
Abstract
Purpose
Autophagy and apoptosis play critical roles in both development and tissue homeostasis in response to (patho)physiological stimuli, such as high-fat diet (HFD) and endurance training (ET). Therefore, we aimed to investigate how ET modulates autophagy and apoptotic-related signaling in visceral adipose tissue of long-standing HFD-fed rats.
Methods
The study was conducted over a 17-week period on Sprague–Dawley rats, which were divided into four groups (n = 8/group): standard diet sedentary (STD+SED), high-fat diet sedentary (HFD+SED), standard diet ET (STD+ET) and high-fat diet ET (HFD+ET). After 9 weeks of dietary regimens, ET groups were trained for 8 weeks on treadmill (5 days/week at 25 m/min for 60 min/day), while maintaining dietary regimens. Autophagy and apoptotic-signaling markers in epididymal white adipose tissue (eWAT) were determined using RT-qPCR, Western blot and spectrometry techniques.
Results
ET reduced body weight, visceral fat mass and HOMA-IR in standard and HF diet-fed animals. Moreover, ET reverted the HFD-induced increases in the percentage of larger adipocytes and also reduced the percentage of smaller adipocytes. The HFD decreased pre-adipocyte factor 1 (DLK1/PREF1) and increased the pro-apoptotic markers (Bax protein and caspase 3-like activity), while having no impact on autophagy markers. However, ET increased DLK1/PREF1 and Bcl-2 in both diet types, while decreasing Bax and caspases 9, 8 and 3-like activities in HFD feeding rats. Additionally, Beclin-1 and p62 protein significantly increased in ET groups of both diet types.
Conclusions
Data demonstrate that 8 weeks of ET was effective in attenuating apoptotic-related signaling in long-standing HFD-fed rats. Moreover, HFD and ET had no impact on VAT autophagy markers.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Western lifestyle, characterized by energy-rich diets and lack of exercise, has led to a global obesity epidemic [1]. In fact, in response to this positive energy balance, visceral adipose tissue (VAT) mass and adipocyte size increase [2]. Both represent the typical features of the emerged concept, adiposopathy or “sick fat” [3], which most likely result in obesity-related cellular metabolic disturbances [4, 5].
Apoptosis is a programmed cell death mediated by a cell-intrinsic machinery, which is important for eliminating damage cells [6], whereas autophagy is a process of lysosomal self-degradation that keeps the homeostatic balance between the synthesis, degradation, and recycling of cellular proteins and organelles [6, 7]. Apoptosis and autophagy share molecular regulatory mechanisms governed by Bcl-2 family proteins, thus regulating the activity of each other [8] under physiological and pathophysiological conditions. Generally, autophagy can support apoptosis in cell death induction or may assist apoptotic program without leading to cell death, thus enhancing cell survival [9,10,11].
In adipose tissue, autophagy actively participates in the regulation of adipogenesis, fat storage and inflammation [12, 13]. Several studies demonstrate that white adipocytes likely exhibit activated autophagic flux as observed in genetic and dietary models of obesity [14], in obese individuals [9, 11] and in obese mice lacking leptin (Lepob) [15]. Obesity is associated with an increased accumulation of autophagosomes and enhanced expression of key autophagy-related molecules LC3II, Beclin-1, ATG5, and ATG7 [11, 16, 17]. Markers of autophagy are correlated with whole-body adiposity, visceral fat distribution, and adipocyte hypertrophy [11, 17]. Moreover, the pathological expansion of the adipose tissue in the onset of obesity contributes to the formation of hypoxic areas, which results in the activation of death receptors (extrinsic) or mitochondrial (intrinsic) pathways leading to the stimulation of effector caspases and adipocyte apoptosis [18, 19]. Reports showed that adipocyte apoptosis is prominent in both obese animals [20, 21] and humans [20]. A complex interplay between adipocyte autophagy and apoptosis determines the degree of obesity-related adipose tissue disturbances.
However, few studies suggest that exercise-induced autophagy activation may have important implications in metabolic tissues, thus protecting against HFD-induced metabolic disease in rodents [22,23,24]. Other studies reported an increase of anti-apoptotic gene expression in mature epididymal adipocytes [25] after acute high-intensity endurance exercise and a reduced rate of apoptosis [26] after 12 weeks of exercise training in lean rats. To our best knowledge, the effects of physical exercise on autophagy and apoptotic signaling in VAT of obese models are poorly investigated. In fact, understanding how exercise modulates autophagy and apoptotic signaling in VAT may contribute to identify potential therapeutic molecular targets to improve adipose tissue function in obesity conditions. Therefore, with the present study, we aimed to analyze the effects of ET on autophagy and apoptotic signaling in VAT of long-standing HFD-fed rats.
Materials and methods
Animals and diets
Six-week-old male Sprague–Dawley rats were purchased (Charles River Laboratories) and housed in cages (2 rats per cage) with an enriched, temperature- and humidity-controlled environment and maintained on a 12:12 h light–dark cycle. Animals (initial body weight 229.9 ± 2.6 g) were divided into the following groups (n = 8/group): standard diet and sedentary (STD+SED); HFD sedentary (HFD+SED), standard diet plus endurance training (STD+ET) and HFD plus endurance training (HFD+ET). Afterwards, animals were fed a nutritionally adequate isoenergetic and isoproteic standard (35 kcal% fat, 47 kcal% carbohydrates, and 18 kcal% protein) or HFD (71 kcal% fat, 11 kcal% carbohydrate, and 18 kcal% protein)-liquid diets purchased from Dyets Inc. (catalog no. 710027 and 712031, respectively) over 17 weeks. The standard diet was given to all animals as an adaptation to the liquid feeding during the first week.
The two diets differed in the amount of corn oil (40 g), which was added and mixed into the diet to obtain high-fat diet (HFD) [27]. As previously reported by our group [28] and others [27], this isoenergetic pair-feeding HFD is a suitable model to induce an accumulation of visceral adipose tissue and obesity-related abnormalities.
Endurance training
Animals of the endurance-trained groups were gradually submitted to a continuous running protocol on a motor-driven rodent treadmill (Le8700, Panlab Harvard Apparatus), 5 days per week for 8 weeks, as previously described [29]. Briefly, exercise intensity was progressively increased from 15 min per day at 15 m min−1 up to 60 min at 25 m min−1 in the last 4 weeks of the program. The exercise-trained animals were euthanized 48 h after the last exercise training session to avoid exercise-related acute effects. The non-exercised groups (SS and HS) were placed on a non-moving treadmill to be exposed to the same potential environment and handling stress.
The study was approved by local Institutional Ethics Committee and followed the guidelines for the care and use of laboratory animals in research advised by the Federation of European Laboratory Animal Science Association (FELASA) and Portuguese Act 129/92.
Body weight, body weight/energy intake ratio, and visceral adiposity assessments
Energy intake (kilocalories) and body weight (g) were recorded daily and weekly, respectively, during the 17 weeks of the experiment. The ratio between body weight (g) and energy intake was calculated. At the end of the study, all animals were fasted overnight for about 12 h with free access to drinking water. Blood was collected from the left ventricle and plasma centrifuged at 3000g for 15 min at 4 °C, and stored at −80 °C for later biochemical analyses. All visceral adipose depots around internal organs were excised and weighted to calculate total visceral fat mass. An aliquot of epididymal white adipose tissue (eWAT) was rapidly removed and stored at −80 °C for further analyses, described below.
Plasma analysis
Using a standardized method for an automated clinical chemistry analyzer (Olympus AU54001) plasma glucose levels were quantified. Plasma insulin levels were determined using enzymatic methods with commercial kits (10-1137-01, Mercodia). The homeostasis model assessment of insulin resistance (HOMA-IR) was measured as follows, fasting plasma insulin X fasting plasma glucose/2.43 [30].
Histological analysis
The adipose tissue samples were fixed in 10% formalin, dehydrated in absolute ethanol, cleared in xylene, and then embedded in paraffin. The paraffin was cut into 5-μm sections that were stained with hematoxylin, counterstained with eosin. The images of histological sections were captured using a light microscope (Zeiss AX10 imager A.1, Oberkochen, Germany) under 40× magnification. Any objects below an area of 350 μm2 were excluded as these may be a mixture of adipocytes and stromal vascular cells, as suggested by [31]. The distribution of adipocyte areas were determined from four sections per rat and four rats per group (>1500 adipocytes counted per group).
Western blot determinations
The eWAT was homogenized in ice-cold RIPA buffer supplemented with protease inhibitors cocktail using a Polytron homogenizer for 30 s. The homogenates were centrifuged at 13,000g for 10 min at 4 °C to remove insoluble material. The infranatant was collected from the upper fat layer to avoid contamination and used for protein quantification by Bradford method [32], then proteins were boiled for 5 min in 2× Laemmli buffer containing 710 mmol/L β-mercaptoethanol. Afterwards, 50 μg of protein extracts obtained from each sample was separated by SDS-PAGE, transferred onto polyvinyldifluoride membranes and blotted with anti-Bax (2772), anti-Bcl-2 (2870) and Beclin-1 (3495) from Cell Signaling Technology, anti-p62 (56416) from Abcam, anti-DLK/PREF1 (2103823) from Sigma Aldrich, anti-LC3 (152-3) from MBL International Corporation. All primary antibodies were at 1:1000 dilution. Control for protein loading was confirmed by Ponceau S staining. The original membrane was stripped and reblotted with β-actin (1616, Santa Cruz Biotechnology) for normalization of quantitative protein. Chemiluminescent detection was performed with horseradish peroxidase-conjugated secondary antibodies [anti-rabbit (SC2317), anti-mouse (SC2317) and anti-goat (SC2020)] in ChemiDoc™ XRS+System and band densities were quantified using Image Lab™ software 262 5.2.1 (Bio-Rad Laboratories, Inc.).
RT-qPCR analysis
Total RNA of eWAT was extracted by homogenization with an Ultra-Turrax® T 25 basic (IKA® Werke GmbH, Staufen, Germany), purified using RNeasy® Lipid Tissue Mini Kit (74804, Qiagen) and treated with DNase I (RNase-free DNase, Qiagen) to remove any trace of genomic DNA. RNA concentration was determined by reading absorbance at 260 nm. One microgram of purified RNA worked as template for cDNA production using 200 units of M-MLV reverse transcriptase (Invitrogen) and random hexamers. Transcript levels for Dlk1 (NM_053744) were quantified by qPCR (7300 Real Time PCR System, Applied Biosystems). Primers and probes were designed using the software Primer Express 2.0 (Applied Biosystems) as follows, Dlk1 (accession no. NM_053744): 5′-AACCTCCCCTGGCTGTGTTAA-3′, and 5′-AGAGGTGCAAGCCCGAATATC-3′ and FAM-AGAACCATGGCAGTGTGTCTGCAAGGA-TAMRA (Taqman® probe) purchased from Genosys (Sigma). Primers or TaqMan® probes encompassing fragments of the areas from the extremes of two exons were designed to ensure the detection of the corresponding transcript avoiding genomic DNA amplification. The cDNA was amplified at the following conditions: 95 °C for 10 min, followed by 45 cycles of 15 s at 95 °C and 1 min at 59 °C, using the TaqMan® Universal PCR Master Mix (Applied Biosystems). The primer and probe concentrations were 300 and 200 nmol/L, respectively. All samples were run in triplicate and the average values were calculated and normalized to 18S rRNA (Applied Biosystems, #4310893E), which was not affected by both interventions, using the comparative C T method (2\( ^{{ - \Delta \Delta C_{\text{T}} }} \)).
Caspase-like activity determination
To measure caspase 3-, 8- and 9-like activities, aliquots of eWAT homogenate were incubated in a reaction buffer containing 25 mM Hepes (pH 7.5), 10% (w/v) sucrose; 10 mM dithiothreitol, 0.1% CHAPS and 100 μM caspase substrate Ac (N-acetyl)-LEHD-pNA (Calbiochem, UK) for 2 h at 37 °C. Caspase 3-, 8- and 9-like activities were determined by following the detection of the chromophore p-nitroanilide after cleavage from the labeled substrate Ac-LEHD-p-nitroanilide at 405 nm. The method was calibrated with known concentrations of p-nitroanilide (Calbiochem, UK). Caspase-like activity was calculated by the p-Na released for equal protein loaded.
Statistical analysis
Data are expressed as mean ± standard error of the mean (SEM). Multiple comparisons between groups were performed using two-way analysis of variance (ANOVA). The Bonferroni post hoc test was applied for post hoc comparisons between groups and differences were considered significant at p ≤ 0.05. Statistical analysis was performed using SPSS 21.0 for Windows (SPSS Inc., Chicago IL, USA).
Results
Body weight, body weight/energy intake ratio, visceral fat mass and HOMA-IR
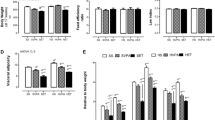
As seen in Fig. 1, the body weight/energy intake ratio was similar between studied groups. Although with no changes on body weight, HFD significantly increased visceral fat mass and HOMA-IR, a surrogate index of IR validated against the clamp technique, compared to standard diet group. ET for 8 weeks decreased body weight, visceral fat mass and HOMA-IR in both diet types compared to sedentary counterparts.

Body weight over a period of 17 weeks (a), final body weight (b), body weight/energy intake ratio (c), visceral fat mass (d) and homeostasis model assessment of insulin resistance (HOMA-IR) (e) of animals submitted to diet and endurance training regimens (n = 8/group). Data are expressed as the mean ± SEM. STD+SED standard diet sedentary, STD+ET standard diet endurance training, HFD+SED high-fat diet sedentary, HFD+ET high-fat diet endurance training. NS not significant, D diet effect, E exercise effect; *p < 0.05 vs. STD+SED; **p < 0.01 vs. STD+SED, ***p < 0.001 vs. STD+SED; # # # p < 0.001 vs. HFD+SED
Adipocyte cellularity
Although without statistical meaning, the HFD tended (p > 0.05) to decrease the percentage of adipocytes area range 1000–2999 μm2 and significantly increased adipocytes greater than 9000 μm2 compared to STD-fed group. On the other hand, the ET increased the percentage of adipocytes area range between 350 and 2999 μm2 and decreased adipocytes area greater than 6000 μm2. The protein expression of pre-adipocyte factor (DLK1/PREF1) decreased in sedentary HFD-fed animals, and its gene and protein expression increased in response to 8 weeks of ET (Fig. 2).

The frequency distribution of adipocyte size (a) and the representative images H&E staining (b). Adipocytes areas were determined from four sections per rat and four rats per group (>1500 adipocytes counted per group). The gene and protein expression of DLK1/PREF1 (c, d) of animals submitted to diet and endurance training regimens (n = 8/group). The representative images blots below. Data are expressed as the mean ± SEM. STD+SED standard diet sedentary, STD+ET standard diet endurance training, HFD+SED high-fat diet sedentary, HFD+ET high-fat diet endurance training. DxE diet and exercise interaction; *p < 0.05 vs. STD+SED; **p < 0.01 vs. STD+SED; ***p < 0.001 vs. STD+SED; ## p < 0.01 vs. HFD+SED; ### p < 0.001 vs. HFD+SED
Autophagy markers in eWAT
As observed in Fig. 3, The HFD regimen had no significant impact in Beclin-1, microtubule-associated protein-light chain 3 (LC3II) or in p62 protein expression compared to standard diet-fed animals. However, 8 weeks of ET increased Beclin-1 protein expression in both diet types (vs. both sedentary groups) and tended to increase LC3II (p = 0.06) in STD-fed animals although without reaching statistical significance. The p62 protein expression significantly increased in both ET groups (vs. both sedentary groups).

The effects of diet and endurance training on the expression of autophagy proteins (n = 8/group). Beclin-1 (a), LC3II (b), p62 (c) and representative images blots of each protein (d). Levels were normalized to β-actin for each sample. Data are expressed as the mean ± SEM. STD+SED standard diet sedentary, STD+ET standard diet endurance training, HFD+SED high-fat diet sedentary, HFD+ET high-fat diet endurance training. NS not significant, DxE diet and exercise interaction, E exercise effect; ***p < 0.001 vs. STD+SED; # p < 0.05 vs. HFD+SED; ## p < 0.01 vs. HFD+SED
Apoptosis markers in eWAT
As seen in Figs. 4 and 5, the HFD significantly increased the pro-apoptotic Bax protein expression, Bax/Bcl-2 ratio and caspase 3-like activity, when compared to standard diet-fed groups, while no alterations were observed in the anti-apoptotic Bcl-2 expression or caspases 8- and 9-like activities. On the other hand, 8 weeks of ET increased protein expression of anti-apoptotic Bcl-2 in both diet types (vs. sedentary groups) and decreased Bax expression, caspases 8 and 9-like activities (only in HFD+ET group), Bax/Bcl-2 ratio and caspase 3-like activity in both STD+ET and HFD+ET groups.

Effect of diet and endurance training on the expression of apoptosis markers (n = 8/group). Bcl-2 (a), Bax (b), Beclin-1/Bcl-2 ratio (c), Bax/Bcl-2 ratio (d) and the representative images blots of each protein (e). Levels were normalized to β-actin for each sample. Data are expressed as the mean ± SEM. STD+SED standard diet sedentary, STD+ET standard diet endurance training, HFD+SED high-fat diet sedentary, HFD+ET high-fat diet endurance training. D diet effect, E exercise effect; **p < 0.01 vs. STD+SED; ***p < 0.001 vs. STD+SED; # p < 0.05 vs. HFD+SED; ## p < 0.01 vs. HFD+SED; ### p < 0.001 vs. HFD+SED

Caspases-like activities. Activity of initiator caspases, caspase-8 and caspase-9 (a, b), and effector caspase-3 (c) in response to diet and endurance training interventions (n = 8/group). Data are expressed as the mean ± SEM. STD+SED standard diet sedentary, STD+ET standard diet endurance training, HFD+SED high-fat diet sedentary, HFD+ET high-fat diet endurance training. D diet effect, E exercise effect; **p < 0.01 vs. STD+SED; ***p < 0.001 vs. STD+SED; ## p < 0.01 vs. HFD+SED; ### p < 0.001 vs. HFD+SED
Discussion
In this study, we intended to analyze how ET modulates autophagy and apoptotic-related signaling in VAT of long-standing HFD-fed rats. The overall findings demonstrate that 8 weeks of ET reverted the apoptotic signaling induced by HFD; however, neither HFD nor ET had any impact in VAT autophagy markers.
Originally, the Lieber–DeCarli diet was established to induce liver damage [27], but the modified version of this diet was developed to induce obesity-related diseases, such as non-alcoholic steatohepatitis [28, 33]. Unlike other diets, the Lieber–DeCarli liquid diet has the advantage of having all ingredients in water, and thus, energy intake is easily monitored. As an isoenergetic pair feeding, the total energy intake and the body weight gain can be efficiently controlled, excluding any effects that may be related to these two parameters. Although with no alterations in body weight, the isoenergetic pair-feeding HFD caused a significant increase of total visceral fat mass, larger adipocytes and HOMA-IR, a surrogate indicator of IR that has been linked to adverse metabolic disturbances, as previously described elsewhere [33,34,35]. Accumulating evidence indicates that an excessive visceral adiposity accumulation, rather than overweight, has been associated with a higher risk of developing obesity-related disorders [4, 5]. In the present study, animals fed with a HFD during 9 weeks were submitted to an ET program to evaluate the therapeutic effects of ET against adverse cellular consequences imposed by a HFD. In accordance with others [35, 36], data revealed that 8 weeks of ET program was effective in attenuating obesity-related features, reducing body weight, visceral fat mass and adipocyte size.
Apoptosis (type I) and autophagy (type II) are considered the two major forms of programed cell death [7]. Nevertheless, in some conditions and till a certain degree of metabolic disorder, both can also function as pro-survival mechanisms in order to maintain cellular homeostasis and metabolism [6, 37]. These are integrated processes as several autophagy-related proteins, such as Beclin-1 [10], are also substrates for caspases during apoptosis [38]. In the present study, despite no statistical significant alterations on autophagy-related proteins were found (discussed below), an upregulation of the apoptotic effector caspase 3-like activity and Bax protein expression in VAT of sedentary obese animals argue in favor of a pro-apoptotic phenotype induced by the HFD. These data are supported by others [20, 21] and reinforce the potential contribution of apoptosis in the pathogenesis of obesity and associated diseases [39]. Some authors have been described that a pathological expansion of adipose tissue may lead to a “pro-apoptotic pressure” during HFD-induced expansion of adipose tissue, which constitutes a key mechanism to the development of inflammation and IR [20]. In the present study, the increased percentage of large adipocytes found in HFD+SED animals may be related with a pro-apoptotic phenotype.
On the other hand, regular physical exercise has been widely recognized as a relevant strategy against several cellular abnormities induced by HFD feeding, such as apoptosis [25, 40]. In this context, we observed that 8 weeks of ET induced an anti-apoptotic impact (including increased Bcl-2) on VAT and also increased DLK1/PREF1 expression in animals fed with both diet types, which are in agreement with other observations [25, 26, 41]. These findings suggest that ET was able to induce remodeling on VAT mass even in the context obesity, likely through the inhibition of adipogenesis of adipocyte precursor cells.
When autophagy is activated, the autophagosome engulfs cytoplasmic constituents and fuses with lysosomes to complete degradation. Although the number of autophagosomes correlates with the level of LC3II, other markers acting upstream (e.g., Beclin-1) or downstream (e.g., p62) are better to understand changes in autophagy flux [7]. The Beclin-1 is required for the initiation of the autophagosome formation and for the recruitment of other autophagy-related proteins [37]. On the other hand, p62 is localized on the autophagosome for degradation, being their levels strictly regulated by continuous degradation through basal autophagy [42]. Thus, a decreased p62 expression indicates an autophagic activity whereas its accumulation indicates defective autophagy [42]. Given the role of autophagy-driven adipocyte differentiation, autophagy activity increases during adipose tissue expansion [7], as observed in visceral fat of obese patients [11]. Surprisingly, our data showed that HFD feeding had no significant impact on any autophagy markers despite a decrease p62 expression (48% vs. STD+SED group). In addition, we observed an increase of hypertrophied adipocytes and decreased DLK1/PREF1 expression, an important gatekeeper of adipogenesis [43] in sedentary HFD-fed animals. These findings are in line with others [11] and suggest that the degree of autophagy activity in VAT correlated with the degree of obesity, visceral fat and adipocyte hypertrophy.
Several others studies have reported that an increased basal autophagy is essential for physical exercise-induced skeletal muscle adaptations and to potentially protect against obesity [24, 44, 45]. However, the role of physical exercise on VAT-related autophagy mechanisms is scarcely known. In the present study, 8 weeks of ET increased p62 expression in VAT of endurance-trained animals submitted to a HFD, which suggest that ET suppressed autophagic activity in eWAT although an increased Beclin-1 was also found. Beclin 1 is a central regulator of autophagy that interacts with several Beclin 1 binding proteins, such as Atg14L, Bif-1, Rubicon, among others [10]. On the other hand p62, a substrate of autophagy, is a cytosolic adaptor protein that regulates diverse cellular processes, including cell survival and cell death [10]. The p62 can facilitate cell death by binding and activating caspase-8, but can also activate the pro-survival transcription factor nuclear kappa B via activation of TRAF6 [46]. Moreover, it has been described that Beclin-1 and p62 were inversely related given that p62 accumulation is a consequence of autophagy inhibition [47]. Nevertheless, in accordance with our data, Tanaka and coworkers [48] reported that exercise training increased the formation of autophagosomes as well as the expression of LC3II, but increased the protein of expression of p62 in eWAT. Although exercise training increased the accumulation of autophagosomes in eWAT, basal autophagy was attenuated. In the same study, autophagy was increased in stromal vascular fraction collected from eWAT, which suggest that autophagic adaptations are depot-specific and may, at least in part, explain some of our conflicting results. In fact, in the present study, we used the whole adipose tissue, composed by distinct cells types, which may had represented a confounding factor to unraveling the role of ET on VAT autophagy activity in the context of obesity.
Conclusions
Data from the present study demonstrate that despite neither HFD nor ET had any significant impact on VAT autophagy markers, 8 weeks of ET was effective in attenuating apoptotic-related signaling in long-standing HFD-fed rats. Therefore, a better understanding of the molecular mechanisms underlying ET-induced adipose tissue autophagy and apoptotic adaptations are still needed to unravel the overall potential therapeutic role of physical exercise on obesity-related diseases.
References
Fruhbeck G, Yumuk V (2014) Obesity: a gateway disease with a rising prevalence. Obes Facts 7(Suppl 2):33–36. doi:10.1159/000361004
McLaughlin T, Craig C, Liu LF, Perelman D, Allister C, Spielman D, Cushman SW (2016) Adipose cell size and regional fat deposition as predictors of metabolic response to overfeeding in insulin-resistant and insulin-sensitive humans. Diabetes 65(5):1245–1254. doi:10.2337/db15-1213
Bays HE, Gonzalez-Campoy JM, Henry RR, Bergman DA, Kitabchi AE, Schorr AB, Rodbard HW (2008) Is adiposopathy (sick fat) an endocrine disease? Int J Clin Pract 62(10):1474–1483. doi:10.1111/j.1742-1241.2008.01848.x
Lopes HF, Correa-Giannella ML, Consolim-Colombo FM, Egan BM (2016) Visceral adiposity syndrome. Diabetol Metab Syndr 8:40. doi:10.1186/s13098-016-0156-2
Jacobs K, Brouha S, Bettencourt R, Barrett-Connor E, Sirlin C, Loomba R (2016) Association of nonalcoholic fatty liver disease with visceral adiposity but not coronary artery calcification in the elderly. Clin Gastroenterol Hepatol 14(9):1337 e1333–1344 e1333. doi:10.1016/j.cgh.2016.01.010
Benbrook DM, Long A (2012) Integration of autophagy, proteasomal degradation, unfolded protein response and apoptosis. Exp Oncol 34(3):286–297
Sarparanta J, Garcia-Macia M, Singh R (2016) Autophagy and mitochondria in obesity and type 2 diabetes. Curr Diabetes Rev [Epub ahead of print]
Pattingre S, Tassa A, Qu X, Garuti R, Liang XH, Mizushima N, Packer M, Schneider MD, Levine B (2005) Bcl-2 antiapoptotic proteins inhibit Beclin 1-dependent autophagy. Cell 122(6):927–939. doi:10.1016/j.cell.2005.07.002
Haim Y, Bluher M, Slutsky N, Goldstein N, Kloting N, Harman-Boehm I, Kirshtein B, Ginsberg D, Gericke M, Guiu Jurado E, Kovsan J, Tarnovscki T, Kachko L, Bashan N, Gepner Y, Shai I, Rudich A (2015) Elevated autophagy gene expression in adipose tissue of obese humans: a potential non-cell-cycle-dependent function of E2F1. Autophagy 11(11):2074–2088. doi:10.1080/15548627.2015.1094597
Kang R, Zeh HJ, Lotze MT, Tang D (2011) The Beclin 1 network regulates autophagy and apoptosis. Cell Death Differ 18(4):571–580. doi:10.1038/cdd.2010.191
Kovsan J, Bluher M, Tarnovscki T, Kloting N, Kirshtein B, Madar L, Shai I, Golan R, Harman-Boehm I, Schon MR, Greenberg AS, Elazar Z, Bashan N, Rudich A (2011) Altered autophagy in human adipose tissues in obesity. J Clin Endocrinol Metab 96(2):E268–E277. doi:10.1210/jc.2010-1681
Zhang Y, Goldman S, Baerga R, Zhao Y, Komatsu M, Jin S (2009) Adipose-specific deletion of autophagy-related gene 7 (atg7) in mice reveals a role in adipogenesis. Proc Natl Acad Sci USA 106(47):19860–19865. doi:10.1073/pnas.0906048106
Singh R, Xiang Y, Wang Y, Baikati K, Cuervo AM, Luu YK, Tang Y, Pessin JE, Schwartz GJ, Czaja MJ (2009) Autophagy regulates adipose mass and differentiation in mice. J Clin Invest 119(11):3329–3339. doi:10.1172/jci39228
Yang L, Li P, Fu S, Calay ES, Hotamisligil GS (2010) Defective hepatic autophagy in obesity promotes ER stress and causes insulin resistance. Cell Metab 11(6):467–478. doi:10.1016/j.cmet.2010.04.005
Jansen HJ, van Essen P, Koenen T, Joosten LA, Netea MG, Tack CJ, Stienstra R (2012) Autophagy activity is up-regulated in adipose tissue of obese individuals and modulates proinflammatory cytokine expression. Endocrinology 153(12):5866–5874. doi:10.1210/en.2012-1625
Kosacka J, Kern M, Kloting N, Paeschke S, Rudich A, Haim Y, Gericke M, Serke H, Stumvoll M, Bechmann I, Nowicki M, Bluher M (2015) Autophagy in adipose tissue of patients with obesity and type 2 diabetes. Mol Cell Endocrinol 409:21–32. doi:10.1016/j.mce.2015.03.015
Rodríguez A, Gomez-Ambrosi J, Catalan V, Rotellar F, Valenti V, Silva C, Mugueta C, Pulido MR, Vazquez R, Salvador J, Malagon MM, Colina I, Frühbeck G (2012) The ghrelin O-acyltransferase–ghrelin system reduces TNF-alpha-induced apoptosis and autophagy in human visceral adipocytes. Diabetologia 55(11):3038–3050. doi:10.1007/s00125-012-2671-5
Sun K, Kusminski CM, Scherer PE (2011) Adipose tissue remodeling and obesity. J Clin Invest 121(6):2094–2101. doi:10.1172/JCI45887
Yin J, Gao Z, He Q, Zhou D, Guo Z, Ye J (2009) Role of hypoxia in obesity-induced disorders of glucose and lipid metabolism in adipose tissue. Am J Physiol Endocrinol Metab 296(2):E333–E342. doi:10.1152/ajpendo.90760.2008
Alkhouri N, Gornicka A, Berk MP, Thapaliya S, Dixon LJ, Kashyap S, Schauer PR, Feldstein AE (2010) Adipocyte apoptosis, a link between obesity, insulin resistance, and hepatic steatosis. J Biol Chem 285(5):3428–3438. doi:10.1074/jbc.M109.074252
Feng D, Tang Y, Kwon H, Zong H, Hawkins M, Kitsis RN, Pessin JE (2011) High-fat diet-induced adipocyte cell death occurs through a cyclophilin D intrinsic signaling pathway independent of adipose tissue inflammation. Diabetes 60(8):2134–2143. doi:10.2337/db10-1411
He C, Bassik MC, Moresi V, Sun K, Wei Y, Zou Z, An Z, Loh J, Fisher J, Sun Q, Korsmeyer S, Packer M, May HI, Hill JA, Virgin HW, Gilpin C, Xiao G, Bassel-Duby R, Scherer PE, Levine B (2012) Exercise-induced BCL2-regulated autophagy is required for muscle glucose homeostasis. Nature 481(7382):511–515. doi:10.1038/nature10758
Cui M, Yu H, Wang J, Gao J, Li J (2013) Chronic caloric restriction and exercise improve metabolic conditions of dietary-induced obese mice in autophagy correlated manner without involving AMPK. J Diabetes Res 2013:852754. doi:10.1155/2013/852754
Greene NP, Lee DE, Brown JL, Rosa ME, Brown LA, Perry RA, Henry JN, Washington TA (2015) Mitochondrial quality control, promoted by PGC-1 alpha, is dysregulated by Western diet-induced obesity and partially restored by moderate physical activity in mice. Physiol Rep. doi:10.14814/phy2.12470
Sakurai T, Takei M, Ogasawara J, Watanabe N, Sanpei M, Yoshida M, Nakae D, Sakurai T, Nakano N, Kizaki T, Ohno H, Izawa T (2005) Exercise training enhances tumor necrosis factor-alpha-induced expressions of anti-apoptotic genes without alterations in caspase-3 activity in rat epididymal adipocytes. Jpn J Physiol 55(3):181–189. doi:10.2170/jjphysiol.R2096
Sertie RA, Andreotti S, Proenca AR, Campana AB, Lima-Salgado TM, Batista ML Jr, Seelaender MC, Curi R, Oliveira AC (1985) Lima FB (2013) Cessation of physical exercise changes metabolism and modifies the adipocyte cellularity of the periepididymal white adipose tissue in rats. J Appl Physiol 115(3):394–402. doi:10.1152/japplphysiol.01272.2012
Lieber CS, Leo MA, Mak KM, Xu Y, Cao Q, Ren C, Ponomarenko A, DeCarli LM (2004) Model of nonalcoholic steatohepatitis. Am J Clin Nutr 79(3):502–509
Goncalves IO, Passos E, Rocha-Rodrigues S, Torrella JR, Rizo D, Santos-Alves E, Portincasa P, Martins MJ, Ascensao A, Magalhaes J (2014) Physical exercise antagonizes clinical and anatomical features characterizing Lieber–DeCarli diet-induced obesity and related metabolic disorders. Clin Nutr 34(2):241–247. doi:10.1016/j.clnu.2014.03.010
Rocha-Rodrigues S, Rodriguez A, Gouveia AM, Goncalves IO, Becerril S, Ramirez B, Beleza J, Fruhbeck G, Ascensao A, Magalhaes J (2016) Effects of physical exercise on myokines expression and brown adipose-like phenotype modulation in rats fed a high-fat diet. Life Sci. doi:10.1016/j.lfs.2016.09.023
Cacho J, Sevillano J, de Castro J, Herrera E, Ramos MP (2008) Validation of simple indexes to assess insulin sensitivity during pregnancy in Wistar and Sprague–Dawley rats. Am J Physiol Endocrinol Metab 295(5):E1269–E1276. doi:10.1152/ajpendo.90207.2008
Parlee SD, Lentz SI, Mori H, MacDougald OA (2014) Quantifying size and number of adipocytes in adipose tissue. Methods Enzymol 537:93–122. doi:10.1016/b978-0-12-411619-1.00006-9
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254
Li X, Lian F, Liu C, Hu KQ, Wang XD (2015) Isocaloric pair-fed high-carbohydrate diet induced more hepatic steatosis and inflammation than high-fat diet mediated by miR-34a/SIRT1 axis in mice. Sci Rep 5:16774. doi:10.1038/srep16774
Diaz-Rua R, Garcia-Ruiz E, Caimari A, Palou A, Oliver P (2014) Sustained exposure to diets with an unbalanced macronutrient proportion alters key genes involved in energy homeostasis and obesity-related metabolic parameters in rats. Food Funct 5(12):3117–3131. doi:10.1039/c4fo00429a
Gollisch KS, Brandauer J, Jessen N, Toyoda T, Nayer A, Hirshman MF, Goodyear LJ (2009) Effects of exercise training on subcutaneous and visceral adipose tissue in normal- and high-fat diet-fed rats. Am J Physiol Endocrinol Metab 297(2):E495–E504. doi:10.1152/ajpendo.90424.2008
Linden MA, Pincu Y, Martin SA, Woods JA, Baynard T (2014) Moderate exercise training provides modest protection against adipose tissue inflammatory gene expression in response to high-fat feeding. Physiol Rep. doi:10.14814/phy2.12071
Salminen A, Kaarniranta K, Kauppinen A (2013) Beclin 1 interactome controls the crosstalk between apoptosis, autophagy and inflammasome activation: impact on the aging process. Ageing Res Rev 12(2):520–534. doi:10.1016/j.arr.2012.11.004
Thorburn A (2008) Apoptosis and autophagy: regulatory connections between two supposedly different processes. Apoptosis 13(1):1–9. doi:10.1007/s10495-007-0154-9
Boutens L, Stienstra R (2016) Adipose tissue macrophages: going off track during obesity. Diabetologia 59(5):879–894. doi:10.1007/s00125-016-3904-9
Goncalves IO, Maciel E, Passos E, Torrella JR, Rizo D, Viscor G, Rocha-Rodrigues S, Santos-Alves E, Domingues MR, Oliveira PJ, Ascensao A, Magalhaes J (2014) Exercise alters liver mitochondria phospholipidomic profile and mitochondrial activity in non-alcoholic steatohepatitis. Int J Biochem Cell Biol 54:163–173. doi:10.1016/j.biocel.2014.07.011
Sakurai T, Endo S, Hatano D, Ogasawara J, Kizaki T, Oh-ishi S, Izawa T, Ishida H, Ohno H (2010) Effects of exercise training on adipogenesis of stromal-vascular fraction cells in rat epididymal white adipose tissue. Acta Physiol (Oxf) 200(4):325–338. doi:10.1111/j.1748-1708.2010.02159.x
Sahani MH, Itakura E, Mizushima N (2014) Expression of the autophagy substrate SQSTM1/p62 is restored during prolonged starvation depending on transcriptional upregulation and autophagy-derived amino acids. Autophagy 10(3):431–441. doi:10.4161/auto.27344
Hudak CS, Sul HS (2013) Pref-1, a gatekeeper of adipogenesis. Front Endocrinol (Lausanne) 4:79. doi:10.3389/fendo.2013.00079
Lira VA, Okutsu M, Zhang M, Greene NP, Laker RC, Breen DS, Hoehn KL, Yan Z (2013) Autophagy is required for exercise training-induced skeletal muscle adaptation and improvement of physical performance. FASEB J 27(10):4184–4193. doi:10.1096/fj.13-228486
Goncalves IO, Passos E, Diogo CV, Rocha-Rodrigues S, Santos-Alves E, Oliveira PJ, Ascensao A, Magalhaes J (2016) Exercise mitigates mitochondrial permeability transition pore and quality control mechanisms alterations in nonalcoholic steatohepatitis. Appl Physiol Nutr Metab 41(3):298–306. doi:10.1139/apnm-2015-0470
Rusten TE, Stenmark H (2010) p62, an autophagy hero or culprit? Nat Cell Biol 12(3):207–209. doi:10.1038/ncb0310-207
Park JM, Huang S, Wu TT, Foster NR, Sinicrope FA (2013) Prognostic impact of Beclin 1, p62/sequestosome 1 and LC3 protein expression in colon carcinomas from patients receiving 5-fluorouracil as adjuvant chemotherapy. Cancer Biol Ther 14(2):100–107. doi:10.4161/cbt.22954
Tanaka G, Kato H, Izawa T (2015) Endurance exercise training induces fat depot-specific differences in basal autophagic activity. Biochem Biophys Res Commun 466(3):512–517. doi:10.1016/j.bbrc.2015.09.061
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Funding sources
This work was supported by a grant of Portuguese Foundation for Science and Technology (FCT) to the Research Center in Physical Activity, Health and Leisure (CIAFEL) (UID/DTP/00617/2013), to JM (PTDC/DTP-DES/7087/2014—POCI-01-0145-FEDER-016690) and to S.R-R (SFRH/BD/89807/2012).
Conflict of interest
The authors declare that there are no conflicts of interest.
Rights and permissions
About this article

Cite this article
Rocha-Rodrigues, S., Gonçalves, I.O., Beleza, J. et al. Effects of endurance training on autophagy and apoptotic signaling in visceral adipose tissue of prolonged high fat diet-fed rats. Eur J Nutr 57, 2237–2247 (2018). https://doi.org/10.1007/s00394-017-1500-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00394-017-1500-5