
Abstract
Purpose
Individuals with vitamin D (VD) insufficiency have a greater tendency to develop obesity and have increased systemic inflammation. Gut microbiota are involved in the regulation of host inflammation and energy metabolism, which plays a role in the pathogenesis of obesity. Thus, we aimed to evaluate the effects of different doses of VD3 on body weight, serum lipids, inflammatory factors, and intestinal barrier function in obese mice and to explore the regulatory effect of VD3 on gut microbiota in obese mice.
Methods
Male C57BL/6 J mice received a normal chow diet (NCD, 10% fat) or high-fat diet (HFD, 60% fat) to induce obesity within 10 weeks. Then, HFD mice were supplemented with 5650, 8475, or 11,300 IU VD3/kg diet for 8 weeks. Finally, 16 s rRNA analysis was performed to analyze gut microbiota composition in cecal contents. In addition, body weight, serum lipids, inflammatory factors, and intestinal barrier function were analyzed.
Results
VD3 supplementation reduced body weight and the levels of TG, TC, HDL-C, TNF-α, IL-1β and LPS, and increased ZO-1 in HFD-fed mice. Moreover, it increased α-diversity, reduced F/B ratio and altered microbiota composition by increasing relative abundance of Bacteroidetes, Proteobacteria, Desulfovibrio, Dehalobacterium, Odoribacter, and Parabacteroides and reducing relative abundance of Firmicutes and Ruminococcus. There were significant differences between HFD and NCD groups in several metabolic pathways, including endotoxin biosynthesis, tricarboxylic acid cycle, lipid synthesis and metabolism, and glycolysis.
Conclusions
Low, medium, and high doses of VD3 inhibited weight gain, reduced levels of blood lipids and inflammatory factors, and improved endotoxemia and gut barrier function in obese mice. It also increased the α-diversity of gut microbiota in obese mice and reduced the relative abundance of some intestinal pathogenic bacteria, increased the relative abundance of some beneficial bacteria, and corrected the intestinal flora disorder of obese mice, with the low- and high-dose groups showing better effects than the medium-dose group.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Obesity is defined as excess lipid accumulation caused by an imbalance between energy intake and consumption. It is one of the main risk factors of type 2 diabetes, hyperlipidemia, cardio-cerebrovascular disease, non-alcoholic fatty liver disease, hypertension, and other related metabolic diseases, making it a major public health problem [1, 2]. The prevalence of obesity and overweight is extremely high worldwide, and it is projected that more than 1 billion people will suffer from obesity by 2030 [3]. Obesity is related to lifestyle, diet, psychological and genetics factors, among which lifestyle and diet are the principal factors. Nevertheless, losing weight through physical exercise requires much time and a stable volition, and weight-loss pills have side effects. Consequently, other secure and feasible measures are of utmost need to prevent or amend obesity.
Obesity is linked to low plasma 25-hydroxy vitamin D3 (25(OH)D3) levels [4, 5]. A previous cross-sectional study demonstrated that the body mass index (BMI) is inversely related to plasma 25(OH)D3 concentrations [6]. However, it remains unclear whether a causal relationship exists between BMI and 25(OH)D3 [7]. To date, the findings from intervention studies remain equivocal, as some studies indicated a positive influence of VD3 on obesity and chronic inflammation [8], while other studies did not observe this effect [9, 10]. In our previous researches, our findings suggested a protective effect of 1,25(OH)2D3 in diabetic retinopathy, due to its immunomodulatory property [11]. Moreover, 1,25(OH)2D3 inhibited lipogenesis and decreased the expression of interleukin-6 (IL-6), tumor necrosis factor-α (TNF-α), and monocyte chemoattractant protein-1 (MCP-1) in the hypertrophic adipocytes [12]. Therefore, VD has potential anti-obesity effects.
Recently, many studies have reported that gut microbiota plays a significant role in the regulation of host inflammatory reactions, energy metabolism, and substrate metabolism, which is involved in the pathogenesis of obesity [13]. The constitution of gut microbiota between normal-weight and obese differ in that the Alpha-diversity (α-diversity) is reduced of individuals with obesity [14]. Obesity is associated with alterations in the gut microbiota composition (dysbiosis) and systemic low-grade inflammation [15, 16]. Chronic inflammation caused by metabolic disorders, known as “metabolic inflammation”, is caused by excessive dietary lipids and is characterized by low-level systemic or local inflammatory responses [17, 18]. Gut microbiota disruption or dysbiosis is marked by increased intestinal permeability and intestinal barrier function impairment, leading to the increased production of lipopolysaccharides (LPS) from gram-negative bacteria [19]. LPS can enter the mesenteric vein and circulation and act on target organs and tissues, increasing the risk of inflammation and causing endotoxemia [20].
The gut microbiota is considered to play a key role in the gut barrier and intestinal immune function, which in turn affects the health of the host [15, 21]. Animal studies have demonstrated the link between changes in the constitution of gut microbiota and its metabolites, and the occurrence of obesity [22, 23]. Numerous studies have shown that gut microbiota dysbiosis, particularly the ratio of Firmicutes to Bacteroidetes (F/B ratio), is strongly associated with the occurrence of obesity [24]. Thus, examining the constitution of gut microbiota could reveal novel targets of anti-obesity treatment. Current evidence suggests that the effects of VD intake on health are partially mediated by the gut microbiota. For instance, VD plays a protective role against colorectal cancer, mainly by regulating the Akkermansia muciniphila content in gut microbiota, which contributes to maintaining the integrity of the colon barrier [25]. A recent study showed that maternal VD deficiency during pregnancy and lactation could aggravate gut microbiota dysbiosis and enhance the progression of obesity among male offspring, which might be regulated by genes associated with barrier function, inflammation, and lipid transportation [26]. However, few studies have explored the effect of regulating the gut microbiota composition through VD supplementation in an obesity model, and it is uncertain whether the effect of VD on obesity is mediated by gut microbiota. The purpose of the study was to ascertain the profitable impacts of VD3 on obese mice induced by HFD as well as the improvement of inflammation and the regulatory role on intestinal gut microbiota. The findings would be relevant for making additional use of VD and delivering a beneficial therapy for obesity.
Materials and methods
Animals and experimental designs
Five-week-old male C57BL/6 J mice were purchased from Huafukang Biotechnology Co. Ltd (Beijing, China) and housed in a specific pathogen-free (SPF) room at the Laboratory Animal Center, Southwest Medical University in an environmentally controlled temperature (23 ± 3 °C), with relative humidity of 50 ± 10%, and a 12-h light/dark cycle. The mice were fed with food and water, and their pad material was replaced every 3 days. After 5 days, they (17.08 ± 0.52 g) were randomly divided into two groups: a normal chow-diet group (NCD; n = 10) and a high-fat diet group (HFD; n = 52). The NCD group was fed with a normal chow diet (TP23302, 10.0% fat, 19.0% protein, 71.0% carbohydrate, caloric value 3.6 kcal·kg−1). The HFD group was fed with HFD (TP23300, 60.0% fat, 19.4% protein, 20.6% carbohydrate, caloric value 5.0 kcal·kg−1) to induce obesity within 10 weeks. After 10 weeks, we selected 40 mice in the HFD group weighing 20% higher than the mean weight in the NCD group and randomly assigned them into four groups [27] (n = 10 per each group). These include the HFD group, HFD_VDL group (HFD diet supplemented with 5650 IU/kg VD3), HFD_VDM group (HFD diet supplemented with 8475 IU/kg VD3), HFD_VDH group (HFD diet supplemented with 11,300 IU/kg VD3) [28, 29]. VD3 was supplemented for 8 more weeks. Body weight was measured once weekly and dietary intake was assessed daily. The rest of the day’s feed was discarded to avoid the fatty acid rancidity and 2–3 g of feed was added to ensure adequate feeding. Feed was purchased from Trophic Animal Feed High-Tech Co., Ltd (Nantong, China). At the end of the experiment, mice were subjected to a 12-h fasting and were anesthetized using pentobarbital sodium (50 mg·kg−1·BW). Blood was collected through cardiac puncture, and serum was separated through centrifugation at 3500 rpm for 12 min and stored at – 80 °C. The cecal contents and epididymal white adipose tissue were collected, weighed, frozen in liquid nitrogen and transferred to – 80 °C refrigerator. All procedures were approved by the Animal Ethics Committee of Southwest Medical University (approval number: 2020716).
Biochemical analysis
Serum glucose, triglyceride (TG), total cholesterol (TC), low-density lipoprotein cholesterol (LDL-C) and high-density lipoprotein cholesterol (HDL-C) were measured using an automatic biochemical analyzer (Mindray, BS-240vet, Shenzhen, China).
ELISA analysis
The serum level of TNF-α (E-MSEL-M0002, Elabscience, Wuhan, China), IL-1β (E-MSEL-M0003, Elabscience, Wuhan, China), MCP-1 (E-EL-M3001, Elabscience, Wuhan, China), LPS (CSB-E13066m, cusabio, Wuhan, China), 25(OH)D3 (E-EL-0015, Elabscience, Wuhan, China) and 25(OH)D3 (ml038442, Enzyme-linked Biotechnology Co.,Ltd, Shanghai, China) in eWAT were determined using enzyme-linked immunosorbent assay (ELISA) kits in compliance with the manufacturer’s instructions.
RNA extraction and quantitative real-time PCR
Total RNA was extracted from the cecum, using RNAiso Plus (Takara Bio, Japan), and cDNA was synthesized through a thermal cycle (Eppendorf, Germany), using a PrimeScript™ RT reagent Kit with gDNA Eraser (Takara Bio, Japan). The quantitative real-time PCR (qRT-PCR) was conducted on a Real-time PCR system (Analytik Jenav, Germany). Each PCR mixture contained synthesized cDNA, TB Green Premix Ex Taq (Takara Bio, Japan), and specific primers for the target gene. The sequences of the primers were listed in Supplementary Table 1. The relative expression levels of the genes were calculated with the 2−ΔΔCt method, using glyceraldehyde 3-phosphate dehydrogenase (GAPDH) as an endogenous reference. Specificity for each designed primer was affirmed by Primer-BLAST in National Center for Biotechnology Information (NCBI) and using melt curve analysis. All primers were synthesized by Shenggong (Shanghai, China).
16 s rRNA analysis
We performed 16 s rRNA analysis of the cecal content samples in Lilai Biotechnology Co., Ltd (Chengdu, Szechwan, China). Total DNA from cecal content bacteria was extracted and DNA was quantified using a NanoDrop spectrophotometer (NanoDrop 8000, Thermo, America). The quality of the extraction was assessed through 1.2% agarose gel electrophoresis. Corresponding primers were designed according to the conservative region (16SV3-V4) of the sequence to amplify the PCR. Sequencing libraries were generated using TruSeq Nano DNA LT Library Prep Kit from Illumina (San Diego, CA, USA) according to the manufacturer’s instructions. The quality of the libraries was evaluated using an Agilent High-Sensitivity DNA Kit on Agilent Bioanalyzer. Illumina MiSeq/NovaSeq platform was used for paired-end sequencing of community DNA fragment. Finally, the QIIME software, PICRUSt2 (phylogenetic investigation of communities by reconstruction of unobserved states) software and R language were performed to analyze gut microbiota.
Statistical analysis
We performed statistical analyses using SPSS Statistics version 26 (IBM SPSS Statistics, Chicago, IL, USA). Normally distributed variables were expressed as the mean ± SEM; otherwise, it is expressed as median and interquartile quartile. Differences among pairwise groups were trialed using student’s t test, and multiple comparisons were performed using ANOVA with post hoc Bonferroni’s correction for statistical significance. The statistical significance of differentially abundant and biologically relevant taxonomical biomarkers between two distinct conditions was measured using a linear discriminant analysis (LDA) effect size (LEfSe). GraphPad Prism 8.02 software was used for data mapping charts. A two-tailed p value < 0.05 was considered statistically significant.
Results
VD3 supplementation inhibits HFD-induced body weight increase and hyperlipidemia in mice
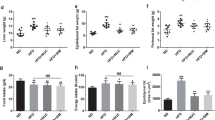
At week 0, there was no difference in body weight between the groups. At the 10th week, the success rate of obesity modeling was 76.92%. After HFD feeding for 18 weeks, body weight significantly increased compared to NCD group (P < 0.001) (Fig. 1A). Three doses of VD3 significantly inhibited body-weight increase of mice fed with HFD between the 10th and 18th week of study (VDL, P < 0.001, VDM, P = 0.007 and VDH, P = 0.015) (Fig. 1A). Besides, HFD feeding resulted in a substantial increase in serum glucose, TG, TC and HDL-C levels as compared to values in the NCD group (Fig. 1B–F). Hyperlipidemia is generally a risk factor for metabolic disease. We observed that levels of serum TG, TC and HDL-C significantly dropped upon VD3 supplementation (Fig. 1C, D and F). There was no difference in serum levels of LDL-C on HFD feeding before and after VD3 supplementation (Fig. 1E). In addition, there was no statistically significant difference in serum glucose under VD3 supplementation (Fig. 1B). These indicated that VD3 supplementation led to a decrease in body weight and hyperlipidemia induced by HFD feeding.

VD3 supplementation inhibits HFD-induced body weight increase and hyperlipidemia in mice. A Body weight. B Levels of serum glucose. C Levels of serum TG. D Levels of serum TC. E Levels of serum LDL-C. F Levels of serum HDL-C. Data are presented as mean ± SEM. In Fig. 1A, HFD group compared to NCD group, ***P < 0.001; HFD_VDL group compared to HFD group, #P < 0.05, ##P < 0.01, ###P < 0.001; HFD_VDM group compared to HFD group, &P < 0.05, &P < 0.01; HFD_VDH group compared to HFD group, $P < 0.05. In Fig. 1B–F, *P < 0.05, ***P < 0.001
VD3 intake and the concentrations of VD3 metabolites
VD3 intake, serum and epididymal adipose tissue 25(OH)D3 concentrations were measured to explore the effects of administering different doses of VD3. As shown in Fig. 2A, the average food intake in the HFD group was significantly decreased compared with that of the NCD group. The effect of VD3 supplementation was not dose-dependent when compared with HFD mice. However, the concentrations of VD3 metabolites in the HFD_VDM group were substantially higher than that in the HFD_VDL or HFD_VDH group. The average daily VD3 intake was calculated during the study. The average VD3 intake was significantly higher in the three supplementation groups than in the HFD group in a dose–response manner (Fig. 2B). Then, we measured the 25(OH)D3 concentrations in serum and epididymal adipose tissue. The serum 25(OH)D3 concentration in HFD group was significantly lower than that in the NCD group (Fig. 2C). However, the 25(OH)D3 concentration in serum and epididymal adipose tissue in the HFD_VDM group were significantly higher than that in the HFD group (Fig. 2C, D).

VD3 intake and the concentrations of VD3 metabolites. A Average food intake. B Average VD3 intake. C Concentrations of serum 25(OH)D3. D Concentrations of epididymal adipose tissue 25(OH)D3. Data are presented as mean ± SEM. *P < 0.05, **P < 0.01, ***P < 0.001
VD3 inhibits the expression of proinflammatory cytokines in serum and caecum of HFD-fed mice
One of the main features of obesity is chronic low-grade systemic inflammation [30]. To examine the anti-inflammatory effects of VD3, serum pro-inflammatory cytokines such as TNF-α, IL-1β, and MCP-1 were measured. The results showed that mice in the HFD group had significantly higher levels of TNF-α, IL-1β and MCP-1 in serum (Fig. 3A, B and C). However, serum TNF-α and IL-1β levels were lower after VD3 supplementation than HFD group (Fig. 3A, B). Moreover, HFD greatly enhanced the mRNA expression of IL-1β in the caecum (Fig. 3E) but did not alter the TNF-α and MCP-1 levels (Fig. 3D, F). On the contrary, the mRNA expressions of TNF-α decreased in the HFD_VDH group (Fig. 3D). In addition, the mRNA expression of IL-1β decreased considerably in the caecum of the three VD3 supplementation groups compared to that in the HFD group (Fig. 3E).

VD3 inhibits the expression of proinflammatory cytokines in serum and caecum of HFD-fed mice. A Serum protein levels of TNF-α determined by ELISA. B Serum protein levels of IL-1β determined by ELISA. C Serum protein levels of MCP-1 determined by ELISA. D Relative mRNA expression of caecum TNF-α determined by qRT-PCR. E Relative mRNA expression of caecum IL-1β determined by qRT-PCR. F Relative mRNA expression of caecum MCP-1 determined by qRT-PCR. Data are presented as means ± SEM. *P < 0.05, **P < 0.01, ***P < 0.001
VD3 improves gut barrier function and endotoxemia in HFD-fed mice
Dysbiosis of gut microbiota leads to the damage of the integrity of tight junctions in intestinal epithelial cells. This causes an influx of LPS into the bloodstream, resulting in endotoxemia and systemic inflammation [31]. The serum LPS concentration was measured using ELISA, and we observed a significant increase in the serum LPS concentration in the HFD group compared to NCD group, while the three doses of VD3 considerably reduced LPS level (Fig. 4A). Next, we investigated the mRNA relative expression of the tight junction proteins, zonula occludens (ZO-1) and Occludin, in the caecum using qRT-PCR. As shown in Fig. 4B, HFD significantly reduced the expression of ZO-1 in the caecum, and this reduction was significantly reversed by VD3 supplementation. However, the mRNA relative expression of Occludin in caecum was similar in the HFD and VD3 supplementation groups as showed by qT-PCR analysis (Fig. 4C). Thus, VD3 prevents endotoxemia and protects gut barrier function, manifested by reduced contents of LPS and increased expressions of tight junction proteins (ZO-1).

VD3 improves gut barrier function and prevents endotoxemia in HFD-fed mice. A Serum protein levels of LPS determined using ELISA. B Relative mRNA expression of caecum ZO-1 determined using qRT-PCR. C Relative mRNA expression of caecum Occludin determined using qRT-PCR. Data are presented as mean ± SEM. *P < 0.05, **P < 0.01, ***P < 0.001
VD3 improves the α-diversity of gut microbiota in HFD-induced obese mice
The gut microbiota are involved in the pathogenesis of obesity and related disorders [32]. We performed a 16S rRNA analysis to explore the effect of VD3 supplementation on gut microbiota using the Illumina MiSeq/NovaSeq platform. After eliminating ineligible sequences and clustering the high-quality sequences at 97% similarity level, 193–850 ASVs/OTUs per sample were obtained.
Chao1 estimator (Chao1), Simpson index (Simpson), Shannon diversity index ( Shannon), Pielou’s Evenness index J (Pielou-e), Observed species richness (Observed-species), Faith’s phylogenetic diversity (Faith-pd), and Good’s non-parametric Coverage estimator (Goods-coverage), were used to evaluate the diversity of bacterial species [33]. Chao1 and observed-species were used to characterize richness. Shannon and Simpson were used to characterize diversity. Faith-PD was used to characterize evolution-based diversity. Pielou-e was used to characterize evenness. Goods-coverage was used to characterize the coverage [34]. In Fig. 5A–G, it could be seen that with Chao1, Simpson, Shannon, Pielou-e, Observed-species and Faith-PD considerably reduced, while Goods-coverage was enhanced in the HFD group. This means that HFD resulted in substantial decline in the microbial richness and diversity of the microflora in the caecum. However, the α-diversity of gut microbiota improved with Chao1, Observed-species, Faith-pd, and Goods-coverage indexes prominently reversed in the HFD_VDL group (Fig. 5A, E, F andG). In the HFD_VDH group, the α-diversity prominently reversed with Chao1, Shannon, Pielou-e, Observed-species, and Faith-pd indexes when compared to HFD-induced obese mice (Fig. 5A, C, D, E and F).

VD3 improves the α-diversity on gut microbiota in HFD-induced obese mice. A Chao1, B Simpson, C Shannon, D Pielou-e, E Observed-species, F Faith-pd, G Goods-coverage. Data are presented as boxplot. ***P < 0.001 was compared with the NCD group. And #P < 0.05, ##P < 0.01 were compared with the HFD group
VD3 has a slight trend on improving the beta-diversity (β-diversity) on gut microbiota in HFD-induced obese mice
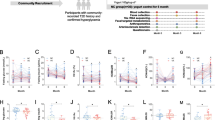
Venn diagrams were plotted to show the mutual bacterial abundance across the groups (Fig. 6A). There are 5540 OTUs obtained in total and 72 OTUs shared in five groups. The NCD group demonstrated the most characteristics taxa with 2424 OTUs, followed by the HFD_VDL, HFD_VDH, HFD_VDM and HFD groups (594, 527, 372 and 318 unique OTUs, respectively). The β-diversity was classified using principal co-ordinate analysis (PCoA), Nonmetric multidimensional scaling (NMDS) and Unweighted pair-group method with arithmetic means (UPGMA) cluster analysis. NMDS is similar to PCoA, but not affected by the numerical value of sample distance. Therefore, in data with complex structure, the sorting result is more stable. The smaller the stress values of NMDS, the better the results. When the value is less than 0.2, the results of NMDS are more reliable. NMDS and UPGMA cluster analysis revealed that the 18-week HFD feeding resulted in a substantial alteration in the gut microbiota as shown by the division between NCD and HFD group. Moreover, the HFD_VDL, HFD_VDM, HFD_VDH groups tended to leave HFD group and approach NCD group (Fig. 6B, C).

VD3 has a slight trend on improving the β-diversity on gut microbiota in HFD-induced obese mice. A Venn, B NMDS 2D sort chart, C UPGMA clustering tree based on sample distance matrix
VD3 improves the gut microbiota disorder induced by HFD at phylum level
Diet is important in the constitution and abundance of gut microbiota. Thus, we assayed the composition of gut microbiota in the trial groups. As demonstrated in Fig. 7A, Firmicutes, Bacteroidetes, and Proteobacteria were the predominant phyla and accounted for over 95% of bacteria. The F/B ratio was significantly higher in the HFD group compared to the HFD_VDL and HFD_VDH groups (Fig. 7B). The abundance of Firmicutes was significantly more in the HFD group than in the NCD group, from 42.29% to 96.13%. Besides, the abundance of Bacteroidetes significantly reduced from 52.65% to 0.71%, Proteobacteria decreased from 2.20% to 1.81%, and TM7 decreased from 0.39% to 0.04%. However, VD3 supplementation significantly decreased the abundance of Firmicutes to 89.54%, 88.80%, and 87.81% in the HFD_VDL, HFD_VDM and HFD_VDH groups, respectively. Furthermore, the abundance of Bacteroidetes was significantly enhanced to 6.80% in the HFD_VDL group and to 4.54% in the HFD_VDH group, as compared to the HFD group. Meanwhile, the abundance of Proteobacteria increased to 4.78% in the HFD_VDM group and to 4.82% in the HFD_VDH group, as compared to the HFD group (P < 0.05).

VD3 improves the gut microbiota disorder induced by HFD at phylum level. A Composition of gut microbiota at phylum level. B Firmicutes/Bacteroidetes, C Relative abundance of Firmicutes, D Relative abundance of Bacteroidetes, E Relative abundance of Proteobacteria, F Relative abundance of TM7. *P < 0.05, **P < 0.01, ***P < 0.001
VD3 improves the gut microbiota disorder induced by HFD at the genus level
At the genus level (Fig. 8), the HFD group obtained 2.62- and 6.59-folds more contents of [Ruminococcus] and unidentified_Clostridiales, and 8.70-, 18.67-, 13.69-, 10.75-, 280.43-, 8.16- and 6.12-folds less abundances of Bacteroides, Odoribacter, Alistipes, Parabacteroides, unidentified_S24-7, unidentified_Ruminococcaceae, and unidentified_Rikenellaceae, respectively, when compared to the NCD group. VD3 improved the gut microbiota dysbiosis by recovering the altered gut microbiota constitutions. Specifically, the level of [Ruminococcus] was decreased, while that of Bacteroides, Odoribacter, Parabacteroides, unidentified_S24-7, unidentified_Ruminococcaceae and unidentified_Rikenellaceae was enhanced by HFD_VDL supplementation compared with that of the HFD group (P < 0.05). Besides, the level of Desulfovibrio, Dehalobacterium and Odoribacter increased after HFD_VDM supplementation compared to the HFD group (P < 0.05). Meanwhile, the abundances of unidentified_ Clostridiales significantly decreased and those of Desulfovibrio, Dehalobacterium, Odoribacter, unidentified_S24-7 and unidentified_Rikenellaceae significantly increased in the HFD_VDH group compared to those in the HD group.

VD3 amends the dysbiosis of gut microbiota induced by HFD at the genus level. A Composition of gut microbiota at genus level, B Desulfovibrio, C [Ruminococcus], [] was used to distinguish the same genus but from different family. In the article, [Ruminococcus] means Ruminococcus from Lachnospiraceae, and Ruminococcus means Ruminococcus from Ruminococcaceae. D Bacteroides, E Dehalobacterium, F Odoribacter, G Alistipes, H Parabacteroides, I unidentified_ Clostridiales, J unidentified_S24-7, K unidentified_Ruminococcaceae, L unidentified_Rikenellaceae. *P < 0.05, **P < 0.01, ***P < 0.001
VD3 improves the biomarker of gut microbiota in HFD-fed mice
To identify the bacteria affected most after HFD and VD3 supplementation, the microbiota constructions within experimental groups were compared using heatmap and LEfSe (LDA > 3.7). Heatmap reveals the calculated relative abundance of gut microbiota at the genus level, as shown in Fig. 9A. The abundances of Allobaculum, Shigella, and [Prevotella] were more highly enriched, while those of Oscillospira, Bacteroides, Parabacteroides, and Odoribacter were impoverished in the HFD group compared to the NCD group. This indicates a disruption of gut symbiosis in HFD-fed mice. In the HFD_VDL group, the dominant bacteria were SMB53, Enterococcus, Bacteroides and Parabacteroides. The abundances of Akkermansia, Sutterella, Desulfovibrio, [Ruminococcus], Bilophila, rc4-4 and Sphingomonas in HFD_VDM group were higher than those in the HFD group. In the HFD_VDH group, the dominant bacteria were Odoribacter, Alistipes, Helicobacter, Desulfovibrio, Roseburia, Lactobacillus, Olsenella, Subdoligranulum, Anoxybacillus, Dorea, Cetobacterium, Dehalobacterium, Clostridium, Adlercreutzia and Streptococcus. In general, there were significant changes in the gut microbiota, and VD3 supplementation reversed the intestinal microbiota disorders caused by HFD. In addition, the results demonstrated that 32 distinctive OTUs (involving 4 phyla, 4 classes, 4 orders, 11 families and 9 genera) represented the significant biomarkers, and the abundance was significantly changed by HFD and VD3 supplementation. As shown in Fig. 9B, C, 13 OTUs were the feature taxa in the NCD group, including Bacteroidetes and TM7 at phylum level and Alistipes, AF12 and Coprococcus at genus level. However, Firmicutes at phylum level had a significant response to HFD intake. Consequently, the pathogeny of HFD-induced disorders is closely related to the changes in the aforementioned bacteria. Bacteroides and Parabacteroides at the genus level in the HFD_VDL group presented as preponderance. Actinobacteria at the phylum level and [Ruminococcus] at the genus level were the principal phenotypes in HFD_VDM group. In the HFD_VDH group, Roseburia, Dehalobacterium and Odoribacter prevailed at the genus level. Therefore, the aforementioned bacteria are potential biomarkers gut microbiota after VD3 supplementation of obese mice.

VD3 influences the biomarkers of gut microbiota in HFD-fed mice. A Heatmap of community abundance at the genus level corresponding to different groups. The blue color or red color, respectively, represents a negative or positive correlation with the microbiota enrichment. B The LEfSe analysis output by Cladogram visualizing. C The most significant diversity in bacterial constitution between groups using LDA. The effect of species abundance on the effect is expressed by the strip length of LDA. The abbreviations: o order, c class, p phylum, f family and g genus
Comparing the metabolic functions of gut microbiota
Dysbiosis of gut microbiota induced by high-fat diet and VD3 supplementation leads to changes in microbial metabolic functions. To investigate this, PICRUSt2 was used to predict the possible changes in functional pathways. As shown in Fig. 10A, the result of Kyoto Encyclopedia of Genes and Genomes (KEGG) secondary functional pathway showed that the relative abundance of gut microbiota exhibited significantly enhanced carbohydrate and amino acid metabolism. In addition, the result of MetaCyc secondary functional pathway showed that the relative abundance of gut microbiota exhibited significantly enhanced cofactor, prosthetic group, electron carrier, vitamin and amino acid biosynthesis (Fig. 10B). After obtaining the abundances of the metabolic pathways, we explored the pathways with significant differences between groups. We found that no pathway was different in the VD3 supplementation groups when compared to the HFD group. However, there were significant differences in the relative abundance of 84 KEGG metabolic pathways and 101 MetaCyc metabolic pathways among the NCD and the HFD groups (P < 0.05). Among the 84 KEGG metabolic pathways, as shown in Fig. 10C, 21 (including endotoxin biosynthesis and tricarboxylic acid cycle) were up-regulated and 63 (including glycerol lipid metabolism and unsaturated fatty acid biosynthesis) were down-regulated in the HFD relative to the NCD group. Among the 101 MetaCyc metabolic pathways, as shown in Fig. 10D, 13 (including lipid biosynthesis and tricarboxylic acid cycle in prokaryotes) were up-regulated, and 88 (including glycolysis and glycogen biosynthesis) were down-regulated in the HFD group relative to the NCD group. The stratified sample metabolic pathway abundance was used to analyze the species composition of the different pathways. The results of species composition in MetaCyc metabolic pathways are shown in Supplement Fig. 1, Bacteroides, [Ruminococcus], Akkermansia, Desulfovibrio, Roseburia, Clostridium, Bilophila, Dehalobacterium, Odoribacter, and Alistipes contributed to the multitudinous metabolic pathways with significant differences in the NCD and HFD groups. However, we did not get the corresponding species composition in KEGG metabolic pathways after analysis.

Comparing metabolic functions of gut microbiota. A KEGG metabolic pathway. B MetaCyc metabolic pathway. C KEGG metabolic pathways in the NCD and HFD groups. The ordinate is different KEGG metabolic pathway labels. D MetaCyc metabolic pathways in the NCD and HFD groups. The ordinate is different MetaCyc metabolic pathway labels. The positive value of logFC represents the upregulation in the HFD group when compared with the NCD group; conversely, the negative value represents the down-regulation. The different degree of significance is expressed in different colors
Discussion
Obesity is becoming one of the most important metabolic disorders worldwide. The morbidity rate of overweight or obesity is consistently on a rise in developed and developing countries [35]. Many studies have revealed that gut microbiota influences the metabolic pathways involved in the pathogenesis of obesity [36]. Some recent meta-analysis demonstrated the association between abdominal obesity and lower serum VD3 levels [4, 37]. However, the mechanism of action of the anti-obesity effect of VD3 is not yet established. In our study, obesity was induced in mice after feeding them with HFD for 10 weeks before VD3 supplementation. Then, we investigated the effect of VD3 supplementation for 8 weeks on gut microbiota of obese C57BL/6 J mice. HFD induced body weight gain, dyslipidemia, systemic chronic inflammation and gut microbiota disorder. These effects were in accordance with previous findings [38,39,40]. Moreover, our present study demonstrated that VD3 mitigates obesity induced by HFD via reducing serum lipid accumulation, improving inflammation, ameliorating endotoxemia and gut barrier function, and stabilizing gut microbiota.
In the study, 18 weeks of HFD feeding caused substantial increases in body weight as well as abnormal lipid accumulation such as higher TG, TC, and HDL-C concentration in serum levels. HDL-C is known as good cholesterol as it clears off the remaining cholesterol in the body, and has a negative correlation with obesity [41]. However, Hayek T et al. [42] found that high-saturated fat and high-cholesterol diets could increase HDL-C by increasing the transport rates and decreasing the fractional catabolic rates of HDL-C ester and apolipoprotein A–I, which might account for the abnormal increase in serum HDL-C concentration of obese mice we also observed. HFD feeding induced the obese phenotype. However, the food intake in the four groups of HFD feeding was significantly lower than that of the NCD group. Perhaps, HFD feeding is prone to feeling of satiety and anorexia, thus reducing the food intake of mice. In addition, fatty rancidity will significantly reduce the food intake in mice. Therefore, higher demand for preservation of HFD feedstuff is needed. We paid close attention to these precautions as we assessed food intake daily and discarded the leftover feedstuff daily to ensure feeding fresh feedstuff. Therefore, improving the taste of HFD feedstuff will confer a better success rate of obesity modeling. VD3 supplementation reduced body weight and hyperlipidemia in obese mice. This reveals a positive influence of VD3 supplementation on body weight and serum lipid levels in obese mice, consistent with previous studies [43]. However, VD3 intervention did not affect glycemia. This was not consistent with a previous study that showed favorable effects of VD3 on glycemic control and cognitive function in rats [44]. Most studies revealed that the profitable influences of VD intervention were primarily targeted at people with VD deficiency [45]. In this study, our mice were not deficient in VD as indicated in the serum 25 (OH)D3 level (Fig. 2C). Therefore, it could be possible to have no profitable effects of VD3 intervention on serum glucose. However, we found that the food intake in HFD_VDM was higher than that in the HFD_VDL and HFD_VDH groups. This could be explained by the taste of HFD and VD3. Consequently, adding VD3 by gavage to make the intake of VD3 more accurate should be considered. Furthermore, the VD3 intake does not represent the actual effective dose, since the HFD_VDM group is the highest effective dose in all groups according to the content of 25(OH)D3 in serum and epididymal adipose tissue. Therefore, the determination of VD3 supplement depends on many factors, not only on the actual intake, and this should be considered in future studies.
Long-term inflammatory response is involved in regulating synthesis and storage of TG in adipose tissue. Hypertrophic adipose cells alter the production of adipocytokines, leading to inflammatory response [46]. Thus, chronic inflammation and lipid synthesis are crucial targets for anti-obesity therapy [47]. We found that HFD feeding enhanced serum inflammatory and mRNA levels in caecum coding for pro-inflammatory factor, consistent with the previous findings [48, 49]. Moreover, VD3 intervention significantly reversed the HFD-induced up-regulation of inflammatory cytokines (TNF-α and IL-1β) in serum and cecal tissues, consistent with previous findings [43, 50]. However, we found that low-, medium-, and high-dose VD3 supplementation did not reduce the level of MCP-1, differing from findings obtained by Chang et al. [51]. They found that intervention with 10,000 IU/kg of VD at 16 weeks significantly reduces the expression of MCP-1 induced by HFD feeding. In this study, VD3 intervention was only performed for 8 weeks, thereby failed to discover the effective inhibitory effect of VD3 on MCP-1.The enteral epithelial barrier is generally sheltered by tight junction proteins, such as ZO-1, occludin, and claudin. They can sustain the integrity of intestinal epithelium, regulate the transport of water and nutrients, and prevent endotoxin and bacteria from entering the systemic circulation from the intestinal cavity [52]. Obesity is associated with dysbiosis and disruption of gut barrier resulting in elevated circulating LPS [53]. LPS is a potent inducer of inflammation. LPS induced by HFD promotes an inflammatory response, as well as bacterial translocation and enhanced gut permeability [54]. The superfluous generation of LPS enhances proinflammatory factors and accelerates the inflammatory response in the intestinal, thereby facilitating the progression of obesity [52]. At the same time, accumulated inflammatory factors inhibit the expression of the tight junction proteins directly, thereby increasing gut permeability [55]. VD deficiency (due to VD deficiency in diet or vitamin D receptor (VDR) knockout) significantly reduces the expression of tight junction proteins (ZO-1 and Occludin) [56, 57]. Our study demonstrated that VD3 supplementation significantly decreased the expression of endotoxin and inverted the decline of the caecum mRNA expression level of ZO-1 caused by HFD feeding, which is in accordance with findings from Liu’s study revealing that exogenous supplementation of VD3 substantially alleviates LPS induced intestinal inflammation by suppressing inflammation and stabilizing tight junctions in the intestine [58]. However, we found that the mRNA level of Occludin in the caecum was similar before and after VD3 supplementation, while VD3 effectively upregulated the expression of Occludin in the intestinal tract, mostly in the colon [44, 59]. Our sample was taken from the caecum for testing, and no beneficial regulatory effect of VD3 on Occludin was observed. In addition, Malaguarnera found that the VD pathway is involved in gut homeostasis [60]. Thus, VD3 exerts anti-obesity effects by inhibiting inflammatory responses and enhancing gut barrier function.
VD regulates the enteric dysbacteriosis and stabilizes intestinal microbiota [61]. In our study, we also found that VD3 has a significant impact on the composition and structure of gut microbiota. In general, NCD and HFD feeding resulted in the substantial alterations in bacterial α-diversity and β-diversity. α-diversity represents taxonomic richness and evenness, and is one of the most common metrics reported in the gut microbiome literature. It is a known marker for microbiome health [34]. After HFD_VDL and HFD_VDH supplementations, the bacterial community was altered, indicating that VD3 affected the gut microbiota of HFD-fed mice. However, HFD_VDM supplementation did not improve the α-diversity of obese mice, and we suspect that excessive VD3 could weaken the improvement of α-diversity. VD3 slightly improved the β-diversity in HFD-fed mice. We could not demonstrate strong findings, possibly because our sample size was inadequate.
VD3 supplementation enhanced the proportion of Bacteroidetes and Proteobacteria, and declined the abundance of Firmicutes and the F/B ratio. Firmicutes are the liveliest bacteria in gut microbiota of people with severe obesity and anomalous raises of the F/B ratio is closely related to obesity [62]. The result indicated that the altered proportion of Firmicutes contributes to the development of obesity induced by HFD according to the LEfSe assay. Our results showed that VD3 supplementation increased the abundance of Bacteroidetes and decreased the relative abundance of Firmicutes in HFD-fed mice, consistent with a previous study [63]. A recent review showed that increased level of Proteobacteria was a potential indicator of dysbiosis and risk of disease [64]. The significant elevation of the abundance of Proteobacteria observed after VD3 supplementation demonstrated the potential disadvantages of excessive VD3 on gut microbiota. Interestingly, at the genus level, VD3 intervention substantially decreased the proportion of [Ruminococcus] and unidentified_ Clostridiales, and enhanced the abundance of Desulfovibrio, Bacteroides, Dehalobacterium, Odoribacter, Parabacteroides, unidentified_S24-7, unidentified_Ruminococcaceae and unidentified_Rikenellaceae. Intestinal dysbiosis featuring abundance of Ruminococcus gnavus is associated with allergic diseases in infants [65]. The colonization of Clostridiales in the intestine is a potential risk factor for colon cancer [66].Desulfovibrio confers resilience to the comorbidity of pain and anxiety in a mouse model of chronic inflammatory pain [67]. Bacteroides is one of the predominant genera in human gut microbiota (20–40%), with beneficial effects in the host. Enhancing the abundance of Bacteroides facilitates digestion, absorption and metabolism of nutrients, and reduces gut inflammation [68]. Bacteroides, Odoribacter, Parabacteroides, Ruminococcaceae and Roseburia are SCFA-producing bacteria [69,70,71]. SCFAs (ethanoate, propionate, butyrate, isobutyric acid and valerate) provide an advantageous environment that inhibits the production of pro-inflammatory factors, and stabilizes the intestinal homeostasis and integrity of the intestinal barrier [72]. Propionate and butyrate are energy sources to colon cells. Studies have shown that they could influence the integrity of the intestinal barrier with tight junction proteins stimulation and mucus secretion, thereby, preventing the onset of diseases, such as obesity, diabetes, inflammatory bowel disease and colorectal cancer [73]. Moreover, ethanoate exerts anti-inflammatory and anti-apoptotic effects and are involved in defending the colon from pathogenic attack as well as prevent colorectal cancer [74]. Prebiotics provide "food" for good bacteria by degrading carbohydrates that normally cannot be digested by the host, converting them into monosaccharides and SCFAs [75]. Dehalobacterium and Rikenellaceae have anti-intestinal inflammatory effects in the study of hericium on alleviating Alzheimer’s disease [76]. In addition, it has been reported that S24_7 affects the metabolism and immune response of the host [77].
Lactobacillus (a probiotic) was significantly enhanced in HFD_VDH group as stated in the heatmap. In the three VD3 supplementation groups, HFD_VDH considerably enhanced the abundance of Lactobacillus, but no difference in the HFD_VDM group and reduced abundance in the HFD_VDL group when compared to the HFD group, although systemic inflammation was remarkably improved in the test groups. VD3 restored HFD-induced gut microbiota dysbiosis by increasing the relative abundance of Lactobacillus and reducing that of Acetatifactor, Oscillibacter and Flavonifractor [63]. In addition, Akkermansia, Bacteroides and Parabacteroides are regarded as intestinal bacteria that generate propanoate and butyrate. The prominent enhancements of Parabacteroides, Bacteroides, and Akkermansia were perceived with the intervention of HFD_VDL or HFD_VDM. Besides, our LEfSe study showed that Clostridiales and Roseburia were the key bacteria among HFD_VDL and HFD_VDH group. Akkermansia is known for its fight against cancer, and can restore the impaired intestinal epithelial barrier, alleviate immune stress and reduce inflammatory response [73]. Liu et al. showed that up-regulated abundance of Alisipes and Rikenella in gut microbiota prevents glucose tolerance, increases SCFAs production and reduces LPS concentration in obese mice [78]. Our heatmap finding was consistent with that in Liu’s study which revealed that the abundance of Alisipes and Rikenella decreased significantly in the HFD group. Our study further demonstrated that VD3 prevents inflammation, enhances intestinal barrier function, and reduces body weight recovering the HFD-altered gut microbiota. Nevertheless, in our study, the regulatory effect of VD3 on gut microbiota was not dose-dependent, and excessive VD3 might confer potential disadvantages. Therefore, the interrelationship of the changes in gut microbiota and the effective anti-obesity dosages of VD3 require further verification.
Most studies on gut microbiota in obesity mainly focus on the composition, diversity and species differences in gut microbiota, and few have explored the relationship between gut microbiota and metabolic pathways. We found that carbohydrate metabolism, lipid biosynthesis and metabolism, amino acid biosynthesis and metabolism, cofactor and vitamin biosynthesis and metabolism, and nucleoside and nucleotide metabolism were significantly different in the NCD and HFD groups. Nevertheless, there was no difference between VD3 and HFD group in terms of metabolic pathways by PICRUSt2 analysis, possibly due to insufficient sample size. From the perspective of obesity, previous researches paid more attention to glycolipid metabolism, especially significant differences in multiple metabolic pathways, such as tricarboxylic acid cycle, glycolysis, lipid synthesis and metabolism. We analyzed the contribution of different bacteria in the metabolic pathways and found that Bacteroides, Akkermansia, [Ruminococcus], Oscillospira, Desulfovibrio, Allobaculum, Alistipes, Odoribacter, Clostridium, Roseburia, unidentified_S24-7, unclassified_Lachnospiraceae and unclassified_Clostridiales were greatly involved in these pathways, which could provide a basis for further research on the development of probiotics and anti-obesity drugs.
Conclusions
Low-, medium- and high-dose VD3 supplementation inhibited body weight gain, prevented hyperlipidemia, reduced levels of TNF-α and IL-1β, improved endotoxemia and preserved gut barrier function in obese mice. Moreover, low- and high-dose VD3 supplementation increased the α-diversity of gut microbiota in obese mice. Furthermore, low-, medium- and high-dose VD3 supplementation reduced the relative abundance of some intestinal pathogenic bacteria, and increased the relative abundance of some beneficial bacteria, which stabilized the intestinal flora disorder often present in obese mice. The low- and high-dose groups showed better stabilizing effects than the medium dose group. Thus, further studies are required to demonstrate and confirm the dose range of VD3 supplementation that exerts anti-obesity and stabilizing effects of the gut microbiota.
Data availability statement
The data presented in this study are available on request from the corresponding author.
References
Lumeng CN, Saltiel AR (2011) Inflammatory links between obesity and metabolic disease. J Clin Investig 121:2111–2117. https://doi.org/10.1172/JCI57132
Singh S, Dulai PS, Zarrinpar A, Ramamoorthy S, Sandborn WJ (2017) Obesity in IBD: epidemiology, pathogenesis, disease course and treatment outcomes. Nat Rev Gastroenterol Hepatol 14:110–121. https://doi.org/10.1038/nrgastro.2016.181
Abarca-Gómez L, Abdeen ZA, Hamid ZA, Abu-Rmeileh NM, Acosta-Cazares B, Acuin C, Adams RJ, Aekplakorn W, Afsana K, Aguilar-Salinas CA et al (2017) Worldwide trends in body-mass index, underweight, overweight, and obesity from 1975 to 2016: a pooled analysis of 2416 population-based measurement studies in 128·9 million children, adolescents, and adults. The Lancet 390:2627–2642. https://doi.org/10.1016/S0140-6736(17)32129-3
Hajhashemy Z, Foshati S, Saneei P (2022) Relationship between abdominal obesity (based on waist circumference) and serum vitamin D levels: a systematic review and meta-analysis of epidemiologic studies. Nutr Rev 80:1105–1117. https://doi.org/10.1093/nutrit/nuab070
Landrier JF, Karkeni E, Marcotorchino J, Bonnet L, Tourniaire F (2016) Vitamin D modulates adipose tissue biology: possible consequences for obesity? Proc Nutr Soc 75:38–46. https://doi.org/10.1017/S0029665115004164
Garcia M, Seelaender M, Sotiropoulos A, Coletti D, Lancha AH (2019) Vitamin D, muscle recovery, sarcopenia, cachexia, and muscle atrophy. Nutrition (Burbank, Los Angeles County, Calif). 60:66–69. https://doi.org/10.1016/j.nut.2018.09.031
Dix CF, Barcley JL, Wright ORL (2018) The role of vitamin D in adipogenesis. Nutr Rev 76:47–59. https://doi.org/10.1093/nutrit/nux056
Lotfi Dizaji L, Mahboob S, Aliashrafi S, Vaghef Mehrabany E, Ebrahimi Mameghani M, Morovati A (2019) Effect of vitamin D supplementation along with weight loss diet on meta-inflammation and fat mass in obese subjects with vitamin D deficiency: A double-blind placebo-controlled randomized clinical trial. Clinical endocrinology (Oxford) 90:94–101. https://doi.org/10.1111/cen.13861
Wiciński M, Ozorowski M, Wódkiewicz E, Otto SW, Kubiak K, Malinowski B (2021) Impact of Vitamin D supplementation on inflammatory markers’levels in obese patients. Curr Issues Mol Biol 43:1606–1622. https://doi.org/10.3390/cimb43030114
Palaniswamy S, Gill D, De Silva NM, Lowry E, Jokelainen J, Karhu T, Mutt SJ, Dehghan A, Sliz E, Chasman DI et al (2020) Could vitamin D reduce obesity-associated inflammation? Observational and Mendelian randomization study. Am J Clin Nutr 111:1036–1047. https://doi.org/10.1093/ajcn/nqaa056
Yi X, Sun J, Li L, Wei Q, Qian Y, Chen X, Ma L (2016) 1,25-Dihydroxyvitamin D3 Deficiency is Involved in the Pathogenesis of Diabetic Retinopathy in the Uygur Population of China. IUBMB Life 68:445–451. https://doi.org/10.1002/iub.1501
Xiang W, Cheng S, Zhou Y, Ma L (2020) Effects of 1,25(OH)2 D3 on lipid droplet growth in adipocytes. BioFactors 46:943–954. https://doi.org/10.1002/biof.1610
Cox AJD, West NPP, Cripps AWP (2015) Obesity, inflammation, and the gut microbiota. Lancet Diabetes Endocrinol 3:207–215. https://doi.org/10.1016/S2213-8587(14)70134-2
Schachter J, Martel J, Lin C, Chang C, Wu T, Lu C, Ko Y, Lai H, Ojcius DM, Young JD (2018) Effects of obesity on depression: a role for inflammation and the gut microbiota. Brain Behav Immun 69:1–8. https://doi.org/10.1016/j.bbi.2017.08.026
Tilg H, Zmora N, Adolph TE, Elinav E (2020) The intestinal microbiota fuelling metabolic inflammation. Nat Rev Immunol 20:40–54. https://doi.org/10.1038/s41577-019-0198-4
Sallam NA, Borgland SL (2021) Insulin and endocannabinoids in the mesolimbic system. J Neuroendocrinol 33:e12965. https://doi.org/10.1111/jne.12965
Liang H, Lum H, Alvarez A, Garduno-Garcia JDJ, Daniel BJ, Musi N, Fürnsinn C (2018) A low dose lipid infusion is sufficient to induce insulin resistance and a pro-inflammatory response in human subjects. Plos One 13:e195810. https://doi.org/10.1371/journal.pone.0195810
Suzuki T, Hara H (2019) Dietary fat and bile juice, but not obesity, are responsible for the increase in small intestinal permeability induced through the suppression of tight junction protein expression in LETO and OLETF rats. Nutr Metab 7:19
Guo S, Li W, Chen F, Yang S, Huang Y, Tian Y, Xu D, Cao N (2021) Polysaccharide of Atractylodes macrocephala Koidz regulates LPS-mediated mouse hepatitis through the TLR4-MyD88-NFkappaB signaling pathway. Int Immunopharmacol 98:107692. https://doi.org/10.1016/j.intimp.2021.107692
Johnson SL, Kirk RD, DaSilva NA, Ma H, Seeram NP, Bertin MJ (2019) Polyphenol microbial metabolites exhibit gut and blood(-)brain barrier permeability and protect murine microglia against LPS-induced inflammation. Metabolites. https://doi.org/10.3390/metabo9040078
de La Serre CB, Ellis CL, Lee J, Hartman AL, Rutledge JC, Raybould HE (2010) Propensity to high-fat diet-induced obesity in rats is associated with changes in the gut microbiota and gut inflammation. Am J Physiol Gastrointest Liver Physiol 299:G440–G448. https://doi.org/10.1152/ajpgi.00098.2010
Gordon JI, Ley RE, Turnbaugh PJ, Klein S (2006) Microbial ecology Human gut microbes associated with obesity. Nature (London) 444:1022–1023. https://doi.org/10.1038/4441022a
Murphy EF, Cotter PD, Healy S, Marques TM, O’Sullivan O, Fouhy F, Clarke SF, O’Toole PW, Quigley EM, Stanton C et al (2010) Composition and energy harvesting capacity of the gut microbiota: relationship to diet, obesity and time in mouse models. Gut 59:1635–1642. https://doi.org/10.1136/gut.2010.215665
Porras D, Nistal E, Martinez-Florez S, Pisonero-Vaquero S, Olcoz JL, Jover R, Gonzalez-Gallego J, Garcia-Mediavilla MV, Sanchez-Campos S (2017) Protective effect of quercetin on high-fat diet-induced non-alcoholic fatty liver disease in mice is mediated by modulating intestinal microbiota imbalance and related gut-liver axis activation. Free Radic Biol Med 102:188–202. https://doi.org/10.1016/j.freeradbiomed.2016.11.037
Zhou X, Chen C, Zhong YN, Zhao F, Hao Z, Xu Y, Lai R, Shen G, Yin X (2020) Effect and mechanism of vitamin D on the development of colorectal cancer based on intestinal flora disorder. J Gastroenterol Hepatol 35:1023–1031. https://doi.org/10.1111/jgh.14949
Li P, Wang Y, Li P, Chen X, Liu Y, Zha L, Zhang Y, Qi K (2023) Maternal vitamin D deficiency aggravates the dysbiosis of gut microbiota by affecting intestinal barrier function and inflammation in obese male offspring mice. Nutrition 105:111837. https://doi.org/10.1016/j.nut.2022.111837
Lei F, Zhang XN, Wang W, Xing DM, Xie WD, Su H, Du LJ (2007) Evidence of anti-obesity effects of the pomegranate leaf extract in high-fat diet induced obese mice. Int J Obes (Lond) 31:1023–1029. https://doi.org/10.1038/sj.ijo.0803502
Jahani R, Fielding KA, Chen J, Villa CR, Castelli LM, Ward WE, Comelli EM (2014) Low vitaminD status throughout life results in an inflammatory prone status but does not alter bone mineral or strength in healthy 3-month-old CD-1 male mice. Mol Nutr Food Res 58:1491–1501. https://doi.org/10.1002/mnfr.201300928
Kim SJ, Cho DH, Lee GY, An JH, Han SN (2022) The effects of dietary vitamin D supplementation and in vitro 1,25 dihydroxyvitamin D3 treatment on autophagy in bone marrow-derived dendritic cells from high-fat diet-induced obese mice. J Nutr Biochem 100:108880. https://doi.org/10.1016/j.jnutbio.2021.108880
Cani PD, Jordan BF (2018) Gut microbiota-mediated inflammation in obesity: a link with gastrointestinal cancer. Nat Rev Gastroenterol Hepatol 15:671–682. https://doi.org/10.1038/s41575-018-0025-6
Hersoug LG, Moller P, Loft S (2016) Gut microbiota-derived lipopolysaccharide uptake and trafficking to adipose tissue: implications for inflammation and obesity. Obes Rev 17:297–312. https://doi.org/10.1111/obr.12370
Ballini A, Scacco S, Boccellino M, Santacroce L, Arrigoni R (2020) Microbiota and Obesity: Where Are We Now? Biology 9:415. https://doi.org/10.3390/biology9120415
Willis AD (2019) Rarefaction, alpha diversity, and statistics. Front Microbiol. https://doi.org/10.1101/231878
Wilmanski T, Rappaport N, Earls JC, Magis AT, Manor O, Lovejoy J, Omenn GS, Hood L, Gibbons SM, Price ND (2019) Blood metabolome predicts gut microbiome alpha-diversity in humans. Nat Biotechnol 37:1217–1228. https://doi.org/10.1038/s41587-019-0233-9
Wang Y, Zhao L, Gao L, Pan A, Xue H (2021) Health policy and public health implications of obesity in China. Lancet Diabetes Endocrinol 9:446–461. https://doi.org/10.1016/S2213-8587(21)00118-2
Scheithauer T, Rampanelli E, Nieuwdorp M, Vallance BA, Verchere CB, van Raalte DH, Herrema H (2020) Gut microbiota as a trigger for metabolic inflammation in obesity and type 2 diabetes. Front Immunol 11:571731. https://doi.org/10.3389/fimmu.2020.571731
Hajhashemy Z, Shahdadian F, Ziaei R, Saneei P (2021) Serum vitamin D levels in relation to abdominal obesity: a systematic review and dose–response meta-analysis of epidemiologic studies. Obes Rev. https://doi.org/10.1111/obr.13134
Zhao D, Cao J, Jin H, Shan Y, Fang J, Liu F (2021) Beneficial impacts of fermented celery (Apium graveolens L.) juice on obesity prevention and gut microbiota modulation in high-fat diet fed mice. Food Funct 12:9151–9164. https://doi.org/10.1039/d1fo00560j
Bian Y, Lei J, Zhong J, Wang B, Wan Y, Li J, Liao C, He Y, Liu Z, Ito K et al (2022) Kaempferol reduces obesity, prevents intestinal inflammation, and modulates gut microbiota in high-fat diet mice. J Nutr Biochem 99:108840. https://doi.org/10.1016/j.jnutbio.2021.108840
Milton-Laskibar I, Marcos-Zambrano LJ, Gomez-Zorita S, Fernandez-Quintela A, Carrillo DSPE, Martinez JA, Portillo MP (2021) Gut microbiota induced by pterostilbene and resveratrol in high-fat-high-fructose fed rats: putative role in steatohepatitis onset. Nutrients. https://doi.org/10.3390/nu13051738
Nur Zati Iwani AK, Jalaludin MY, Yahya A, Mansor F, Md Zain F, Hong JYH, Wan Mohd Zin RM, Mokhtar AH (2022) TG: HDL-C ratio as insulin resistance marker for metabolic syndrome in children with obesity. Front Endocrinol (Lausanne) 13:852290. https://doi.org/10.3389/fendo.2022.852290
Hayek T, Ito Y, Azrolan N, VERDERY RB, AALTO-SETÄLÄ K, WALSH A, BRESLOW JL, (1993) Dietary fat increases high density lipoprotein (HDL) levels both by increasing the transport rates and decreasing the fractional catabolic rates of HDL cholesterol ester and apolipoprotein (Apo) A-I : presentation of a new animal model and mechanistic studies in human Apo A-I transgenic and control mice. J Clin Investig 91:1665–1671. https://doi.org/10.1172/JCI116375
Cordeiro MM, Biscaia PB, Brunoski J, Ribeiro RA, Franco GCN, Scomparin DX (2021) Vitamin D supplementation decreases visceral adiposity and normalizes leptinemia and circulating TNF-α levels in western diet-fed obese rats. Life Sci 278:119550. https://doi.org/10.1016/j.lfs.2021.119550
Hussein HM, Elyamany MF, Rashed LA, Sallam NA (2022) Vitamin D mitigates diabetes-associated metabolic and cognitive dysfunction by modulating gut microbiota and colonic cannabinoid receptor 1. Eur J Pharm Sci 170:106105. https://doi.org/10.1016/j.ejps.2021.106105
Mirhosseini N, Vatanparast H, Mazidi M, Kimball SM (2017) The effect of improved serum 25-hydroxyvitamin D status on glycemic control in diabetic patients: a meta-analysis. J Clin Endocrinol Metab 102:3097–3110. https://doi.org/10.1210/jc.2017-01024
Deledda A, Annunziata G, Tenore GC, Palmas V, Manzin A, Velluzzi F (2021) Diet-derived antioxidants and their role in inflammation. Obesity Gut Microb Modul Antiox 10:708. https://doi.org/10.3390/antiox10050708
Yang X, Mo W, Zheng C, Li W, Tang J, Wu X (2020) Alleviating effects of noni fruit polysaccharide on hepatic oxidative stress and inflammation in rats under a high-fat diet and its possible mechanisms. Food Funct 11:2953–2968. https://doi.org/10.1039/d0fo00178c
Rohm TV, Fuchs R, Muller RL, Keller L, Baumann Z, Bosch A, Schneider R, Labes D, Langer I, Pilz JB et al (2021) Obesity in humans is characterized by gut inflammation as shown by pro-inflammatory intestinal macrophage accumulation. Front Immunol 12:668654. https://doi.org/10.3389/fimmu.2021.668654
Rogero MM, Calder PC (2018) Obesity, inflammation, toll-like receptor 4 and fatty acids. Nutrients 10:432. https://doi.org/10.3390/nu10040432
Marziou A, Philouze C, Couturier C, Astier J, Obert P, Landrier J, Riva C (2020) Vitamin D supplementation improves adipose tissue inflammation and reduces hepatic steatosis in obese C57BL/6J mice. Nutrients 12:342. https://doi.org/10.3390/nu12020342
Chang E (2022) Effects of vitamin D supplementation on adipose tissue inflammation and NF-κB/AMPK activation in obese mice fed a high-fat diet. Int J Mol Sci 23:10915. https://doi.org/10.3390/ijms231810915
Qu L, Ma X, Fan D (2021) Ginsenoside Rk3 suppresses hepatocellular carcinoma development through targeting the gut-liver axis. J Agr Food Chem 69:10121–10137. https://doi.org/10.1021/acs.jafc.1c03279
Cani PD, Bibiloni R, Knauf C, Waget A, Neyrinck AM, Delzenne NM, Burcelin R (2008) Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes 57:1470–1481. https://doi.org/10.2337/db07-1403
Rohr MW, Narasimhulu CA, Rudeski-Rohr TA, Parthasarathy S (2020) Negative effects of a high-fat diet on intestinal permeability: a review. Adv Nutr 11:77–91. https://doi.org/10.1093/advances/nmz061
Huang K, Dong W, Liu W, Yan Y, Wan P, Peng Y, Xu Y, Zeng X, Cao Y (2019) 2-O-beta-d-Glucopyranosyl-l-ascorbic Acid, an Ascorbic Acid Derivative Isolated from the Fruits of Lycium Barbarum L., Modulates Gut Microbiota and Palliates Colitis in Dextran Sodium Sulfate-Induced Colitis in Mice. J Agric Food Chem 67:11408–11419. https://doi.org/10.1021/acs.jafc.9b04411
Chen H, Lu R, Zhang Y, Sun J (2018) Vitamin D receptor deletion leads to the destruction of tight and adherens junctions in lungs. Tissue Barriers 6:1–13. https://doi.org/10.1080/21688370.2018.1540904
Meckel K, Li YC, Lim J, Kocherginsky M, Weber C, Almoghrabi A, Chen X, Kaboff A, Sadiq F, Hanauer SB et al (2016) Serum 25-hydroxyvitamin D concentration is inversely associated with mucosal inflammation in patients with ulcerative colitis. Am J Clin Nutr 104:113–120. https://doi.org/10.3945/ajcn.115.123786
Liu Y, Meng F, Wang S, Xia S, Wang R (2021) Vitamin D 3 mitigates lipopolysaccharide-induced oxidative stress, tight junction damage and intestinal inflammatory response in yellow catfish, Pelteobagrus fulvidraco. Comparative biochemistry and physiology. Toxicol Pharmacol 243:108982
Wang Z, Yang H, Lv H, Huang C, Qian J (2022) Vitamin D receptor-dependent protective effect of moderate hypoxia in a mouse colitis model. Front Physiol 13:876890. https://doi.org/10.3389/fphys.2022.876890
Malaguarnera L (2020) Vitamin D and microbiota: Two sides of the same coin in the immunomodulatory aspects. Int Immunopharmacol 79:106112. https://doi.org/10.1016/j.intimp.2019.106112
Battistini C, Ballan R, Herkenhoff ME, Saad SMI, Sun J (2021) Vitamin D modulates intestinal microbiota in inflammatory bowel diseases. Int J Mol Sci 22:362–384. https://doi.org/10.3390/ijms22010362
Sanchez-Carrillo S, Ciordia S, Rojo D, Zubeldia-Varela E, Mendez-Garcia C, Martinez-Martinez M, Barbas C, Ruiz-Ruiz S, Moya A, Garriga M et al (2021) A body weight loss- and health-promoting gut microbiota is established after bariatric surgery in individuals with severe obesity. J Pharm Biomed Anal 193:113747. https://doi.org/10.1016/j.jpba.2020.113747
Zhang X, Shang X, Jin S, Ma Z, Wang H, Ao N, Yang J, Du J (2021) Vitamin D ameliorates high-fat-diet-induced hepatic injury via inhibiting pyroptosis and alters gut microbiota in rats. Arch Biochem Biophys 705:108894. https://doi.org/10.1016/j.abb.2021.108894
Shin N, Whon TW, Bae J (2015) Proteobacteria: microbial signature of dysbiosis in gut microbiota. Trends Biotechnol 33:496–503. https://doi.org/10.1016/j.tibtech.2015.06.011
Chua HH, Chou HC, Tung YL, Chiang BL, Liao CC, Liu HH, Ni YH (2018) Intestinal dysbiosis featuring abundance of ruminococcus gnavus associates with allergic diseases in infants. Gastroenterology 154:154–167. https://doi.org/10.1053/j.gastro.2017.09.006
Drewes JL, Chen J, Markham NO, Knippel RJ, Domingue JC, Tam AJ, Chan JL, Kim L, McMann M, Stevens C et al (2022) Human colon cancer-derived clostridioides difficile strains drive colonic tumorigenesis in mice. Cancer Discov 12:1873–1885. https://doi.org/10.1158/2159-8290.CD-21-1273
Wu Z, Liu H, Yan E, Zhang X, Wang Y, Huang C, He T, Miao L, Yang L, Jiang R et al (2023) Desulfovibrio confers resilience to the comorbidity of pain and anxiety in a mouse model of chronic inflammatory pain. Psychopharmacology 240:87–100. https://doi.org/10.1007/s00213-022-06277-4
Suo H, Shishir MRI, Xiao J, Wang M, Chen F, Cheng K (2021) Red wine high-molecular-weight polyphenolic complex: an emerging modulator of human metabolic disease risk and gut microbiota. J Agr Food Chem 69:10907–10919. https://doi.org/10.1021/acs.jafc.1c03158
Zhang N, Wang Q, Lin F, Zheng B, Huang Y, Yang Y, Xue C, Xiao M, Ye J (2022) Neoagarotetraose alleviates high fat diet induced obesity via white adipocytes browning and regulation of gut microbiota. Carbohyd Polym 296:119903. https://doi.org/10.1016/j.carbpol.2022.119903
Hiippala K, Barreto G, Burrello C, Diaz-Basabe A, Suutarinen M, Kainulainen V, Bowers JR, Lemmer D, Engelthaler DM, Eklund KK et al (2020) Novel odoribacter splanchnicus strain and its outer membrane vesicles exert immunoregulatory effects in vitro. Front Microbiol 11:575455. https://doi.org/10.3389/fmicb.2020.575455
Xie J, Li LF, Dai TY, Qi X, Wang Y, Zheng TZ, Gao XY, Zhang YJ, Ai Y, Ma L et al (2022) Short-chain fatty acids produced by ruminococcaceae mediate alpha-linolenic acid promote intestinal stem cells proliferation. Mol Nutr Food Res 66:e2100408. https://doi.org/10.1002/mnfr.202100408
Raja G, Gupta H, Gebru YA, Youn GS, Choi YR, Kim HS, Yoon SJ, Kim DJ, Kim TJ, Suk KT (2021) Recent advances of microbiome-associated metabolomics profiling in liver disease: principles, mechanisms, and applications. Int J Mol Sci. https://doi.org/10.3390/ijms22031160
Yang W, Ren D, Zhao Y, Liu L, Yang X (2021) Fuzhuan brick tea polysaccharide improved ulcerative colitis in association with gut microbiota-derived tryptophan metabolism. J Agr Food Chem 69:8448–8459. https://doi.org/10.1021/acs.jafc.1c02774
Ma H, Zhao Y, Li L, Liu Y (2021) Effects of combinations of goat milk and oligosaccharides on altering the microbiota, immune responses, and short chain fatty acid levels in the small intestines of mice. J Agr Food Chem 69:8828–8837. https://doi.org/10.1021/acs.jafc.1c03408
Wang L, Cheng R, Sun X, Zhao Y, Ge W, Yang Y, Gao Y, Ding Z, Liu J, Zhang J (2021) Preparation and gut microbiota modulatory property of the oligosaccharide riclinoctaose. J Agric Food Chem 69:3667–3676. https://doi.org/10.1021/acs.jafc.0c07783
Zhu X, Zhang Z, Yang X, Qi L, Guo Y, Tang X, Xie Y, Chen D (2022) Improvement of extraction from Hericium erinaceus on the gut-brain axis in AD-like mice. Brain Res 1793:148038. https://doi.org/10.1016/j.brainres.2022.148038
Zhang H, Xu Z, Chen W, Huang F, Chen S, Wang X, Yang C (2022) Algal oil alleviates antibiotic-induced intestinal inflammation by regulating gut microbiota and repairing intestinal barrier. Front Nutr (Lausanne) 9:1081717. https://doi.org/10.3389/fnut.2022.1081717
Liu J, Hao W, He Z, Kwek E, Zhao Y, Zhu H, Liang N, Ma KY, Lei L, He WS et al (2019) Beneficial effects of tea water extracts on the body weight and gut microbiota in C57BL/6J mice fed with a high-fat diet. Food Funct 10:2847–2860. https://doi.org/10.1039/c8fo02051e
Funding
This research was funded by the Szechwan Province Science and Technology Program in 2019, Grant Number: 2019YJ0483 and the Luzhou Key Research and Development of Science and Technology Plan in 2018, Grant Number: 2018-SYF-20.
Author information
Authors and Affiliations
Contributions
Conceptualization, LX, YZ and LM; data collection, LX and LM; formal analysis, LX, TD and JZ; funding acquisition, YZ and LM; investigation, LX, TD, JZ, YZ, YZ and YZ; methodology, LX, JZ and LM; project administration, LX, YZ and LM; resources, LM; supervision, YZ and LM; validation, YZ and YZ; conceptualization, LX and TD; writing—original draft, LX; writing—review & editing, LX, TD, YZ and LM. All authors have read and approved the published version of this manuscript.
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Informed consent statement
Not applicable.
Institutional review board statement
The study was approved by the Animal Ethics Committee of Southwest Medical University (approval number: 2020716).
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Xiang, L., Du, T., Zhang, J. et al. Vitamin D3 supplementation shapes the composition of gut microbiota and improves some obesity parameters induced by high-fat diet in mice. Eur J Nutr 63, 155–172 (2024). https://doi.org/10.1007/s00394-023-03246-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00394-023-03246-1