1999
Alvarez de Toledo G, Fernandez-Chacon R, and Fernandez JM. Release of secretory products during transient vesicle fusion. Nature 363: 554–558, 1993
Ardiles AO, Maripillan J, Lagos VL, Toro R, Mora IG, Villarroel L, Ales E, Borges R, Cardenas AM (2006) A rapid exocytosis mode in chromaffin cells with a neuronal phenotype. J Neurochem 99:29–41
Barg S, Olofsson CS, Schriever-Abeln J, Wendt A, Gebre-Medhin S, Renstrom E, Rorsman P (2002) Delay between fusion pore opening and peptide release from large dense-core vesicles in neuroendocrine cells. Neuron 33:287–299
Bauer RA, Khera RS, Lieber JL, Angleson JK (2004) Recycling of intact dense core vesicles in neurites of NGF-treated PC12 cells. FEBS Lett 571:107–111
Bevington A, Briggs RW, Radda GK, Thulborn KR (1984) Phosphorus-31 nuclear magnetic resonance studies of pig adrenal glands. Neuroscience 11:281–286
Borges R (2013) The ATP or the natural history of neurotransmission. Purinergic Signal 9:5–6
Borges R, Machado JD, Alonso C, Brioso MA, and Gomez JF. Functional role of chromogranins. The intragranular matrix in the last phase of exocytosis Adv Exp Med Biol 482: 69–81, 2000
Borges R, Machado JD, Betancor G, Camacho M (2002) Pharmacological regulation of the late steps of exocytosis. In: Chromaffin cell: transmitter biosynthesis, storage, release, actions, and informatics, vol 971, pp 184–192
Borges R, Machado JD, Betancor G, Camacho M (2002) Pharmacological regulation of the late steps of exocytosis. Ann N Y Acad Sci 971:184–192
Borges R, Travis ER, Hochstetler SE, Wightman RM (1997) Effects of external osmotic pressure on vesicular secretion from bovine adrenal medullary cells. J Biol Chem 272:8325–8331
Camacho M, Machado JD, Alvarez J, Borges R (2008) Intravesicular calcium release mediates the motion and exocytosis of secretory organelles: a study with adrenal chromaffin cells. J Biol Chem 283:22383–22389
Camacho M, Machado JD, Montesinos MS, Criado M, Borges R (2006) Intragranular pH rapidly modulates exocytosis in adrenal chromaffin cells. J Neurochem 96:324–334
Chow RH, von Ruden L, Neher E (1992) Delay in vesicle fusion revealed by electrochemical monitoring of single secretory events in adrenal chromaffin cells. Nature 356:60–63
Colliver TL, Pyott SJ, Achalabun M, Ewing AG (2000) VMAT-mediated changes in quantal size and vesicular volume. J Neurosci 20:5276–5282
Corcoran JJ, Korner M, Caughey B, Kirshner N (1986) Metabolic pools of ATP in cultured bovine adrenal medullary chromaffin cells. J Neurochem 47:945–952
Crout JR, Muskus AJ, Trendelenburg U (1962) Effect of tyramine on isolated guinea-pig atria in relation to their noradrenaline stores. Br J Pharmacol Chemother 18:600–611
Diaz-Vera J, Camacho M, Machado JD, Dominguez N, Montesinos MS, Hernandez-Fernaud JR, Lujan R, Borges R (2012) Chromogranins A and B are key proteins in amine accumulation, but the catecholamine secretory pathway is conserved without them. FASEB J 26:430–438
Diaz-Vera J, Morales YG, Hernandez-Fernaud JR, Camacho M, Montesinos MS, Calegari F, Huttner WB, Borges R, Machado JD (2010) Chromogranin B gene ablation reduces the catecholamine cargo and decelerates exocytosis in chromaffin secretory vesicles. J Neurosci 30:950–957
Dominguez N, Estevez-Herrera J, Borges R, Machado JD (2014) The interaction between chromogranin A and catecholamines governs exocytosis. FASEB J 28:4657–4667
Elhamdani A, Azizi F, Artalejo CR (2006) Double patch clamp reveals that transient fusion (kiss-and-run) is a major mechanism of secretion in calf adrenal chromaffin cells: high calcium shifts the mechanism from kiss-and-run to complete fusion. J Neurosci 26:3030–3036
Estevez-Herrera J, Dominguez N, Pardo MR, Gonzalez-Santana A, Westhead EW, Borges R, Machado JD (2016) ATP: the crucial component of secretory vesicles. Proc Natl Acad Sci U S A 113:E4098–E4106
Falkensammer G, Fischer-Colbrie R, Winkler H (1985) Biogenesis of chromaffin granules: incorporation of sulfate into chromogranin B and into a proteoglycan. J Neurochem 45:1475–1480
Fathali H, Dunevall J, Majdi S, Cans AS (2017) Extracellular osmotic stress reduces the vesicle size while keeping a constant neurotransmitter concentration. ACS Chem Neurosci 8:368–375
Fon EA, Pothos EN, Sun BC, Killeen N, Sulzer D, Edwards RH (1997) Vesicular transport regulates monoamine storage and release but is not essential for amphetamine action. Neuron 19:1271–1283
Fulop T, Radabaugh S, Smith C (2005) Activity-dependent differential transmitter release in mouse adrenal chromaffin cells. J Neurosci 25:7324–7332
Gerhardt G, Adams RN (1982) Determination of diffusion-coefficients by flow-injection analysis. Analyt Chem 54:2618–2620
Helle KB, Reed RK, Pihl KE, Serck-Hanssen G (1985) Osmotic properties of the chromogranins and relation to osmotic pressure in catecholamine storage granules. Acta Physiol Scand 123:21–33
Henkel AW, Meiri H, Horstmann H, Lindau M, Almers W (2000) Rhythmic opening and closing of vesicles during constitutive exo- and endocytosis in chromaffin cells. EMBO J 19:84–93
Henry JP, Sagne C, Bedet C, Gasnier B (1998) The vesicular monoamine transporter: from chromaffin granule to brain. Neurochem Int 32:227–246
Kasai H, Kishimoto T, Nemoto T, Hatakeyama H, Liu TT, Takahashi N (2006) Two-photon excitation imaging of exocytosis and endocytosis and determination of their spatial organization. Adv Drug Deliv Rev 58:850–877
Lang T, Wacker I, Steyer J, Kaether C, Wunderlich I, Soldati T, Gerdes HH, Almers W (1997) Ca2+-triggered peptide secretion in single cells imaged with green fluorescent protein and evanescent-wave microscopy. Neuron 18:857–863
Liu TT, Kishimoto T, Hatakeyama H, Nemoto T, Takahashi N, Kasai H (2005) Exocytosis and endocytosis of small vesicles in PC12 cells studied with TEPIQ (two-photon extracellular polar-tracer imaging-based quantification) analysis. J Physiol 568:917–929
Lollike K, Borregaard N, Lindau M (1998) Capacitance flickers and pseudoflickers of small granules, measured in the cell-attached configuration. Biophys J 75:53–59
Machado JD, Alonso C, Morales A, Gomez JF, Borges R (2002) Nongenomic regulation of the kinetics of exocytosis by estrogens. J Pharmacol Exper Ther 301:631–637
Machado JD, Diaz-Vera J, Dominguez N, Alvarez CM, Pardo MR, Borges R (2010) Chromogranins A and B as regulators of vesicle cargo and exocytosis. Cell Mol Neurobiol 30:1181–1187
Machado JD, Gomez JF, Betancor G, Camacho M, Brioso MA, Borges R (2002) Hydralazine reduces the quantal size of secretory events by displacement of catecholamines from adrenomedullary chromaffin secretory vesicles. Circ Res 91:830–836
Machado JD, Montesinos MS, Borges R (2008) Good practices in single cell amperometry. Methods Mol Biol 440:297–313
Machado JD, Morales A, Gomez JF, Borges R (2001) cAMP modulates exocytotic kinetics and increases quantal size in chromaffin cells. Mol Pharmacol 60:514–520
Machado JD, Segura F, Brioso MA, Borges R (2000) Nitric oxide modulates a late step of exocytosis. J Biol Chem 275:20274–20279
Montesinos MS, Camacho M, Machado JD, Viveros OH, Beltran B, Borges R (2010) The quantal secretion of catecholamines is impaired by the accumulation of beta-adrenoceptor antagonists into chromaffin cell vesicles. Br J Pharmacol 159:1548–1556
Montesinos MS, Machado JD, Camacho M, Diaz J, Morales YG, Alvarez de la Rosa D, Carmona E, Castaneyra A, Viveros OH, O'Connor DT, Mahata SK, and Borges R. The crucial role of chromogranins in storage and exocytosis revealed using chromaffin cells from chromogranin A null mouse J Neurosci 28: 3350–3358, 2008
Mundorf ML, Hochstetler SE, Wightman RM (1999) Amine weak bases disrupt vesicular storage and promote exocytosis in chromaffin cells. J Neurochem 73:2397–2405
Mundorf ML, Troyer KP, Hochstetler SE, Near JA, Wightman RM (2000) Vesicular Ca2+ participates in the catalysis of exocytosis. J Biol Chem 275:9136–9142
Nelson N, Harvey WR (1999) Vacuolar and plasma membrane proton-adenosinetriphosphatases. Physiol Rev 79:361–385
Obermuller S, Lindqvist A, Karanauskaite J, Galvanovskis J, Rorsman P, Barg S (2005) Selective nucleotide-release from dense-core granules in insulin-secreting cells. J Cell Sci 118:4271–4282
Oleinick A, Hu R, Ren B, Tian Z, Svir I, Amatore C (2016) Theoretical model of neurotransmitter release during in vivo vesicular exocytosis based on a grainy biphasic nano-structuration of chromogranins within dense core matrixes. J Electrochem Soc 163:H3014–H3024
Omiatek DM, Bressler AJ, Cans AS, Andrews AM, Heien ML, Ewing AG (2013) The real catecholamine content of secretory vesicles in the CNS revealed by electrochemical cytometry. Sci Rep 3:1447
Perrais D, Kleppe IC, Taraska JW, Almers W (2004) Recapture after exocytosis causes differential retention of protein in granules of bovine chromaffin cells. J Physiol 560:413–428
Philippu A, Schumann HJ (1965) Effect of alpha-methyldopa, alpha-methyldopamine, and alpha-methyl-norepinephrine on the norepinephrine content of the isolated heart. Life Sci 4:2039–2046
Pihel K, Travis ER, Borges R, Wightman RM (1996) Exocytotic release from individual granules exhibits similar properties at mast and chromaffin cells. Biophys J 71:1633–1640
Rutter GA, Tsuboi T (2004) Kiss and run exocytosis of dense core secretory vesicles. Neuroreport 15:79–81
Sawada K, Echigo N, Juge N, Miyaji T, Otsuka M, Omote H, Yamamoto A, Moriyama Y (2008) Identification of a vesicular nucleotide transporter. Proc Natl Acad Sci U S A 105:5683–5686
Schroeder TJ, Borges R, Finnegan JM, Pihel K, Amatore C, Wightman RM (1996) Temporally resolved, independent stages of individual exocytotic secretion events. Biophys J 70:1061–1068
Sen R, Sharp RR (1980) The soluble components of chromaffin granules. A carbon-13 NMR survey. Biochim Biophys Acta 630:447–458
Sulzer D, Maidment NT, Rayport S (1993) Amphetamine and other weak bases act to promote reverse transport of dopamine in ventral midbrain neurons. J Neurochem 60:527–535
Taraska JW, Perrais D, Ohara-Imaizumi M, Nagamatsu S, Almers W (2003) Secretory granules are recaptured largely intact after stimulated exocytosis in cultured endocrine cells. Proc Natl Acad Sci U S A 100:2070–2075
Troyer KP, Mundorf ML, Fries HE, and Wightman RM. Separating vesicle fusion and exocytosis in hypertonic conditions. In: Chromaffin cell: transmitter biosynthesis, storage, release, actions, and informatics 2002, p. 251–253
Troyer KP, Wightman RM (2002) Temporal separation of vesicle release from vesicle fusion during exocytosis. J Biol Chem 277:29101–29107
Verdugo P (1990) Goblet cells secretion and mucogenesis. Annu Rev Physiol 52:157–176
von Grafenstein H, Knight DE (1992) Membrane recapture and early triggered secretion from the newly formed endocytotic compartment in bovine chromaffin cells. J Physiol 453:15–31
von Grafenstein H, Knight DE (1993) Triggered exocytosis and endocytosis have different requirements for calcium and nucleotides in permeabilized bovine chromaffin cells. J Membr Biol 134:1–13
Wightman RM, Jankowski JA, Kennedy RT, Kawagoe KT, Schroeder TJ, Leszczyszyn DJ, Near JA, Diliberto EJ, Viveros OH (1991) Temporally resolved catecholamine spikes correspond to single vesicle release from individual chromaffin cells. Proc Nat Acad Sci USA 88:10754–10758
Winkler H, Westhead E (1980) The molecular organization of adrenal chromaffin granules. Neuroscience 5:1803–1823
Yoo SH, Albanesi JP (1990) Ca2(+)-induced conformational change and aggregation of chromogranin A. J Biol Chem 265:14414–14421
Acknowledgements
Supported by Spanish MINECO (BFU2013-45253).
Author information
Authors and Affiliations
Corresponding author
Additional information
This article is part of the special issue on Chromaffin Cells in Pflügers Archiv—European Journal of Physiology
Rights and permissions
About this article

Cite this article
Mark Wightman, R., Domínguez, N. & Borges, R. How intravesicular composition affects exocytosis. Pflugers Arch - Eur J Physiol 470, 135–141 (2018). https://doi.org/10.1007/s00424-017-2035-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00424-017-2035-6