
Abstract
Alzheimer’s disease (AD) is characterized by β-amyloid plaques and intraneuronal τ aggregation usually associated with cerebral amyloid angiopathy (CAA). Both β-amyloid plaques and CAA deposits contain fibrillar aggregates of the amyloid β-peptide (Aβ). Aβ plaques and CAA develop first in neocortical areas of preclinical AD patients and, then, expand in a characteristic sequence into further brain regions with end-stage pathology in symptomatic AD patients. Aβ aggregates are not restricted to amyloid plaques and CAA. Soluble and several types of insoluble non-plaque- and non-CAA-associated Aβ aggregates have been described. Amyloid fibrils are products of a complex self-assembly process that involves different types of transient intermediates. Amongst these intermediate species are protofibrils and oligomers. Different variants of Aβ peptides may result from alternative processing or from mutations that lead to rare forms of familial AD. These variants can exhibit different self-assembly and aggregation properties. In addition, several post-translational modifications of Aβ have been described that result, for example, in the production of N-terminal truncated Aβ with pyroglutamate modification at position 3 (AβN3pE) or of Aβ phosphorylated at serine 8 (pSer8Aβ). Both AβN3pE and pSer8Aβ show enhanced aggregation into oligomers and fibrils. However, the earliest detectable soluble and insoluble Aβ aggregates in the human brain exhibit non-modified Aβ, whereas AβN3pE and pSer8Aβ are detected in later stages. This finding indicates the existence of different biochemical stages of Aβ aggregate maturation with pSer8Aβ being related mainly to cases with symptomatic AD. The conversion from preclinical to symptomatic AD could thereby be related to combined effects of increased Aβ concentration, maturation of aggregates and spread of deposits into additional brain regions. Thus, the inhibition of Aβ aggregation and maturation before entering the symptomatic stage of the disease as indicated by the accumulation of pSer8Aβ may represent an attractive treatment strategy for preventing disease progression.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Alzheimer’s disease (AD) is a neurodegenerative disorder that leads to cognitive decline [5, 85]. Definite AD diagnosis depends on the postmortem neuropathological analysis of brain sections and the detection of extracellular plaques that consist of amyloid β-peptide (Aβ) fibrils as well as intracellular neurofibrillary tangles (NFTs) with abnormally phosphorylated τ-protein [5, 47, 58, 81]. The clinical diagnosis of symptomatic AD in living patients is currently based upon signs of dementia and positive AD biomarkers, such as amyloid positron emission tomography (PET) and pathological Aβ or τ-levels in the cerebrospinal fluid [25, 85]. This symptomatic phase of the disease is preceded by preclinical AD (preAD), which can be discriminated from non-AD cases by the presence of the above-mentioned biomarkers [25, 125, 145]. Pathologically defined preclinical AD (p-preAD) encounters all cases that exhibit AD pathology at autopsy, i.e., amyloid plaques and NFTs, but did not exhibit any signs of cognitive decline during life [102, 133].
Aβ is a 36–43 amino acid peptide that results from β- and γ-secretase-mediated cleavage of the amyloid precursor protein (APP) [48, 65, 99, 106]. The most abundant forms of Aβ are the 40- and 42-residue peptide variants Aβ1–40 and Aβ1–42. In addition, several post-translational modifications of Aβ have been identified, including N-terminal truncations and pyroglutamate modifications at residues 3 or 11 (AβN3pE and AβN11pE) as well as phosphorylation at Serine residue 8 and 26 (pSer8Aβ and pSer26Aβ) [69, 89, 111, 112]. Although Aβ occurs mainly in the extracellular space, it can also be found within neurons [36, 42, 153]. Intraneuronal Aβ aggregates may colocalize with tau and contain AβN3pE or pSer8Aβ. They could also impair neuronal function and correlate with neurodegeneration [41, 72, 104, 128, 152].
Aβ peptides have intrinsic tendency to self-assemble into a range of different aggregates that are termed oligomers, protofibrils or mature amyloid fibrils based on their appearance by electron or atomic force microscopy [38, 52, 75]. Another way of defining different forms of Aβ extracted from human brains is based on the separation of soluble and insoluble fractions by differential ultracentrifugation and centrifugation steps with different solvents [121]. The species found in the Tris-buffered supernatant after 175,000×g ultracentrifugation will be referred to as ‘soluble Aβ’ and those in the pellet as ‘insoluble Aβ’. The ‘dispersible Aβ’ fraction belongs to the insoluble Aβ fraction but remains in the supernatant of Tris-buffered brain homogenates after centrifugation at 14,000×g [101]. ‘Membrane-associated Aβ’ is the other part of the ‘insoluble Aβ fraction’ that remains in the pellet after 14,000×g centrifugation and requires extraction with sodium dodecyl sulfate (SDS) or Triton X. Plaque-associated Aβ can only be recovered from the 14,000×g pellet by formic acid treatment [101, 121]. Aβ plaques as well as all types of soluble and insoluble non-plaque-associated, dispersible and membrane-associated Aβ oligomers, protofibrils and fibrils are found in symptomatic AD as well as in p-preAD cases [7, 96, 102, 137, 139]. A detailed protocol for the centrifugation steps required to separate the soluble, dispersible, membrane-associated and plaque-associated fractions can be found in the literature, e.g., Rijal Upadhaya et al. [102].
Here, we will review the neuropathological, molecular structural and biochemical aspects of Aβ aggregation and discuss its relation to AD neuropathology and the clinical stages of the disease.
Neuropathology of amyloid plaque deposition and cerebral amyloid angiopathy (CAA)
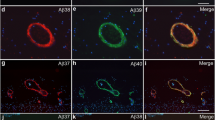
Amyloid plaques and CAA are morphologically detectable correlatives of Aβ aggregation in AD (Fig. 1), whereas soluble Aβ oligomers as well as dispersed Aβ oligomers, protofibrils and fibrils are usually not detected by immunohistochemistry with conventional anti-Aβ antibodies [38, 81, 139]. Figure 1a–e shows the most prominent plaque types occurring in the human brain with all relating to AD [91, 138]. The morphological appearance of distinct plaque types is related to their anatomical distribution. For example, fleecy amyloid is restricted to the layers pri-α, pri-β, pri-γ of the entorhinal cortex, and to the CA1-subiculum region, while lake-like amyloid occurs exclusively in the presubicular region [138]. CAA can occur in all types of vessels including capillaries (Fig. 1f, g). However, amyloid plaques and CAA are not specific for symptomatic AD and can be seen in non-demented individuals [7, 96, 137, 139]. For the neuropathological diagnosis of AD, as published by the National Institute of Aging and the Alzheimer Association, all cases in which Aβ plaques are found in the brain are to be diagnosed with AD pathology regardless of their clinical status [91]. However, we classify non-demented cases with pathologically detectable AD pathology as p-preAD cases in contrast to symptomatic AD cases [102, 133, 139]. This definition does not imply that all of these cases would have necessarily converted into symptomatic AD given the chance to live longer but rather describes that non-demented patients can have AD pathology.

Different types of Aβ plaques in the human brain [138]. All are today considered to represent AD pathology [91]. a Diffuse and cored plaques. b Presubicular lake-like amyloid. c Subpial band-like amyloid (arrows). d Fleecy amyloid (arrows). e Neuritic plaques (arrowheads), diffuse non-neuritic plaque (arrow). f Artery with Aβ deposition in the vessel wall, i.e., CAA. g Capillary Aβ deposition (arrows). Stainings: a–d, f, g anti-Aβ17–24 (4G8; Covance, Dedham, USA); e anti-Aβ8–17 (6F/3D, Novocastra, Newcastle upon Tyne, UK) counterstained with Gallyas silver staining, calibration bar in g valid for: a 30 µm, b 160 µm, c 55 µm, d 145 µm, e, g 25 µm, f 45 µm. CA1 Ammon’s horn sector CA1, ER entorhinal regions, OCC occipital isocortex, TI temporal isocortex
Neuritic plaques (Fig. 1e) comprise a distinct subgroup of amyloid plaques that is characterized by the combined occurrence of Aβ deposits and dystrophic neurites [22, 90]. These plaques have been considered to have specific pathological value for AD and are, therefore, included in the neuropathological criteria for the postmortem diagnosis of AD [58, 90]. Two types of dystrophic neurites can be associated with neuritic plaques: (1) APP-containing dystrophic neurites and (2) paired helical filaments containing dystrophic neurites consisting of abnormal τ protein [22, 148]. Neuritic plaques can contain both or exclusively either of these types of dystrophic neurites [22, 148], whereby τ-positive neuritic plaques appear more frequently than τ-negative neuritic plaques [28]. Since neuritic plaques occur later in the development of AD-related pathology than diffuse, non-neuritic Aβ plaques [45, 132, 135, 138], it is in our opinion more likely that the occurrence of dystrophic neurites within neuritic plaques represents a reactive lesion due to axonal damage. Similarly, APP-positive dystrophic neurites are observed after head trauma, brain infarction or artificial brain tissue damage by laser irradiation [43, 63, 84, 119]. A further argument that Aβ aggregates could trigger APP-positive dystrophic neurites is the finding in APP transgenic mice that CAA lesions attract sprouting of APP-positive dystrophic neurites [95]. Interactions between Aβ and APP with abnormal τ are likely to ensue because APP accumulation and τ aggregation can occur in the same dystrophic neurite [28, 136] and because dystrophic neurites develop in the close vicinity of extracellular Aβ [22, 23, 154]. At synapses a colocalization of intracellular τ and Aβ has been described [15, 128] probably pointing to the synapse as a critical anatomical correlative for the AD-related neurodegeneration process with synapse loss as a well-known neuropathological feature of AD [21, 130].
Amyloid plaque pathology as well as CAA pathology usually starts in neocortical brain regions before they expand first into allocortical areas and then into the rest of the brain [134, 137]. Neocortical amyloid plaques define the first phase of amyloid plaque pathology, while phase 2 shows an additional involvement of allocortical areas, such as the entorhinal cortex, hippocampus, and cingulate gyrus. In phase 3, further amyloid plaques become detectable in the striatum, hypothalamus, thalamus, and the basal forebrain, while phase 4 shows additional plaque pathology in the midbrain and the medulla oblongata. Finally, in phase 5, plaques are also seen in the cerebellum and the pons [137] (Fig. 2). Similarly, stage 1 of CAA begins in the cortical and leptomeningeal vessels of neocortical areas and then expands into allocortical regions and the cerebellum (CAA stage 2). Finally, vascular Aβ deposits spread into vessels of the basal ganglia, diencephalon, brain stem and/or the white matter (CAA stage 3) [134] (Fig. 2).

Schematic representation of the expansion of Aβ plaques (Aβ phases) [137], vascular Aβ deposition in CAA in the human brain (CAA stages) [134] and biochemical maturation stages of Aβ aggregates as seen in the soluble, dispersible, membrane-associated, and plaque-associated fractions of brain homogenates (B-Aβ stages) from p-preAD and symptomatic AD cases [102]. For Aβ plaque deposition newly involved brain regions are marked in red, already affected areas are painted in black. For CAA newly involved brain regions are marked in blue, already affected areas are black. Except for the cerebellum the regional expansion patterns of Aβ plaques and CAA are quite similar. Parts of this figure are reproduced with permission [102, 137]
The progression from p-preAD to the symptomatic phase of AD is associated with the spread of Aβ pathology and changes in the composition of Aβ aggregates. Biochemically, Aβ42 is the first Aβ species (Fig. 3) to accumulate in the human brain [59, 76]. Aβ40 is detected subsequently, followed by N-terminal truncated and pyroglutamate-modified AβN3pE and/or AβN11pE (Fig. 3; Table 1). These modified forms of Aβ are frequently detected in plaques of p-preAD cases and in all AD cases [59, 60, 76, 102]. In some studies, AβN3pE and Aβ42 occurred together within plaques in every case [60]. However, in our sample (n = 74) we found few p-preAD cases (n = 3) with Aβ plaques that did not exhibit AβN3pE (Table 1). pSer8Aβ in plaques was less frequently observed in p-preAD cases than AβN3pE and AβN11pE (Figs. 2, 3). It was mainly restricted to symptomatic AD cases [102]. The α-secretase-cleaved P3 fragment (Fig. 3) was also found mainly in plaques of symptomatic AD patients [60] (Table 1). Biochemical extraction from human neocortex homogenates also revealed a similar differential distribution of Aβ and its modified forms in soluble, dispersible and membrane-associated Aβ aggregates [102]. Notably, the occurrence of modified forms of Aβ in biochemical isolates of soluble and insoluble Aβ aggregates correlated well with its neuropathological detection in plaques following a specific sequence: (1) non-modified Aβ, (2) AβN3pE and (3) pSer8Aβ. This sequence of Aβ aggregate maturation was considered to represent sequential changes in the biochemical composition of Aβ aggregates allowing the identification of three stages of the biochemical composition of Aβ aggregates (B-Aβ stages) [102] (Fig. 2).

Cleavage of APP and production of Aβ, P3 by β-, γ-, and α-secretase cleavage and its modified forms. The distinct Aβ species are ranked by their sequence of occurrence in amyloid plaques. The species found in preclinical (p-preAD) and clinical AD (AD) cases are indicated
Mechanism of Aβ aggregation and structure of Aβ fibrils and intermediates
Mature amyloid fibrils are the major compound of amyloid plaques or Aβ-derived deposits in CAA [38, 81]. These aggregates represent the terminal states of the Aβ fibrillation process, at least in vitro [18, 92]. Fibrils have a width of ~10–20 nm and a length of usually more than 1 μm [110]. Cryo transmission electron microscopy (TEM)-based reconstructions of the three-dimensional electron densities of in vitro-formed Aβ fibrils revealed one or several protofilaments that construct the full-scale fibril. These protofilaments share a relatively conserved cross-sectional architecture in Aβ1–40 and Aβ1–42 fibrils [108, 118], which suggests the presence of similar conformations among the different fibrils. The fibrils are constructed from intermolecular β-sheets in which peptides are arranged with β-strands. These strands are oriented perpendicular to the main fibril axis with the backbone hydrogen bonds located parallel. This type of assembly is termed a cross-β structure. The Aβ residues forming this structure are located within the peptide center and at the C-terminus (approximately residues 16–20 and 31–36) [31]. The N-terminus is conformationally flexible in many, but probably not all types of fibrils [31]. These data have come from the analysis of Aβ filaments that were formed in vitro, while the detailed structures of fibrils from AD brain tissue have remained largely elusive. Fibril polymorphism is another phenomenon, which is also mainly known from in vitro-formed Aβ fibrils. Aβ has been shown to adopt multiple fibril structures that can even be observed within the same sample tube [39, 86]. Polymorphic fibrils can differ in the number of their protofilaments, the relative protofilament–protofilament orientation or in their detailed peptide conformations [30]. Changing the conditions of fibril formation may affect this fibril spectrum and create broader or narrower outcomes or even induce alternate fibril structures [67]. An interesting implication of the observation of different fibril morphologies is that it raises the possibility of a strain-like behavior with Aβ aggregates. The term strain as used in the prion field refers to different phenotypic traits of prion protein aggregates in transmissible spongiform encephalopathies that are transmissible from donor to recipient upon prion infection [1]. Prion strains may have their molecular basis in different prion protein conformations and/or aggregate structures. Observation of Aβ fibril morphologies suggests that different biological effects could arise from differently structured fibrils and that these may be potentially transmissible, at least under laboratory conditions. Indeed, there is evidence for such a strain-type behavior of Aβ aggregates in APP transgenic mouse models for Aβ pathology [55, 127]. One study compared Aβ fibril structures in brain homogenates of two human AD cases and reported case-specific Aβ fibril structures distinguishing between the two individual cases [79], indicating support for strain-type behavior of Aβ in animal models.
Amyloid fibrils arise from a complex self-assembly reaction. Monomeric Aβ peptide is the theoretical precursor of all aggregates and has a random coil-like conformation in aqueous solutions at low peptide concentrations as well as in the absence of salts, as demonstrated by circular dichroism spectroscopy [131]. Molecular dynamic simulations found such Aβ monomers to adopt a collapsed micelle-like conformation where hydrophobic side chains are located mainly within the interior of the otherwise rather flexible peptide conformation [143]. In vitro, monomeric Aβ may be stable for sometime in strong denaturants, such as high molar concentrations of guanidine hydrochloride or neat trifluoroacetic acid. However, it is not very stable in aqueous solutions which mimic physiological conditions by their salt, lipid or sugar composition, and are prone to aggregate into β-sheet conformations. This process is exaggerated if the peptide concentration is relatively high [56]. These effects should be kept in mind when Aβ is claimed to be present as monomers. Apolar environments, such as detergent micelles or lipid bilayers, induce α-helical conformations [131] which reflect the natural origin of the Aβ sequence within the α-helical transmembrane domain of APP.
Monitoring the kinetics of Aβ fibril formation in vitro typically shows three phases: an initial lag phase of little fibril formation, a subsequent growth phase of rapid fibril assembly and a final stationary phase where fibril formation and fibril dissociation are at equilibrium (Fig. 4). Mathematical modeling of experimental kinetic data suggested that three main steps account for these kinetic effects: primary nucleation, fibril elongation and secondary nucleation [68]. Fibril nucleation describes the slow initial assembly of Aβ peptide into nuclei that allow the fast subsequent outgrowth of elongated fibrils by monomer addition. Secondary nucleation describes the ability of already formed fibrils to potentiate the formation of new fibrils. This amplification occurs because preformed fibrils disintegrate and expand the number of active sites that enable fibril elongation or alternatively, the surfaces of the preformed fibrils act as scaffolds to accelerate the generation of new fibril nuclei which then elongate into additional filaments [68]. Experimental evidence which supports the effectiveness of a nucleation–polymerization mechanism is provided by studies in which the addition of preformed fibrils was found to strongly accelerate fibril formation in vitro and to reduce or to eliminate the observable lag phase [51] (Fig. 4).

Schematic representation of the kinetics of fibril formation. Blue spontaneous fibril formation from monomeric precursors; red fibril formation, seeded by preformed fibrils added to the solution of monomers
Interestingly, several of these general features of fibril formation reactions in vitro are remodeled by cell culture systems of amyloid plaque formation suggesting the effectiveness of similar mechanisms. Measurement of the formation of single Aβ amyloid plaques in a cell model revealed a growth kinetics consisting of lag, growth and a stationary phase [32]. In addition, plaque formation can be accelerated by the addition of preformed amyloid fibrils to the cell model of Aβ plaque formation [32] or by injection of such material into the brain or peritoneum of Aβ-producing mice [27, 64, 87, 149]. So far, only very little is known about the molecular structure of these fibrillation seeds but it is likely that they capture structural features seen in fibrils.
While monomeric peptides and fibrils mark the starting point and the end products of fibrillation, ‘intermediates’ represent the structural states in between these two. Intermediates either occur in the course of fibril assembly or they are structurally in between a monomer and a fibril. Investigating the time course of Aβ peptide assembly into fibrils with TEM or atomic force microscopy, in vitro, revealed a range of different intermediates from protofibrils to oligomers with low and high molecular weights [29]. In addition, there are several protocols reported to prepare specific intermediates inside the test tube or to extract them from AD brains [49]. Detailed nuclear magnetic resonance-based insights into the structure of such intermediates are so far only available for a few oligomer preparations [2, 20, 53] and for protofibrils [114]. Furthermore, there are data indicating a β-barrel assembly for certain oligomeric states [74].
There are several specific oligomers, e.g., Aβ dimers, Aβ trimers, Aβ*56, and Aβ globulomers, that received special interest and were considered to have specific toxic properties that other intermediates may not have [8, 35, 57, 77, 78, 83, 121]. Some oligomeric Aβ intermediates enter the fibril-forming pathway, e.g., Aβ dimers, whereas others will accumulate off-pathway as stable non-fibrillar oligomers, e.g., Aβ trimers or globulomers [35, 40, 82, 94]. Aβ intermediates, are particularly interesting as they possess a higher specific in vitro toxicity than Aβ fibrils [37]. One argument against the critical role of Aβ plaques or fibrils in the development of AD is that NFTs provide a better neuropathological correlate to the progression of clinical deficits than Aβ plaques [6]. In the light of (1) end-stage Aβ plaque pathology in all symptomatic AD cases [137], (2) the significant increase in soluble oligomers and dispersible Aβ aggregates from p-preAD to symptomatic AD cases [102, 139] and (3) the toxic properties of dispersible Aβ aggregates in APP transgenic mouse models [101] soluble or dispersible Aβ oligomers or protofibrils rather than Aβ plaques seem to be responsible for Aβ toxicity and to contribute to disease progression. Moreover, some, though not all, oligomers appear to be capable of accelerating the lag phase for Aβ fibril formation [10]. Nevertheless, it is likely that fibrils and plaques also contribute to AD as they are able to release toxic Aβ intermediates [113]. In favor of this hypothesis, it has been considered that plaque-associated oligomers or protofibrils induce neuritic changes near amyloid plaques in APP transgenic mouse models [11, 126, 140]. As such, the current data appear to support a critical role for Aβ intermediates such as oligomers and protofibrils in AD pathogenesis probably in “collaboration” with NFTs. Whether Aβ intermediates and τ pathology have impact on one another, whether tau or Aβ is the main driver of the disease is still matter of discussion [6, 13, 50, 124]. Anyway, considering the reports on Aβ and τ effects on neurotoxicity there is strong evidence that both pathologies have impact on the disease [6, 13, 50, 71, 75, 120, 124].
However, it is not yet clear whether Aβ intermediate toxicity is restricted to distinct oligomers, e.g., Aβ dimers or Aβ*56 [78, 83] or whether all kinds of high molecular weight intermediates seen by blue-native-polyacryl gel electrophoresis in symptomatic AD cases [103] contribute to neurodegeneration. Additionally, fibrillar Aβ in CAA-affected vessels appears to damage the vessel wall [17, 144] and to alter perivascular drainage [151] as a consequence of the displacement and consecutive destruction of the vessel wall just by its physical presence. This is consistent with Aβ’s mechanical properties [109].
Modification of the Aβ peptide and its impact on aggregation and toxicity
The conformation, self-assembly and aggregation of Aβ is determined by its amino acid sequence. This is well documented for the two abundant species Aβ40 and Aβ42 [49, 146]. The longer Aβ42 variant aggregates much faster than Aβ40 and is the major species initially detected in AD-associated plaques [59, 105], even though the proteolytic processing of APP generates mainly Aβ40. Several mutations in APP have been associated with familial early onset AD. These mutations are located within or close to the Aβ domain of APP and favor the production and/or aggregation of Aβ [66, 129] (Fig. 5). Interestingly, another mutation localized close to the N-terminus of the Aβ domain (A673T) appears to be protective against AD and decreases the proteolytic generation of Aβ [62]. Thus, genetic evidence strongly supports a critical role of Aβ in the pathogenesis of AD. However, mutations in APP that cause familial AD are very rare and only account for a minority of all AD cases. Here, we focus on variants of Aβ that derive from the most abundant ‘wild-type’ form of APP by post-translational modifications and their effects on aggregate formation and their potential roles in AD pathogenesis (Fig. 6).

Localization of post-translational modifications and disease-causing mutations within and close to the Aβ domain. The amino acid sequence of Aβ42 (boxed) and neighboring N- and C-terminal sequences are given in single letter code. Positions of pyroglutamate (pE) and phosphoserine residues (P) are indicated by yellow triangles and red circles, respectively. Mutations associated with familial forms of early onset AD (black letters) or with cerebral hemorrhages or stroke and severe CAA pathology (red letters) are indicated below the amino acid sequence in single letter code. Notably, all identified disease-causing mutations in APP are localized within or close to the Aβ domain. These mutations commonly affect the proteolytic generation and/or the aggregation of Aβ. The table below lists the position of different mutations and their effects on production and aggregation of Aβ (?, effect not reported or unclear). The effects of the mutations on aggregation are predominantly deduced from in vitro experiments. Thus, it would be interesting to test how the mutations affect the solubility of the different aggregates in vivo. Mutation A673T is associated with a decreased risk of AD and considered to be protective (marked in blue). In vitro, this mutation decreased production of Aβ (Table is adapted from http://www.alzforum.org/mutations). The mutations outside the Aβ domain do not directly affect the aggregation propensity of the monomeric Aβ peptide itself, but by increasing the total production of Aβ or altering the ratio of Aβ42/40 promote oligomerization and fibrillization

Potential functional links of Aβ maturation, propagation and increase in concentration. Propagation of Aβ plaque pathology as given by the phases of Aβ deposition is associated with an increase of the Aβ content in the brain (a) as well as with maturation of Aβ aggregates in a given regions (b, c) as depicted for the hypothetical propagation of Aβ pathology from the neocortex (b) to the entorhinal cortex (c) between Aβ phases 1 and 2 (blue box in a). Mature Aβ aggregates that are predominantly found in symptomatic AD cases consist of modified forms of Aβ that increase the stability of the aggregates and, in so doing, impair its clearance and finally cause further Aβ aggregation and increased toxicity of the Aβ aggregates
A large variety of post-translational modifications has been shown for Aβ including non-covalent interactions with metal ions, lipids and other proteins or peptides [26, 54, 147]. The binding of Aβ to these ligands could modify the aggregation behavior of Aβ and hence, contribute to the pathogenesis of AD. The Aβ peptide itself can also be modified covalently by oxidation, racemization, isomerization, and as mentioned above by pyroglutamate formation and phosphorylation [73, 93, 111, 112, 122, 123].
We and others have shown that Aβ species with pyroglutamate and phosphoserine are abundantly present in both human AD and APP transgenic mice brains. Pyroglutamate-modified Aβ was initially identified in human AD brains [93, 111, 112]. The formation of pyroglutamate requires precedent trimming of the N-terminus of Aβ by exopeptidases or alternative cleavage of APP by endoproteases to expose the glutamate residue at positions 3 or 11 (Fig. 3). The enzyme glutaminyl cyclase catalyzes formation of a lactam ring resulting in Aβ with pyroglutamate at their newly formed N-termini. When compared to unmodified Aβ, AβN3pE has an increased propensity to form oligomeric and fibrillar assemblies [117]. The use of specific antibodies for AβN3pE revealed the common presence of these species in human AD and p-preAD brains [60, 102, 111, 112]. Studies with transgenic mice and cultured neurons also showed that AβN3pE exerts increased toxicity and deposits early in extracellular plaques and intraneuronal aggregates [4, 9]. In vitro, the lag phase for AβN3pE is much shorter in comparison to that for non-modified Aβ [115].
Aβ species phosphorylated at Ser8 have also been well documented in brains of transgenic mice and human AD cases [69, 72, 102, 104]. Ser8 is efficiently phosphorylated by purified cAMP-dependent protein kinase A (PKA) in vitro. Notably, an Aβ phosphorylating PKA-like activity has also been detected on the surface of primary cortical neurons and in human cerebrospinal fluids [69]. These data suggest that secreted Aβ could undergo phosphorylation by extracellular forms of PKA. In vitro studies using Thioflavin T or Congo red, showed that phosphorylation at Ser8 promotes the formation of Aβ fibrils. Kinetically, Ser8 phosphorylation decreases the lag phase of aggregation, indicating an increased formation of oligomeric nuclei. It is also interesting to note that oligomers of pSer8Aβ not only promote fibrillization of phosphorylated Aβ, but can also seed for the assembly of non-phosphorylated Aβ [69]. Studies with phosphorylation state-specific antibodies have shown co-deposition of phosphorylated and unphosphorylated Aβ species in both extracellular plaques and intraneuronal aggregates. However, pSer8Aβ appears to be selectively enriched in the core of individual plaques suggesting that it might also seed aggregation of other Aβ species in vivo.
In summary, phosphorylation of Ser8 has very similar effects on the aggregation behavior as the pyroglutamate modification strongly favoring the formation of oligomeric and fibrillar Aβ assemblies. Aβ can also be phosphorylated in vitro at Ser26 [89]. Notably, synthetic pSer26Aβ peptides assemble rapidly into “off-pathway” oligomers without proceeding to fibrillar aggregates [100]. It remains unknown if pSer26Aβ is also found in human brain and whether it plays an important role during AD pathogenesis.
The impact of Aβ modification on its aggregate stability and resistance against degradation
Aggregation of modified pyroglutamate and phosphorylated Aβ species results in structures that show, compared with monomeric Aβ, an increased resistance against proteolytic degradation or other clearance mechanisms of the brain. The N-terminal modification of Aβ by pyroglutamate formation could also decrease its proteolytic degradation by amino- or endopeptidases [116]. We have also shown that phosphorylation at Ser8 strongly decreased the cleavage of monomeric Aβ by the insulin-degrading enzyme (IDE) and angiotensin-converting enzyme [70]. Thus, these post-translational modifications increase the biostability of Aβ and eventually their concentration in the brain, which could further promote their aggregation. Whether pyroglutamate modification or phosphorylation could also affect other clearance mechanisms including: phagocytosis and subsequent intracellular degradation or the drainage via the blood brain barrier or the perivascular space, remains to be investigated.
Post-translational modifications of Aβ might also affect the physicochemical characteristics of oligomeric and fibrillar assemblies and thus, their dissociation during biochemical extraction from brains. We showed that pSer8Aβ extracted from brains of APP transgenic mice and separated by SDS-PAGE has increased stability under denaturing conditions as compared to unmodified Aβ [69]. The different characteristics of unmodified and modified Aβ species during extraction from brain tissue and the detection by Western blotting, ELISA or mass spectrometry have to be considered for quantitative analyses. Different reactivities of generic Aβ antibodies for modified and unmodified Aβ species in the monomeric or aggregated state could further complicate the interpretation of experimental results for quantification of different Aβ species.
Mechanisms for the progression of Aβ pathology in the pathogenesis of AD
As described above, Aβ pathology (1) expands from the neocortex into further brain regions following neuronal connections [134, 137], (2) follows a maturation sequence during which the concentration of soluble Aβ increases, post-translational modifications take place and insoluble aggregates accumulate [60, 76, 102], and (3) is associated with an increase in total Aβ amount [19, 102, 139, 141, 142]. However, the critical events that trigger or cause these processes are still under debate.
In the event that expansion of Aβ pathology from one brain region into another is critical for disease progression, the question still remains whether specific forms of aggregates are crucial for propagation of Aβ pathology. Expansion of Aβ pathology means that there is a specific sequence in which Aβ aggregates emerge in different brain regions as extracellular plaques in the brain parenchyma and in blood vessels. In vitro Aβ fibrillization as well as plaque formation in cell models is characterized by a lag phase in which there is only very little amyloid formation [32, 51]. Interestingly, addition of seeds composed of preformed fibrils can shorten this lag phase significantly (Fig. 4). Hence, such a seeding mechanism is a probable explanation for the observed expansion of Aβ plaque pathology from one brain region into the next. An argument favoring such a seeding hypothesis is the observation of a strain-type behavior in mouse models injected with distinct amyloid structures [55, 127] that remained detectable after expansion from one brain region into another [149]. Amyloid seeds might distribute in the brain by passive diffusion and/or by active transport in nerve or glial cells. Diffusion might be particularly relevant for soluble extracellular Aβ aggregates. It has been demonstrated that neuron-derived Aβ accumulates in CAA-affected leptomeningeal vessels without direct contact with the CNS neurons [14] and in cortex grafts from non-transgenic animals transplanted into the cortex of APP transgenic mice [88]. The finding that different brain regions connected with one another by axons become sequentially affected by Aβ plaque pathology [137] does not argue against diffusion as a major mechanism for Aβ propagation because axons may facilitate directed diffusion of Aβ aggregates. An alternative hypothesis is that cells carry and transport Aβ. Intracellular Aβ has not only been detected in neurons but also in astrocytes and microglial cells [3, 33, 42, 156]. Therefore, migrating glial cells could also contribute to the propagation of Aβ. Microglial cells and astrocytes can take up Aβ [80, 155] and are both capable of migrating into other brain regions [12, 61]. For astrocytes, migration along myelinated fiber tracts has been reported [12] which would be compatible with an expansion along axonal tracts. Thus, these cells may be able to distribute Aβ or its assemblies. For astrocytes, it has been shown that they are able to clear Aβ into the perivascular space [107]. Here, Aβ can interact with the vessel wall and ultimately result in the development of CAA [16, 151]. Cell–cell propagation of different Aβ species has been demonstrated in neuroblastoma cell culture [24] and may depend on the activity of exosomes that may contain Aβ or Aβ aggregates [98]. Whether microglial cells indeed contribute to Aβ propagation is not clear, as their deletion had no effect on plaque pathology in APP transgenic mice [44]. However, microglial cells can play an important role in AD pathogenesis as they can phagocytose Aβ and modulate neuroinflammatory processes in the brain [34, 46, 97, 150]. Thus, it is very likely that multiple mechanisms, such as passive diffusion and active transport by neurons and glial cells, contribute to the propagation of Aβ pathology throughout the brain. Therefore, it will be necessary to further dissect the relative contribution of these mechanisms to Aβ propagation and AD pathogenesis in the future.
The maturation of Aβ aggregates from deposits containing mainly non-modified Aβ into aggregates encompassing significant amounts of AβN3pE and pSer8Aβ is associated with the propagation of Aβ pathology from the neocortex to further brain regions as well as with the conversion from preclinical to symptomatic AD. Although current findings already suggest that the maturation stage of Aβ aggregates could be crucial for the toxic properties of Aβ, it will be important to further confirm whether the maturation stage of Aβ aggregates indeed determines their toxicity and if it critically affects AD pathogenesis. The presence of AβN3pE and pSer8Aβ may boost aggregation of Aβ [69, 117]. Our finding of an Aβ phase 3-like distribution pattern indicating expansion of Aβ plaques from the neocortex into other brain regions in a case with the earliest stage of biochemical Aβ aggregate maturation (B-Aβ stage 1), i.e., Aβ aggregates without detectable modified Aβ species may argue against an essential contribution of advanced Aβ aggregate maturation to the spreading and seeding of Aβ pathology. Another case exhibited mature Aβ aggregates (B-Aβ stage 3) containing Aβ, AβN3pE and pAβ only in the neocortex (Table 2). These examples imply that modified Aβ forms are no prerequisite for the propagation of Aβ aggregates. Whether distinct “off-pathway” Aβ intermediates with high toxicity are produced at distinct steps in Aβ maturation is not clear. However, the overall increase of high molecular weight Aβ intermediates in AD cases, as seen by BN-PAGE analysis of brain lysates [103], coincides with the increase of toxic intermediates. It will be interesting to further assess whether AD-related neurodegeneration is driven by several specific Aβ intermediates or whether many kinds of fibrillar and non-fibrillar Aβ aggregates contribute to the development of the disease.
In addition to the maturation of soluble and insoluble Aβ aggregates, their concentration in the neocortex increased from p-preAD cases to AD cases [102, 139]. AβN3pE and pSer8Aβ have an increased stability against proteolytic degradation [69, 117]. Accordingly, aggregates containing these variants might also have a lower tendency to dissociate. Therefore, these post-translational modifications may decrease the clearance of Aβ and contribute to increased Aβ levels during AD pathogenesis. Increased concentration of Aβ aggregates may promote spreading of Aβ pathology and further accumulation of posttranslationally modified Aβ peptides. The increase in Aβ concentration, its maturation and expansion into further brain regions could be functionally linked to the pathogenesis of AD. Once the aggregation of Aβ is initiated, further seeding and maturation of Aβ aggregates may increase the total Aβ levels in the brain and thereby accelerate the propagation of Aβ into unaffected brain regions and promote disease progression.
Conclusions
In this review, we have demonstrated that the progression from the p-preAD to the symptomatic AD phase is accompanied by an increase in concentration, maturation and expansion (spreading/propagation) of Aβ aggregate pathology. Based upon biochemical and biophysical properties of Aβ together with neuropathological findings, we hypothesize that modified Aβ species such as AβN3pE and pSer8Aβ play critical roles for Aβ accumulation, plaque maturation, seeding, and propagation in the pathogenesis of AD and for its conversion from the preclinical to symptomatic stage. Accordingly, the maturation step of Aβ aggregates while converting from p-preAD to symptomatic AD, i.e., the additional accumulation of pSer8Aβ, may be an attractive target to inhibit the progression of p-preAD to the symptomatic stage. The simple view on Aβ plaque pathology at autopsy or by amyloid PET does not automatically permit conclusions on Aβ aggregate maturation status or on the concentration of soluble and dispersible Aβ aggregates. Accordingly, for monitoring Aβ pathology in clinical studies amyloid PET and postmortem neuropathological assessments might be supplemented by maturation stage and Aβ concentration as additional outcome measures.
References
Aguzzi A, Heikenwalder M, Polymenidou M (2007) Insights into prion strains and neurotoxicity. Nat Rev Mol Cell Biol 8:552–561
Ahmed M, Davis J, Aucoin D, Sato T, Ahuja S, Aimoto S, Elliott JI, Van Nostrand WE, Smith SO (2010) Structural conversion of neurotoxic amyloid-beta(1–42) oligomers to fibrils. Nat Struct Mol Biol 17:561–567
Akiyama H, Schwab C, Kondo H, Mori H, Kametani F, Ikeda K, McGeer PL (1996) Granules in glial cells of patients with Alzheimer’s disease are immunopositive for C-terminal sequences of beta-amyloid protein. Neurosci Lett 206:169–172
Alexandru A, Jagla W, Graubner S, Becker A, Bauscher C, Kohlmann S, Sedlmeier R, Raber KA, Cynis H, Ronicke R, Reymann KG, Petrasch-Parwez E, Hartlage-Rubsamen M, Waniek A, Rossner S, Schilling S, Osmand AP, Demuth HU, von Horsten S (2011) Selective hippocampal neurodegeneration in transgenic mice expressing small amounts of truncated Abeta is induced by pyroglutamate-Abeta formation. J Neurosci 31:12790–12801
Alzheimer A (1907) Ueber eine eigenartige Erkrankung der Hirnrinde. Allg Zschr Psych 64:146–148
Arriagada PV, Growdon JH, Hedley-Whyte ET, Hyman BT (1992) Neurofibrillary tangles but not senile plaques parallel duration and severity of Alzheimer’s disease. Neurology 42:631–639
Arriagada PV, Marzloff K, Hyman BT (1992) Distribution of Alzheimer-type pathologic changes in nondemented elderly individuals matches the pattern in Alzheimer’s disease. Neurology 42:1681–1688
Barghorn S, Nimmrich V, Striebinger A, Krantz C, Keller P, Janson B, Bahr M, Schmidt M, Bitner RS, Harlan J, Barlow E, Ebert U, Hillen H (2005) Globular amyloid beta-peptide oligomer—a homogenous and stable neuropathological protein in Alzheimer’s disease. J Neurochem 95:834–847
Bayer TA, Wirths O (2014) Focusing the amyloid cascade hypothesis on N-truncated Abeta peptides as drug targets against Alzheimer’s disease. Acta Neuropathol 127:787–801
Benseny-Cases N, Cocera M, Cladera J (2007) Conversion of non-fibrillar beta-sheet oligomers into amyloid fibrils in Alzheimer’s disease amyloid peptide aggregation. Biochem Biophys Res Commun 361:916–921
Bittner T, Fuhrmann M, Burgold S, Ochs SM, Hoffmann N, Mitteregger G, Kretzschmar H, LaFerla FM, Herms J (2010) Multiple events lead to dendritic spine loss in triple transgenic Alzheimer’s disease mice. PLOS One 5:e15477
Booss J, Solly KS, Collins PV, Jacque C (1991) Migration of xenogenic astrocytes in myelinated tracts: a novel probe for immune responses in white matter. Acta Neuropathol 82:172–177
Braak H, Thal DR, Ghebremedhin E, Del Tredici K (2011) Stages of the pathological process in Alzheimer’s disease: age categories 1 year to 100 years. J Neuropathol Exp Neurol 70:960–969
Calhoun ME, Burgermeister P, Phinney AL, Stalder M, Tolnay M, Wiederhold KH, Abramowski D, Sturchler-Pierrat C, Sommer B, Staufenbiel M, Jucker M (1999) Neuronal overexpression of mutant amyloid precursor protein results in prominent deposition of cerebrovascular amyloid. Proc Natl Acad Sci USA 96:14088–14093
Capetillo-Zarate E, Gracia L, Yu F, Banfelder JR, Lin MT, Tampellini D, Gouras GK (2011) High-resolution 3D reconstruction reveals intra-synaptic amyloid fibrils. Am J Pathol 179:2551–2558
Carare RO, Bernardes-Silva M, Newman TA, Page AM, Nicoll JA, Perry VH, Weller RO (2008) Solutes, but not cells, drain from the brain parenchyma along basement membranes of capillaries and arteries: significance for cerebral amyloid angiopathy and neuroimmunology. Neuropathol Appl Neurobiol 34:131–144
Carare RO, Hawkes CA, Jeffrey M, Kalaria RN, Weller RO (2013) Review: cerebral amyloid angiopathy, prion angiopathy, CADASIL and the spectrum of protein elimination failure angiopathies (PEFA) in neurodegenerative disease with a focus on therapy. Neuropathol Appl Neurobiol 39:593–611
Caughey B, Lansbury PT (2003) Protofibrils, pores, fibrils, and neurodegeneration: separating the responsible protein aggregates from the innocent bystanders. Annu Rev Neurosci 26:267–298
Chetelat G, La Joie R, Villain N, Perrotin A, de La Sayette V, Eustache F, Vandenberghe R (2013) Amyloid imaging in cognitively normal individuals, at-risk populations and preclinical Alzheimer’s disease. Neuroimage Clin 2:356–365
Chimon S, Shaibat MA, Jones CR, Calero DC, Aizezi B, Ishii Y (2007) Evidence of fibril-like beta-sheet structures in a neurotoxic amyloid intermediate of Alzheimer’s beta-amyloid. Nat Struct Mol Biol 14:1157–1164
DeKosky ST, Scheff SW (1990) Synapse loss in frontal cortex biopsies in Alzheimer’s disease: correlation with cognitive severity. Ann Neurol 27:457–464
Dickson DW (1997) The pathogenesis of senile plaques. J Neuropathol Exp Neurol 56:321–339
Dickson DW, Farlo J, Davies P, Crystal H, Fuld P, Yen SH (1988) Alzheimer’s disease. A double-labeling immunohistochemical study of senile plaques. Am J Pathol 132:86–101
Domert J, Rao SB, Agholme L, Brorsson AC, Marcusson J, Hallbeck M, Nath S (2014) Spreading of amyloid-beta peptides via neuritic cell-to-cell transfer is dependent on insufficient cellular clearance. Neurobiol Dis 65:82–92
Dubois B, Feldman HH, Jacova C, Dekosky ST, Barberger-Gateau P, Cummings J, Delacourte A, Galasko D, Gauthier S, Jicha G, Meguro K, O’Brien J, Pasquier F, Robert P, Rossor M, Salloway S, Stern Y, Visser PJ, Scheltens P (2007) Research criteria for the diagnosis of Alzheimer’s disease: revising the NINCDS-ADRDA criteria. Lancet Neurol 6:734–746
Duce JA, Bush AI (2010) Biological metals and Alzheimer’s disease: implications for therapeutics and diagnostics. Prog Neurobiol 92:1–18
Eisele YS, Obermuller U, Heilbronner G, Baumann F, Kaeser SA, Wolburg H, Walker LC, Staufenbiel M, Heikenwalder M, Jucker M (2010) Peripherally applied Abeta-containing inoculates induce cerebral beta-amyloidosis. Science 330:980–982
Elobeid A, Rantakomi S, Soininen H, Alafuzoff I (2014) Alzheimer’s disease-related plaques in nondemented subjects. Alzheimers Dement 10:522–529
Fändrich M (2012) Oligomeric intermediates in amyloid formation: structure determination and mechanisms of toxicity. J Mol Biol 421:427–440
Fändrich M, Meinhardt J, Grigorieff N (2009) Structural polymorphism of Alzheimer Abeta and other amyloid fibrils. Prion 3:89–93
Fändrich M, Schmidt M, Grigorieff N (2011) Recent progress in understanding Alzheimer’s beta-amyloid structures. Trends Biochem Sci 36:338–345
Friedrich RP, Tepper K, Ronicke R, Soom M, Westermann M, Reymann K, Kaether C, Fändrich M (2010) Mechanism of amyloid plaque formation suggests an intracellular basis of Abeta pathogenicity. Proc Natl Acad Sci USA 107:1942–1947
Funato H, Yoshimura M, Yamazaki T, Saido TC, Ito Y, Yokofujita J, Okeda R, Ihara Y (1998) Astrocytes containing amyloid beta-protein (Abeta)-positive granules are associated with Abeta40-positive diffuse plaques in the aged human brain. Am J Pathol 152:983–992
Gandy S, Heppner FL (2013) Microglia as dynamic and essential components of the amyloid hypothesis. Neuron 78:575–577
Gellermann GP, Byrnes H, Striebinger A, Ullrich K, Mueller R, Hillen H, Barghorn S (2008) A beta-globulomers are formed independently of the fibril pathway. Neurobiol Dis 30:212–220
Gimenez-Llort L, Blazquez G, Canete T, Johansson B, Oddo S, Tobena A, LaFerla FM, Fernandez-Teruel A (2007) Modeling behavioral and neuronal symptoms of Alzheimer’s disease in mice: a role for intraneuronal amyloid. Neurosci Biobehav Rev 31:125–147
Glabe CG (2008) Structural classification of toxic amyloid oligomers. J Biol Chem 283:29639–29643
Glenner GG, Wong CW (1984) Alzheimer’s disease: initial report of the purification and characterization of a novel cerebrovascular amyloid protein. Biochem Biophys Res Commun 120:885–890
Goldsbury CS, Wirtz S, Muller SA, Sunderji S, Wicki P, Aebi U, Frey P (2000) Studies on the in vitro assembly of a beta 1–40: implications for the search for a beta fibril formation inhibitors. J Struct Biol 130:217–231
Gorman PM, Yip CM, Fraser PE, Chakrabartty A (2003) Alternate aggregation pathways of the Alzheimer beta-amyloid peptide: Abeta association kinetics at endosomal pH. J Mol Biol 325:743–757
Gouras GK, Tampellini D, Takahashi RH, Capetillo-Zarate E (2010) Intraneuronal beta-amyloid accumulation and synapse pathology in Alzheimer’s disease. Acta Neuropathol 119:523–541
Gouras GK, Tsai J, Naslund J, Vincent B, Edgar M, Checler F, Greenfield JP, Haroutunian V, Buxbaum JD, Xu H, Greengard P, Relkin NR (2000) Intraneuronal Abeta42 accumulation in human brain. Am J Pathol 156:15–20
Graham DI, Adams JH, Nicoll JA, Maxwell WL, Gennarelli TA (1995) The nature, distribution and causes of traumatic brain injury. Brain Pathol 5:397–406
Grathwohl SA, Kalin RE, Bolmont T, Prokop S, Winkelmann G, Kaeser SA, Odenthal J, Radde R, Eldh T, Gandy S, Aguzzi A, Staufenbiel M, Mathews PM, Wolburg H, Heppner FL, Jucker M (2009) Formation and maintenance of Alzheimer’s disease beta-amyloid plaques in the absence of microglia. Nat Neurosci 12:1361–1363
Griffin WS, Sheng JG, Roberts GW, Mrak RE (1995) Interleukin-1 expression in different plaque types in Alzheimer’s disease: significance in plaque evolution. J Neuropathol Exp Neurol 54:276–281
Griffin WS, Stanley LC, Ling C, White L, MacLeod V, Perrot LJ, White CL 3rd, Araoz C (1989) Brain interleukin 1 and S-100 immunoreactivity are elevated in Down syndrome and Alzheimer disease. Proc Natl Acad Sci USA 86:7611–7615
Grundke-Iqbal I, Iqbal K, Quinlan M, Tung YC, Zaidi MS, Wisniewski HM (1986) Microtubule-associated protein tau. A component of Alzheimer paired helical filaments. J Biol Chem 261:6084–6089
Haass C, Kaether C, Thinakaran G, Sisodia S (2012) Trafficking and proteolytic processing of APP. Cold Spring Harb Perspect Med 2:a006270
Haass C, Selkoe DJ (2007) Soluble protein oligomers in neurodegeneration: lessons from the Alzheimer’s amyloid beta-peptide. Nat Rev Mol Cell Biol 8:101–112
Hardy J, Selkoe DJ (2002) The amyloid hypothesis of Alzheimer’s disease: progress and problems on the road to therapeutics. Science 297:353–356
Harper JD, Lansbury PT Jr (1997) Models of amyloid seeding in Alzheimer’s disease and scrapie: mechanistic truths and physiological consequences of the time-dependent solubility of amyloid proteins. Annu Rev Biochem 66:385–407
Harper JD, Wong SS, Lieber CM, Lansbury PT (1997) Observation of metastable Abeta amyloid protofibrils by atomic force microscopy. Chem Biol 4:119–125
Haupt C, Leppert J, Ronicke R, Meinhardt J, Yadav JK, Ramachandran R, Ohlenschlager O, Reymann KG, Gorlach M, Fändrich M (2012) Structural basis of beta-amyloid-dependent synaptic dysfunctions. Angew Chem Int Ed Engl 51:1576–1579
Hayashi H, Kimura N, Yamaguchi H, Hasegawa K, Yokoseki T, Shibata M, Yamamoto N, Michikawa M, Yoshikawa Y, Terao K, Matsuzaki K, Lemere CA, Selkoe DJ, Naiki H, Yanagisawa K (2004) A seed for Alzheimer amyloid in the brain. J Neurosci 24:4894–4902
Heilbronner G, Eisele YS, Langer F, Kaeser SA, Novotny R, Nagarathinam A, Aslund A, Hammarstrom P, Nilsson KP, Jucker M (2013) Seeded strain-like transmission of beta-amyloid morphotypes in APP transgenic mice. EMBO Rep 14:1017–1022
Hortschansky P, Schroeckh V, Christopeit T, Zandomeneghi G, Fändrich M (2005) The aggregation kinetics of Alzheimer’s beta-amyloid peptide is controlled by stochastic nucleation. Protein Sci 14:1753–1759
Hung LW, Ciccotosto GD, Giannakis E, Tew DJ, Perez K, Masters CL, Cappai R, Wade JD, Barnham KJ (2008) Amyloid-beta peptide (Abeta) neurotoxicity is modulated by the rate of peptide aggregation: Abeta dimers and trimers correlate with neurotoxicity. J Neurosci 28:11950–11958
Hyman BT, Phelps CH, Beach TG, Bigio EH, Cairns NJ, Carrillo MC, Dickson DW, Duyckaerts C, Frosch MP, Masliah E, Mirra SS, Nelson PT, Schneider JA, Thal DR, Thies B, Trojanowski JQ, Vinters HV, Montine TJ (2012) National Institute on Aging–Alzheimer’s Association guidelines for the neuropathologic assessment of Alzheimer’s disease. Alzheimers Dement 8:1–13
Iwatsubo T, Odaka A, Suzuki N, Mizusawa H, Nukina N, Ihara Y (1994) Visualization of A beta 42(43) and A beta 40 in senile plaques with end-specific A beta monoclonals: evidence that an initially deposited species is A beta 42(43). Neuron 13:45–53
Iwatsubo T, Saido TC, Mann DM, Lee VM, Trojanowski JQ (1996) Full-length amyloid-beta (1–42(43)) and amino-terminally modified and truncated amyloid-beta 42(43) deposit in diffuse plaques. Am J Pathol 149:1823–1830
Jensen MB, Gonzalez B, Castellano B, Zimmer J (1994) Microglial and astroglial reactions to anterograde axonal degeneration: a histochemical and immunocytochemical study of the adult rat fascia dentata after entorhinal perforant path lesions. Exp Brain Res 98:245–260
Jonsson T, Atwal JK, Steinberg S, Snaedal J, Jonsson PV, Bjornsson S, Stefansson H, Sulem P, Gudbjartsson D, Maloney J, Hoyte K, Gustafson A, Liu Y, Lu Y, Bhangale T, Graham RR, Huttenlocher J, Bjornsdottir G, Andreassen OA, Jonsson EG, Palotie A, Behrens TW, Magnusson OT, Kong A, Thorsteinsdottir U, Watts RJ, Stefansson K (2012) A mutation in APP protects against Alzheimer’s disease and age-related cognitive decline. Nature 488:96–99
Kalaria RN, Bhatti SU, Palatinsky EA, Pennington DH, Shelton ER, Chan HW, Perry G, Lust WD (1993) Accumulation of the beta amyloid precursor protein at sites of ischemic injury in rat brain. NeuroReport 4:211–214
Kane MD, Lipinski WJ, Callahan MJ, Bian F, Durham RA, Schwarz RD, Roher AE, Walker LC (2000) Evidence for seeding of beta -amyloid by intracerebral infusion of Alzheimer brain extracts in beta -amyloid precursor protein-transgenic mice. J Neurosci 20:3606–3611
Kang J, Lemaire HG, Unterbeck A, Salbaum JM, Masters CL, Grzeschik KH, Multhaup G, Beyreuther K, Muller-Hill B (1987) The precursor of Alzheimer’s disease amyloid A4 protein resembles a cell-surface receptor. Nature 325:733–736
Kennedy JL, Farrer LA, Andreasen NC, Mayeux R, St George-Hyslop P (2003) The genetics of adult-onset neuropsychiatric disease: complexities and conundra? Science 302:822–826
Klement K, Wieligmann K, Meinhardt J, Hortschansky P, Richter W, Fändrich M (2007) Effect of different salt ions on the propensity of aggregation and on the structure of Alzheimer’s Abeta(1–40) amyloid fibrils. J Mol Biol 373:1321–1333
Knowles TP, Waudby CA, Devlin GL, Cohen SI, Aguzzi A, Vendruscolo M, Terentjev EM, Welland ME, Dobson CM (2009) An analytical solution to the kinetics of breakable filament assembly. Science 326:1533–1537
Kumar S, Rezaei-Ghaleh N, Terwel D, Thal DR, Richard M, Hoch M, Mc Donald JM, Wullner U, Glebov K, Heneka MT, Walsh DM, Zweckstetter M, Walter J (2011) Extracellular phosphorylation of the amyloid beta-peptide promotes formation of toxic aggregates during the pathogenesis of Alzheimer’s disease. EMBO J 30:2255–2265
Kumar S, Singh S, Hinze D, Josten M, Sahl HG, Siepmann M, Walter J (2012) Phosphorylation of amyloid-beta peptide at serine 8 attenuates its clearance via insulin-degrading and angiotensin-converting enzymes. J Biol Chem 287:8641–8651
Kumar S, Tepper K, Kaniyappan S, Biernat J, Wegmann S, Mandelkow EM, Muller D, Mandelkow E (2014) Stages and conformations of tau repeat domain during aggregation and effects on neuronal toxicity. J Biol Chem 289:20318–20332
Kumar S, Wirths O, Theil S, Gerth J, Bayer TA, Walter J (2013) Early intraneuronal accumulation and increased aggregation of phosphorylated Abeta in a mouse model of Alzheimer’s disease. Acta Neuropathol 125:699–709
Kummer MP, Hermes M, Delekarte A, Hammerschmidt T, Kumar S, Terwel D, Walter J, Pape HC, Konig S, Roeber S, Jessen F, Klockgether T, Korte M, Heneka MT (2011) Nitration of tyrosine 10 critically enhances amyloid beta aggregation and plaque formation. Neuron 71:833–844
Laganowsky A, Liu C, Sawaya MR, Whitelegge JP, Park J, Zhao M, Pensalfini A, Soriaga AB, Landau M, Teng PK, Cascio D, Glabe C, Eisenberg D (2012) Atomic view of a toxic amyloid small oligomer. Science 335:1228–1231
Lambert MP, Barlow AK, Chromy BA, Edwards C, Freed R, Liosatos M, Morgan TE, Rozovsky I, Trommer B, Viola KL, Wals P, Zhang C, Finch CE, Krafft GA, Klein WL (1998) Diffusible, nonfibrillar ligands derived from Abeta1–42 are potent central nervous system neurotoxins. Proc Natl Acad Sci USA 95:6448–6453
Lemere CA, Blusztajn JK, Yamaguchi H, Wisniewski T, Saido TC, Selkoe DJ (1996) Sequence of deposition of heterogeneous amyloid beta-peptides and APO E in Down syndrome: implications for initial events in amyloid plaque formation. Neurobiol Dis 3:16–32
Lesne S, Koh MT, Kotilinek L, Kayed R, Glabe CG, Yang A, Gallagher M, Ashe KH (2006) A specific amyloid-beta protein assembly in the brain impairs memory. Nature 440:352–357
Lesne SE, Sherman MA, Grant M, Kuskowski M, Schneider JA, Bennett DA, Ashe KH (2013) Brain amyloid-beta oligomers in ageing and Alzheimer’s disease. Brain 136:1383–1398
Lu JX, Qiang W, Yau WM, Schwieters CD, Meredith SC, Tycko R (2013) Molecular structure of beta-amyloid fibrils in Alzheimer’s disease brain tissue. Cell 154:1257–1268
Mandrekar S, Jiang Q, Lee CY, Koenigsknecht-Talboo J, Holtzman DM, Landreth GE (2009) Microglia mediate the clearance of soluble Abeta through fluid phase macropinocytosis. J Neurosci 29:4252–4262
Masters CL, Simms G, Weinman NA, Multhaup G, McDonald BL, Beyreuther K (1985) Amyloid plaque core protein in Alzheimer disease and Down syndrome. Proc Natl Acad Sci USA 82:4245–4249
Matsumura S, Shinoda K, Yamada M, Yokojima S, Inoue M, Ohnishi T, Shimada T, Kikuchi K, Masui D, Hashimoto S, Sato M, Ito A, Akioka M, Takagi S, Nakamura Y, Nemoto K, Hasegawa Y, Takamoto H, Inoue H, Nakamura S, Nabeshima Y, Teplow DB, Kinjo M, Hoshi M (2011) Two distinct amyloid beta-protein (Abeta) assembly pathways leading to oligomers and fibrils identified by combined fluorescence correlation spectroscopy, morphology, and toxicity analyses. J Biol Chem 286:11555–11562
Mc Donald JM, Savva GM, Brayne C, Welzel AT, Forster G, Shankar GM, Selkoe DJ, Ince PG, Walsh DM (2010) The presence of sodium dodecyl sulphate-stable Abeta dimers is strongly associated with Alzheimer-type dementia. Brain 133:1328–1341
McKenzie KJ, McLellan DR, Gentleman SM, Maxwell WL, Gennarelli TA, Graham DI (1996) Is beta-APP a marker of axonal damage in short-surviving head injury? Acta Neuropathol 92:608–613
McKhann GM, Knopman DS, Chertkow H, Hyman BT, Jack CR Jr, Kawas CH, Klunk WE, Koroshetz WJ, Manly JJ, Mayeux R, Mohs RC, Morris JC, Rossor MN, Scheltens P, Carrillo MC, Thies B, Weintraub S, Phelps CH (2011) The diagnosis of dementia due to Alzheimer’s disease: recommendations from the National Institute on Aging–Alzheimer’s Association workgroups on diagnostic guidelines for Alzheimer’s disease. Alzheimers Dement 7:263–269
Meinhardt J, Sachse C, Hortschansky P, Grigorieff N, Fändrich M (2009) Abeta(1–40) fibril polymorphism implies diverse interaction patterns in amyloid fibrils. J Mol Biol 386:869–877
Meyer-Luehmann M, Coomaraswamy J, Bolmont T, Kaeser S, Schaefer C, Kilger E, Neuenschwander A, Abramowski D, Frey P, Jaton AL, Vigouret JM, Paganetti P, Walsh DM, Mathews PM, Ghiso J, Staufenbiel M, Walker LC, Jucker M (2006) Exogenous induction of cerebral beta-amyloidogenesis is governed by agent and host. Science 313:1781–1784
Meyer-Luehmann M, Stalder M, Herzig MC, Kaeser SA, Kohler E, Pfeifer M, Boncristiano S, Mathews PM, Mercken M, Abramowski D, Staufenbiel M, Jucker M (2003) Extracellular amyloid formation and associated pathology in neural grafts. Nat Neurosci 6:370–377
Milton NG (2001) Phosphorylation of amyloid-beta at the serine 26 residue by human cdc2 kinase. NeuroReport 12:3839–3844
Mirra SS, Heyman A, McKeel D, Sumi SM, Crain BJ, Brownlee LM, Vogel FS, Hughes JP, van Belle G, Berg L (1991) The consortium to establish a registry for Alzheimer’s disease (CERAD). Part II. Standardization of the neuropathologic assessment of Alzheimer’s disease. Neurology 41:479–486
Montine TJ, Phelps CH, Beach TG, Bigio EH, Cairns NJ, Carrillo MC, Dickson DW, Duyckaerts C, Frosch MP, Masliah E, Mirra SS, Nelson PT, Schneider JA, Thal DR, Thies B, Trojanowski JQ, Vinters HV, Hyman BT (2012) National Institute on Aging–Alzheimer’s Association guidelines for the neuropathologic assessment of Alzheimer’s disease: a practical approach. Acta Neuropathol 123:1–11
Morgado I, Fändrich M (2011) Assembly of Alzheimer’s Aβ peptide into nanostructured amyloid fibrils. Curr Opin Colloid Interface Sci 16:508–514
Mori H, Takio K, Ogawara M, Selkoe DJ (1992) Mass spectrometry of purified amyloid beta protein in Alzheimer’s disease. J Biol Chem 267:17082–17086
Necula M, Kayed R, Milton S, Glabe CG (2007) Small molecule inhibitors of aggregation indicate that amyloid beta oligomerization and fibrillization pathways are independent and distinct. J Biol Chem 282:10311–10324
Phinney AL, Deller T, Stalder M, Calhoun ME, Frotscher M, Sommer B, Staufenbiel M, Jucker M (1999) Cerebral amyloid induces aberrant axonal sprouting and ectopic terminal formation in amyloid precursor protein transgenic mice. J Neurosci 19:8552–8559
Price JL, Davis PB, Morris JC, White DL (1991) The distribution of tangles, plaques and related immunohistochemical markers in healthy aging and Alzheimer’s disease. Neurobiol Aging 12:295–312
Prokop S, Miller KR, Heppner FL (2013) Microglia actions in Alzheimer’s disease. Acta Neuropathol 126:461–477
Rajendran L, Honsho M, Zahn TR, Keller P, Geiger KD, Verkade P, Simons K (2006) Alzheimer’s disease beta-amyloid peptides are released in association with exosomes. Proc Natl Acad Sci USA 103:11172–11177
Reinhard C, Hebert SS, De Strooper B (2005) The amyloid-beta precursor protein: integrating structure with biological function. EMBO J 24:3996–4006
Rezaei-Ghaleh N, Amininasab M, Giller K, Kumar S, Stundl A, Schneider A, Becker S, Walter J, Zweckstetter M (2014) Turn plasticity distinguishes different modes of amyloid-beta aggregation. J Am Chem Soc 136:4913–4919
Rijal Upadhaya A, Capetillo-Zarate E, Kosterin I, Abramowski D, Kumar S, Yamaguchi H, Walter J, Fändrich M, Staufenbiel M, Thal DR (2012) Dispersible amyloid β-protein oligomers, protofibrils, and fibrils represent diffusible but not soluble aggregates: their role in neurodegeneration in amyloid precursor protein (APP) transgenic mice. Neurobiol Aging 33:2641–2660
Rijal Upadhaya A, Kosterin I, Kumar S, Von Arnim C, Yamaguchi H, Fändrich M, Walter J, Thal DR (2014) Biochemical stages of amyloid β-peptide aggregation and accumulation in the human brain and their association with symptomatic and pathologically-preclinical Alzheimer’s disease. Brain 137:887–903
Rijal Upadhaya A, Lungrin I, Yamaguchi H, Fändrich M, Thal DR (2012) High-molecular weight Aβ-oligomers and protofibrils are the predominant Aβ-species in the native soluble protein fraction of the AD brain. J Cell Mol Med 16:287–295
Rijal Upadhaya A, Scheibe F, Kosterin I, Abramowski D, Gerth J, Kumar S, Liebau S, Yamaguchi H, Walter J, Staufenbiel M, Thal DR (2013) The type of Abeta-related neuronal degeneration differs between amyloid precursor protein (APP23) and amyloid beta-peptide (APP48) transgenic mice. Acta Neuropathol Commun 1:77
Roher AE, Lowenson JD, Clarke S, Wolkow C, Wang R, Cotter RJ, Reardon IM, Zurcher-Neely HA, Heinrikson RL, Ball MJ, Greenberg BD (1993) Structural alterations in the peptide backbone of beta-amyloid core protein may account for its deposition and stability in Alzheimer’s disease. J Biol Chem 268:3072–3083
Roher AE, Lowenson JD, Clarke S, Woods AS, Cotter RJ, Gowing E, Ball MJ (1993) beta-Amyloid-(1–42) is a major component of cerebrovascular amyloid deposits: implications for the pathology of Alzheimer disease. Proc Natl Acad Sci USA 90:10836–10840
Rolyan H, Feike AC, Upadhaya AR, Waha A, Van Dooren T, Haass C, Birkenmeier G, Pietrzik CU, Van Leuven F, Thal DR (2011) Amyloid-beta protein modulates the perivascular clearance of neuronal apolipoprotein E in mouse models of Alzheimer’s disease. J Neural Transm 118:699–712
Sachse C, Fändrich M, Grigorieff N (2008) Paired beta-sheet structure of an Abeta(1–40) amyloid fibril revealed by electron microscopy. Proc Natl Acad Sci USA 105:7462–7466
Sachse C, Grigorieff N, Fändrich M (2010) Nanoscale flexibility parameters of Alzheimer amyloid fibrils determined by electron cryo-microscopy. Angew Chem Int Ed Engl 49:1321–1323
Sachse C, Xu C, Wieligmann K, Diekmann S, Grigorieff N, Fändrich M (2006) Quaternary structure of a mature amyloid fibril from Alzheimer’s Abeta(1–40) peptide. J Mol Biol 362:347–354
Saido TC, Iwatsubo T, Mann DM, Shimada H, Ihara Y, Kawashima S (1995) Dominant and differential deposition of distinct beta-amyloid peptide species, A beta N3(pE), in senile plaques. Neuron 14:457–466
Saido TC, Yamao-Harigaya W, Iwatsubo T, Kawashima S (1996) Amino- and carboxyl-terminal heterogeneity of beta-amyloid peptides deposited in human brain. Neurosci Lett 215:173–176
Sanchez L, Madurga S, Pukala T, Vilaseca M, Lopez-Iglesias C, Robinson CV, Giralt E, Carulla N (2011) Abeta40 and Abeta42 amyloid fibrils exhibit distinct molecular recycling properties. J Am Chem Soc 133:6505–6508
Scheidt HA, Morgado I, Rothemund S, Huster D, Fändrich M (2011) Solid-state NMR spectroscopic investigation of Abeta protofibrils: implication of a beta-sheet remodeling upon maturation into terminal amyloid fibrils. Angew Chem Int Ed Engl 50:2837–2840
Schilling S, Lauber T, Schaupp M, Manhart S, Scheel E, Bohm G, Demuth HU (2006) On the seeding and oligomerization of pGlu-amyloid peptides (in vitro). Biochemistry 45:12393–12399
Schilling S, Zeitschel U, Hoffmann T, Heiser U, Francke M, Kehlen A, Holzer M, Hutter-Paier B, Prokesch M, Windisch M, Jagla W, Schlenzig D, Lindner C, Rudolph T, Reuter G, Cynis H, Montag D, Demuth HU, Rossner S (2008) Glutaminyl cyclase inhibition attenuates pyroglutamate Abeta and Alzheimer’s disease-like pathology. Nat Med 14:1106–1111
Schlenzig D, Manhart S, Cinar Y, Kleinschmidt M, Hause G, Willbold D, Funke SA, Schilling S, Demuth HU (2009) Pyroglutamate formation influences solubility and amyloidogenicity of amyloid peptides. Biochemistry 48:7072–7078
Schmidt M, Sachse C, Richter W, Xu C, Fändrich M, Grigorieff N (2009) Comparison of Alzheimer Abeta(1–40) and Abeta(1–42) amyloid fibrils reveals similar protofilament structures. Proc Natl Acad Sci USA 106:19813–19818
Schulze PC, Thal DR, Bettag M, Schober R (1998) Brain tissue damage and regeneration monitored by β-amyloid precursor protein in experimental laser-induced interstitial thermotherapy. Neuropathology 18:55–61
Shankar GM, Bloodgood BL, Townsend M, Walsh DM, Selkoe DJ, Sabatini BL (2007) Natural oligomers of the Alzheimer amyloid-beta protein induce reversible synapse loss by modulating an NMDA-type glutamate receptor-dependent signaling pathway. J Neurosci 27:2866–2875
Shankar GM, Li S, Mehta TH, Garcia-Munoz A, Shepardson NE, Smith I, Brett FM, Farrell MA, Rowan MJ, Lemere CA, Regan CM, Walsh DM, Sabatini BL, Selkoe DJ (2008) Amyloid-beta protein dimers isolated directly from Alzheimer’s brains impair synaptic plasticity and memory. Nat Med 14:837–842
Shapira R, Austin GE, Mirra SS (1988) Neuritic plaque amyloid in Alzheimer’s disease is highly racemized. J Neurochem 50:69–74
Shimizu T, Fukuda H, Murayama S, Izumiyama N, Shirasawa T (2002) Isoaspartate formation at position 23 of amyloid beta peptide enhanced fibril formation and deposited onto senile plaques and vascular amyloids in Alzheimer’s disease. J Neurosci Res 70:451–461
Small SA, Duff K (2008) Linking Abeta and tau in late-onset Alzheimer’s disease: a dual pathway hypothesis. Neuron 60:534–542
Sperling RA, Aisen PS, Beckett LA, Bennett DA, Craft S, Fagan AM, Iwatsubo T, Jack CR Jr, Kaye J, Montine TJ, Park DC, Reiman EM, Rowe CC, Siemers E, Stern Y, Yaffe K, Carrillo MC, Thies B, Morrison-Bogorad M, Wagster MV, Phelps CH (2011) Toward defining the preclinical stages of Alzheimer’s disease: recommendations from the National Institute on Aging–Alzheimer’s Association workgroups on diagnostic guidelines for Alzheimer’s disease. Alzheimers Dement 7:280–292
Spires TL, Meyer-Luehmann M, Stern EA, McLean PJ, Skoch J, Nguyen PT, Bacskai BJ, Hyman BT (2005) Dendritic spine abnormalities in amyloid precursor protein transgenic mice demonstrated by gene transfer and intravital multiphoton microscopy. J Neurosci 25:7278–7287
Stohr J, Condello C, Watts JC, Bloch L, Oehler A, Nick M, DeArmond SJ, Giles K, DeGrado WF, Prusiner SB (2014) Distinct synthetic Abeta prion strains producing different amyloid deposits in bigenic mice. Proc Natl Acad Sci USA 111:10329–10334
Takahashi RH, Capetillo-Zarate E, Lin MT, Milner TA, Gouras GK (2010) Co-occurrence of Alzheimer’s disease beta-amyloid and tau pathologies at synapses. Neurobiol Aging 31:1145–1152
Tanzi RE (2012) The genetics of Alzheimer disease. Cold Spring Harb Perspect Med 2:a006296
Terry RD (2000) Cell death or synaptic loss in Alzheimer disease. J Neuropathol Exp Neurol 59:1118–1119
Terzi E, Holzemann G, Seelig J (1997) Interaction of Alzheimer beta-amyloid peptide(1–40) with lipid membranes. Biochemistry 36:14845–14852
Thal DR, Arendt T, Waldmann G, Holzer M, Zedlick D, Rüb U, Schober R (1998) Progression of neurofibrillary changes and PHF-tau in end-stage Alzheimer’s disease is different from plaque and cortical microglial pathology. Neurobiol Aging 19:517–525
Thal DR, Attems J, Ewers M (2014) Spreading of amyloid, tau, and microvascular pathology in Alzheimer’s disease: findings from neuropathological and neuroimaging studies. J Alzheimers Dis 42:S421–S429
Thal DR, Ghebremedhin E, Orantes M, Wiestler OD (2003) Vascular pathology in Alzheimer’s disease: correlation of cerebral amyloid angiopathy and arteriosclerosis/lipohyalinosis with cognitive decline. J Neuropathol Exp Neurol 62:1287–1301
Thal DR, Glas A, Schneider W, Schober R (1997) Differential pattern of beta-amyloid, amyloid precursor protein and apolipoprotein E expression in cortical senile plaques. Acta Neuropathol (Berl) 94:255–265
Thal DR, Härtig W, Schober R (1998) Stage-correlated distribution of type 1 and 2 dystrophic neurites in cortical and hippocampal plaques in Alzheimer’s disease. J Hirnforsch 39:175–181
Thal DR, Rüb U, Orantes M, Braak H (2002) Phases of Abeta-deposition in the human brain and its relevance for the development of AD. Neurology 58:1791–1800
Thal DR, Rüb U, Schultz C, Sassin I, Ghebremedhin E, Del Tredici K, Braak E, Braak H (2000) Sequence of Abeta-protein deposition in the human medial temporal lobe. J Neuropathol Exp Neurol 59:733–748
Thal DR, von Arnim C, Griffin WS, Yamaguchi H, Mrak RE, Attems J, Rijal Upadhaya A (2013) Pathology of clinical and preclinical Alzheimer’s disease. Eur Arch Psychiatry Clin Neurosci 263(Suppl 2):S137–S145
Tsai J, Grutzendler J, Duff K, Gan WB (2004) Fibrillar amyloid deposition leads to local synaptic abnormalities and breakage of neuronal branches. Nat Neurosci 7:1181–1183
Vandenberghe R, Van Laere K, Ivanoiu A, Salmon E, Bastin C, Triau E, Hasselbalch S, Law I, Andersen A, Korner A, Minthon L, Garraux G, Nelissen N, Bormans G, Buckley C, Owenius R, Thurfjell L, Farrar G, Brooks DJ (2010) 18F-flutemetamol amyloid imaging in Alzheimer disease and mild cognitive impairment: a phase 2 trial. Ann Neurol 68:319–329
Villemagne VL, Pike KE, Darby D, Maruff P, Savage G, Ng S, Ackermann U, Cowie TF, Currie J, Chan SG, Jones G, Tochon-Danguy H, O’Keefe G, Masters CL, Rowe CC (2008) Abeta deposits in older non-demented individuals with cognitive decline are indicative of preclinical Alzheimer’s disease. Neuropsychologia 46:1688–1697
Vitalis A, Caflisch A (2010) Micelle-like architecture of the monomer ensemble of Alzheimer’s amyloid-beta peptide in aqueous solution and its implications for Abeta aggregation. J Mol Biol 403:148–165
Vonsattel JP, Myers RH, Hedley-Whyte ET, Ropper AH, Bird ED, Richardson EP Jr (1991) Cerebral amyloid angiopathy without and with cerebral hemorrhages: a comparative histological study. Ann Neurol 30:637–649
Vos SJ, Xiong C, Visser PJ, Jasielec MS, Hassenstab J, Grant EA, Cairns NJ, Morris JC, Holtzman DM, Fagan AM (2013) Preclinical Alzheimer’s disease and its outcome: a longitudinal cohort study. Lancet Neurol 12:957–965
Walsh DM, Selkoe DJ (2007) A beta oligomers—a decade of discovery. J Neurochem 101:1172–1184
Walter J, van Echten-Deckert G (2013) Cross-talk of membrane lipids and Alzheimer-related proteins. Mol Neurodegener 8:34
Wang D, Munoz DG (1995) Qualitative and quantitative differences in senile plaque dystrophic neurites of Alzheimer’s disease and normal aged brain. J Neuropathol Exp Neurol 54:548–556
Watts JC, Condello C, Stohr J, Oehler A, Lee J, DeArmond SJ, Lannfelt L, Ingelsson M, Giles K, Prusiner SB (2014) Serial propagation of distinct strains of Abeta prions from Alzheimer’s disease patients. Proc Natl Acad Sci USA 111:10323–10328
Weekman EM, Sudduth TL, Abner EL, Popa GJ, Mendenhall MD, Brothers HM, Braun K, Greenstein A, Wilcock DM (2014) Transition from an M1 to a mixed neuroinflammatory phenotype increases amyloid deposition in APP/PS1 transgenic mice. J Neuroinflammation 11:127
Weller RO, Massey A, Newman TA, Hutchings M, Kuo YM, Roher AE (1998) Cerebral amyloid angiopathy: amyloid beta accumulates in putative interstitial fluid drainage pathways in Alzheimer’s disease. Am J Pathol 153:725–733
Wirths O, Breyhan H, Cynis H, Schilling S, Demuth HU, Bayer TA (2009) Intraneuronal pyroglutamate-Abeta 3–42 triggers neurodegeneration and lethal neurological deficits in a transgenic mouse model. Acta Neuropathol 118:487–496
Wirths O, Multhaup G, Czech C, Blanchard V, Moussaoui S, Tremp G, Pradier L, Beyreuther K, Bayer TA (2001) Intraneuronal Abeta accumulation precedes plaque formation in beta-amyloid precursor protein and presenilin-1 double-transgenic mice. Neurosci Lett 306:116–120
Wisniewski HM, Terry RD (1973) Reexamination of the pathogenesis of the senile plaque. In: Zimmerman HM (ed) Progress in neuropathology, vol 2. Grane & Stratton, New York, pp 1–26
Wyss-Coray T, Loike JD, Brionne TC, Lu E, Anankov R, Yan F, Silverstein SC, Husemann J (2003) Adult mouse astrocytes degrade amyloid-beta in vitro and in situ. Nat Med 9:453–457
Yamaguchi H, Sugihara S, Ogawa A, Saido TC, Ihara Y (1998) Diffuse plaques associated with astroglial amyloid beta protein, possibly showing a disappearing stage of senile plaques. Acta Neuropathol 95:217–222
Acknowledgments
The authors thank Bill Close (Institute for Pharmaceutical Biotechnology, Center for Biomedical Research, University of Ulm, Germany) for reading the manuscript. This study was supported by DFG-Grants TH624/6-1 (DRT), WA1477/6 (JW), FA456/12-1 (MF) and Alzheimer Forschung Initiative Grants #10810, 13803 (DRT).
Conflict of interest
DRT received consultancies from Covance Laboratories (UK) and GE-Healthcare (UK), received a speaker honorarium from GE-Healthcare (UK) and collaborated with Novartis Pharma Basel (Switzerland).
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article

Cite this article
Thal, D.R., Walter, J., Saido, T.C. et al. Neuropathology and biochemistry of Aβ and its aggregates in Alzheimer’s disease. Acta Neuropathol 129, 167–182 (2015). https://doi.org/10.1007/s00401-014-1375-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00401-014-1375-y