
Abstract
Background
As important as the vertebral ligaments are in maintaining the integrity of the spinal column and protecting the contents of the spinal canal, a single detailed review of their anatomy and function is missing in the literature.
Methods
A literature search using online search engines was conducted.
Results
Single comprehensive reviews of the spinal ligaments are not found in the extant medical literature.
Conclusions
This review will be useful to those who treat patients with pathology of the spine or who interpret imaging or investigate the anatomy of the ligaments of the vertebral column.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The sub-axial ligaments of the spinal column help stabilize the vertebral column joints. The three main joints in the vertebral column being stabilized by the sub-axial ligaments are the vertebral joints, costovertebral joints, and the sacroiliac joints. The vertebral joints consist of the intervertebral synovial and cartilaginous joints, both of which are the major components in the formation of a functional spinal unit. The costovertebral joints, only associated with the thoracic vertebrae, are a vital component to allowing adequate movement of the ribcage necessary for ventilation yet are able to provide a sturdy encasement for the protection of the thoracic organs. The sacroiliac joints attach the sacrum to the left and right ilium.
In particular, the sub-axial ligaments allow the vertebrae significant mobility within a predetermined range while still providing the structural integrity necessary to safely house the spinal cord in the vertebral canal. There are two classifications of spinal sub-axial ligaments: the intrasegmental and intersegmental systems (Fig. 1). The intrasegmental system consists of ligaments that hold individual adjacent vertebrae together and include such ligaments as the ligamentum flavum, interspinous ligament, and the intertransverse ligament. Meanwhile, the intersegmental system has the responsibility of holding together multiple vertebrae using just a single ligament, but its ligaments are much longer in length compared to those of the intrasegmental system. The intersegmental system includes ligaments such as the anterior and posterior longitudinal ligaments as well as the supraspinous ligament. Several ligaments of the sub-axial spine are very well known among the medical community because of the pathology associated with rupture, calcification, etc. of these ligaments. These ligaments include the anterior longitudinal ligament known for its involvement in the diagnosis of cervical acceleration-deceleration injury, i.e., whiplash as well as the ligamentum flavum known for its characteristic yellow color due to its composition.

Anterior view of the spinal column with associated liagments
While many of these ligaments are relatively large compared to the smaller ligaments associated with the craniocervical junction, it is important to individually study and dissect these sub-axial ligaments in order to better understand how each of the sub-axial ligaments contributes to the impressive mobility yet sturdy design of the 24 vertebrae and sacrum and coccyx. In this paper, we will review the literature associated with the sub-axial ligaments regarding its anatomy, biomechanics, pathology, radiology, and surgical aspects of each where applicable. The literature review included the use of search engines such as PubMed and appropriate textbooks looking for keywords such as ligament, sub-axial, vertebral joints, sacroiliac joint, costovertebral joint, anatomy, pathology, vertebrae, and intervertebral discs. By the end of this paper, one should have a detailed understanding of how all of these sub-axial ligaments work in concert with one another to not only assist the vertebrae with providing protection for the spinal cord and its nerve roots but to also allow for the extraordinary amount of movement seen in the vertebral column.
Functional spinal unit
The functional spinal unit, i.e., the spinal motion segment, is the smallest physiological motion unit of the spine to exhibit the biomechanical characteristics applicable to the entire spine. It consists of two adjacent vertebral bodies held together by the two facet synovial joints known as the zygapophysial joints formed by each vertebra’s articular processes, the intervertebral disc between the two vertebral bodies, and all adjoining ligaments; however, the functional spinal unit excludes any other connective tissues as muscles and tendons. These three joints allow for vertebral motion such as flexion, extension, rotation, and lateral bending at the level of the functional spinal unit.
The zygapophysial joint is a synovial joint between the inferior articular process of the superior vertebrae and the superior articular process of the inferior vertebrae in each functional spinal unit [19]. There are two zygapophysial joints in each functional spinal unit, one on either side of the midline of the vertebrae [19]. A synovial joint is formed by articulating bones coated with articular cartilage united by a joint/articular capsule spanning and enclosing the articular cavity; the capsule is composed of an outer fibrous layer lined by a serous synovial membrane [19]. The synovial joint cavity is a potential space that contains a small amount of lubricating synovial fluid, which is secreted by the synovial membrane [19]. Any non-articulating surface of a synovial joint is covered by synovial membrane [19].
The other joint involved in the functional spinal unit is the intervertebral disc between the adjacent vertebral bodies. These discs provide a strong attachment between the vertebral bodies thus uniting them into a continuous semi-rigid column [19]. These discs account for approximately 20–25 % of the height of the vertebral column [19]. The intervertebral disc is composed of an outer layer of concentric lamellae of fibrocartilage known as the annulus fibrosis while the inner portion of the intervertebral disc encased by the annulus fibrosis is the nucleus pulposus [19]. The nucleus pulposus consists mostly of water giving the nucleus pulposus a semifluid nature thus permitting the disc to have significant compressibility allowing for a wide array of movements [19].
Biomechanics of the vertebral joints
The zygapophysial joints determine the types of movements permitted and restricted between adjacent of each region (cervical, thoracic, or lumbar) [19]. Although movements between adjacent vertebrae are relatively small, it is the aggregate of all the individual movements that gives the vertebral column its extensive range of movement [19].
This diarthrodial facet joint is a crucial anatomic region of the spine due to its biomechanical role in facilitating articulation of the vertebrae of the spinal column [14]. Together with the intervertebral disc, the bilateral facet joints transfer loads and guide and constrain motions in the spine due to their geometry and mechanical function [14]. This mechanical behavior ensures the normal health and function of the spine during physiologic loading but can also lead to its dysfunction when the tissues of the facet joint are altered either by injury, degeneration, or as a result of surgical modification of the spine [14]. In general, the inclination angle of the articular surfaces of the facet joint in the sagittal plane ranges from 20° to 78° in the cervical region, 55° to 80° in the thoracic region, and 82° to 86° in the lumbar region [14]. The angle between the articulating surfaces in the axial plane range from 70° to 96°, 85° to 120°, and 15° to 70° off of the midline in the cervical, thoracic, and lumbar regions, respectively, with increasing orientation angles moving towards the lower levels in the lumbar spine [14].
Regarding the cervical zygapophysial joints, the superior facets face superoposteriorly while the inferior facets face directly inferoanteriorly [19]. The unique characteristics of the cervical vertebrae and their articulations give this region the greatest range and array of movements among all the vertebral regions [19]. Based upon a study done by Onan et al. in 1998 utilizing fresh frozen cervical vertebrae, they determined that isolated cervical facet joints allowed flexion up to 19°, extension up to 14°, lateral bending up to 28°, and rotation up to 17° [24]. However, this study does elaborate that joint movements within the constraints of the facet capsule were less than if the joints were isolated [24].
The thoracic zygapophysial joints consist of superior facets that face posteriorly and slightly laterally while the inferior facets face anteriorly and slightly medially [19]. The thoracic vertebrae have the least amount of the mobility of the three regions mainly due to the attachment of the rib cage to the vertebrae as well as the almost vertical orientation of the facet joint and the superimposing spinous processes [19]. These attachments limit the amount of flexion, extension, and lateral bending in the thoracic region; however, the most amount of rotation per joint in the vertebral column is seen in the thoracic region [19].
The lumbar zygapophysial joints involve superior facets that are directed posteromedially with inferior facets that are directed anterolaterally [19]. This arrangement of facets allows for a joint that permits flexion and extension as well as lateral flexion while inhibiting rotation [19]. In 2007, Serhan et al. showed that destabilization of a lumbar facet joint leading to an isolated lumbar zygapophysial joint that allowed for 12.2° of flexion-extension increased from 8.7° with the joint intact in its capsule [32]. Lateral bending in an isolated lumbar facet joint allows for 10.2°, increased from 9.0° with the intact joint capsule [32]. Axial rotation in an isolated lumbar zygapophysial joint is 7.2°, which decreases to 3.8° in an intact lumbar facet joint [32].
The intervertebral disc is very prominent for its function as a shock absorber due to its resilient deformability; however it is also a vertebral joint in that it permits movement between adjacent vertebrae [19]. Due to the fibrogelatinous nucleus pulposus occupying the center of the intervertebral disc surrounded by the annulus fibrosus, compressive forces on the vertebral column cause the nucleus pulposus to flatten out and the annulus fibrosus to bulge. This allows for the shock absorbance from the weight bearing force such as that exhibited by standing or lifting weights to be by the disc rather than the bony portions of the vertebral column [19]. During flexion or extension, the intervertebral disc will bulge anteriorly or posteriorly, respectively, such that the nucleus pulposus acts as a fulcrum [19]. During lateral bending, the nucleus pulposus will cause a lateral bulge towards the side of the movement.
Thus the overall function of the facet joints is to prevent the two adjacent vertebrae joined by the facet joint from engaging in relative motions that could overload and damage any surrounding spinal structures such as the intervertebral disc, the nerve roots exiting the spinal column, and the spinal cord itself [14]. The intervertebral disc absorbs approximately 75–97 % of the compressive load applied to the lumbar spine while 3–25 % is absorbed by the posterior elements of the vertebral column; this posterior force is also known as the “facet force” [14].
Anterior longitudinal ligament
Anatomy
The anterior longitudinal ligament (ALL) (Figs. 1, 2, 3, and 4) is composed of a strong band of collagenous fibers that extends along the anterior aspect of the vertebral column [3, 4, 6, 16]. The ALL is attached to the basilar occipital bone rostrally, extends to the anterior tubercle of the atlas (C1) as well as the front of the body of the axis (C2), and runs caudally to the anterior of the sacrum [3]. The ALL is narrower and thicker over the vertebral bodies than when it passes over the intervertebral (IV) discs [3, 6]. The ALL is also thicker and narrower in the thoracic region, as compared to its presence in the cervical and lumbar regions, and broadens as it travels caudally [3]. The average width of the ALL in the lumbar spine in 25 cadaveric specimens was measured to be 37.1 mm with a mean cross-sectional area of 32.4 mm2 with a range from 10.6 to 52.5 mm2 [28].

Anterolateral view of the thoracic spine illustrating the anterior longitudinal ligament, radiate ligament, and superior costotransverse ligament. The interarticular ligament is also seen at the lower rib. (Gray’s Anatomy)

Cranial part of the anterior longitudinal ligament. (Gray’s Anatomy)

Posterior aspect of thoracic vertebral ligaments (left) and anterolateral view (right)
The ALL consists of numerous collagen fibers that are densely packed and constitute three distinct layers: superficial, intermediate, and deep [3, 4, 16]. The superficial fibers are the longest, spanning anywhere from three to five vertebrae; the intermediate fibers spanning between two to three vertebrae; and the deep fibers spanning from one vertebral body to the next, connecting adjacent vertebrae [3, 16]. These longitudinal fibers attach strongly to the IV discs and the anterior margins of the vertebral bodies [3, 4, 6]. These fibers form looser, secondary attachments to the mid-portion of the vertebral bodies, filling their concave surfaces and flattening the appearance of anterior surface of the vertebral column [4]. The deep fibers of the ALL blend with the periosteum at this point, while the more superficial fibers bridge the concavity [4]. The fibers from the ALL also blend with the external surface of the annulus fibrosis, which has led anatomists to have varying opinions on the origins of these fibers [4]. While some authors believe these fibers to be part of the annulus fibrosis and others feel it constitutes a “disc capsule,” Bogduk demonstrated that the deep fibers of the ALL should not be considered part of the annulus fibrosis due to differences in the embryological origin of these two structures [4]. Accordingly, the fibers of the ALL have attachments to cortical bone, which is always the case with ligaments [4]. In 1997, Bogduk demonstrated that the annulus fibrosis does not attach to cortical bone, but rather to the vertebral end plate [4]. Bogduk also illustrated some key differences in ALL structure at varying regions of the vertebral column such as the thoracic region was the only location where the ALL was not associated with any of the prevertebral muscles and therefore stood alone [4]. Another distinction was made in the lumbar spine at the point where the crura of the diaphragm attached to the first three lumbar vertebrae [4]. In this region, the tendons of the crura extend caudally beyond the three vertebrae and form much of what is described as the ALL in this region [4]. This peculiarity has led Bogduk to suggest that the ALL may, to some extent, be a continuation of a tendon attachment in addition to its ligamentous nature [4].
Biomechanics
The ALL functions primarily to maintain the stability of the vertebral column by restricting the motion of the spinal segments during extension [4, 19]. While Neumann et al. in 1992 suggested that the ALL functions during extension, lateral bending, and axial rotation, Zander et al. in 2004 believed that the ALL is only loaded during extension and is unloaded during other movements of the spinal column [23, 43]. The ALL is the strongest and largest of the spinal ligaments and is the only one that limits extension; the remaining IV ligaments limit flexion movements [19, 23, 43]. In a study conducted by Neumann et al. in 1992, they found the stiffness of the ALL to be significantly greater than that of the posterior spinal ligaments [23]. They also noted that the ALL strength decreased slightly in subjects between the ages of 21 to 43 years [23]. In a subsequent study in 1993, Neumann et al. found the mean failure loads of bone-ALL-bone complexes to be 802 N [22]. This result is drastically different from other studies, in which the mean values ranged from 330 to 473 N [22]. Neumann and colleagues explained this difference by the properties of the specimens used in their study; they suggested that the difference is due to the fact that they used specimens from younger, and presumably less sedentary specimens, whereas other studies used ALL specimens from older subjects [22]. They also found that younger ALL specimens were more than twice as strong as older specimens [22]. However, they proposed that bone mineral content is a more accurate predictor of bone-ligament-bone structural properties than is age, as there is a linear increase in ultimate force, stiffness, and energy of this complex with increasing bone density [22]. In 1992, Pintar et al. reported the mean stiffness value for the ALL in the lumbar spine to be 33.0 N/mm with a margin of 15.7 N/mm [28]. The highest values of energy to failure were observed for the ALL and the supraspinous ligament (SSL), with values ranging from 0.82 to 8.68 J for the ALL and 3.18–11.64 J for the SSL [28]. According to Panjabi et al. in 1982, this failure is due to these ligaments needing to withstand functionally more force and deformation [25]. Failure stresses were found to be similar between the ALL and posterior longitudinal ligament (PLL) at 8.2–16.1 and 7.2–28.4 MPa, respectively [28]. The values of stress and strain at failure reported by Pintar et al. in 1992 were similar to the values reported by Chazal et al. in 1985 [5, 28].
Posterior longitudinal ligament
Anatomy
The PLL (Figs. 5 and 6) is a band of longitudinal fibers that runs within the vertebral canal along the posterior aspect of the vertebral bodies [3, 4, 6, 16]. Rostrally, the PLL is continuous with the tectorial membrane, a broad and strong band inside the vertebral canal whose superficial and deep laminae are attached to the posterior surface of the axial body with the superficial laminae expanding as it travels rostral to the upper surface of the basilar occipital bone and attaching above the foramen magnum and blending with the cranial dura mater, and attaches to the body of the axis (C2) before extending along the vertebral column to the sacrum [3]. In the cervical and upper thoracic regions, the PLL is broad and uniform in width, measuring 10–15 mm in width, but becomes almost saw-toothed in appearance as it reaches the lower thoracic and lumbar regions [3, 4, 6, 16]. This is due to the fact that the ligament narrows as it passes over the vertebral bodies and expands over the IV discs caudally [16]. In 1993, Lang described the changing course of the fibers as they reach the thoracolumbar transition; rather than running in a caudal direction and attaching to the tip of the vertebral body below, the lateral deep fibers at this level take a horizontal path and extend out to the IV foramen, covering the lower half of the annulus fibrosus and attaching at its lateral opening [16]. The average width of the PLL in the lumbar spines of 21 cadaveric specimens was 33.3 mm, with a mean cross-section area of 5.2 mm2 [28].

Cadaveric view of the posterior longitudinal ligament after removing the posterior elements of the vertebral column

Posterior view illustrating the PLL and its continuation, the tectorial membrane, onto the clivus
The fibers of the PLL are more compact than those of the ALL [6]. Between the ligamentous fibers lay the basivertebral veins and venous rami that drain them into anterior internal vertebral plexuses [3]. Early studies also describe an abundance of free and encapsulated nerve endings within the PLL [16]. Similar to the ALL, the superficial fibers of the PLL can span anywhere from three to five vertebrae and the deepest fibers between adjacent vertebrae [3, 4]. However, these layers are more pronounced in the immediate postnatal years [3]. Examination of the layers of the PLL showed that the superficial fibers measure a length of 0.5–1.0 cm [16]. The fibers of the PLL attach more firmly to the IV discs than the vertebral bodies, with deep fibers fused to the annuli fibrosi in the adult spine and the superficial fibers being more easily separable from the deeper fibers when over the vertebral bodies [3, 16]. The deep layer of the PLL will form a narrow ligament at the middle of the vertebral body, but, as it travels caudally and approaches the upper edge of the IV space, it will diverge towards the lip of the next lower vertebral body as well as a portion of the pedicle of that next lower vertebral body [16]. As per Bogduk, the deep fibers of the PLL fuse with the fibers of the annulus fibrosus but ultimately penetrate through them to attach to the posterior margins of the vertebral bodies [4]. In this way, these deep fibers act much like “paravertebral ligaments” [3].
Biomechanics
The PLL functions to resist hyperflexion of the vertebral column by resisting separation of the posterior ends of the vertebral bodies [4, 19]. However, due to the polysegmental nature of this ligament, its force is applied over several inter-body joints as opposed to just one [4]. The PLL also functions to prevent posterior herniation of the nucleus pulposus [19]. It contains nociceptive nerve endings and is the source of pain from IV disc herniation as the annulus fibrosis impinges on the ligament [19]. Pintar et al. reported the PLL has the smallest cross-sectional area with a mean of 5.2 mm2, compared to the other lumbar spinal ligaments [28]. This ligament had the lowest value of energy to failure, at a range of 0.07–0.33 J, as well as the least strain at failure, ranging between 11.3 and 16.2 % [28]. Failure stresses were found to be similar between the ALL and PLL at 8.2–16.1 and 7.2–28.4 MPa, respectively [28]. Similar values for stress and strain at failure were reported by Chazal et al. in 1985 [5, 28]. The mean stiffness value for the PLL in the lumbar spine was 20.4 ± 11.9 N mm−1 [28].
Ligamentum flavum
Anatomy
The ligamenta flava (LF) (Fig. 7) are a series of ligaments that connect the lamina of adjacent vertebrae to form alternating sections of the posterior wall of the vertebral canal [3, 4, 6, 19]. The LF are present bilaterally at each vertebral level, converging in the midline [4, 19]. This ligament’s perpendicular fibers attach to the anterior surface of the lower edge of the lamina above as well as the inferior aspect of the pedicle and descend to the posterior surface of the upper edge of the lamina below [3, 4, 6, 19, 42]. As this ligament descends inferiorly, it will divide into a medial and lateral portion with the medial portion passing to the back of the next lower lamina to attach to the rough area on the upper quarter of the dorsal surface of the lamina and the lateral portion passing in front of the zygapophysial joint formed by two vertebrae that the ligament connects [4]. Inferiorly, the lateral portion of each ligament extends to the midpoint between two pedicles and forms the anterior capsule of the zygapophysial joint as it attaches to the anterior aspects of the inferior and superior articular processes of the zygapophysial joint [3, 4, 42]. In fact, the most lateral fibers will extend along the root of the superior articular process as far as the next lower pedicle to which they are attached [4]. Where the posterior margins of the ligament meet as the ligament’s attachments extend from the zygapophysial capsule to where the laminae fuse to form spines, the ligament is only partially united with intervals being left for veins connecting the internal to posterior external posterior vertebral venous plexus [3]. In 1992, Pintar and colleagues measured the lumbar LF in 22 cadaveric specimens and found that on average the LF measures 15.2 mm in length with a mean cross-sectional area of 84.2 mm2 [28]. In the cervical region, the LF are thin and broad, thickening as they descend caudally with the thickest ligaments found in the lumbar region [3, 19]. The average thickness of the LF, both at midline and laterally, was noted to be 2–3 mm by Yong-Hing and colleagues in 1976 [42].

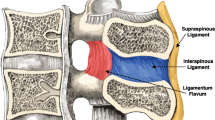
Internal and lateral view of the spinal column illustrating the ligamentum flavum, ALL, PLL, interspinous ligament, and supraspinous ligament from Gray's Anatomy
Biomechanics
The LF functions to oppose separation of the vertebral laminae during spinal flexion [3, 4, 6, 19]. This is accomplished by arresting abrupt flexion and aiding in restoring the vertebral column to an erect posture, thereby preserving the normal curvature of spine and protecting the IV discs from injury [3, 4, 6, 19, 40]. The elastic nature of this ligament plays an important role in maintaining this function as a collagenous ligament would be able to resist separation but would buckle when the laminae were approximated [4]. Buckling is minimal with an elastic ligament, protecting the contents of the spinal canal from damage [4]. The elastic nature of this ligament aids in restoring a flexed lumbar spine to its extended position while the lateral division of the ligament will prevent the anterior capsule of the zygapophysial joint from being nipped within the joint capsule during movement [4]. The LF exerts a disc pressure of 0.7 kg/cm2 and serves to pre-stress the IV disc; however, the biological significance of this is unknown [4]. The lateral portion of the ligament prevents injury to the anterior capsule of the zygapophysial joint during movement [4]. While some believe that the LF assists in extension of the spine with the help of the erector spinae muscles, Bogduk questions the importance of this ligament in producing extension movements [4, 42]. Pintar et al. reported the LF exhibited the greatest cross-sectional area compared to the other lumbar spinal ligaments, with a mean area of 84.2 ± 17.9 mm2, similar to the values reported by Panjabi et al. in 1991 [26, 28]. The LF had the lowest overall failure stresses at 1.3–4.1 MPa, with findings similar to those reported by Nachemson and Evans in 1968 [21, 28]. Similar values for stress and strain at failure were reported by Chazal et al. in 1985 [5, 28]. Panjabi et al. in 1991 reported the LF lengths ranging from 11.6 to 16.0 mm, which is again similar to the range (13.0–18.0 mm) reported by Pintar et al. in 1992 [26, 28]. The mean stiffness value for the LF was 27.2 ± 9.2 N mm−1 [28].
There have been no reported disabilities in patients with ligamentum flava excised at single or multiple sites; this is most likely due to the location rather than the function of the ligament as the ligament lies immediately behind the vertebral canal and is therefore immediately adjacent to nervous structures within the canal [4]. The lateral extensions of the ligamentum flava had elasticity of medial and anterior capsule of the posterior joints that balanced the elasticity of the intervertebral disc [42].
Supraspinous ligament
Anatomy
The supraspinous ligament (SSL) (Fig. 8a) is composed of a strong fibrous cord connecting the tips of spinous processes of the vertebral column [3, 4, 6, 19]. Fibrocartilage has been noted to be present where the ligament attaches to the spinous process [6]. There are inconsistencies in the description of the SSL among anatomists with many anatomists describing the SSL as beginning at the seventh cervical vertebra, where it merges/becomes with the nuchal ligament (Fig. 8b) to extend to the occipital protuberance, and extending in a caudal direction along the length of the vertebral column to the sacrum [3, 6, 19]. As this ligament passes caudally, it will thicken and become broader in the lumbar region where it will blend intimately with the neighboring fascia [3]. While there is no disagreement about where the ligament begins cranially, Parke, Heylings, and Bogduk all suggest that the SSL extends caudally only as far as the fifth lumbar vertebra [3, 4, 12, 27]. According to Bogduk, the SSL bridges the L4-L5 interspace in only 5 % of individuals [4]. He suggests that the ligament ends at the third lumbar spinous process in 22 % of individuals, and at L4 in the remaining 73 % [4]. Caudal to L5, the SSL is replaced by medial tendons of the erector spinae muscle [12]. The average length of the SSL in the lumbar spine is 25.2 mm with a mean cross-sectional area of 25.2 mm2 based on a study of 22 cadaveric specimens [28].

a Posterior view of the lower thoracic and lumbosacral spine noting the supraspinous ligaments. b Lateral view of the cervical spine noting the cranial extension of the supraspinous ligament, the nuchal ligament
The SSL consists of three layers: superficial, middle, and deep [4]. Similar to the ALL, the superficial fibers are composed of longitudinally oriented collagen fibers and extend over three to four spinous processes [3, 4]. This layer ranges in size from a few thin bundles of fibers to very thick bundles measuring 5–6 mm wide and 3–4 mm thick, with the majority ranging somewhere between these extremes [4]. Deep to this layer comes the middle layer, which spans two to three spinous processes, has a thickness of 1 mm, and is composed of fibers of the dorsal layer of the thoracolumbar fascia and the aponeurosis of the longissimus thoracis muscle [4]. The deepest layer of SSL connects adjacent spinous processes and consists of strong tendinous fibers of the aponeurosis of the longissimus thoracis muscle [4]. This layer in reinforced by tendon of the multifidus muscle [4]. Given the tendinous nature of the middle and deep layers of the SSL, Bogduk questions the validity of the SSL as a true ligament [4].
Biomechanics
The SSL functions to limit flexion, but provides little resistance to separation of the spinous processes [4, 12]. Heylings suggests that the absence of the SSL at the lumbosacral junction may account for the increased mobility of the spine in this region [12]. He also suggests that this absence may play a role in the etiology of low back pain and the musculoskeletal disorders that often affect the lumbosacral junction [12]. In 1999, Gudavalli and Triano used computer modeling to assess the lumbar spinal ligaments and found that the SSL carries the greatest load during flexion, followed by the LF, capsular ligament, intertransverse, and interspinous ligaments [9]. They also found that the SSL undergoes the greatest increase in length [9]. Hindle and colleagues in 1990 found that the supraspinous and interspinous ligaments carried very little load during the first half of flexion but resisted a much greater load towards the end of flexion [13]. They suggest that while these ligaments are useful in limiting passive flexion when acting in concert with other support structures, they lack much mechanical function when isolated [13]. Due to its continuous nature, the SSL was found to have one of the greatest original length, next to the ALL and PLL, compared to other spinal ligaments [28]. The highest values of energy to failure were observed for the ALL and the SSL, with values ranging from 0.82 to 8.68 J and 3.18–11.64 J, respectively [28]. Pintar et al. also reported the SSL with the highest strain at failure (70.6–115.0 %) [28]. The mean stiffness value reported for the SLL was 23.7 ± 10.9 N mm−1 [28].
Interspinous ligament
Anatomy
The interspinous ligament (ISL) (Fig. 9) is a thin and almost membranous ligament connecting adjacent vertebral spinous processes [3, 4, 6, 19]. Its attachments extend from the root to apex of each spinous process, meeting the SSL dorsally and the LF ventrally [3, 6, 19]. The ISL is poorly developed in the neck, elongated and narrow in the thoracic region, and thicker and broader in the lumbar region [3, 6]. There are inconsistencies in the description of the fibers of the ISL [12]. With regards to the cervical bundles, some anatomists describe these bundles of the ISL as part of the ligamentum nuchae while others describe them as distinct interspinous fascicles [3]. Also major discrepancies exist regarding the lumbar supraspinous and interspinous ligaments in man [12]. Many anatomists describe these fibers as crossing the interspinous space in a posterocaudal direction [3, 27, 34]. However, Heylings, Rissanen, Grant, and Bogduk describe the fibers as traversing in a posterocranial direction especially in the lumbar spine where, anteriorly the ISL is essentially a paired structure with ligaments on each side split by a mid-line cavity filled with fat [1, 4, 12, 31]. However, this cavity is not present more posteriorly [4]. Pintar et al. reported the average length in of the ISL in the lumbar spine of 18 studied specimens as 16 mm with a standard deviation of 3.2 and a mean cross-sectional area of 35.1 mm2 with a standard deviation of 15.0 [28].

Posterolateral view of the thoracic spine noting the interspinous ligament
Heylings in 1978 postulated that in order to allow the ligament to maintain a physiologically desirable degree of tensile control throughout the movement from extension to flexion but to still be stable enough to bring this movement to a halt, the ligaments needed to directed almost radially with respect to the axis of movement as seen in the collateral ligaments of the joints in the limbs [12]. The fibers that cross the interspinous space in a posterocranial direction fulfill these criteria, but if the fibers were running in the posterocaudal direction, they would either limit flexion too early if the fibers ran a straight course between their attachments, or they would be so lax in extension that the ligament would be useless in controlling in the spine except in the movement of flexion [12].
Biomechanics
The ISL functions along with the LF and SSL to provide protection and stability to the vertebral joints and store muscular energy during movement [40]. It functions weakly to limit flexion especially in the lumbar spine; the ISL also serves to resist separation of the spinous processes because the fibers run almost perpendicular to the direction of separation of the spinous processes [4, 12]. Hindle and colleagues found that the supraspinous and interspinous ligaments carried very little load during the first half of flexion but resisted a much greater load towards the end of flexion [13]. They suggest that while these ligaments are useful in limiting passive flexion when acting in concert with other support structures, they lack much mechanical function when isolated [13]. They also noted that when the SSL was removed, the ISL resisted 75 % of the load on its own [13]. The ISL is the only spinal ligament that is not loaded during lateral bending [43]. When compared to the other lumbar ligaments, this ligament was recorded to exhibit the lowest overall stiffness, at a value of 11.5 ± 6.6 N mm−1, with the joint capsule giving the highest stiffness at a value of 33.9 ± 10.7 N mm−1 as well as the lowest stress value, ranging between 1.8 and 5.9 MPa [28].
Intertransverse ligament
Anatomy
The intertransverse ligament (ITL) (Figs. 1 and 8) is a complicated ligament, interpreted in a variety of ways, that consists of sheets of connective tissue that extend from the superior border of one transverse process to the inferior border of the transverse process above [3, 4, 6, 19]. In the cervical region, this ligament consists of a few irregular and scattered fibers interspersed and within and largely replaced by the intertransverse muscles [3, 19]. The ITL is composed of rounded fibrous cords in the thoracic region intimately blended with adjacent muscles, and thin and membranous fibers in the lumbar region [3, 6, 19]. Thus, Bogduk states that the ITL is not a true ligament but rather a membranous continuation of muscular fibers with the medial and lateral continuations comprising part of a complex fascial system that separates certain paravertebral compartments [4]. In fact, Bogduk states that the only true ligament in the intertransverse area is the ligament of Bourgery; this ligament connects the base of the transverse process to the mammillary process below [4].
The ITL is not considered a true ligament, as it lacks distinct medial and lateral borders, and its collagen fibers are not as densely packed nor are they as regularly oriented as the fibers of true ligaments [4, 15]. Another reason why this ligament has been questioned as a true ligament is because its fibrous fascicles between the transverse processes of the adjacent vertebrate do not directly attach to the upper and lower transverse processes as Jiang et al. described in 1994 using a study of 32 human cadavers [15]. This study suggested that the ligament actually consisted of tendinous fascicles of the semispinalis thoracis muscle and tendons of the levatore costrarum longi muscle with the former fascicles arising from the transverse process and inserting via tendons into the spinous process and the latter fascicles arising from the ends of the transverse process and passing obliquely downward and laterally until eventually inserting into the outer surface of the rib caudal to the vertebrae from which it originated [15]. This hypothesis was corroborated by the fact that these fasicular fibers thought to be ligamentous fibers were inseparable from the muscular fibers during dissection of the 32 human cadavers [15]. These features suggest that the ITL resembles a membrane, rather than a ligament with its ligamentous appearance produced by the interweaving of muscle tendons between adjacent transverse processes [15].
In the lumbar spine, the ITL forms a septum dividing the anterior musculature from the posterior musculature in the intertransverse spaces [4]. Laterally, the ligament divides into two layers: an anterior layer of thoracolumbar fascia that covers the front of the quadratus lumborum muscle, and a posterior layer that blends with the tranverse abdominus aponeurosis to form the middle layer of the thoracolumbar fascia [4]. Medially, the ITL splits into dorsal and ventral leaves [4]. The dorsal leaf extends medially and attaches to the lateral margin of the lamina of the vertebra lying opposite to the intertransverse space while inferiorly blending with the capsule of the adjacent facet joint [4]. Meanwhile, the ventral leaf curves forward and extends over the lateral surface of vertebral bodies and blends with lateral margins of the ALL [4]. By covering the lateral aspect of the vertebral column, the ventral leaf forms a membranous sheet that closes the outer end of the interventricular foramen [4]. As such, this ventral leaf has two noted perforations that transmit structures into and out of the IV foramen [4]. Running through the superior opening are nerve branches to the psoas muscle and running through the inferior opening are the ventral rami of spinal nerves as well as spinal branches of lumbar arteries and veins [4]. Sandwiched between the dorsal and ventral leaves is a wedge-shaped space called the superior articular recess that serves to accommodate movements of the sub-adjacent facet joint [4]. This recess is filled with fat that is continuous with the intra-articular in the joint below via the foramen in the superior capsule [4].
Biomechanics
Similar to most of the other ligaments of the subaxial spine, the ITL is loaded during flexion [43]. The ITL and ALL carry the highest load during lateral bending movements of the vertebral column [43]. However, the ITL is the only ligament that is not loaded during torsion. Gudavalli and Triano assessed ligament loads using a computer model and concluded that the SSL undergoes the maximum increase in length during torsion, followed by the ISL, LF, facet capsular ligament, and finally, the ITL [9].
Facet capsular ligament
Anatomy
The facet capsular ligament (Fig. 3) fully encases the facet joint by covering the joint in a rostral to caudal direction with a non-uniform thickness [14]. The lumbar facet capsule has been reported to be approximately 2.0 mm thick in the posterior region while being as much as 3.2 mm thick in the anterior region [14]. The superior and inferior regions of the capsule have been reported to be approximately 2.4 mm thick [14]. Gray’s Anatomy identifies three distinct types of intracapsular structures: adipose tissue fat pads (anterosuperior, posteroinferior, or a combination of the two), fibroadipose meniscoids (located at the superior or inferior pole or at both poles of the capsule), and connective tissue rims (either anterior or posterior or both) [3]. While the fat pads of the facet capsules are similar to those found in many other joint capsules, the connective tissue rims of the zygapophysial joint are representative of the capsular ligament (CL) [3]. Each joint is surrounded by a thin, loose CL, which attaches to the margins of the articular processes of adjacent vertebrae [6, 19]. The average length of the CL in the lumbar spine in 24 individuals’ capsular ligaments was measured to be 16.4 mm (±2.9) and an average cross-sectional area of 43.8 mm2 (±28.3) [28].
Biomechanics
Together with other accessory ligaments of the vertebral column, the capsular ligaments help stabilize the vertebral column and its joints. In the cervical region, the loose nature of this ligament allows for a wide range of movement [19]. However, in the sacral spine, the CL functions to prevent the lumbar vertebra from sliding anteriorly down the incline of the sacrum [19]. This is because the facets on S1 face posteromedial and interlock with the anterolateral facing inferior facets of the L5 vertebra [19].
Due to the make-up of the facet capsular ligament, it can only provide mechanical resistance when the two adjacent vertebrae articulating in the zygapophysial joint undergo movement relative to one another [14]. The role of the ligament was demonstrated in studies using C3–C7 cadaveric spine segments subjected to 100 N of compression with either 2 nm of sagittal bending or 5 nm of axial torsion, before and after graded bilateral removal of the capsular ligament [14]. After removal of 50 % of the facet capsule, axial rotation increased by 19 % when torsion was applied while the vertical distance between the C4 and C6 spinous processes increased by 5 % under flexion [14]. The extent of axial torsion and posterior displacement increased by 25 and 32 %, respectively, after 75 % of the facet capsule was removed [14]. The increases in the range of motion observed after capsular transection or removal supports the hypothesis that the facet capsular ligament provides substantial contribution to constraining vertebral motion, particularly flexion and lateral bending/torsion [14].
Iliolumbar ligament
Anatomy
The iliolumbar ligament (Figs. 1 and 8) is attached to the tip of anteroinferior aspect of the L5 transverse process bilaterally as well as the L4 transverse process in some cases and radiates laterally to attach to the pelvis [3, 4]. The iliolumbar ligament will begin to radiate as it passes laterally from the transverse process and will split into two bands, upper and lower, to attach to the pelvis [3]. The lower bands (the lumbosacral portion of the iliolumbar ligament) will originate from the inferior aspect of the L5 transverse process and travel to the anterosuperior lateral surface of the sacrum and eventually blend in with the anterior sacroiliac ligament [3]. The upper bands will traverse laterally and attach to the inner lip of the iliac crest just anterior to the sacroiliac articulation with the superior portion of the upper bands becoming continuous with the lumbodorsal fascia [3, 4]. The psoas major muscle surrounds the iliolumbar ligament anteriorly; the muscles occupying the vertebral groove surround the iliolumbar ligament posteriorly and superiorly by the quadratus lumborum [3].
Early anatomists postulated that the iliolumbar ligament consisted of five parts [4]. The anterior part was a well-developed ligamentous band whose fibers span the entire anteroinferior border of the L5 transverse process (from the border of L5 vertebral body to the tip of the L5 transverse process) [4]. As the fibers from the medial end of the L5 transverse process pass laterally, they will begin to cover the fibers arising from the lateral end and the tip of the L5 transverse process such that all the fibers become continuous as it leaves the transverse process and passes posterolaterally to attach to the ilium [4]. The upper surface of this anterior bundle of the iliolumbar ligament forms the site of attachment of the ligament to the inferior portion of the quadratus lumborum muscle [4]. The superior bundle of the iliolumbar ligament is thought to be formed from the anterior and posterior thickenings of the fascia that surround the base of the quadratus lumborum muscle; these thickenings will attach to the anterosuperior border of the L5 transverse process near the lateral tip [4]. As this bundle travels laterally, it will separate to pass in front of and behind the quadratus lumborum muscle and ultimately attach to the ilium superiorly and inferiorly blend with the anterior ligament to form a channel from which the quadratus lumborum muscle arise [4]. The posterior bundle of the iliolumbar ligament arises from the tip and posterior border of the L5 transverse process and will insert into the ligamentous area of the ilium behind the origin of the quadratus lumborum; of a unique side note, the deepest fibers of the longissimus lumborum arise from the posterior bundle of the iliolumbar ligament [4]. The fourth part of the iliolumbar ligament is the inferior bundle, which arises from the lower border of the transverse process and the body of L5 vertebra [4]. These fibers will pass downwards and laterally across the surface of the anterior sacroiliac ligament to attach to the superoposterior part of the iliac fossa [4]. In order to distinguish these fibers from the anterior sacroiliac ligament, one needs to observe the oblique orientation of the inferior bundle of the iliolumbar ligament [4]. The final portion of the iliolumbar ligament is the vertical bundle, which arises from the anteroinferior border of the L5 transverse process and will descend almost vertically to attach to the posterior end of the iliopectineal line of the pelvis [4]. The significance of this bundle is that it forms the lateral margin of the channel through which the L5 ventral ramus enters the pelvis [4].
According to Bogduk, a recent study confirmed the presence of anterior and posterior parts of the iliolumbar ligament but denied the existence of the superior part while not commenting on the inferior and vertical part [4]. This study stated that the superior part is actually the anterior fascia of the quadratus lumborum muscle and as such lacks the characteristic features of a true ligament that includes the oriented collagen fibers passing directly from one bone to another [4]. The vertical and inferior bundles of the iliolumbar ligament were overlooked as part of the ventral sacroiliac ligament because these bundles attach to the lumbar portion and ilium and not the sacrum and ilium as previously thought; thus, these bundles are now thought to deserve the name “iliolumbar ligament” and not “ventral sacroiliac ligament” [4].
In 1998, Hanson et al. found that the mean length of the iliolumbar ligament in Caucasians was approximately 33.2 mm while the length in African Americans was approximately 61.8 mm [11]. These differences were also observed to be true in both sexes, i.e., African American females had iliolumbar ligaments that were longer than those of Caucasian females [11].
Thus, the numerous insertions and fiber directions of the iliolumbar ligament suggest a diversity of biomechanical functions for the iliolumbar ligament [30].
Biomechanics
The iliolumbar ligament is the most important ligament to restraining excess movement and to maintaining stability at the lumbosacral junction [17, 30]. The iliolumbar ligament is composed of two bands, anterior and posterior band, both of which serve different functions [17]. While its overall function is multifocal, the iliolumbar ligament as a whole is thought to be a ligament that prevents forward sliding of the L5 vertebrae onto the sacrum [4]. The anterior band of the iliolumbar ligament is thought to be responsible for “squaring” the L5 vertebrae on the sacrum and preventing the tilt of the vertebrae in the coronal plane as well as to prevent lateral bending while the posterior band is thought to prevent anterior slipping of the L5 vertebrae over the sacrum during weight-bearing movements as well helping to resist forward bending [4, 17, 33]. Together with the strong intervertebral disc, the iliolumbar ligament helps stabilize the lumbosacral junction and may have a significant role in maintaining the lumbosacral stability in patients with lumbosacral pathology [4, 17].
During flexion, the iliolumbar ligament will significantly limit anterior displacement of L5 vertebra [33]. When the iliolumbar was divided bilaterally, there was a 77.5 % increase noted in anterior flexion of L5 with the posterior bands accounting for the majority, approximately 61.2 %, of the increase and the anterior bands contributing only a small amount to the anterior stability of the L5 [33]. Meanwhile during extension, there was also a 20.41 % increase noted in flexion of L5 after bilateral division of the ligament; however, in this case, it was the anterior band that contributed to all of the stability during extension [33]. Lateral bending with bilateral division of the ligament did result in a dramatic increase of 141.7 % in flexion of L5 during contralateral bending with much of this increase being attributed to the anterior bands [33]. During torsion of the L5 vertebrae, division of the iliolumbar ligament did not produce much change in the flexion of L5 but both bands together did restrict torsion by 5.3 % [33]. These results by Sims and Moorman suggest that the iliolumbar ligament plays a very insignificant role in limiting the displacement of L5 onto the sacrum during torsion thus suggesting that other structures are primarily responsible for limiting this displacement during torsion [33]. Thus, Sims and Moorman theorized that the iliolumbar ligament is the probable cause of a majority of chronic low back pain [33].
Yamamoto et al. performed in vitro studies on four fresh cadavers of males aged 25–45 (total of eight ligament specimens) to evaluate the biomechanical function of the iliolumbar ligament both intact and after transection of the ligament [41]. This study involved applying a load of 10 N-m to the intact iliolumbar ligament, a right transected iliolumbar ligament, and bilateral transected iliolumbar ligaments with the movements of the vertebral column tested before and after the transaction [41]. Flexion increased by 0.9° or approximately 12 % after removal of right iliolumbar ligament and by 1.7° or approximately 23 % after bilateral iliolumbar ligament removal [41]. Extension increased by 0.6° (11 %) with right iliolumbar ligament transection and by 1.1° (20 %) with bilateral iliolumbar ligament transaction [41]. Iliolumbar ligaments significantly restricted the motion of the vertebral column to the contralateral side when under torsional load [41]. Thus, bilateral iliolumbar ligaments will significantly restrict the movements of the lumbosacral junction in the directions of flexion and extension with a slight increase in destabilization seen with unilateral transection and a statistically significant increase in destabilization seen with bilateral transection; however, when comparing all the movements of flexion, extension, right and left lateral bending, and right and left axial rotation, the iliolumbar ligament seems to be most effective in resisting lateral bending of the vertebral column [41].
In addition to stabilizing the lumbosacral junction, the iliolumbar ligament may play an important role in also stabilizing the sacroiliac joint by restraining excess movement and thus possibly helping to prevent onset of low back pain [29, 30]. In his 2003 study, Pool-Goudzwaard defined stability as the ability of a joint to bear loading forces without allowing for any uncontrolled displacements [29]. He states that the ligaments of this lumbopelvic and sacroiliac region contribute to stability by controlling the relative positions of the joint thus restricting the joint to only those positions that the joint can handle when loading forces are applied [29]. The existence of the sacroiliac part of the iliolumbar ligament, verified by MRI and cryosectioning of the pelvis in coronal and transverse plane, reinforces the thinking that the iliolumbar ligament has a direct restraining effect on movements in the sacroiliac joints [30]. In 2001, Pool-Goudzwaard et al. studied 12 human specimens and applied incremental moment to the sacroiliac joint to induce rotation in the sagittal plane [29, 30]. The results of this study indicated that the sacroiliac joint mobility in the sagittal plane increased monumentally after the total transection of both iliolumbar ligaments with most of the increase seen after cutting the ventral band of the ligament [29]. This suggests that the iliolumbar ligaments restrict sacroiliac joint mobility in the sagittal plane with the ventral band of the iliolumbar ligament contributing to most of this restriction [29]. What seems strange is that the locality of the ventral band of the iliolumbar ligament seems to draw skepticism as to how this band is able to restrict the sacroiliac joint’s mobility in the sagittal plane [29]. However, Pool-Goudzwaard offered an explanation in his 2003 study that postulated that this phenomenon could be due to the orientation of the auricular surfaces of the sacroiliac joint with respect to the fiber direction of the ventral band of the iliolumbar ligament [29]. The shape of the sacrum is a wedge with the ventral (anterior) aspect of the sacrum being larger than the dorsal (posterior) aspect of the sacrum [29]. With the auricular surfaces of sacroiliac joint being oriented dorsomedially and ventrolaterally, one would expect most of the restriction from the ligaments during sagittal rotation of the sacroiliac joint to be parallel to this orientation of the auricular surfaces [29]. Since the most ventral fibers of the ventral band of the iliolumbar ligament are arranged more parallel to the auricular surfaces of the sacroiliac joint compared to the fibers of the other components of the iliolumbar ligament, loading of the ventral band of the iliolumbar ligament is likely to be stimulated by sacroiliac rotation in the sagittal plane [29].
While several recent studies of the lumbopelvic region have demonstrated support for the hypothesis that loss of the stability of the sacroiliac joints is crucial in the etiology of non-specific low back pain, it is now believed that a variety of factors in addition to loss of ligamentous support such as decrease in muscle forces and dysfunction of the mechanoreceptors in the ligamentous system of the lumbosacral area that are important for activation of the muscles necessary for posture control all play a role in the development of non-specific low back pain [29].
Costovertebral/costotransverse joint
The costovertebral and costotransverse joints are the articulations that connect the 12 pairs (24 total) of ribs to the thoracic vertebrae. When these ribs attach to the bodies of the thoracic vertebrae, they attach at two separate places on the rib: the head and the tubercle of the rib [19]. Where the head attaches to the thoracic vertebrae, it will actually attach to two convex facets that are from adjacent vertebrae [19]. The head of the rib will articulate with the superior costal facet of the corresponding vertebrae as well as the inferior costal facet of the vertebrae superior to it [19]. The crest of the head of the rib will articulate with the corresponding intervertebral disc [19]. Due to this bi-vertebral attachment, a synovial (gliding) joint is formed that contributes to the mobility of ribs to allow for expansion and retraction during the process of breathing [19]. However, regarding the heads of ribs 1, 10, 11, and 12, they will only articulate with a single vertebra, T1, T10, T11, and T12, respectively [3].
Meanwhile, the tubercle of the rib articulates with facets on the tips of the transverse process of the vertebrae that matches its numbering, i.e., the head of the sixth rib will articulate with the superior costal facet of the T6 vertebral body and inferior costal facet of the T5 vertebral body as well as the intervertebral disc between T5 and T6 vertebral bodies and the tubercle of the sixth rib will articulate with the transverse process of the 6th rib [19]. This joint is referred to as the costotransverse joint and is also a synovial joint [19]. Just as there is an exception for the rib heads articulations, there is such an exception for the tubercles of the ribs as the 11th and 12th ribs do not articulate with the transverse processes and as a result have freer movements [3].
In order to inhibit many movements and allow for just slight gliding movements through the costovertebral and costotransverse joints, there are several strong costotransverse ligaments that constrain these joints [19]. The articular surfaces on the tubercles of the superior six ribs are convex and will fit into the concavities on the transverse processes, allowing for rotation around a transverse axis [19]. This results in elevation and depression movements of the sternal ends of the ribs and the sternum in the sagittal plane, a dynamic that is known as a pump-handle movement [19]. The gliding movements seen as a result of the articulations of the 7th–10th ribs flat tubercles with the transverse processes of T7–T10 allows for elevation and depression of the lateral-most portions of these ribs in the transverse plane, resulting in the so-called bucket-handle movement [19]. These movements in conjunction with one another during inspiration will increase the AP and transverse diameter of the thorax allowing for expansion of the cavity that will draw air into the lungs [19].
The fibrous capsules of the heads of ribs are composed of short and strong fibers that connect the costal heads to the circumference of articular surfaces formed by interventricular discs and demifacets of adjacent vertebrae [3, 6, 19]. There are 12 fibrous capsules on each side of the body (one for each rib head) equaling a total of 24 fibrous capsules. The fibrous capsule is the strongest in the anterior section, a site where the fibrous capsule will form a radiate ligament of head of rib [19]. However, the capsule is most distinct at the superior and inferior portions of the costal head articulation [6]. The upper fibers will traverse the corresponding intervertebral foramina to blend with the posterior aspects of the intervertebral discs to form a symphysis, also known as an amphiarthrosis, which is a slightly moveable joint [3]. Meanwhile, the posterior fibers are continuous with the costotransverse ligaments [3].
Radiate ligament of the costovertebral joint
Anatomy
The costovertebral radiate ligament (Figs. 1, 2, and 4) fans out from the anterior portion of the head of a rib to connect the rib to the bodies of two adjacent vertebrae as well as the intervertebral disc between the two adjacent vertebral bodies [3, 6, 19]. This ligament consists of three flat fasciculi, which are attached to the anterior part of the head of the rib just beyond the articular surface [3]. The superior fasciculus ascends to the vertebral body above while the inferior fasciculus to the vertebral body below the articulation [3]. Meanwhile, the intermediate fasciculus has the shortest and least distinct fibers of the three fasciculi with the fibers running horizontally and attaching to the corresponding intervertebral disc [19].
Some exceptions to this attachment do exist. For example regarding the first rib, the radiate ligament, which is not divided into the three fasciculi, will attach to the C7 vertebral body and the T1 vertebral body [3]. Regarding the joints of the 10th–12th rib heads, which only articulate with a single vertebral body, the ligament does not have the tri-fascicular arrangement [3]. Instead, the fibers of the ligament will attach to the vertebral body above the corresponding rib as well as the corresponding rib, i.e., the 11th rib radiate ligament will attach to the T10 vertebral body as well as the 11th rib [3].
The radiate ligaments are located along the spinal column and as such have a relationship with neurovascular structures in this area [6]. The radiate ligament is located anterior to the thoracic ganglia of the sympathetic trunk and the pleura [6]. On the right side of the body, the radiate ligaments are located anterior to the azygos vein [6]. Located posterior to the radiate ligaments at each vertebral level are the inter-articular ligaments and the synovial membranes [6].
Biomechanics
The radiate ligament connects the rib head to the vertebral bodies so closely that only slight gliding movements will occur at the facets [19]. Even with this tight connection, the slightest movement here may produce a relatively large excursion of the distal/sternal/anterior end of a rib [19].
Intra-articular ligament of head of rib
Anatomy
The intra-articular (inter-articular in older texts) ligament (Fig. 2) of the head of the rib is a short and flat ligament that attaches the crest of the head of the rib to the intervertebral disc within the joint capsule [3, 6, 19]. This ligament will attach laterally to the crest of the head of the rib separating the costal articular facets and will attach medially to the intervertebral disc; in fact, this ligament will divide the joint space into two synovial cavities [3, 6, 19]. This ligament, however, is not seen in the 1st rib head’s joint capsule as well as the 10th–12th rib head s’ joint capsule [3, 6, 19]. These ribs’ heads will articulate with their corresponding vertebral bodies without the use of an intra-articular ligament [19]. Thus in these four ribs stated above, there is only one cavity synovial cavity in the joint as well as only one synovial membrane [6].
Biomechanics
While the main function of the intra-articular ligament of the head of the rib is to divide the joint capsule into two synovial cavities, the intra-articular ligament will act as the axis around which rotation will occur [19]. While the costotransverse ligaments will limit movements of the ribs to slight gliding, the articular surfaces on the tubercles of the superior six ribs are convex and as such will fit into the concavities on the transverse processes of the vertebrae [19]. As a result, rotation will occur around a transverse axis that traverses the intra-articular ligament and the head and neck of the rib that results in elevation and depression movements of the sternal ends of the ribs and the sternum in the sagittal plane; this movement is the pump-handle movement responsible for assisting in increasing the anterior-posterior diameter of the chest during inspiration [19].
Superior costotransverse ligament
Anatomy
The superior costotransverse ligament (Figs. 2 and 4) is a broad ligament, with a varied texture ranging from a woven/membrane-like texture to a tendon-like texture, that joins the crest of the neck of the rib to the transverse process superior to the rib [15, 19]. With its fibers oriented superolaterally to inferomedially except for a small bundle that runs inferolaterally, the superior costotransverse ligament connects the transverse process superior to the rib to the costal neck, with the rib already being connected to its corresponding transverse process by two separate joints, the lateral costotransverse ligament, the costotransverse ligament, and the radiate ligament [15] . The superior costotransverse ligament, classified as a true ligament, is sometimes divided by anatomists into a strong “anterior costotransverse ligament” and a weak “posterior costotransverse ligament” between which the levator costae and external intercostal muscles lie [6, 15, 19]. The anterior portion will attach between the crest of the costal neck and the lower aspect of the transverse process above the rib with its lateral end blending in with the internal intercostals’ membrane as it is crossed by the intercostal vessels and nerve and its medial border forming a free edge [3, 15]. While Jiang did not mention the existence of a posterior layer, Gray’s Anatomy states that the posterior layer will attach on the posterior surface of costal neck and ascend postero-medially to the transverse process above with its lateral end blending in the external intercostal muscles [3, 15]. The aperture bounded by this ligament and the vertebrae permits the passage of the spinal nerve as well as the posterior branch of the intercostal artery [19]. Posterior to this ligament passes the dorsal ramus of the thoracic nerve while the ventral ramus of the intercostal nerve passes anterior to the superior costotransverse ligament [6].
However, on the first rib, no superior costotransverse ligament exists [3]. And on the shaft of the 12th rib near its head arises the lumbocostal ligament, which will attach to the base of the 1st lumbar transverse process; this lumbocostal ligament is located in series with the superior costotransverse ligaments [3].
The superior costotransverse ligament is the only spinal ligament that will connect adjacent vertebrae on the lateral aspect of the spine; the lateral costotransverse ligament will only connect the transverse process to its own rib [15]. Jiang found that the average length of the superior costotransverse ligament did not differ with sex but did increase significantly as one moves down the vertebral levels with an average length of 8 ± 2 mm at T7 vertebral level and an average length of 13 ± 3 mm at T10 vertebral level [15]. The average cross-sectional area between the levels was found to be 5 ± 2 mm2 at T7 level and 8 ± 5 mm2 at T10 level [15].
Biomechanics
The superior costotransverse ligament along with the costotransverse and lateral costotransverse ligament bind the costotransverse joint that limits these joints’ movements to slight gliding [19]. However, the superior costotransverse ligament was found by Jiang et al. to have the characteristics appropriate for involvement in the active lateral balancing of the spine [15]. The fact that the superior costotransverse ligament is the only ligament that connects adjacent vertebrae on the lateral aspect of the spine further reaffirmed its importance in maintaining lateral stability between adjacent vertebrae [15]. In their 1994 study, Jiang et al. offered evidence that the role of the superior costotransverse ligament in maintaining the lateral balance of the spine is only seen in species with a bipedal stance [15]. They tested the spinal ligaments of quadrupeds such as dogs, rats, mice, and hamsters, and found that these species lack superior costotransverse ligaments [15]. They concluded that the spines of quadrupeds do not require the balance laterally that bipedal organisms so require; as such, there may be no need for the superior costotransverse ligament in quadrupeds [15]. Because of the lack of supplementary structure of the lateral aspect of the spine, the costotransverse ligament is susceptible to damage from excessive strain [15].
Costotransverse ligament
Anatomy
The costotransverse ligament is a ligament composed of many short fibers that stretch from the rough posterior aspect of the neck of the rib to the anterior aspect of the adjacent transverse process thus filling the costotransverse foramen [3, 6, 19]. However, as seen with other ribs, there are exceptions that exist with the costotransverse ligament. In the 11th and 12th ribs, the costotransverse ligament is either rudimentary or completely absent [3].
Biomechanics
The costotransverse ligament will strengthen the anterior aspect of the costotransvserse joint [19].
Lateral costotransverse ligament
Anatomy
The lateral costotransverse ligament (Figs. 4 and 8) is a short but strong, thick ligament that passes obliquely from the dorsal apex of the transverse process to the rough non-articular part of the adjacent costal tubercle [3, 6, 19]. Each of these true ligaments, as it travels superiorly and laterally from the transverse process to the rib tubercle, forms an acute angle with the vertical line of the spine [15]. The lateral costotransverse ligament of the upper ribs, which are shorter and more oblique compared to the lateral costotransverse ligament of the lower ribs, will ascend from the transverse processes while the lateral costotransverse ligament of the lower ribs will descend from the transverse processes [3].
Biomechanics
The lateral costotransverse ligament will strengthen the posterior aspect of the costotransverse joint while the costotransverse ligament will strengthen the anterior aspect of the costotransverse joint as previously stated [19]. Because of the connection between the ribs and the transverse processes, these ligaments formed a rigid bond compared to the bond formed by the connection between the upper transverse processes and the lower ribs by the superior costotranverse ligament [15]. Due to this almost rigid connection, the lateral costotranverse ligament does not appear to allow for very much flexibility [15]. However, this ligament did not seem to have the characteristics necessary for significant involvement in the active lateral balancing of the spine [15]. As such, this ligament is unlikely to be strained during lateral bending of the spine [15].
Accessory ligament
Anatomy
The accessory ligament, if present, is usually medial to the superior costotransverse ligament [3]. While it is variable in its attachments, the bands of the ligament will usually pass from a depression located on the rib medial to a costal tubercle and span to the inferior articular process immediately above this depression [3]. However, some fibers of the ligament will also pass to the base of the transverse process [3]. In order to differentiate this ligament from the superior costotransverse ligament, one must look for the dorsal ramus of a thoracic spinal nerve and its accompanying vessels, as these structures will clearly demarcate the separation of these two ligaments from one another [3].
Biomechanics
The major function of the accessory ligament is to strengthen the costovertebral joint [3].
Sacroiliac joint
Anatomy
The sacroiliac joint is an extremely strong joint in the pelvis between the sacrum and the ilium of the pelvis joined by strong ligaments such as anterior, interosseous, and posterior sacroiliac ligaments [19]. The sacroiliac joints link and support the axial skeleton, i.e., the vertebral column with the inferior appendicular skeleton, i.e., the skeleton of the lower limb [19]. These weight-bearing joints, located on the left and the right side of the body, consist of anterior synovial joints and posterior syndesmotic joints [19]. The anterior synovial joint is formed between the sacrum’s ear-shaped auricular surfaces, which are irregular but have congruent elevations and depressions that interlock, and the ilium and is covered with articular cartilage [19]. The sacroiliac joints differ from most synovial joints in that very limited mobility is allowed, which is a consequence of their role in transmitting the weight of most of the body to the hipbones [19]. Meanwhile, the posterior syndesmosis joint is formed between the tuberosities of the same bones [19]. The sacroiliac joints are covered by two different kinds of cartilage: hyaline cartilage covering the sacral surface and fibrocartilage covering the ilial surface [38].
The sacroiliac joints’ surfaces start out flat or planar in early life; however as we begin walking, the sacroiliac joints’ surfaces begin to form distinct angular orientations while losing their planar/flat landscape [2]. These joints will also develop an elevated ridge along the ilial surface and a depression along the sacral surface, which along with the very strong ligaments increase the sacroiliac joints’ stability and makes dislocations very rare [2].
Biomechanics
The sacroiliac joints’ function is mainly that of shock absorption for the spine as well as weight transfer from the trunk and upper extremities to the lower extremities. However, the amount of this shock absorption depends on the amount of available motion at the sacroiliac joint [7]. Motion at the sacroiliac joint is limited by the interlocking of the articulating bones and the sacroiliac ligaments to slight gliding and rotary movements [19]. During weight-bearing exercises, tremendous force is transmitted through the lumbar vertebrae bodies to the superior end of the sacrum, which is pushed inferiorly and anteriorly because the weight transfer occurs anterior to the axis of the sacroiliac joint [19]. Despite the rotation of the superior sacrum, the sacrum’s movement is counterbalanced by strong ligaments that anchor the inferior end of the sacrum to the ischium thus preventing any superior and posterior rotation of the inferior sacrum [19]. By preventing too much upward movement of the inferior sacrum relative to the hip bones, resilience is provided to the sacroiliac region when the vertebral columns sustains sudden increases in force or weight [19].
In addition to shock absorption, the SI joint will also perform torque conversion to allow for transverse rotations that take place in the lower extremities to be transmitted up the spine. This provides a “self-locking” mechanism that helps with stability during the push-off phase of walking as the joint will lock on one side as weight is transferred from one leg to the other through the sacrum to the other hip bone [7]. Motion of the sacroiliac joint is limited to minute amounts of rotation and translation about the three major axes with normal ranges for rotation equal to approximately 2° in males and 2–4° in females (allowing for increased mobility during parturition) [8]. Movements of the sacroiliac joint were identified using motion-palpation, passive motion, and provocation tests by performing an analysis of 25 patients with sacroiliac disorders [35]. Sturesson et al. determined the mean maximal mobility between end points in flexion and extension on each sacroiliac joint was 2.5° ± 0.5° (1.6–3.9°), and a mean translation of 0.7 mm (0.1–1.6 mm) along the axes of rotation [35].
There are five major motions of the sacroiliac joint. Two of these movements are the anterior and posterior innominate tilt of both hipbones on the sacrum in which both the left and right coxal bones move together as a unit in the anterior or posterior direction, respectively [7]. Another movement is an anterior innominate tilt of one hipbone while the opposite hipbone tilts posteriorly on the sacrum; this movement is seen during gait [7]. During sacral flexion, the motion of the sacrum will occur simultaneously with the motion of the ilium while sacral extension involves opposite movements of the sacrum and the ilium [7].
Anterior sacroiliac ligament
Anatomy
The anterior sacroiliac ligament (Fig. 10) is the anterior part of the fibrous capsule of the synovial part of the joint [19]. This ligament is considered to be just an antero-inferior capsular thickening covering the ventral aspect of the joint [1, 3, 4]. While the ligament is mostly thin in nature, it is particularly well developed near the arcuate line and posterior inferior iliac spine where it will connect the 3rd sacral segment to the lateral side of the preauricular sulcus [3]. The ligament consists of long and transversely oriented fibers that extend from the ala and the anterior surface of the sacrum all the way to the anterior surface of the ilium [4]. In fact, these ligamentous fibers are attached to these bony landmarks for considerable distances beyond the margins of the joint [4].

Anterior view of the lower lumbar and sacrococcygeal spine and associated ligaments from Gray's Anatomy
Biomechanics
While the anterior sacroiliac ligament binds the ilium to the sacrum, it is able to prevent diastasis of the sacroiliac joint due to its anterior location [4]. However during pregnancy, relaxation of the sacroiliac ligaments due to the effects of the hormone relaxin causes the interlocking mechanism of the sacroiliac joint to become less rigid thus allowing greater rotation of the pelvis during parturition; in addition, this relaxation will also help contribute to the lordotic posture assumed during pregnancy with the change in the center of gravity during childbirth [19]. But this relaxation of the ligaments of the sacroiliac joint is not just limited to the pelvis because of the increased possibility of other joint dislocations late during pregnancy [19].
Interosseous sacroiliac ligament
Anatomy
While the interosseous sacroiliac ligament is not officially recognized as a true ligament, it may in fact be the most important ligament of the sacroiliac joint [3, 4]. This is the largest syndesmotic joint, a fibrous articulation in which bony surfaces are bound together by an interosseous ligament, a slender fibrous cord, or an aponeurotic membrane that allows for slight movement between the attachments, in the body and lies deep in the narrow recess between the sacrum and ilium posterior to the cavity of the joint [3, 4]. This “ligament,” which consists of a dense and thick collection of short collagen fibers, is the major bond between the ligamentous surfaces of the sacrum and ilium filling the irregular space postero-superior to the sacroiliac joint [3, 4]. While the posterior sacroiliac ligament covers this ligament superficially, its deeper part has superior and inferior bands that pass from depressions posterior to the sacral auricular surface to depressions on the iliac tuberosity [3]. The superior band of the interosseous sacroiliac ligament will connect the superior articular process and the lateral crest of the first two sacral segments to the neighboring ilium [3, 4]. This superior band is thusly termed the “short posterior sacroiliac ligament” as it acts in concert with the interosseous connection between the ilium and the sacrum to prevent posterior diastasis of the joint [3, 4]. In order to fully appreciate the true thickness of this ligament, one must look at transverse sections of the sacrum and ilium where this ligament is most clearly evident grossly [4].
Biomechanics
The primary function of the interosseous sacroiliac ligament is to strongly bind the ilium to the sacrum to form a secure bony interlocking mechanism that inhibits excessive movement between the sacrum and the ilium [4]. Thus, this interosseous sacroiliac ligament tightly binds the primary structures involved in transferring the weight of the upper body from the axial skeleton (vertebral column and sacrum) to the two ilia of the appendicular skeleton [19].
Posterior sacroiliac ligament
Anatomy
The posterior sacroiliac ligament (Fig. 11) is the chief ligament involved in the union of the sacrum and the ilium [3]. While overlying the interosseous ligament, the posterior sacroiliac ligament has numerous fasciculi that run in multiple directions [3, 4]. It will connect the intermediate and the lateral sacral crests to the posterior superior iliac spine and the posterior end of the internal lip of the iliac crest [3, 4]. As a posterior external continuation of the same mass of fibrous tissue, fibers of the posterior sacroiliac ligament and the interosseous ligament run obliquely superolaterally from the sacrum [19].

Posterior view of the sacrococcygeal spine and associated ligaments from Gray's Anatomy
The posterior sacroiliac ligament consists of an upper part or a short posterior sacroiliac ligament and a lower part or a long posterior sacroiliac ligament (LPSL) [3]. The upper part consists of nearly horizontal fibers that pass from the first and second transverse tubercles on the back of the sacrum to the tuberosity of the ilium [3]. Meanwhile, the lower part’s fibers are oblique in direction and run from the transverse tubercle between the 3rd and 4th sacral segments on the back of the sacrum to the posterior superior spine of the ilium and the posterior end of the internal lip of the iliac crest [3, 4, 18]. The lower part or long posterior sacroiliac ligament is continuous laterally with part of the sacrotuberous ligament and medially with the thoracolumbar fascia’s posterior lamina [3].
In 2009, McGrath et al. did histological experiments on the long posterior sacroiliac ligament that showed this part of the sacroiliac ligament was actually divided into three morphologically distinct regions [18]. Two of these regions were attached to osseous structures: the posterior superior iliac spine superiorly and the third sacral transverse tubercle inferiorly [18]. Between the proximal and distal regions of osseous attachment, the middle portion of the long posterior sacroiliac ligament, i.e., the third region of the long posterior sacroiliac ligament was observed to be a confluence of three layers: the erectors spinae aponeurosis, the “deep fasical layer,” and the gluteal aponeurosis [18]. The lateral branches of the dorsal sacral rami (medial cluneal nerves) were found to be within a layer of adipose and loose connective tissue that lies deep to the “deep fascial layer” with the middle posterior sacroiliac ligament providing a pathway for these rami between the posterior sacral region and the gluteal region [18]. With the discovery of these morphological findings, a possible pathoanatomical basis for localized pain in the posterior sacroiliac region due to an entrapment neuropathy of the lateral branches of the middle cluneal nerves was discovered [18].
Running among the posterior sacroiliac ligaments are the dorsal rami of the sacral spinal nerves, which innervate the ligament, and vessels with the lateral branches of the first three sacral dorsal rami sharing an intimate relationship with the long posterior sacroiliac ligament and the sacrotuberous ligament [3, 18, 39]. The lateral division of the S1 nerve will pass deep into the long posterior sacroiliac ligament and run between this ligament and the sacroiliac joint [39]. Meanwhile, the lateral branches of S2 and S3 unite and pass through a tunnel created by the sacrotuberous ligament and an outer sheath of thoracolumbar fascia [39].
Biomechanics
The major function of the posterior sacroiliac ligament is to unite the sacrum and the ilium [19]. Due to the fibers of the interosseous and posterior sacroiliac ligaments being obliquely oriented superolaterally from the sacrum, the axial weight pushing down on the sacrum will actually pull the ilium medially so that the ligaments compress the sacrum between them [19]. This will lock the irregular but congruent surfaces of the sacroiliac joints together along with some help from the iliolumbar ligaments [19]. In fact, it is the long posterior sacroiliac ligament that prevents backward rocking (counter-nutation) of the sacrum with respect to the ilium [4].
Sacrotuberous ligament
Anatomy
The sacrotuberous ligament (Fig. 11) attaches the sacrum, ilium, and the coccyx to the ischial tuberosity [1]. The dorsal part of this ligament will originate cranially at the sacrum and insert caudally at the ischial tuberosity while the ventral part of this ligament will originate caudally at the sacrum and insert cranially at the ischial tuberosity; as a result, the sacrotuberous ligament is considered to be composed of two triangles that connect at their tips [10]. The sacrotuberous ligament is formed from the inferior portions of the posterior sacroiliac ligaments as well as fibers extending from the posterior margin of the ilium (between the posterior superior and posterior inferior iliac spines) and the base of the coccyx [19]. The sacrotuberous ligament arises from the transverse tubercles of the lower sacral segments, from the lateral margin of the sacrum where it will blend with the sacrospinous ligament, and broadly from the posterior superior iliac spine (PSIS) where it will blend with the long posterior sacroiliac ligament [3, 4]. From this very broad origin, the ligament’s oblique fibers will descend laterally to form a thick narrow band which will widen again as it traverses laterally to attach to the medial margin of the ischial ramus/tuberosity [3, 4]. As this ligament passes the posterior ilium and the lateral sacrum and coccyx on its way to its attachment on the ischial tuberosity, the sciatic notch of the hipbone is transformed into a large sciatic foramen [19]. Where the ligament attaches to the ischial tuberosity is called the “falciform process” whose concave edge blends with the fascial sheath of the internal vessels and pudendal nerve [3].
In 2009, Hammer et al. determined that the sacrotuberous ligament in men had a mean length of 64 mm and 70 mm in women [10].
There are various muscle fibers that are continuous with the sacrotuberous ligament. Superficial fibers of this ligament are continuous with the tendon of the biceps femoris while fibers of the posterior surface of this ligament are attached to the lowest fibers of the gluteus maximus [3, 4, 36]. It was also found that a thin fascia covering the dorsal aspect of the piriformis muscle was continuous with the sacrotuberous ligament [36]. The attachment of these muscles (gluteus maximus, biceps femoris long head) to the sacrotuberous ligaments can dynamically influence movement and stability of sacroiliac joints [37].
The coccygeal branches of the inferior gluteal artery, the perforating cutaneous nerve, and filaments of the coccygeal plexus all perforate the sacrotuberous ligament [3]. In addition to sharing an intimate relationship with the long posterior sacroiliac ligament, the lateral branches of the first three sacral dorsal rami are involved with the sacrotuberous ligament [39]. As mentioned earlier in the paper, the lateral division of S1 passes deep into long posterior sacroiliac ligament, coursing between it and the sacroiliac joint; however, pressure on or swelling of the LPSL is not only capable of compressing the S1 nerve but also the S2 and S3 since these nerves also pass in close proximity to this ligament [39]. In fact, such a pressure could even compromise the microvasculature that accompanies these nerves thus potentially creating ischemic zones, which could represent the origin of local tenderness when this LPSL ligament is palpated during physical examination [39]. The lateral branches of the S2 and S3 will unite and pass through a tunnel created by the sacrotuberous ligament internally and an outer sheath of thoracolumbar fascia externally; compression of this tunnel by congestion or edema could also potentially result in low back or buttocks pain [39].
Biomechanics
Due to the orientation of the sacrotuberous ligament fibers, upward tilting of the lower end of sacrum, i.e., nutation is prevented due to its anchoring to the ischium along with the sacrospinous ligament [4, 37]. The sacrotuberous ligament rotates from its origin at the sacrum towards its insertion at the ischial tuberosity as cited by multiple authors such as Fick in 1904, Waldeyer and Fangha¨nel in 2003, and Kapandji and Koebke in 2006 [10]. An interesting fact about the sacrospinous and sacrotuberous ligaments is that both are remnants of tail muscles seen in primates [10]. Since the ligaments unroll like a contorted rope under tension, the sacrotuberous ligament will raise the ischium in relation to the sacrum while the sacrospinous will lower the ischium in relation to the sacrum; since these ligaments do not rotate in the same direction, it is assumed that the ligaments exert oppositional effects on the pelvis [10]. Even though these two ligaments have opposite effects, they are still considered to be a couple as both are very closely correlated with regards to the volumes and cross-sectional areas of both ligaments [10].
The sacrotuberous ligament can be tensed using the infamous straight leg raise test that causes shortening of the muscles [36]. The straight leg raise test alters the position of the sacrum because the sacrotuberous ligament bridges the sacroiliac joint [36]. Lavignolle et al. demonstrated that the straight leg raise test causes distinct movements in the sacroiliac joint when the patient lies supine and the hip is flexed [36]. As a result, pain elicited by the straight leg raise test could be caused by a multitude of factors including a sacroiliac joint strain [36].
Sacrospinous ligament
Anatomy
The sacrospinous ligament (Figs. 10 and 11) is a thin, triangular ligament that extends broadly from the lateral margins of the sacrum and coccyx below the sacroiliac joint to ischial spine and, together with the sacrotuberous ligament, divides the greater sciatic notch into the greater sciatic foramen and the lesser sciatic notch into the lesser sciatic foramen [1, 3, 4, 19]. This ligament will attach to the coccyx anterior to the sacrotuberous ligament with which it will blend [3]. The lateral portion of the upper border of the sacrospinous ligament will form the lower border of the greater sciatic foramen while the lateral portion of the lower border of this ligament will form the upper border of the lesser sciatic foramen [3]. While the posterior half of the sacrospinous ligament is covered by the sacrotuberous ligament, the anterior half of the sacrospinous ligament is muscular and constitutes the coccygeus muscle; in fact, many anatomists regard the sacrospinous ligament as the degenerate part of the coccygeus muscle [3].
A 2009 study by Hammer et al. found that the sacrospinous ligament, interspersed with coccygeus skeletal muscle fibers, in men had a mean length of 38 mm, while in women the mean length was found to be 46 mm [10]. They described the sacrospinous ligament as a triangle with a medial base that attaches to the sacrum and coccyx and a lateral apex that attaches to the ischial spine [10].
Biomechanics
The sacrospinous ligament has fibers oriented to prevent upward tilting of the lower end of the sacrum; in order to accomplish this task, the sacrospinous ligament anchors to the ischium with the help of the sacrotuberous ligament [4]. As a result of this tilted orientation, these two ligaments oppose upward movement of the lower part of the sacrum under downward thrust at the sacrum’s upper end [3]. Since these two ligaments do not rotate in the same direction, a suggestion has been made that these two ligaments have oppositional effects on the pelvis [10]. If these ligaments unroll like a “contorted rope under tension,” then it can be considered that the sacrotuberous ligament will raise the ischium in relation to the sacrum while the sacrospinous ligament will lower the ischium in relation to the sacrum thus preventing any posterior rotation of the ilium with respect to the sacrum [10]. Even though these ligaments perform different functions, the two must be thought of as a couple since they are both closely correlated in regards to the volumes and cross-sectional areas of both ligaments [10].
Anterior sacrococcygeal ligament
Anatomy
The anterior sacrococcygeal ligament (Fig. 10) consists of irregular fibers that descend onto the pelvic surfaces of both the sacrum and coccyx [3]. It is the sacrococcygeal joint equivalent of the anterior longitudinal ligament as it attaches anterior to the sacrum and the coccyx [3].
Biomechanics
This ligament acts in concert with the posterior sacrococcygeal ligament to reinforce this joint [19].
Superficial posterior sacrococcygeal ligament
Anatomy
The superficial posterior sacrococcygeal ligament (Fig. 11) is a flat ligament that passes from the margin of the sacral hiatus to the dorsal coccygeal surface thus forming the roof of the lower sacral canal [3]. This ligament is the sacral equivalent of the ligamentum flavum [3].
Biomechanics
This ligament acts in concert with the posterior sacrococcygeal ligament to reinforce this joint while also covering the lower portion of the sacral canal [3, 19].
Deep dorsal sacrococcygeal ligament
Anatomy
The deep posterior sacrococcygeal ligament is the short part of the posterior sacrococcygeal ligament that passes from the back of the 5th sacral vertebral body to the dorsum of the coccyx [3]. This ligament is actually a continuation of the posterior longitudinal ligament [3].
Biomechanics
This ligament will connect the sacrum to the coccyx as well as acting in concert with the posterior sacrococcygeal ligament to reinforce this joint [3, 19].
Lateral sacrococcygeal ligament
Anatomy
There are two lateral sacrococcygeal ligaments, one on each side of the sacrum, that connect the first coccygeal transverse process to the inferolateral sacral angle [3]. These lateral sacrococcygeal ligaments will complete a foramen through which the 5th sacral spinal nerve will pass [3].
Biomechanics
Together with the anterior, posterior, and intercornual sacrococcygeal ligaments, the lateral sacrococcygeal ligaments help to stabilize the sacrococcygeal symphysis [20].
Intercornual sacrococcygeal ligaments
Anatomy
The intercornual ligaments will connect the sacral and coccygeal cornua on each side [3]. The cornua of the coccyx are superiorly projecting processes that arise from the upper part of coccyx [3]. The coccygeal cornua will articulate with the sacral cornua to form synovial joints [3]. There is also a fasciculus that will connect the sacral cornua to the coccygeal transverse processes [3].
Biomechanics
Together with the anterior, posterior, and lateral sacrococcygeal ligaments, the intercornual sacrococcygeal ligaments help to stabilize the sacrococcygeal symphysis [20].
Conclusions
This review will be useful to those who treat patients with pathology of the spine or who interpret imaging or investigate the anatomy of the ligaments of the vertebral column.
References
Agur AMR, Dalley AF (2005) Grant’s atlas of anatomy, 11th edn. Lippincott Williams and Wilkins, Baltimore
Alderink GJ (1991) The sacroiliac joint: review of anatomy, mechanics, and function. J Orthop Sports Phys Ther 13(2):71–84
Bannister LH, Berry MM, Collins P, Dyson M, Dussek JE, Ferguson MWJ (eds) (1996) Gray’s anatomy, 38th edn. Churchill Livingston, New York
Bogduk N (1997) Clinical anatomy of the lumbar spine and sacrum, 3rd edn. Churchill Livingston, New York
Chazal J, Tanguy A, Bourges M, Gaurel G, Escande G, Guillot M, Vanneuville G (1985) Biomechanical properties of spinal ligaments and a histological study of the supraspinal ligament in traction. J Biomech 18(3):167–176
Dickman CA, Rosenthal DJ, Perin NI (1999) Thoracoscopic spine surgery. Thieme Medical Publishers, New York
DonTigny RL (1985) Function and pathomechanics of the sacroiliac joint: a review. Phys Ther 65(1):35–44
Goode A, Hegedus E, Sizer P Jr, Brismee JM, Linberg A, Cook CE (2008) Three-dimensional movements of the sacroiliac joint: a systematic review of the literature and assessment of clinical utility. J Man Manip Ther 16(1):25–38
Gudavalli MR, Triano JJ (1999) An analytical model of lumbar motion segment in flexion. J Manipulative Physiol Ther 22:201–208
Hammer N, Steinke H, Slowik V, Josten C, Stadler J, Bohme J, Spanel-Borowski K (2009) The sacrotuberous and the sacrospinous ligament—a virtual reconstruction. Ann Anat 191(4):417–425
Hanson P, Magnusson SP, Sorensen H, Simonsen EB (1998) Differences in the iliolumbar ligament and the transverse process of the L5 vertebra in young white and black people. Acta Anat 163(4):218–223
Heylings DJA (1978) Supraspinous and interspinous ligaments of the human lumbar spine. J Anat 125(1):127–131
Hindle RJ, Pearcy MJ, Cross A (1990) Mechanical function of the human lumbar interspinous and supraspinous ligaments. J Biomed Eng 12:340–344
Jaumard NV, Welch WC, Winkelstein BA (2011) Spinal facet joint biomechanics and mechanotransduction in normal, injury and degenerative conditions. J Biomech Eng 133(7):071010
Jiang H, Raso JV, Moreau MJ, Russell G, Hill DL, Bagnall KM (1994) Quantitative morphology of the lateral ligaments of the spine: assessment of their importance in maintaining lateral stability. Spine 19(23):2676–2682
Lang J (1993) Clinical anatomy of the cervical spine. Thieme Medical Publishers, New York
Luk KDK, Ho HC, Leong JCY (1986) The iliolumbar ligament: a study of its anatomy, development, and clinical significance. J Bone Joint Surg 68:197–200
McGrath C, Hurst NH (2009) The long posterior sacroiliac ligament: a histological study of morphological relations in the posterior sacroiliac region. Joint Bone Spine 76(1):57–62
Moore KL, Dalley AF, Agur AMR (2010) Clinically oriented anatomy, 6th edn. Lippincott Williams & Wilkins, Baltimore
Morris CE (2005) Low back syndromes: integrated clinical management. McGraw-Hill, New York
Nachemson AL, Evans JH (1968) Some mechanical properties of the third human lumbar interlaminar ligament (ligamentum flavum). J Biomech 1(3):211–220
Neumann P, Keller T, Ekstrom L, Hult E, Hansson T (1993) Structural properties of the anterior longitudinal ligament. Spine 18(5):637–645
Neumann P, Keller TS, Ekstrom L, Perry L, Hansson TH, Spengler DM (1992) Mechanical properties of the human lumbar anterior longitudinal ligament. J Biomech 25(10):1185–1194
Onan OA, Heggeness MH, Hipp JA (1998) A motion analysis of the cervical facet joint. Spine 23(4):430–439
Paniabi MM, Goel VK, Takata K (1982) Physiologic strains in the lumbar spinal ligaments. Spine 7(3):192–203
Panjabi MM, Greenstein G, Duranceau J, Nolte LP (1991) Three-dimensional quantitative morphology of lumbar spinal ligaments. J Spinal Disord 4(l):54–62
Parke WW (1975) The spine, vol 1. W. B Saunders Co, Philadelphia
Pintar FA, Yoganandan N, Myers T, Elhagediab A, Sances A Jr (1992) Biomechanical properties of human lumbar spine ligaments. J Biomech 25(11):1351–1356
Pool-Goudzwaard AL, Hoek van Dijke G, Mulder P, Spoor C, Snijders CJ, Stoeckart R (2003) The iliolumber ligament: its influence on stability of the sacroiliac joint. Clin Biomech 18(2):99–105
Pool-Goudzwaard AL, Kleinrensink GJ, Snijders CJ, Entius C, Stoeckart R (2001) The sacroiliac part of the iliolumbar ligament. J Anat 199:457–463
Rissanen, PM (1976) The surgical anatomy and pathology of the supraspinous and interspinous ligaments of the lumbar spine, with special reference to ligament ruptures. Acta Orthop Scand (Suppl. 46): 1–100
Serhan HA, Varnavas G, Dooris AP, Patwardhan A, Tzsermiadonis M (2007) Biomechanics of the posterior lumbar articulating elements. Neurosurg Focus 22(1):E1
Sims JA, Moorman SJ (1996) The role of the iliolumbar ligament in low back pain. Med Hypotheses 46:511–515
Spalteholz W (1943) Hand atlas of human anatomy, 7th edn, vol 1. JB Lippincott Co, Philadelphia, London
Sturesson B, Selvik G, Uden A (1989) Movements of the sacroiliac joints: a roentgen stereophotogrammetric analysis. Spine 14:162–165
Vleeming A, Stoeckart R, Snijders CJ (1989) The sacrotuberous ligament: a conceptual approach to its dynamic role in stabilizing the sacroiliac joint. Clin Biomech 4(4):201–203
Vleeming A, Van Wingerden JP, Snijders CJ, Stoeckart R, Stijnen T (1989) Load application to the sacrotuberous ligament; influences on sacroiliac joint mechanics. Clin Biomech 4(4):204–209
Wilder DG, Pope MH, Frymoyer JW (1980) The functional topography of the sacroiliac joint. Spine 5(6):575–579
Willard FH, Carreiro JE, Manko W (1998) The long posterior interosseous ligament and the sacrococcygeal plexus. In: Procedings of the third interdisciplinary world congress on low back and pelvic pain. p. 207–209
Yahia LH, Garzon S, Strykowski H, Rivard CH (1990) Ultrastructure of the human interspinous ligament and ligamentum flavum: a preliminary study. Spine 15(4):262–268
Yamamoto I, Panjabi MM, Oxland TR, Crisco JJ (1990) The role of the iliolumbar ligament in the lumbosacral junction. Spine 15:1138–1141
Yong-Hing K, Reilly J, Kirkaldy-Willis WH (1976) The ligamentum flavum. Spine 1(4):226–234
Zander T, Rohlmann A, Bergmann G (2004) Influence of ligament stiffness on the mechanical behavior of a functional spinal unit. J Biomech 37(7):1107–1111
Conflict of interest
The authors have no coflict of interest to report.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Butt, A.M., Gill, C., Demerdash, A. et al. A comprehensive review of the sub-axial ligaments of the vertebral column: part I anatomy and function. Childs Nerv Syst 31, 1037–1059 (2015). https://doi.org/10.1007/s00381-015-2729-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00381-015-2729-z