
Abstract
Purpose
The present study identified the physiological and performance characteristics that are deterministic during a maximal 1500-m time trial and in paced 1500-m time trials, with an all-out last lap.
Methods
Thirty-two trained middle-distance runners (n = 21 male, VO2peak: 72.1 ± 3.2; n = 11, female, VO2peak: 61.2 ± 3.7 mL kg−1 min−1) completed a 1500-m time trial in the fastest time possible (1500FAST) as well as a 1500MOD and 1500SLOW trial whereby mean speed was reduced during the 0–1100 m by 5% and 10%, respectively. Anaerobic speed reserve (ASR), running economy (RE), the velocity corresponding with VO2peak (VVO2peak), maximal sprint speed (MSS) and maximal accumulated oxygen deficit (MAOD) were determined during additional testing. Carnosine content was quantified by proton magnetic resonance spectroscopy in the gastrocnemius and expressed as a Z-score to estimate muscle fibre typology.
Results
1500FAST time was best explained by RE and VVO2peak in female runners (adjusted r2 = 0.80, P < 0.001), in addition to the 0–1100-m speed relative to VVO2peak in male runners (adjusted r2 = 0.72, P < 0.001). Runners with a higher gastrocnemius carnosine Z-score (i.e., higher estimated percentage of type II fibres) and greater MAOD, reduced their last lap time to a greater extent in the paced 1500-m trials. Neither ASR nor MSS was associated with last lap time in the paced trials.
Conclusion
These findings suggest that VVO2 peak and RE are key determinants of 1500-m running performance with a sustained pace from the start, while a higher carnosine Z-score and MAOD are more important for last lap speed in tactical 1500-m races.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In elite 1500-m running competitions, championship races (i.e., World Athletics Championships and Olympic Games) can be won in relatively fast times (≤ 3:34.00), moderately fast (> 3:34.00– ≤ 3:41.99) or even slow times (≥ 3:42.00) (Sandford et al. 2019a), but with an extremely fast last lap (e.g., < 0:51.00). The tactical racing strategy that produces fast race times resides from a relatively even pacing profile with a sustained pace from the start (Tucker et al. 2006). Conversely, slower race times result from slow initial laps and a greater number of race surges, that precede a ‘last lap kick’ (Sandford et al. 2019a). Indeed, the ‘last lap kick’ may differentiate between successful and unsuccessful competitors in 1500-m championship races (Sandford et al. 2019a; Casado and Renfree 2018; Hanley et al. 2019); however, the physiological and performance characteristics that underpin last lap speed in tactical 1500-m races have not been fully elucidated.
Many attempts have been made to characterise the key physiological determinants of 1500-m running performance (Iwaoka et al. 1988; Ingham et al. 2008; Ferri et al. 2012; Billat et al. 1996, 2009). The prevailing conclusion from these studies is that aerobic characteristics, such as VO2peak (Ingham et al. 2008), the minimal running velocity that elicits VO2peak (vVO2peak) (Billat et al. 1996) and running economy (RE) (Ingham et al. 2008; Billat et al. 1996), are the most deterministic characteristics relating to 1500-m performance. Other studies that have profiled the energy system contribution to simulated (Spencer and Gastin 2001) and actual track running (Duffield et al. 2005) 1500-m time trials report that the predominant energy system contribution is derived from aerobic metabolism (77–85%). However, one caveat of this literature is that 1500-m running performance has been assessed under conditions whereby the subjects are likely to be attempting to run the fastest time possible, such as a maximal time trial (Ferri et al. 2012; Duffield et al. 2005), simulated maximal time trial on a treadmill (Spencer and Gastin 2001), a season best (Billat et al. 1996; Iwaoka et al. 1988) or recent 1500-m track performance (Ingham et al. 2008; Billat et al. 2009). During these scenarios, the key physiological and performance characteristics may not align with the key attributes that are required to perform a tactical race, characterized by an extremely fast last lap, but slower initial pace.
One concept that may underpin last lap speed is the anaerobic speed reserve (ASR), which is the difference between a runner's maximum sprinting speed (MSS) and vVO2peak (Bundle et al. 2003). While the ASR is negatively associated with 1500-m running speed (i.e., larger ASR equates to a slower mean 1500-m speed; r = − 0.68 ± 0.54) (Sandford et al. 2019c), it has been suggested that a greater ASR may be an important determinant of a fast last lap in a tactical, slow 1500-m race (Sandford et al. 2019b; Mytton et al. 2015), but this is yet to be tested experimentally. Alternatively, another concept that may underpin performance in tactical and maximal approaches to 1500-m racing arises from the parameters of the hyperbolic relationship between running speed and the tolerable duration, which is defined by the speed asymptote (critical speed; CS) and the curvature constant (D’) (Monod and Scherrer 1965). It could be purported that a 1500-m runner with a higher CS (but lower D’) would benefit from a higher sustained pace from the start of the race as another competitor with a lower CS (but greater D’) would be required to rely on a larger portion of their D' to maintain pace. In contrast, this competitor would likely be more suited to a reduced pace in the initial laps, to take advantage of a superior D’ for the ‘last lap kick’ (Pettitt 2016). However, despite the attractiveness of this proposal, further interrogation of the relationship between these parameters and 1500-m running performance with different pacing strategies is warranted. Lastly, it is conceivable that muscle fibre typology may be an important characteristic that could relate to preferential pacing strategies in middle-distance runners. Classical work (Costill et al. 1976) has shown that international level middle-distance runners possess a much greater between-athlete variation in muscle fibre typology (percentage type I fibres) in the gastrocnemius (mean; range: 52%; 41–69%) compared to long-distance runners (69%; 63–74%) and sprinters (24%; 21–27%). Given the superior capacity for anaerobic metabolism (Pette 1985), greater buffering capacity (Parkhouse et al. 1985) and superior mechanical properties (Harber et al. 2004) in type II compared to type I fibres, possessing a greater proportion of type II fibres relative to another competitor may be a key attribute for last lap speed when the initial laps are slow, but this proposal needs further examination. Therefore, the purpose of this study was to identify the physiological and performance characteristics that are deterministic during a maximal 1500-m time trial (i.e., fastest time possible) and in paced 1500-m time trials with either moderate or slow-paced initial laps, but an all-out last lap.
Methods
Subjects
Thirty-two trained middle-distance runners (n = 21 men, VO2peak: 72.1 ± 3.2 mL kg−1 min−1; n = 11 women, VO2peak: 61.2 ± 3.7 mL kg−1 min−1) participated in this study who had a consistent training history of at least 2 years in the 1500-m event and were without major injury interruption for the previous 3 months. The male runners had season best performance times during outdoor 1500-m track competition of 3:57.50 ± 0:09.70 (3:40.00–4:14.08 s), while the female runners achieved times of 4:39.20 ± 0:14.52 (4:17.44–5:08.10). At the time of the study, the male runners had a mean running training volume of 81.6 ± 16.2 km week−1, while the female runners ran 64.6 ± 12.1 km week−1. All runners provided written informed consent prior to participating in this study which was approved by the Griffith University Human Research Ethics Committee.
Study design
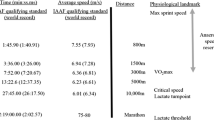
Subjects completed a 1500-m time trial in the fastest time possible (1500FAST) with lap split times recorded, and two pacing trials on an outdoor athletics track. In the pacing trials, runners were paced for the 0–1100 m, whereby speed was reduced on average by 5% (1500MOD) and 10% (1500SLOW) relative to 1500FAST, while the last 400 m was completed in the fastest time possible. In addition, subjects also completed laboratory treadmill running tests to determine RE, VO2peak, VVO2peak and the maximal accumulated oxygen deficit (MAOD). Subjects also completed additional outdoor time trials (2200 and 3000 m) for the quantification of CS and D’ and a MSS assessment (flying 50 m sprint) for the determination of the ASR. All trials were conducted on separate days across a 5-week period. Carnosine content was quantified by proton magnetic resonance spectroscopy in the gastrocnemius medialis muscle and was expressed as a sex-specific Z-score to estimate muscle fibre typology (Baguet et al. 2011).
Running time trials
The running time trials were conducted on an outdoor 400 m synthetic athletics track. All running trials were preceded by a standardized warmup that consisted of a 10-min self-paced jog which was followed by a 5-min bout of submaximal running that increased to a rating of perceived exertion of 5 (CR-10 scale) and six repetitions of 10-s strides, with a walk-back recovery (∼60 s), whereby strides were defined as bouts of fast running which were to be completed at each subjects perceived 1500-m race pace. The warm-up procedures were followed by 10 min of recovery. During 1500FAST, subjects ran individually and were not permitted to view their race split times and were instructed to complete the trial in the fastest time possible. Electronic timing gates (Smartspeed, Fusion Sport, Australia) were placed at the start and at the 300 m mark of the first lap to obtain each lap split time (0–400 m, 400–800 m, 800–1100 m and 1100–1500 m). In the paced trials (1500MOD and 1500SLOW), runners were paced for the 0–1100 m, whereby the 0–400 m lap speed was reduced by 10% (1500MOD) and 15% (1500SLOW), the 400–800 m split speed was reduced by 5% (1500MOD) and 15% (1500SLOW), while the 800–1100 m split speed remained equivalent to 1500FAST in both trials. Subjects were instructed to complete the final 400 m in the fastest time possible. These lap paces were chosen given that they reflected the relative speed decrement of moderately fast (> 3:34.00– ≤ 3:41.99) and slow (≥ 3:42.00) races relative to fast races (≤ 3:34.00), as classified as in the study of Sandford et al. (2019a). While these are clearly world class standard races, we employed the relative difference in lap times from these races to prescribe the split speeds of the paced trials in the present study. To assist with pacing from the 0–1100 m mark in the paced trials, each runner had a pacer who ran 4 m in front of the subject and had access to target split times and a wristwatch (Garmin Forerunner 235, Canton of Schaffhausen, Switzerland). Members of the research team also assisted with pacing by providing splits verbally at each 200 m mark. All subjects completed a familiarization of the paced trials on a separate day prior to the actual recorded paced trials. During the paced trials, if a subject recorded a lap split time within the 0–1100 m that deviated from the prescribed split by ≥ 1.00 s, the trial was discarded and performed on another day. This occurred on two occasions (both 1500SLOW trials).
Subjects also completed additional outdoor track time trials of 2200 and 3000 m to determine CS and D’. These distances were chosen to yield finishing times between 2 and 12 min (Hughson et al. 1984). These trials were performed with the same instructions as 1500FAST and CS and D’ were determined using a linear distance–time model. Subjects also performed a 50 m sprint for the determination of MSS. Linear sprint speed was evaluated over 50 m using electronic timing gates positioned at the start line and at 40 and 50 m. In addition to the standardized warmup procedure, subjects also performed two 50 m running efforts at 75% and 90% of each subject’s perceived maximal sprinting pace. MSS was determined as the mean speed derived from the 40 to 50 m split time. ASR was defined as the difference between the MSS and the vVO2peak, estimated from the submaximal and maximal incremental running test (see below). Air temperature, relative humidity and wind speed/direction were recorded using a thermal environment monitor (Questemp-15 Area Heat Stress Monitor, Quest Technologies, WI). Testing was conducted at an identical time of day (6:00–8:00 am) for all subjects which assisted in achieving relatively consistent atmospheric conditions (air temperature range 23.8–25.9 °C; relative humidity 66.0–71.0%; wind speed 4.6–7.6 km h–1). Eight subjects (n = 4 male and female runners) from this study performed duplicate trials on separate days one week apart to determine the test–retest reliability for the 1500-m time trials. The coefficient of variation (CV) for 1500FAST performance time and 1500MOD and 1500SLOW last lap times were 1.8%, 2.2% and 2.1%, respectively.
Maximal incremental running test
Following a warm-up (10 min at 8–14 km h−1) and a 5-min rest period, each subject performed an incremental treadmill (HP cosmos Saturn, Traunstein, Germany) run to volitional exhaustion starting at 10 km h−1 (women) or 12 km h−1 (men) and 1% gradient with the speed increasing by 1 km h−1 each min until volitional exhaustion. Respiratory variables were measured using a Cosmed Quark b2 (Rome, Italy), which had been calibrated according to the manufacturer’s instructions (Quinton Instruments, USA). Exhaled air was analysed to determine VO2 and VCO2. The gas exchange threshold (GET) was determined using the simplified V-slope method previously described (Schneider et al. 1993). VO2peak was determined as the highest VO2 value using a rolling 1-min average of breath-by-breath data. The inter-day CV for GET and VO2peak measurements in our lab is 2.00% and 3.13%, respectively (n = 26; measurements taken three weeks apart).
Submaximal laboratory running test
On a separate day, subjects completed six, 4-min submaximal incremental stages on a motorised treadmill which were completed in 5% incremental speeds ranging 85–110% of the GET which was determined from the maximal incremental running test. The treadmill belt was also set at 1% gradient (Jones and Doust 1996). Pulmonary (breath-by-breath) gas exchange was monitored throughout each stage, whereby the highest VO2 value was obtained using a rolling 1-min average of breath-by-breath data. Immediately following the final stage, an earlobe blood sample was taken for the determination of blood lactate concentration ([La]b) with a Lactate Pro 2 device (Arkray inc. Japan).
Calculation of running economy and vVO2peak
Oxygen uptake (VO2) and carbon dioxide production (VCO2) during the final minute of each submaximal stage were used to determine RE (kcal kg−1 km−1). RE was expressed as an energy cost, rather than an oxygen cost, as this accounts for differences in substrate oxidation within and between subjects and is more sensitive to changes in running speed (Fletcher et al. 2009). Updated non-protein respiratory quotient equations (Peronnet and Massicotte 1991) were used to estimate substrate utilisation (g min−1) during the final minute. The energy derived from each substrate was then calculated by multiplying fat and carbohydrate use by 9.75 kcal and 4.07 kcal, respectively, reflecting the mean energy content of the metabolised substrates during moderate to high-intensity exercise. The vVO2peak was calculated by solving the regression equation describing the individual VO2-running speed relationship based on the mean VO2 values and during the final min of each 4-min submaximal incremental stage and VO2peak measured from the maximal incremental running test using linear regression. The inter-day CV% for RE and vVO2peak in our lab is 2.3% and 3.1%, respectively (n = 26; measurements taken 3 weeks apart).
Supramaximal laboratory running test
The supramaximal running test was performed at a running speed equivalent to 110% of vVO2peak. Subjects performed a 15-min warm-up consisting of 5 min at 8—14 km h−1, 5 min at 140% of the GET and 2 × 6 s running bouts at 90% of vVO2peak, followed by 10 min of rest. Each subject wore a safety harness during the supramaximal effort which was attached to their chest to ensure maximal effort. Time to exhaustion was measured to the nearest second and blood lactate concentration was determined from earlobe samples taken at 1, 3, 5, and 7 min after the completion of the test, with the highest value obtained considered the peak blood lactate concentration.
Maximal accumulated oxygen deficit
As originally described (Medbo et al. 1988), the individual VO2-running speed relationship was extrapolated to estimate the VO2 demand corresponding to 110% of vVO2peak. The accumulated VO2 demand was estimated by multiplying the time to exhaustion by the VO2 demand and the MAOD was calculated by subtracting the accumulated VO2, measured during the exercise bout, from the estimated accumulated VO2 demand. Absolute MAOD values were reduced by 9% to correct for reductions in the oxygen stores of the body (Medbo et al. 1988). The CV for supramaximal TTE and MAOD calculations in our lab is 4.0% and 7.1%, respectively (n = 7; 4 men; measurements taken 1 week apart).
Carnosine quantification via 1H-MRS
Muscle carnosine content was measured by 1H-MRS in the gastrocnemius medialis muscle of each subject’s right limb to estimate muscle fibre typology. We chose to estimate the muscle fiber typology of the gastrocnemius medialis because (i) we can measure carnosine reliably in this muscle, (ii) carnosine content in the gastrocnemius medialis muscle has been positively correlated with the percentage area occupied by type II muscle fibers (Baguet et al. 2011), and (iii) the gastrocnemius medialis is an extremely active muscle during running, whereby the mean and maximal electromyographic activities relative to a maximal voluntary contraction is the highest compared to other prominent lower limb muscles (Tsuji et al. 2015). 1H-MRS measurements were performed on a 3-T whole body MRI scanner (Philips Medical Systems Best, The Netherlands). Subjects were lying in a supine position, while their lower leg was fixed in a spherical knee-coil. All the spectra were acquired using single voxel point-resolved spectroscopy (PRESS) with the following parameters: repetition time of 2000 ms, echo time of ~ 40 ms, number of excitations was 128 (carnosine) and 16 (water), spectral bandwidth was 2048 Hz, and an acquisition time of 4 min 16 s (carnosine) and 32 s (water). The voxel size was 40 mm × 15 mm × 20 mm. Spectral data analysis was carried out using jMRUI (version 6.0) with carnosine peaks fitted and expressed relative to the internal water signal.
Carnosine content (mM) was calculated using following formula:
where Cm is the carnosine concentration, CS is the carnosine signal, H2OS is the water signal, CT1r, CT2r, H2OT1r, H2OT2r are the relaxation correction factors for carnosine (earlier described by Baguet et al. (Baguet et al. 2011)) and water (earlier described by MacMillan et al. (MacMillan et al. 2016)), H2Omuscle is the concentration of water in muscle, which was deducted from the molar concentration of water (55,000 mM) and the approximate water content of skeletal muscle tissue (0.7 L/kg wet weight of tissue) and H2Oproton is the number of protons in water. The CV for test–retest inter-day carnosine measurements in our laboratory is 4.3% (gastrocnemius; n = 15 subjects). In the present study, the carnosine concentration was converted to a Z-score based on the normal distribution of our reference population. The absolute value of Z-score represents the distance between the individual value of a participant and the reference population mean value in units of the standard deviation. The reference sample population consisted of 40 recreationally active male and 33 female subjects who were scanned in our laboratory in the past 12 months.
Statistical analysis
Results are expressed as mean ± SD. Multiple stepwise regression was performed to identify the physiological and performance characteristics for which the majority of the variance in the overall time and last lap time of 1500FAST could be attributed to as well as the last lap time and improvement in last lap time in 1500MOD and 1500SLOW, relative to 1500FAST. Coefficients of determination were identified for the physiological and performance characteristics using univariate analyses and were included in the regression model if a significant r2 value (p < 0.05) was reported. The magnitude of correlations was classified as small: 0.10–0.29, moderate: 0.30–0.49, large: 0.50–0.69, very large: 0.70–0.89 and extremely large: 0.90–1.00. A two-way analysis of variance with Tukey’s post hoc test was conducted to compare segment speed times (i.e., 0–400, 400–800, 800–1100 and 1100–1500 m) across trials (i.e., 1500FAST, 1500MOD and 1500SLOW).
Results
1500-m trials
Table 1 displays the segment split times for each 1500-m trial. Given the manipulation in pacing, the 0–400 m and 400–800 m split times were faster in 1500FAST compared to 1500MOD and 1500MSLOW, while the 800–1100 m split was similar. However, the last lap was significantly faster in both 1500MOD and 1500MSLOW compared to 1500FAST. Table 2 displays the mean and range values for the physiological and performance variables.
1500FAST
The mean speed of 1500FAST relative to vVO2peak and CS was 101.2% ± 3.2% and 120.1% ± 7.0, respectively. Multiple stepwise regression analysis revealed that vVO2peak, RE and the speed that the 0–1100 m was completed relative to vVO2peak (adjusted r2 = 0.72, P < 0.0001) explained the most variation in 1500FAST performance time in males (Fig. 1). In female runners, vVO2peak and RE best explained the variance in 1500FAST performance time (adjusted r2 = 0.80, P < 0.001). The ASR was associated with slower 1500FAST performance times in male runners (r2 = 0.25, P = 0.02) and female runners (r2 = 0.89, P < 0.001), while MSS was also associated with slower times in females (r2 = 0.76, P < 0.001). In male runners, the variance in 1500FAST last lap time was best explained by a two-parameter model including vVO2peak and RE (adjusted r2 = 0.63, P < 0.0001). In female runners, vVO2peak and RE also provided the best model (adjusted r2 = 0.53, P = 0.02), while ASR was associated with slower times.

Determinants of maximal 1500-m time trial performance time (1500FAST; panel a: male, panel b: female) and 1500FAST last lap time (panel c: male, panel d: female). Bars indicate the magnitude of explained variance (% r2) obtained from Pearson’s correlations. Black bars indicate determinants associated with faster times, while grey bars indicate variables that are associated with slower times. Closed bars indicate a significance (P < 0.05). Multiple stepwise regression analysis revealed a combination of predictors, which are presented at the top of each panel
1500MOD
The mean speed of the 0–1100 m of 1500MOD relative to vVO2peak and CS was 97.8% ± 3.5% and 116.1% ± 7.0, respectively. Multiple stepwise regression analysis for the determinants of last lap time in 1500MOD in male runners revealed that vVO2peak, MAOD and gastrocnemius carnosine Z-score provided the best model (adjusted r2 = 0.56, P < 0.0001; Fig. 2a–d), while in female runners, vVO2peak and MAOD were associated with the best model (adjusted r2 = 0.75, P < 0.001), but gastrocnemius carnosine Z-score did not further improve the model (P = 0.184). The ASR and the mean speed of the 0–1100 m relative to vVO2peak (both P > 0.05) were associated with a slower 1500MOD last lap in female runners. When the improvement in 1500MOD last lap time was expressed relative to 1500FAST last lap time, gastrocnemius carnosine Z-score and MAOD provided the best model in male runners (adjusted r2 = 0.60, P = 0.017), whereas in female runners, gastrocnemius carnosine Z-score and supramaximal TTE provided the best model (adjusted r2 = 0.66, P = 0.026).

Determinants of last lap time in the moderately paced trial (1500MOD; panel a: male, panel b: female) and the improvement in 1500MOD lap time relative to 1500FAST (panel c: male, panel d: female). Bars indicate the magnitude of explained variance (% r2) obtained from Pearson’s correlations. Black bars indicate determinants associated with faster times, while grey bars indicate variables that are associated with slower times. Closed bars indicate a significance (P < 0.05). Multiple stepwise regression analysis revealed a combination of predictors, which are presented at the top of each panel
1500SLOW
The mean speed of the 0–1100 m of 1500SLOW mean speed relative to vVO2peak and CS was 92.7% ± 3.3% and 110.1% ± 6.7, respectively. Multiple stepwise regression analysis for the determinants of last lap time in 1500SLOW in male runners revealed that vVO2peak, gastrocnemius carnosine Z-score and MAOD provided the best model (adjusted r2 = 0.63, P < 0.001; Fig. 3a–d) which was also similar in female runners (adjusted r2 = 0.80, P < 0.0001). When the improvement in 1500SLOW last lap time was expressed relative to 1500FAST last lap time, gastrocnemius carnosine Z-score and MAOD provided the best model in male runners (adjusted r2 = 0.76, P = 0.017), whereas in female runners, gastrocnemius carnosine Z-score and MAOD provided the best model (adjusted r2 = 0.81, P < 0.001).

Determinants of last lap time in the slow-paced trial (1500SLOW; panel a: male, panel b: female) and the improvement in 1500SLOW lap time relative to 1500FAST (panel c: male, panel d: female). Bars indicate the magnitude of explained variance (% r2) obtained from Pearson’s correlations. Black bars indicate determinants associated with faster times, while grey bars indicate variables that are associated with slower times. Closed bars indicate a significance (P < 0.05). Multiple stepwise regression analysis revealed a combination of predictors, which are presented at the top of each panel
Muscle fibre typology
The middle-distance runners included in the present study predominantly had negative gastrocnemius carnosine Z-score values (28/32 runners; Fig. 4). Figure 5 shows the association between gastrocnemius carnosine Z-score and RE at 110% GET in both male (r2 = 0.39, P < 0.01) and female runners (r2 = 0.46, P = 0.02). This association was comparable for all running speeds that were tested (85–110% GET), whereby r2 values ranged 0.35–0.47. All runners had a blood lactate concentration of ≤ 2.1 mM suggesting that energy contribution of anaerobic metabolism was minimal. Gastrocnemius carnosine Z-score was also associated with MSS in both male (r2 = 0.23, P = 0.02) and female runners (r2 = 0.35, P = 0.05), CS in male runners (r2 = 0.24, P = 0.02), as well as supramaximal TTE (r2 = 0.22, P < 0.01; male runners and female runners combined). There was no significant association between gastrocnemius carnosine Z-score and vVO2peak.

Individual and mean (95%CI) carnosine Z-score values of the gastrocnemius of runners in the present study, as well as the non-athlete control population. The absolute carnosine concentration for the runners was converted to a sex- specific Z-score relative to an age-matched control population of active, healthy non-athletes (males: n = 40; females: n = 33)

Relationship between muscle fibre typology (carnosine Z-score) and the energy cost of running at 110% of the gas exchange threshold in male (a) and female (b) subjects
Discussion
Novel findings from the present study show that the improvements in last lap time in the paced trials were largely explained by MAOD and muscle fibre typology in both male and female runners. That is, runners with a higher gastrocnemius carnosine Z-score (i.e., higher estimated percentage of type II fibres) and greater MAOD, reduced their last lap time to a greater extent in the paced 1500-m trials. Furthermore, a lower carnosine Z-score (i.e., estimated higher percentage of type I fibres) was associated with superior RE (lower energy cost). We also confirm that RE and VVO2peak largely explain the variation in maximal 1500-m time trial time, while VVO2peak is also a key determinant of last lap time in the paced trials. These findings suggest that middle-distance runners who possess diverse physiological profiles may be best suited to adopting polarized racing strategies to have the best chance of winning. The extent to which the diversity in the athlete profiles relate to differences in training characteristics and subsequent adaptations or inherent differences in genetic predisposition requires further examination.
RE and vVO2peak were the most deterministic characteristics of 1500FAST running performance which agrees with previous studies (Ingham et al. 2008; Sandford et al. 2019c). The mean speed of the 1500FAST trial was ~ 101% of vVO2peak, which is slightly lower than previous reports (~ 105–115% vVO2peak) (Billat et al. 2009; Sandford et al. 2019c; Billat 2001), but this can be explained by the faster times achieved in competition racing against competitors (Billat et al. 2009; Sandford et al. 2019c; Billat 2001) compared to completing individual time trials (i.e., 1500FAST), as well as differences in the methodology used to determine vVO2peak. We observed that a greater ASR was associated with slower 1500FAST performance times. These findings also agree with previous research (Sandford et al. 2019c), which was conducted in senior and junior national and international level middle-distance runners, whereby the magnitude of ASR was associated with slower 1500-m running velocity (r = − 0.68 ± 0.54) (Sandford et al. 2019c). Given that vVO2peak was a key determinant of 1500-m performance time in the present study and in previous research (Sandford et al. 2019c), it is likely that the association between ASR and slower 1500-m performance times manifests from a lower vVO2peak, rather than greater MSS. For example, if one athlete has a larger ASR compared to another athlete, but both are matched for MSS, the athlete with the larger ASR would simply have a lower vVO2peak without possessing superior mechanical speed capabilities.
In the present study, vVO2peak was also a key determinant of the last lap time in the paced trials (i.e., 1500MOD and 1500SLOW), in addition to MAOD. These findings are consistent with the supposition that 1500-m racing is likely to elicit a maximum rate of aerobic metabolism and exhaust the finite capacity of the anaerobic energy systems (Billat et al. 2009; Ward-Smith 1999). Apart from 1500MOD in female runners, gastrocnemius carnosine Z-score and MAOD significantly contributed to the regression models explaining some of the variation associated with last lap time and the improvement in last lap time in the paced trials. That is, runners with a higher carnosine Z-score (i.e., higher estimated percentage of type II fibres) and greater MAOD, reduced their last lap time to a greater extent in the paced 1500-m trials. This indicates an advantage for 1500-m runners who possesses a higher proportion of type II fibres, given that these fibres not only underpin speed and power performance (Tesch et al. 1989; Pette 1985; Harber et al. 2004), but also have the ability to adapt to high oxidative demands which are not necessarily subservient to type I fibres in well-trained endurance athletes (Jansson and Kaijser 1977; Ørtenblad et al. 2018). For example, succinate dehydrogenase activity levels have been shown to be similar between type I and II fibres in the gastrocnemius muscle of elite cross-country runners (Jansson and Kaijser 1977), while mitochondrial volume, average capillary-to-fibre ratio and maximal citrate synthase activity are comparable in the highly trained type I and II fibres of elite cross-country skiers (Ørtenblad et al. 2018). In addition to a higher carnosine Z-score, a larger anaerobic capacity (i.e., MAOD) also significantly contributed to the best regression model explaining the variance in absolute last lap time and the relative improvement in the last lap time in the paced trials. The significant contribution of MAOD may have arisen due to the slower relative speed of the 0–1100 m in 1500MOD and 1500SLOW (~ 98% and ~ 93% of vVO2peak and 116% and 110% of CS, respectively), compared to 1500FAST (~ 103% vVO2peak and 121% CS). Given the slower running speeds relative to vVO2peak and CS, it could be expected that that the capacity of anaerobic energy provision, as reflected by the remaining portion of the MAOD, could be preserved to enhance last lap running speed. Previous research in elite middle-distance runners has shown that at any point within a 1500-m time trial, running speed is controlled by the remaining anaerobic energy store (defined as the difference between end-race and instantaneous oxygen deficits) (Billat et al. 2009). Thus, in a group of athletes with a relatively homogenous vVO2peak and CS, preservation of MAOD during the initial laps of a 1500-m race is likely to result in a greater last lap performance improvement for athletes with a larger MAOD.
It has been suggested that a greater ASR may be an important determinant of a fast last lap in a tactical, slow 1500-m race (Sandford et al. 2019b; Mytton et al. 2015). This supposition may be based on the premise that if the speed of the initial laps of a 1500-m race are slower, an athlete with a larger ASR may be able to utilize their superior speed reserve to run a faster last lap than their competitors (Sandford et al. 2019b; Mytton et al. 2015). Despite this, we found little evidence that ASR or MSS was associated with the last lap time in paced trials or the relative improvement in last lap time relative to 1500FAST given that these values did not significantly contribute to the regression models explaining the variation in performance in the paced trials. While MSS has previously been associated with 800-m time trial performance (Bachero-Mena et al. 2017), it is unlikely that athletes would reach running speeds during 1500-m races that are close to their MSS, thus reducing the importance of MSS for 1500-m running performance. For example, the fastest mean 100 m sector for male 800-m championship medallists is ~ 8.9 m s−1, whereas 1500-m championship medallists only reach mean 100 m sector speeds of ~ 7.0 m s−1 (Hanley et al. 2019). In relation to ASR, it is unlikely that this construct represents an anaerobic energy reserve given that its upper boundary (i.e., MSS) is more likely to be limited from a mechanical standpoint rather than an energetics perspective (Bundle and Weyand 2012). The lower boundary of the ASR, demarcated by vVO2peak is thought to represent the minimal intensity that would elicit VO2peak (Morgan et al. 1989) and certainly does not represent a threshold above which energy metabolism is solely accounted for by anaerobic metabolism. As such, the magnitude of ASR does not necessarily represent a surrogate measure of anaerobic capacity which has been previously proposed (Bundle et al. 2003). Indeed, we did not find a strong association between MAOD and ASR in the present study. It is plausible that the importance of ASR for the last lap speed in slower paced 1500-m trials may be related to the running speed relative to vVO2peak (or CS) during the initial laps (i.e., 0–1100 m), but we found no such association. A classical study (Blondel et al. 2001) showed that time to exhaustion at 120% and 140% of vVO2peak and the velocity expressed relative to the ASR had a high correlation (r = − 0.83, P < 0.01 and r = − 0.94, both P < 0.001), suggesting that ASR may be a meaningful parameter when the intensity of exercise is supramaximal. However, the trials conducted in this study (Blondel et al. 2001) are in stark contrast to the 1500-m trials of the present study, whereby the supramaximal last lap speed is preceded by relatively high energetic requirements of the 0–1100 m. As such, in the context of our study, we did not find any substantial evidence that the ASR substantially contributes to last lap speed, or the improvement in last lap speed, in paced 1500-m trials in a group of trained 1500-m runners. It is likely that ASR may be a more important determinant of 800-m running performance, whereby the duration (122 ± 27 s and 65 ± 17 s, respectively) and running intensity (120% and 140% of vVO2peak) of the supramaximal trials featured in Blondel et al. (2001) more closely reflect 800-m running duration and intensity (Billat et al. 2009; Sandford et al. 2019c; Billat 2001). This would also be in accordance with previous research showing a strong relationship between 800-m performance times and ASR in a group of ten elite 800-m runners (800-m personal best ≤ 1:47.50) (Sandford et al. 2018).
Contrary to how D’ may theoretically contribute to last lap performance during paced trials (Pettitt 2016), we did not find that D′ was associated with the relative improvement or absolute last lap time during the paced trials. Furthermore, we did not find a strong association between MAOD and D’. This may seem surprising given that some investigators consider the magnitude of D’ (Gaesser and Poole 1996; Hill and Smith 1993) to be synonymous with anaerobic capacity. However, there is still a great deal of uncertainty regarding the physiological determinants of D’ (Vanhatalo et al. 2010). It has recently been demonstrated that W’ during knee extension exercise, which is synonymous with D’ during running, is reduced with the inspiration of hyperoxic gas (Vanhatalo et al. 2010), whereby the reduction in W’ was inversely associated with the increase in critical power in hyperoxic conditions. These findings (Vanhatalo et al. 2010) suggest that W′/D’ may not simply reflect an anaerobic energy store given that a particular measure of anaerobic capacity should be O2-independent and insensitive to interventions which alter muscle O2 delivery. Other research shows that the magnitude of the VO2 slow component (Murgatroyd et al. 2011) and mental fatigue (Salam et al. 2018) may influence W’, indicating that the mechanistic basis of W’/D’ is quite complex, and the compatibility of this parameter as a measure of anaerobic capacity seems unreasonable. Nonetheless, in the context of the present study, D’ did not relate to any aspect of performance in maximal or paced 1500-m trials.
An interesting finding from the present study was that gastrocnemius carnosine Z-score was positively correlated with RE, whereby subjects with a lower gastrocnemius carnosine Z-score (higher estimated proportion of type I fibres) were more economical (i.e., have a lower energetic cost of running) while running at speeds close to the GET (85–110%). This finding agrees with some (Bosco et al. 1987), but not all research (Kyrolainen et al. 2003) that has investigated the relationship between muscle fibre typology and RE in highly trained runners. However, the purported inverse relationship between the proportion of type II fibres and RE (Kyrolainen et al. 2003) likely manifested from the slower VO2 on-kinetics associated with type II fibres (Pringle et al. 2003) given that the running speed (~ 25.2 km h−1) and duration (1 min) employed were not suitable to achieve a steady state which is a necessary requirement for the valid measurement of RE. In contrast, we assessed subjects running at a relative running speed equivalent to 110% GET (~ 16.8 km h−1), whereby all blood lactate concentration values were ≤ 2.0 mmol L−1. While the relevance of this submaximal speed to 1500-m race pace could be questioned, the validity of RE measurements is dependent upon steady state submaximal running speeds. The superior RE in subjects with slow-twitch muscle typology in the present study could possibly be related to the higher muscular efficiency (i.e., ATP formation per oxygen used) of type I fibres, but is more likely related to the greater contractile efficiency (i.e., work per ATP usage) of type I fibres (Bottinelli and Reggiani 2000).
The aim of the present study was to provide more insight to the underpinning physiological and performance characteristics that dictate last lap speed in tactical 1500-m races. It should be acknowledged that all 1500-m races are somewhat unique, and it is impossible to truly replicate tactical 1500-m scenarios. In the present study, the 0–1100 m in 1500MOD and 1500SLOW were paced relative to 1500FAST and did not include additional mid race surges that seem to be a characteristic of tactical 1500-m races (Sandford et al. 2019a). Furthermore, other factors, such as drafting, re-positioning and running wide on bends or close to the rail, would also influence the physiological requirements of a given race (Sandford et al. 2019a; Jones and Whipp 2002), and subsequently, last lap speed. Lastly, the results of the present study should be contextualised in light of the performance level of the subjects that participated. As is often the case in experimental studies, we recruited trained, but not elite 1500-m runners. Whether similar findings would be present in a group of elite 1500-m runners required further evaluation.
Perspective
The findings from the present study suggest that the diversity in the physiological profile of middle-distance runners could inform a preference for different racing strategies. While the majority of this diversity may be due to training characteristics and subsequent training adaptations, we highlight that muscle fibre typology explains a large degree of the variation in RE which is a key determinant of maximal 1500-m performance, but also underpins last lap speed in tactical 1500-m racing. Importantly, muscle fibre typology is largely dependent on genetic predisposition and is not thought to change substantially in response to training (Ingalls 2004). As such, there is a complex interplay between how much training induced adaptations and genetic predisposition contributes to the diversity in the physiological profiles of middle-distance runners. Coaches should continue to maximise aerobic characteristics (i.e., RE and vVO2peak) through training as these were important determinants of both maximal and tactical 1500-m racing. Once a given level of aerobic capability has been acquired, other characteristics, such as anaerobic capacity and higher proportions of type II muscle fibres, may be important for last lap speed, particularly in slower paced races. As such, developing anaerobic capacity and speed endurance should not be overlooked. While athletes with lower gastrocnemius carnosine Z-scores (i.e., higher estimated proportion of type I fibres) have the advantage of better RE and CS (at least in male runners), athletes with higher gastrocnemius carnosine Z-scores (i.e., higher estimated proportion of type II fibres) may possess the benefits of superior speed endurance and greater capacity for anaerobic energy provision. It could be suggested that runners with a lower gastrocnemius carnosine Z-score may be best suited to a racing strategy characterized by a sustained pace from the start to take advantage of superior RE, while runners with a higher gastrocnemius carnosine Z-score may be best suited to controlling the pace of the race in the initial laps, to take advantage of the superior last lap speed.
Abbreviations
- RE:
-
Running economy
- VVO2peak :
-
Minimal velocity that elicits VO2peak
- ASR:
-
Anaerobic speed reserve
- MSS:
-
Maximal sprinting speed
- CS:
-
Critical speed
- D’:
-
Curvature constant
- VO2peak :
-
Peak oxygen uptake
- 1500FAST :
-
Maximal 1500-m time trial
- 1500MOD :
-
Moderately paced 1500-m time trial
- 1500SLOW :
-
Slow-paced 1500-m time trial
- MAOD:
-
Maximal accumulated oxygen deficit
- d :
-
Distance run
- t:
-
Time
- CV:
-
Coefficient of variation
- VO2 :
-
Oxygen uptake
- GET:
-
Gas exchange threshold
- TTE:
-
Time to exhaustion
- 1H-MRS:
-
Proton magnetic resonance spectroscopy
- MRI:
-
Magnetic resonance imaging
- SD:
-
Standard deviation
References
Bachero-Mena B, Pareja-Blanco F, Rodríguez-Rosell D, Yáñez-García JM, Mora-Custodio R, González-Badillo JJ (2017) Relationships between sprint, jumping and strength abilities, and 800 m performance in male athletes of national and international levels. J Hum Kinet 58(1):187–195. https://doi.org/10.1515/hukin-2017-0076
Baguet A, Everaert I, Hespel P, Petrovic M, Achten E, Derave W (2011) A new method for non-invasive estimation of human muscle fiber type composition. PLoS ONE 6(7):e21956. https://doi.org/10.1371/journal.pone.0021956
Billat LV (2001) Interval training for performance: a scientific and empirical practice. Sports Med 31(1):13–31. https://doi.org/10.2165/00007256-200131010-00002
Billat V, Beillot J, Jan J, Rochcongar P, Carre F (1996) Gender effect on the relationship of time limit at 100% VO2max with other bioenergetic characteristics. Med Sci Sports Exerc 28(8):1049–1055. https://doi.org/10.1097/00005768-199608000-00016
Billat V, Hamard L, Koralsztein JP, Morton RH (2009) Differential modeling of anaerobic and aerobic metabolism in the 800-m and 1,500-m run. J Appl Physiol 107(2):478–487. https://doi.org/10.1152/japplphysiol.91296.2008
Blondel N, Berthoin S, Billat V, Lensel G (2001) Relationship between run times to exhaustion at 90, 100, 120, and 140% of vVO2max and velocity expressed relatively to critical velocity and maximal velocity. Int J Sports Med 22(1):27–33. https://doi.org/10.1055/s-2001-11357
Bosco C, Montanari G, Ribacchi R, Giovenali P, Latteri F, Iachelli G, Faina M, Colli R, Dal Monte A, La Rosa M, Cortili G, Saibene F (1987) Relationship between the efficiency of muscular work during jumping and the energetics of running. Eur J Appl Physiol Occup Physiol 56(2):138–143. https://doi.org/10.1007/BF00640636
Bottinelli R, Reggiani C (2000) Human skeletal muscle fibres: molecular and functional diversity. Prog Biophys Mol Biol 73(2):195–262. https://doi.org/10.1016/S0079-6107(00)00006-7
Bundle MW, Weyand PG (2012) Sprint exercise performance: does metabolic power matter? Exerc Sport Sci Rev 40(3):174–182. https://doi.org/10.1097/JES.0b013e318258e1c1
Bundle MW, Hoyt RW, Weyand PG (2003) High-speed running performance: a new approach to assessment and prediction. J Appl Physiol 95(5):1955–1962. https://doi.org/10.1152/japplphysiol.00921.2002
Casado A, Renfree A (2018) Fortune favors the brave: tactical behaviors in the middle-distance running events at the 2017 IAAF World Championships. Int J Sports Physiol Perform 13(10):1386–1391. https://doi.org/10.1123/ijspp.2018-0055
Costill DL, Daniels J, Evans W, Fink W, Krahenbuhl G, Saltin B (1976) Skeletal muscle enzymes and fiber composition in male and female track athletes. J Appl Physiol 40(2):149–154. https://doi.org/10.1152/jappl.1976.40.2.149
Duffield R, Dawson B, Goodman C (2005) Energy system contribution to 1500- and 3000-metre track running. J Sports Sci 23(10):993–1002. https://doi.org/10.1080/02640410400021963
Ferri A, Adamo S, La Torre A, Marzorati M, Bishop DJ, Miserocchi G (2012) Determinants of performance in 1,500-m runners. Eur J Appl Physiol 112(8):3033–3043. https://doi.org/10.1007/s00421-011-2251-2
Fletcher JR, Esau SP, MacIntosh BR (2009) Economy of running: beyond the measurement of oxygen uptake. J Appl Physiol 107(6):1918–1922. https://doi.org/10.1152/japplphysiol.00307.2009
Gaesser GA, Poole DC (1996) The slow component of oxygen uptake kinetics in humans. Exerc Sport Sci Rev 24(1):35–70
Hanley B, Stellingwerff T, Hettinga FJ (2019) Successful pacing profiles of Olympic and IAAF World Championship middle-distance runners across qualifying rounds and finals. Int J Sports Physiol Perform 14(7):894–901. https://doi.org/10.1123/ijspp.2018-0742
Harber MP, Gallagher PM, Creer AR, Minchev KM, Trappe SW (2004) Single muscle fiber contractile properties during a competitive season in male runners. Am J Physiol Regul Integr Comp Physiol 287(5):R1124–R1131. https://doi.org/10.1152/ajpregu.00686.2003
Hill DW, Smith JC (1993) A comparison of methods of estimating anaerobic work capacity. Ergonomics 36(12):1495–1500. https://doi.org/10.1080/00140139308968017
Hughson R, Orok C, Staudt L (1984) A high velocity treadmill running test to assess endurance running potential. Int J Sports Med 5(01):23–25. https://doi.org/10.1055/s-2008-1025875
Ingalls CP (2004) Nature vs. nurture: can exercise really alter fiber type composition in human skeletal muscle? J Appl Physiol 97(5):1591–1592. https://doi.org/10.1152/classicessays.00010.2004
Ingham SA, Whyte GP, Pedlar C, Bailey DM, Dunman N, Nevill AM (2008) Determinants of 800-m and 1500-m running performance using allometric models. Med Sci Sport Exerc 40(2):345–350. https://doi.org/10.1249/mss.0b013e31815a83dc
Iwaoka K, Hatta H, Atomi Y, Miyashita M (1988) Lactate, respiratory compensation thresholds, and distance running performance in runners of both sexes. Int J Sports Med 09(05):306–309. https://doi.org/10.1055/s-2007-1025028
Jansson E, Kaijser L (1977) Muscle adaptation to extreme endurance training in man. Acta Physiol Scand 100(3):315–324. https://doi.org/10.1111/j.1748-1716.1977.tb05956.x
Jones AM, Doust JH (1996) A 1% treadmill grade most accurately reflects the energetic cost of outdoor running. J Sports Sci 14(4):321–327. https://doi.org/10.1080/02640419608727717
Jones AM, Whipp BJ (2002) Bioenergetic constraints on tactical decision making in middle distance running. Br J Sports Med 36(2):102–104. https://doi.org/10.1136/bjsm.36.2.102
Kyrolainen H, Kivela R, Koskinen S, McBride J, Andersen JL, Takala T, Sipila S, Komi PV (2003) Interrelationships between muscle structure, muscle strength, and running economy. Med Sci Sports Exerc 35(1):45–49. https://doi.org/10.1249/01.mss.0000046149.03322.bb
MacMillan EL, Bolliger CS, Boesch C, Kreis R (2016) Influence of muscle fiber orientation on water and metabolite relaxation times, magnetization transfer, and visibility in human skeletal muscle. Magn Reson Med 75(4):1764–1770. https://doi.org/10.1002/mrm.25778
Medbo JI, Mohn AC, Tabata I, Bahr R, Vaage O, Sejersted OM (1988) Anaerobic capacity determined by maximal accumulated O2 deficit. J Appl Physiol 64(1):50–60. https://doi.org/10.1152/jappl.1988.64.1.50
Monod H, Scherrer J (1965) The work capacity of a synergic muscular group. Ergonomics 8(3):329–338. https://doi.org/10.1080/00140136508930810
Morgan DW, Baldini FD, Martin PE, Kohrt WM (1989) Ten kilometer performance and predicted velocity at VO2max among well-trained male runners. Med Sci Sports Exerc 21(1):78–83. https://doi.org/10.1249/00005768-198902000-00014
Murgatroyd SR, Ferguson C, Ward SA, Whipp BJ, Rossiter HB (2011) Pulmonary O2 uptake kinetics as a determinant of high-intensity exercise tolerance in humans. J Appl Physiol 110(6):1598–1606. https://doi.org/10.1152/japplphysiol.01092.2010
Mytton GJ, Archer DT, Turner L, Skorski S, Renfree A, Thompson KG, Gibson ASC (2015) Increased variability of lap speeds: differentiating medalists and nonmedalists in middle-distance running and swimming events. Int J Sports Physiol Perform 10(3):369–373. https://doi.org/10.1123/ijspp.2014-0207
Ørtenblad N, Nielsen J, Boushel R, Söderlund K, Saltin B, Holmberg H-C (2018) The muscle fiber profiles, mitochondrial content, and enzyme activities of the exceptionally well-trained arm and leg muscles of elite cross-country skiers. Front Physiol. https://doi.org/10.3389/fphys.2018.01031
Parkhouse WS, McKenzie DC, Hochachka PW, Ovalle WK (1985) Buffering capacity of deproteinized human vastus lateralis muscle. J Appl Physiol 58(1):14–17. https://doi.org/10.1152/jappl.1985.58.1.14
Peronnet F, Massicotte D (1991) Table of nonprotein respiratory quotient: an update. Can J Sport Sci 16(1):23–29
Pette D (1985) Metabolic heterogeneity of muscle fibres. J Exp Biol 115(1):179–189
Pettitt RW (2016) Applying the critical speed concept to racing strategy and interval training prescription. Int J Sports Physiol Perform 11(7):842–847. https://doi.org/10.1123/ijspp.2016-0001
Pringle JSM, Doust JH, Carter H, Tolfrey K, Campbell IT, Jones AM (2003) Oxygen uptake kinetics during moderate, heavy and severe intensity “submaximal” exercise in humans: the influence of muscle fibre type and capillarisation. Eur J Appl Physiol 89(3):289–300. https://doi.org/10.1007/s00421-003-0799-1
Salam H, Marcora SM, Hopker JG (2018) The effect of mental fatigue on critical power during cycling exercise. Eur J Appl Physiol 118(1):85–92. https://doi.org/10.1007/s00421-017-3747-1
Sandford GN, Allen SV, Kilding AE, Ross A, Laursen PB (2018) Anaerobic speed reserve: a key component of elite male 800m running. Int J Sports Physiol Perform 14(4):501–508. https://doi.org/10.1123/ijspp.2018-0163
Sandford GN, Day BT, Rogers SA (2019a) Racing fast and slow: defining the tactical behavior that differentiates medalists in elite men’s 1,500 m championship racing. Front Sports Act Living. https://doi.org/10.3389/fspor.2019.00043
Sandford GN, Kilding AE, Ross A, Laursen PB (2019b) Maximal sprint speed and the anaerobic speed reserve domain: the untapped tools that differentiate the world’s best male 800 m runners. Sports Med 49(6):843–852. https://doi.org/10.1007/s40279-018-1010-5
Sandford GN, Rogers SA, Sharma AP, Kilding AE, Ross A, Laursen PB (2019c) Implementing anaerobic speed reserve testing in the field: validation of vVO2max prediction from 1500-m race performance in elite middle-distance runners. Int J Sports Physiol Perform 14(8):1147–1150. https://doi.org/10.1123/ijspp.2018-0553
Schneider DA, Phillips SE, Stoffolano S (1993) The simplified V-slope method of detecting the gas exchange threshold. Med Sci Sports Exerc 25(10):1180–1184
Spencer MR, Gastin PB (2001) Energy system contribution during 200-to 1500-m running in highly trained athletes. Med Sci Sport Exerc 33(1):157–162. https://doi.org/10.1097/00005768-200101000-00024
Tesch PA, Thorsson A, Fujitsuka N (1989) Creatine phosphate in fiber types of skeletal muscle before and after exhaustive exercise. J Appl Physiol 66(4):1756–1759. https://doi.org/10.1152/jappl.1989.66.4.1756
Tsuji K, Ishida H, Oba K, Ueki T, Fujihashi Y (2015) Activity of lower limb muscles during treadmill running at different velocities. J Phys Ther Sci 27(2):353–356. https://doi.org/10.1589/jpts.27.353
Tucker R, Lambert MI, Noakes TD (2006) An analysis of pacing strategies during men’s world-record performances in track athletics. Int J Sports Physiol Perform 1(3):233–245. https://doi.org/10.1123/ijspp.1.3.233
Vanhatalo A, Fulford J, DiMenna FJ, Jones AM (2010) Influence of hyperoxia on muscle metabolic responses and the power–duration relationship during severe-intensity exercise in humans: a 31P magnetic resonance spectroscopy study. Exp Physiol 95(4):528–540. https://doi.org/10.1113/expphysiol.2009.050500
Ward-Smith AJ (1999) The bioenergetics of optimal performances in middle-distance and long-distance track running. J Biomech 32(5):461–465. https://doi.org/10.1016/S0021-9290(99)00029-9
Author information
Authors and Affiliations
Contributions
PB, WD, EL, BA and CM conceived and designed research. PB, BK, HR and BA conducted experiments. WD and EL analysed 1H-MRS data. PB analysed training and performance data. PB wrote the manuscript. All authors read and approved the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that we received internal funding from Griffith University which supported MR related costs for the data collection. The authors have no competing financial or non-financial interests to disclose. The results of the present study are presented clearly, honestly, and without fabrication.
Additional information
Communicated by Philip D. Chilibeck.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Bellinger, P., Derave, W., Lievens, E. et al. Determinants of last lap speed in paced and maximal 1500-m time trials. Eur J Appl Physiol 121, 525–537 (2021). https://doi.org/10.1007/s00421-020-04543-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-020-04543-x