
Abstract
Purpose
The purpose of the present study was to examine the effects of unilateral fatigue of the knee extensors at different movement velocities on neuromuscular performance in the fatigued and non-fatigued leg.
Methods
Unilateral fatigue of the knee extensors was induced in 11 healthy young men (23.7 ± 3.8 years) at slower (60°/s; FAT60) and faster movement velocities (240°/s; FAT240) using an isokinetic dynamometer. A resting control (CON) condition was included. The fatigue protocols consisted of five sets of 15 maximal concentric knee extensions using the dominant leg. Before and after fatigue, peak isokinetic torque (PIT) and time to PIT (TTP) of the knee extensors as well as electromyographic (EMG) activity of vastus medialis, vastus lateralis, and biceps femoris muscles were assessed at 60 and 240°/s movement velocities in the fatigued and non-fatigued leg.
Results
In the fatigued leg, significantly greater PIT decrements were observed following FAT60 and FAT240 (11–19%) compared to CON (3–4%, p = .002, d = 2.3). Further, EMG activity increased in vastus lateralis and biceps femoris muscle following FAT240 only (8–28%, 0.018 ≤ p ≤ .024, d = 1.8). In the non-fatigued leg, shorter TTP values were found after the FAT60 protocol (11–15%, p = .023, d = 2.4). No significant changes were found for EMG data in the non-fatigued leg.
Conclusion
The present study revealed that both slower and faster velocity fatiguing contractions failed to show any evidence of cross-over fatigue on PIT. However, unilateral knee extensor fatigue protocols conducted at slower movement velocities (i.e., 60°/s) appear to modulate torque production on the non-fatigued side (evident in shorter TTP values).
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Many training and rehabilitation programs include exercises that require prolonged or repetitive unilateral muscle actions (e.g., single leg squats, one arm biceps curls). Thus, unilateral resistance training has been recommended as it incorporates greater core or trunk muscle activation (Behm et al. 2005) and overcomes voluntary force deficits evident during bilateral contraction (Van Dieen et al. 2003). An additional benefit is that training one side of the body has been shown to enhance muscle strength of the contralateral homologous muscles, a phenomenon which is known as cross-education (Hortobagyi et al. 1997; Carroll et al. 2006). In fact, a meta-analysis by Munn et al. (2004) indicated that unilateral resistance training produces moderate improvements (7.8%) in contralateral strength in healthy, young individuals. However, there is evidence that unilateral fatiguing contractions would lead to a transient modulation of neural activation patterns and motor performance in the non-fatigued, contralateral homologous (Todd et al. 2003; Kawamoto et al. 2014) and heterologous (Kennedy et al. 2013; Sidhu et al. 2014; Sambaher et al. 2016) muscle groups.
For instance, Kawamoto et al. (2014) assessed the effect of four sets of unilateral dynamic knee extensions at 40 or 70% of maximal isometric voluntary contractions and showed moderate-to-large decrements in force output (4–7%) of the non-fatigued contralateral homologous muscles. Indeed, unilateral contraction can modulate the responsiveness of corticomotor pathways innervating the non-exercised limbs (Aboodarda et al. 2015, 2016). However, contrary to the concept of non-local muscle fatigue, several lines of evidence indicate that dynamic fatiguing contractions show only minimal effects on contralateral muscle force output (Grabiner and Owings 1999; Regueme et al. 2007; Berger et al. 2010; Amann et al. 2013). Accordingly, Halperin et al. (2015) suggested that the cross-over fatigue effects appear to be dependent on several factors such as the affected muscle group, fatigue protocol or the testing protocol for the non-fatigued muscle groups. For instance, greater cross-over fatigue effects were observed in fatigue protocols using higher compared to lower intensities or isometric compared to dynamic muscle actions (Halperin et al. 2015).
In terms of muscle action type, it has to be noted that the majority of the studies on dynamic fatiguing muscle actions have assessed force output in the rested muscles using isometric contractions. However, it can be argued that isometric contractions are not pertinent to examine neuromuscular fatigue associated with dynamic movements involved in daily life activities as well as training programs. Therefore, further research is required to investigate the effect of unilateral dynamic fatiguing contractions on contralateral muscle performance evaluated during different movement velocities. In fact, the specific movement velocity during dynamic muscle actions may influence cross-over motor responses, given that this variable is known to affect force/torque output (Thorstensson et al. 1976; Babault et al. 2002), neural drive (Babault et al. 2002; Morel et al. 2015), and metabolic stress (Douris 1993; Morel et al. 2015). For instance, Babault et al. (2002) found that voluntary activation (as determined by twitch interpolation technique) was significantly lower in the knee extensors during maximal concentric knee extensions at angular velocities of 60°/s compared to 120°/s. Moreover, Morel et al. (2015) examined the effects of movement velocity during unilateral concentric fatiguing knee extensions on neuromuscular performance measures. The authors reported that force decrements and accumulated metabolites within the muscle were significantly larger following a fatigue protocol of the knee extensors when performed at 240°/s compared to 30°/s, whereas decrements in voluntary activation of knee extensor muscles were significantly greater following fatiguing knee extensions at 30°/s (Morel et al. 2015). Of note, alterations in the neural drive to the muscles have been suggested to account for some of the observed cross-over modulation in motor task performance (Todd et al. 2003; Halperin et al. 2015). Consequently, it could be speculated that the neural stress associated with slower versus faster movement velocities may have larger impact upon the cross-over responses in torque production. On the other hand, research on training programs using different movement velocities during exercises has demonstrated that the greatest adaptations occur at or near the velocities trained (Behm and Sale 1993). According to the principle of training specificity (Behm and Sale 1993), it is of interest whether performance changes in the fatigued and non-fatigued muscle groups are more pronounced if the velocity during testing complies with the velocity during fatiguing dynamic muscle actions. To the best of our knowledge, there is no study available that examined the effects of different movement velocities during fatiguing muscle actions (e.g., knee extensions) on dynamic muscle performance of the fatigued and non-fatigued (homologous) muscle groups.
Thus, the purpose of the present study was to examine the effects of unilateral fatigue of the knee extensors induced at different movement velocities (e.g., 60, 240°/s) on neuromuscular performance (i.e., peak isokinetic torque, time to peak isokinetic torque, muscle activity, and neuromuscular efficiency) of the ipsilateral fatigued and contralateral non-fatigued homologous muscles measured at different velocities (e.g., 60, 240°/s). Based on the aforementioned literature (Behm and Sale 1993; Babault et al. 2002; Morel et al. 2015), we hypothesized that (1) torque production of the exercised knee extensors would be impaired following unilateral muscle fatigue particularly when using faster movement velocities, whereas torque output of the non-exercised knee extensors would be lower following unilateral muscle fatigue particularly when using slower movement velocities, and (2) specific effects of the fatigue protocols are more pronounced in the fatigued and non-fatigued leg if the movement velocity during the test complies with the velocity used during the fatigue protocol.
Methods
Participants
Eleven healthy young men (age: 23.7 ± 3.8 years, height: 182.1 ± 6.0 cm, body mass: 78.0 ± 10.2 kg, body fat: 13.3 ± 3.8%) volunteered to participate in the study. With reference to the study of Morel and colleagues (2015), an a priori power analysis with an assumed Type I error of 0.05 and a Type II error rate of 0.20 (80% statistical power) revealed that this number of participants would be sufficient for finding a statistically significant condition × time interaction in the fatigued leg. All subjects were classified as physically active (physical activity: 11.8 ± 5.5 h/week) according to the Freiburg questionnaire for everyday and sports-related activities (Frey et al. 1999). None had any history of musculoskeletal, neurological, or orthopedic disorder that might have affected their ability to execute the experimental protocol. Written informed consent was obtained before the start of the study. Local ethical permission was given (approval number: 34/2014), and all experiments were conducted according to the latest version of the Declaration of Helsinki.
Experimental procedure
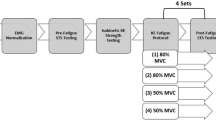
A single-group, randomized cross-over design was used to examine the acute effects of unilateral fatiguing knee extensions on the neuromuscular performance of the contralateral homologous muscle. Participants were required to attend the lab on three different occasions separated by at least 48 h. The three testing sessions included unilateral fatiguing knee extensions at slower angular velocities (i.e., 60°/s), higher angular velocities (i.e., 240°/s), and a control condition (i.e., no intervention) in a randomized order. Fatigue protocols were performed with the dominant leg as determined by the lateral preference inventory (Coren 1993). Each testing session started with locating the surface electrodes followed by a standardized warm-up protocol. Before and immediately after the fatigue protocol, torque and electromyographic (EMG) data were assessed for both the fatigued and non-fatigued leg. For testing and fatigue protocols, a motorized isokinetic dynamometer (Isomed2000, D&R Ferstl GmbH, Hemau, Germany) was used. Acceleration and deceleration of the dynamometer’s lever arm were kept constant for each participant and condition throughout the entire experimental conditions (maximal acceleration/ deceleration: 8500°/s²). To get accustomed to the isokinetic device and the procedures, one familiarization session was conducted on a separate occasion before the start of the study. During familiarization session, subjects’ body height was assessed using a wall-mounted scale. In addition, body mass and body composition (i.e., skeletal muscle mass and body fat) were registered by means of a bioimpedance analysis system (InBody 720, BioSpace, Seoul, Korea).
Fatigue protocols
The fatigue protocols used in this study consisted of 5 sets of 15 maximal unilateral concentric knee extensions on the isokinetic dynamometer either at angular velocities of 60°/s (FAT60) or 240°/s (FAT240). After individual adjustment of the dynamometer, the participants were seated with the hip angle adjusted at 90° and straps attached to the isokinetic device to firmly fix the upper body and the hip. The shank of the dominant leg was attached to the lever arm of the dynamometer to control for knee extension velocity and to record the torque applied by the knee extensors. The range of motion at the knee joint was 0–90° with 0° angle corresponding to full knee extension. Knee flexions were performed passively by having an assistant moving the leg back to the initial position. Further, the participants were instructed to relax their leg muscles during knee flexions. A brief rest of 15 s was allowed between each set. During the control condition (CON), the participants rested on the dynamometer for 7 min.
Measurement of maximal isokinetic knee extension
For all testing sessions, the same isokinetic device and positioning were used as during the fatigue protocols. Prior to pre-tests, participants performed the general and a specific warm-up including 5 min of cycling with 60–70 revolutions/min on an ergometer (100 W) followed by two sets of 10 submaximal unilateral concentric knee extensions (one set at 60°/s and one set at 240°/s) at approximately 50% of peak torque with each leg. During pre-tests, two trials of maximal unilateral concentric knee extensions were conducted each at 60 and 240°/s with the non-fatigued followed by the fatigued leg. The order of test velocity was randomized. A third trial was performed if the difference between the first two trials was >5%. During post-tests, one maximal unilateral concentric knee extension at 60°/s and one at 240°/s in a randomized order were executed with the fatigued as well as non-fatigued leg. In accordance with the fatigue protocol, the range of motion at the knee joint was defined between 0–90°. Due to methodological reasons, the post-tests for the fatigued leg were organized immediately (i.e., 10 s) following set four of the fatigue protocol. By monitoring the extent of fatigue of the exercised leg after set four, we minimized the time for recovery between completion of the fatiguing knee extensions and the contralateral non-fatigued leg post-tests. Given that the dynamometer was moved from the fatigued to the non-fatigued limb following set five, post-tests for the non-fatigued leg were conducted 30 s, 3 and 5 min following set five of the fatigue protocol. A 30-s rest was provided between test trials of one leg. All torque signals were individually corrected for the effect of gravity on the lower leg and finally sampled by the isokinetic device at a frequency of 200 Hz. For later analysis, peak isokinetic torque (PIT), knee joint angle at PIT (on the following referred to as PIT angle), and time to PIT (TTP) were determined. PIT was defined as the maximal value of the torque time curve, whereas TTP was calculated as the time needed to reach PIT from onset of torque. Onset of torque was defined as the time point at which torque development exceeded 2.5% of PIT (Prieske et al. 2014).
Measurement of leg muscle activity
During maximal isokinetic torque measurements, EMG activity of the vastus medialis (VM), vastus lateralis (VL), and biceps femoris (BF) muscles of the fatigued and non-fatigued leg were assessed using circular bipolar surface electrodes (Ambu®, type Blue Sensor P-00-S/50, Ag/AgCl, 13.2 mm, center-to-center distance 25 mm, Ballerup, Denmark). Electrodes were positioned on the muscle bellies according to the European recommendations for surface electromyography (Hermens et al. 1999). Inter-electrode impedance was kept below 5 kΩ by shaving, slightly roughening, degreasing, and disinfecting the skin. The location of the electrodes was marked with permanent ink for identical position on the three occasions. The EMG signals were amplified, transmitted telemetrically (TeleMyo 2400 G2, Noraxon®, Scottsdale, AZ, USA), and finally stored at a sampling frequency of 1,500 Hz. Synchronization of EMG, torque, and angle data was achieved by analog-to-digital conversion of the torque signal (TeleMyo 2400R Analog Output Receiver, Noraxon®, Scottsdale, AZ, USA). For later offline analysis, raw signals were digitally band-pass filtered (10–750 Hz) followed by a moving-root-mean-square filter with a time constant of 50 ms (Prieske et al. 2014) running the MyoResearch XP Master edition software (version 1.08.17, Noraxon®, Scottsdale, AZ, USA). In the present study, EMG was defined as the mean amplitude voltage identified for time intervals ranging from onset to offset of torque. EMG data were normalized to the respective pre-test EMG value in the time interval 50 ms before and after PIT at 240°/s. Additionally, neuromuscular efficiency (NME) was calculated as the ratio of PIT to EMG for VM and VL (Kawamoto et al. 2014). A decrease in NME for instance indicates altered muscle contractile function. In other words, higher EMG activity is needed to produce similar force/torque output (Bigland-Ritchie 1981).
Statistical analyses
Data are presented as group mean values and standard deviations. After normal distribution was examined (i.e., Shapiro–Wilk Test), a 3 (condition: FAT60, FAT240, CON) × 2 (time: pre, post) × 2 (test velocity: 60°/s, 240°/s) analysis of variance (ANOVA) with repeated measures for condition, time, and test velocity was used to analyze absolute torque and EMG parameters in the fatigued leg. In terms of the non-fatigued leg, a 3 (condition: FAT60, FAT240, CON) × 4 (time: pre, post 30 s, post 3 min, post 5 min) × 2 (test velocity: 60°/s, 240°/s) ANOVA with repeated measures for condition, time, and test velocity was conducted. Homogeneity of variance was examined using the Mauchly sphericity test for the repeated measures. The Greenhouse-Geisser correction was applied for further analyses, if homogeneity was violated. Post hoc tests with the Bonferroni-adjusted α were conducted to identify the comparisons that were statistically significant. The significance level was set at p < .05. Additionally, the effect size was calculated by converting partial eta-squared to Cohens d (Cohen 1988) to indicate whether a statistically significant difference is a difference of practical concern. According to Cohen (1988), the magnitude of effect size can be classified as small (0.2 ≤ d < 0.5), medium (0.5 ≤ d < 0.8), and large (d ≥ 0.8). All analyses were performed using Statistical Package for Social Sciences (SPSS) version 23.0 (IBM Corp., Armonk, NY, USA).
Results
Maximal isokinetic knee extension in the fatigued leg
Means and standard deviations of isokinetic performance measures (i.e., PIT, PIT angle, TTP) in the fatigued leg are presented in Table 1. Figure 1 shows representative recordings of maximal unilateral knee extension torque with the exercised leg before and after fatigue. Significant main effects of (1) time (p ≤ .036, 1.5 ≤ d ≤ 3.3), (2) condition (p ≤ .01, 1.5 ≤ d ≤ 2.0), and/or (3) test velocity (p < .001, 3.4 ≤ d ≤ 9.9) were found for PIT, PIT angle, and TTP. Additionally, a significant condition × time interaction occurred for PIT in the fatigued leg (p = .002, d = 2.3). PIT decreased significantly from pre- to post-tests by 11–13% during FAT60 (p = .016, d = 1.8) and by 19% during FAT240 (p < .001, d = 4.9), but only by 3–4% during CON (p = .001, d = 2.9), irrespective of test velocity (Fig. 2a). Moreover, baseline PIT values were not statistically different between conditions. However, post-test PIT values following FAT60 and FAT240 were significantly lower compared to post-test PIT values of CON condition (p ≤ .008, 2.1 ≤ d ≤ 3.9). Additionally, post-test PIT measures of FAT240 condition were significantly lower compared to FAT60 post-test values (p = .032, d = 1.6). Further, significant time × test velocity (0.011 ≤ p ≤ .024, 1.7 ≤ d ≤ 2.0) interactions were identified for PIT angle and TTP in the fatigued leg. Post-hoc analyses showed significantly increased TTP values (7–15%, p = .014, d = 1.9) and decreased PIT angles (i.e., more extended knee joints; 2–6%, p = .013, d = 1.9) from pre to post in the 60°/s test velocity condition, irrespective of condition (Fig. 2b).

Representative recordings of maximal unilateral concentric knee extension torque with the exercised leg at 60°/s before (pre) and after the fatigue protocols (FAT60 fatiguing muscle actions at 60°/s, FAT240 = fatiguing muscle actions at 240°/s, CON control). Note the decrease in torque production particularly after the FAT240 protocol

Changes in a peak isokinetic torque (ΔPIT) and b time to PIT (ΔTTP) during maximal unilateral concentric knee extensions in the fatigued leg at 60°/s and 240°/s following unilateral fatiguing knee extensions. Data are presented as percentages relative to the respective pre-test values. FAT60 fatiguing muscle actions at 60°/s, FAT240 fatiguing muscle actions at 240°/s, CON control. Significantly different for decrements in torque: *p < .05, **p < .01, ***p < .001
Muscle activity in the fatigued leg
Means and standard deviations of EMG measures in the fatigued leg are presented in Table 2. A significant main effect of time was found for NME in VL (p = .018, d = 2.3). In addition, the statistical analysis revealed significant main effects of test velocity for all EMG (0.001 ≤ p ≤ .007, 3.3 ≤ d ≤ 4.3) and NME (p < .001, 5.3 ≤ d ≤ 5.7) measures. Significant condition × time interactions were observed for EMG (i.e., VL, BF) (0.008 ≤ p ≤ .046, 1.8 ≤ d ≤ 2.0) and NME (p ≤ .032, 1.9 ≤ d ≤ 3.0) of the fatigued leg. Subsequent post hoc tests indicated significantly increased muscle activation levels of VL and BF (8–28%, 0.018 ≤ p ≤ .024, d = 1.8) as well as decreased NME measures of VM and VL (17–29 %, p < .001, 4.1 ≤ d ≤ 4.3) following the FAT240 protocol only (Fig. 3). Significant time × test velocity interactions for NME of VM and VL in the fatigued leg (0.002 ≤ p ≤ .041, 1.9 ≤ d ≤ 3.6). Post hoc tests indicated significant pre–post reductions in NME at a test velocity of 60°/s (4–24%, 0.002 ≤ p ≤ .005, 2.4 ≤ d ≤ 2.9), irrespective of the applied fatigue condition (Fig. 4).

Changes in time to peak isokinetic torque (ΔTTP) during maximal unilateral concentric knee extensions in the non-fatigued leg at 60°/s and 240°/s 30 s, 3, and 5 min following unilateral fatiguing knee extensions. Data are presented as percentages relative to the respective pre-test values. FAT60 fatiguing muscle actions at 60°/s, FAT240 fatiguing muscle actions at 240°/s, CON control. Significantly different from pre-test, irrespective of test velocity: # p < .10, *p < .05

Changes in neuromuscular efficiency (NME) of a m. vastus medialis and b m. vastus lateralis during maximal unilateral concentric knee extensions in the fatigued leg at 60 and 240°/s following unilateral fatiguing knee extensions. Data are presented as percentages relative to the respective pre-test values. FAT60 fatiguing muscle actions at 60°/s, FAT240 fatiguing muscle actions at 240°/s, CON control. Significantly different from pre-test, irrespective of test velocity: ***p < .001
Maximal isokinetic knee extension in the non-fatigued leg
Means and standard deviations of isokinetic performance measures (i.e., PIT, PIT angle, TTP) in the non-fatigued leg are presented in Table 3. Significant main effects of time (p < .001, d = 1.8) and/or test velocity (p < .001, 3.4 ≤ d ≤ 8.9) were found for PIT, PIT angle, and TTP. In terms of time effects for PIT, post hoc tests revealed a strong trend for a reduction 30 s after the fatigue protocol (p = .057). In addition, when compared to PIT pre-test values, significant differences were found 3 min after the fatigue protocol (p = .010) (Fig. 5). Additionally, a significant condition × time interaction was observed for TTP (p = .026, d = 1.0). Post hoc tests showed a tendency toward decreased TTP values 30 s after the fatigue protocol (9–15%, p = .062, d = 2.0) and significantly lower TTP values 5 min (11–15%, p = .023, d = 2.4) following the FAT60 protocol when compared to pre-test (Fig. 3).

Changes in peak isokinetic torque (ΔPIT) of maximal unilateral concentric knee extensions in the non-fatigued leg at 60 and 240°/s 30 s, 3, and 5 min following unilateral fatiguing knee extensions. Reductions are presented as percentage values relative to the respective pre-test values. FAT60 fatiguing muscle actions at 60°/s, FAT240 fatiguing muscle actions at 240°/s, CON control. Significantly different from pre-test, irrespective of condition and test velocity: # p < .10, **p < .01
Muscle activity in the non-fatigued leg
Means and standard deviations of EMG measures in the non-fatigued leg are presented in Table 4. Main effects of test velocity were observed only for EMG and NME of VL (0.008 ≤ p ≤ .027, 3.4 ≤ d ≤ 4.9). In VM and BF, no changes in EMG were observed.
Discussion
The main goal of the present study was to examine the effects of unilateral isokinetic fatiguing knee extensions at 60°/s and 240°/s movement velocities on isokinetic neuromuscular performance measures of the contralateral homologous muscle quantified at 60 and 240°/s velocities. The main findings were that (1) maximal torque and NME decreased whereas EMG (i.e., VL, BF) increased in the fatigued knee extensors following unilateral exercise-induced fatigue particularly when using faster movement velocities, (2) maximal torque of the non-fatigued knee extensors was not impaired following unilateral exercise-induced fatigue, (3) the TTP was reduced in the non-fatigued contralateral leg after FAT60 only, and (4) the changes in neuromuscular performance of the fatigued and non-fatigued leg following fatiguing contractions at 60 and 240°/s were independent of the testing velocity (i.e., 60, 240°/s).
In the present study, the results of sigificnantly larger PIT decrements in the fatigued leg following unilateral knee extensions at 240°/s velocities indicated that fatigue was induced in the exercised leg particularly when using faster movement velocities. In fact, muscle fatigue can be defined as any exercise-induced reduction in force and power production of the neuromuscular system regardless of whether or not the task can be sustained (Bigland-Ritchie and Woods 1984). In support of our findings, several previous studies showed greater fatigue effects (i.e., lower force/torque/power output) following repetitive dynamic multi-joint (Bogdanis et al. 2007; Ide et al. 2011) and single-joint (Mathiassen 1989; Douris 1993; Morel et al. 2015) movements at faster compared to slower velocities. For instance, Ide et al. (2011) reported that decrements in leg press one-repetition maximum were significantly larger after leg press and leg extension exercises (each 5 × 12 repetitions) when execution time for each repetition was 1.5 s compared to 6 s. Further, in a recent study of Morel and colleagues (2015), it was demonstrated that unilateral fatiguing knee extensions at 240°/s contributed to significantly lower maximal isometric force levels when compared to fatiguing knee extensions at 0 and 30°/s. Additionally, these authors found that fatigue-induced metabolic stress (i.e., blood lactate accumulation) and deficits in contractile function were more pronounced at faster compared to slower movement velocities during application of the fatigue protocol (Morel et al. 2015). Similarly, the present study revealed that NME of knee extensor muscles was significantly more affected when using faster movement velocities during the fatigue protocol. Of note, changes in NME are associated with altered muscle contractile function (Bigland-Ritchie 1981). With respect to the present findings, significantly lower NME values (in VM and VL) indicated reduced torque output at a given muscle activation level only following the FAT240 protocol. This may imply that fatigue induced by faster maximal concentric knee extensions was generated more by changes within the muscle of the exercised limb (e.g., propagation of action potential, excitation contraction coupling, calcium kinetics, and greater ATP turnover at higher contraction speed; for review see Stainsby and Barclay 1976, and Enoka and Duchateau 2016) versus neural based modulation.
Notably, Morel et al. (2015) further reported that voluntary activation of the knee extensors was significantly more reduced following the fatigue protocol when using slower compared to higher movement velocities. Thus, it was hypothesized that the neural stress associated with slower movement velocities during dynamic unilateral fatiguing knee extensions may particularly contribute to enhanced cross-over responses in torque production of the non-fatigued leg when compared to faster movement velocities. However, in the present study, no cross-over fatigue effects were found in the contralateral non-exercised knee extensors although maximal unilateral concentric knee extensions led to significant performance decrements in the exercised limb compared to control condition. Indeed, this result is in agreement with previous findings, in which dynamic unilateral fatigue protocols in the lower limbs did not affect maximal force/torque output on contralateral muscles (Grabiner and Owings 1999; Regueme et al. 2007; Berger et al. 2010; Amann et al. 2013). For instance, in the study of Grabiner and Owings (1999), subjects performed either concentric or eccentric unilateral fatiguing knee extensions. Although fatigue was induced in the exercised limb, no changes in torque production (i.e., concentric mode) or even significantly increased torque output (i.e., eccentric mode) were found in the non-exercised limb following fatigue. However, in contrast to the studies mentioned above, Kawamoto et al. (2014) reported moderate-to-large-sized decrements in force output (4–7 %) of the non-fatigued contralateral homologous muscles following dynamic unilateral knee extensions at 40 or 70 % of maximal isometric voluntary contractions. The reason for these inconsistencies in the literature is unclear; however, divergence of the results may be attributed to parameters such as the recruited fatigue protocols, the testing protocols for the non-fatigued muscle groups or affected muscle groups. For instance, a recent systematic review on cross-over or non-local muscle fatigue showed that higher exercise intensities (i.e., force/torque level) during unilateral fatiguing muscle contractions tend to increase modulating effects in non-exercised muscle groups (Halperin et al. 2015). In this regard, it is well-established that maximal force output increases progressively when decreasing movement velocity (i.e., Hill’s muscle model) (Hill 1938; Thorstensson et al. 1976; Westing et al. 1991). Moreover, from a mechanistic point of view, alterations in the neural drive to the muscles appear to account for some of the observed cross-over effects in the non-exercised muscles (Todd et al. 2003; Halperin et al. 2015). Interestingly, Morel and colleagues (2015) demonstrated that movement velocities of ≤30°/s during unilateral fatiguing knee extensions induced significant decrements in voluntary muscle activation (i.e., knee extensors) whereas faster movement velocities (240°/s) did not. In the present study, unilateral fatiguing knee extensions were conducted at 60 and 240°/s. Thus, it could be speculated that the movement velocities used during the fatigue protocols of this study provided insufficient changes in the neural drive to consistently induce cross-over fatigue in the lower limbs. At last, the limited cross-over fatigue effects in the present study may be attributed not only to the fatigue protocol but also to the testing conditions used. Repetitive/prolonged compared to single testing trials demand more persistent neural input, which could augment muscle activation patterns (e.g. inter-hemispheric and/or corticospinal inhibition) demonstrating cross-over fatigue effects more clearly (Behm 2004).
Although our data did not show any evidence of cross-over fatigue (i.e., PIT levels in the non-fatigued leg were not reduced), the pattern of maximal torque production was modulated in the non-fatigued contralateral leg after slower velocity unilateral fatiguing contractions (i.e., FAT60) and demonstrated significantly lower values for TTP (Fig. 3). The exact mechanism responsible for the alteration in contralateral torque production is unclear. However, it could be speculated that unilateral fatiguing knee extensions could have modulated central nervous circuitries which determine neural control and motor programming of torque production in the non-fatigued leg. This explanation is supported by those studies which have demonstrated alterations of the corticomotor responses recorded from neural pathways which innervating non-exercised limb (Takahashi et al. 2011; Aboodarda et al. 2016). The alteration of neural control in non-exercised contralateral limb could be mediated through transcallosal connection, which is the underlying pathway for transfer of signals between homologous muscles in the two hemispheres (Hortobagyi et al. 2003; Carson et al. 2004). However, it is unclear why the observed change in TTP was more evident at 5 min post-intervention without changes in PIT angle (the 30 s post-test showed a trend toward significance p < .10). This result could be due to inter-individual variability among subjects which is supported by the large standard deviation presented in Fig. 3. Therefore, further research is required to determine the effect of unilateral neuromuscular fatigue induced at different movement velocities on motor programming and task control of the contralateral limb.
In terms of velocity specificity, the present study failed to show that the fatigue effects in both legs were more pronounced if the movement velocity during testing complies with the velocity used during the fatigue protocol. This result is partly in line with the findings of Rebai et al. (2012) who investigated the effect of 5 × 10 maximal unilateral concentric knee extensions at angular velocity of 120°/s on maximal torque output at different angular velocities (i.e., 60, 120, and 180°/s). In their study, PIT and muscle activation levels significantly decreased following fatigue for each movement velocity during testing in the exercised knee extensors. Interestingly, PIT decrements were even significantly larger using the velocity of 60°/s compared to 120 and 180°/s. It has to be noted that the evidence of velocity specificity has predominantly emerged from resistance training studies (Behm and Sale 1993). However, the present study and that of Rebai et al. (2012) are cross-sectional studies investigating acute effects of fatiguing muscle actions. Thus, it appears reasonable to assume that the concept of velocity specificity becomes less distinct for acute compared to long-term adaptations to training stimuli.
The methodological considerations which are central to this study are the following (1) participants were asked to produce maximal force output during each trial (either fatiguing contractions or testing trials at both movement velocities 60 and 240°/s). An increase in EMG activity was observed for the VL and BF after FAT240 (irrespective of the test velocity). The exact mechanism for this finding is unresolved. However, a potential explanation is that the amplitude of the signal can change due to activation shift/interval (muscle on or muscle off) and not necessarily as a consequence of a different level of overall muscle activation (Ivanenko et al. 2004). Another potential explanation is that the warm-up protocol used in our study did not adequately potentiate the central and peripheral mechanisms controlling the high intensity contractions. Therefore, participants could adopt neural activation strategies to propagate more central motor command following FAT240. (2) As previously described, post-intervention trials were performed for the exercised leg after set four and for the non-exercised leg after set five of the fatigue protocol (see "Methods" section). The prolonged resting period between sets four and five (approximately 45 s) may indeed have provided some recovery effect.
Conclusion
In summary, the present study revealed that unilateral fatiguing knee extensions did not produce any impairment of maximal torque output in the contralateral homologous muscles. However, slower movement velocities (i.e., 60°/s) induced large-sized cross-over modulations in torque production of the non-fatigued leg (i.e., reduced TTP) lasting up to 5 min following the fatigue protocol. Further, torque production changes following repetitive slow and fast muscle actions in the fatigued and non-fatigued leg appear to be independent of movement velocities between fatigue protocols and testing procedures. This indicates that the concept of velocity specificity is less distinct for acute adaptations to training stimuli. Based on the present findings, it can be postulated that unilateral fatiguing knee extensions at slower movement velocities modulate the neural strategies of torque production in the contralateral non-fatigued leg. These neural changes appear to be responsible for shorter TTP values in the non-fatigued leg. It is suggested that coaches and practitioners should particularly consider the exercise order during (resistance) training programs when using unilateral muscle actions or exercises at slower rather than faster movement velocities.
Abbreviations
- ANOVA:
-
Analysis of variance
- BF:
-
m. biceps femoris
- CON:
-
Control condition
- EMG:
-
Electromyographic
- FAT60:
-
Fatigue protocol at angular velocities of 60°/s
- FAT240:
-
Fatigue protocol at angular velocities of 240°/s
- NME:
-
Neuromuscular efficiency
- PIT:
-
Peak isokinetic torque
- TTP:
-
Time to peak isokinetic torque
- VL:
-
m. vastus lateralis
- VM:
-
m. vastus medialis
References
Aboodarda SJ, Copithorne DB, Power KE, Drinkwater E, Behm DG (2015) Elbow flexor fatigue modulates central excitability of the knee extensors. Appl Physiol Nutr Metab 40(9):924–930. doi:10.1139/apnm-2015-0088
Aboodarda SJ, Šambaher N, Behm DG (2016) Unilateral elbow flexion fatigue modulates corticospinal responsiveness in non-fatigued contralateral biceps brachii. Scand J Med Sci Sports 26(11):1301–1312. doi:10.1111/sms.12596
Amann M, Venturelli M, Ives SJ, McDaniel J, Layec G, Rossman MJ, Richardson RS (2013) Peripheral fatigue limits endurance exercise via a sensory feedback-mediated reduction in spinal motoneuronal output. J Appl Physiol 115(3):355–364. doi:10.1152/japplphysiol.00049.2013
Babault N, Pousson M, Michaut A, Ballay Y, van Hoecke J (2002) EMG activity and voluntary activation during knee-extensor concentric torque generation. Eur J Appl Physiol 86(6):541–547. doi:10.1007/s00421-002-0579-3
Behm DG (2004) Force maintenance with submaximal fatiguing contractions. Can J Appl Physiol 29(3):274–290
Behm DG, Sale DG (1993) Velocity specificity of resistance training. Sports Med 15(6):374–388
Behm DG, Leonard AM, Young WB, Bonsey, W Andrew C, MacKinnon SN (2005) Trunk muscle electromyographic activity with unstable and unilateral exercises. J Strength Cond Res 19(1):193–201. doi:10.1519/1533-4287(2005)19<193:TMEAWU>2.0.CO;2
Berger LL, Regueme SC, Forestier N (2010) Unilateral lower limb muscle fatigue induces bilateral effects on undisturbed stance and muscle EMG activities. J Electromyogr Kinesiol 20(5):947–952. doi:10.1016/j.jelekin.2009.09.006
Bigland-Ritchie B (1981) EMG/force relations and fatigue of human voluntary contractions. Exerc Sport Sci Rev 9:75–117
Bigland-Ritchie B, Woods JJ (1984) Changes in muscle contractile properties and neural control during human muscular fatigue. Muscle Nerve 7(9):691–699. doi:10.1002/mus.880070902
Bogdanis GC, Papaspyrou A, Theos A, Maridaki M (2007) Influence of resistive load on power output and fatigue during intermittent sprint cycling exercise in children. Eur J Appl Physiol 101(3):313–320. doi:10.1007/s00421-007-0507-7
Carroll TJ, Herbert RD, Munn J, Lee M, Gandevia SC (2006) Contralateral effects of unilateral strength training: evidence and possible mechanisms. J Appl Physiol 101(5):1514–1522. doi:10.1152/japplphysiol.00531.2006
Carson RG, Riek S, Mackey DC, Meichenbaum DP, Willms K, Forner M, Byblow WD (2004) Excitability changes in human forearm corticospinal projections and spinal reflex pathways during rhythmic voluntary movement of the opposite limb. J Physiol 560(3):929–940. doi:10.1113/jphysiol.2004.069088
Cohen J (1988) Statistical power analysis for the behavioral sciences, 2nd edn. Erlbaum, Hillsdale
Coren S (1993) The lateral preference inventory for measurement of handedness, footedness, eyedness, and earedness: Norms for young adults. Bull Psychon Soc 31(1):1–3
Douris PC (1993) The effect of isokinetic exercise on the relationship between blood lactate and muscle fatigue. J Orthop Sports Phys Ther 17(1):31–35. doi:10.2519/jospt.1993.17.1.31
Enoka RM, Duchateau J (2016) Translating fatigue to human performance. Med Sci Sports Exerc. doi:10.1249/MSS.0000000000000929
Frey I, Berg A, Grathwohl D, Keul J (1999) Freiburg Questionnaire of physical activity-development, evaluation and application. (Freiburg Questionnaire of physical activity–development, evaluation and application). Soz Praventivmed 44(2):55–64
Grabiner MD, Owings TM (1999) Effects of eccentrically and concentrically induced unilateral fatigue on the involved and uninvolved limbs. J Electromyogr Kinesiol 9(3):185–189
Halperin I, Chapman DW, Behm DG (2015) Non-local muscle fatigue. Effects and possible mechanisms. Eur J Appl Physiol 115(10):2031–2048. doi:10.1007/s00421-015-3249-y
Hermens HJ, Merletti R, Freriks B (1999) SENIAM: European recommendations for surface electromyography results of the SENIAM project, 2nd edn. Roessingh Research and Development, Enschede
Hill AV (1938) The heat of shortening and the dynamic constants of muscle. Proc R Soc Lond B 126(843):136–195
Hortobagyi T, Lambert NJ, Hill JP (1997) Greater cross education following training with muscle lengthening than shortening. Med Sci Sports Exerc 29(1):107–112
Hortobagyi T, Taylor JL, Petersen NT, Russell G, Gandevia SC (2003) Changes in segmental and motor cortical output with contralateral muscle contractions and altered sensory inputs in humans. J Neurophysiol 90(4):2451–2459. doi:10.1152/jn.01001.2002
Ide BN, Leme TCF, Lopes CR, Moreira A, Dechechi CJ, Sarraipa MF, Da Mota GR, Brenzikofer R, Macedo DV (2011) Time course of strength and power recovery after resistance training with different movement velocities. J Strength Cond Res 25(7):2025–2033. doi:10.1519/JSC.0b013e3181e7393f
Ivanenko YP, Poppele RE, Lacquaniti F (2004) Five basic muscle activation patterns account for muscle activity during human locomotion. J Physiol 556(Pt 1):267–282. doi:10.1113/jphysiol.2003.057174
Kawamoto J-E, Aboodarda SJ, Behm DG (2014) Effect of differing intensities of fatiguing dynamic contractions on contralateral homologous muscle performance. J Sports Sci Med 13(4):836–845
Kennedy A, Hug F, Sveistrup H, Guevel A (2013) Fatiguing handgrip exercise alters maximal force-generating capacity of plantar-flexors. Eur J Appl Physiol 113(3):559–566. doi:10.1007/s00421-012-2462-1
Mathiassen SE (1989) Influence of angular velocity and movement frequency on development of fatigue in repeated isokinetic knee extensions. Eur J Appl Physiol Occup Physiol 59(1–2):80–88
Morel B, Clémençon M, Rota S, Millet GY, Bishop DJ, Brosseau O, Rouffet DM, Hautier CA (2015) Contraction velocity influence the magnitude and etiology of neuromuscular fatigue during repeated maximal contractions. Scand J Med Sci Sports 25(5):e432–e441. doi:10.1111/sms.12358
Munn J, Herbert RD, Gandevia SC (2004) Contralateral effects of unilateral resistance training: a meta-analysis. J Appl Physiol 96(5):1861–1866. doi:10.1152/japplphysiol.00541.2003
Prieske O, Wick D, Granacher U (2014) Intrasession and intersession reliability in maximal and explosive isometric torque production of the elbow flexors. J Strength Cond Res 28(6):1771–1777. doi:10.1519/JSC.0000000000000321
Rebai H, Zarrouk N, Sahli S, Tabka Z, Hug F, Dogui M (2012) Effect of heavy isokinetic intermittent exercise on acute neuromuscular fatigue in knee extensors. Isokin Exerc Sci 20(2):121–128
Regueme SC, Barthelemy J, Nicol C (2007) Exhaustive stretch–shortening cycle exercise: no contralateral effects on muscle activity in maximal motor performances. Scand J Med Sci Sports 17(5):547–555. doi:10.1111/j.1600-0838.2006.00614.x
Sambaher N, Aboodarda SJ, Behm DG (2016) Bilateral knee extensor fatigue modulates force and responsiveness of the corticospinal pathway in the non-fatigued, dominant elbow flexors. Front Hum Neurosci 10:18. doi:10.3389/fnhum.2016.00018
Sidhu SK, Weavil JC, Venturelli M, Garten RS, Rossman MJ, Richardson RS, Gmelch BS, Morgan DE, Amann M (2014) Spinal mu-opioid receptor-sensitive lower limb muscle afferents determine corticospinal responsiveness and promote central fatigue in upper limb muscle. J Physiol 592(22):5011–5024. doi:10.1113/jphysiol.2014.275438
Stainsby WN, Barclay JK (1976) Effect of initial length on relations between oxygen uptake and load in dog muscle. Am J Physiol 230(4):1008–1012
Takahashi K, Maruyama A, Hirakoba K, Maeda M, Etoh S, Kawahira K, Rothwell JC (2011) Fatiguing intermittent lower limb exercise influences corticospinal and corticocortical excitability in the nonexercised upper limb. Brain Stimul 4(2):90–96. doi:10.1016/j.brs.2010.07.001
Thorstensson A, Grimby G, Karlsson J (1976) Force–velocity relations and fiber composition in human knee extensor muscles. J Appl Physiol 40(1):12–16
Todd G, Petersen NT, Taylor JL, Gandevia SC (2003) The effect of a contralateral contraction on maximal voluntary activation and central fatigue in elbow flexor muscles. Exp Brain Res 150(3):308–313. doi:10.1007/s00221-003-1379-7
Van Dieen JH, Ogita F, Haan A de (2003) Reduced neural drive in bilateral exertions: a performance-limiting factor? Med Sci Sports Exerc 35(1):111–118. doi:10.1249/01.MSS.0000043476.34920.AF
Westing SH, Cresswell AG, Thorstensson A (1991) Muscle activation during maximal voluntary eccentric and concentric knee extension. Eur J Appl Physiol Occup Physiol 62(2):104–108
Acknowledgements
No sources of funding were used to assist in the preparation of this study.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflicts of interest that are directly relevant to the content of this study.
Additional information
Communicated by Nicolas Place.
Rights and permissions
About this article

Cite this article
Prieske, O., Aboodarda, S.J., Benitez Sierra, J.A. et al. Slower but not faster unilateral fatiguing knee extensions alter contralateral limb performance without impairment of maximal torque output. Eur J Appl Physiol 117, 323–334 (2017). https://doi.org/10.1007/s00421-016-3524-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-016-3524-6