
Abstract
Bats use brief calls for echolocation, suggesting that they might be more sensitive to brief sounds than non-echolocating mammals. To investigate this possibility, absolute thresholds for brief tones were determined for four species of bats: The Common vampire bat (Desmodus rotundus) and the Greater spear-nosed bat (Phyllostomus hastatus), both of which use frequency-modulated calls, the Egyptian fruit bat (Rousettus aegyptiacus), an echolocator that uses tongue-clicks rather than laryngeal calls, and the Dog-faced fruit bat (Cynopterus brachyotis), a non-echolocating species. Norway rats and a human were tested for comparison using the same acoustic stimuli. Contrary to expectations, the echolocating bats were not superior to non-echolocating mammals in detecting brief tones in the frequency range of their echolocation calls. Instead, all four species of bats were remarkably less sensitive than non-bats to brief sounds of 10 kHz and below. This implies that temporal summation in the mammalian auditory system can show large species differences, and that the detection of brief sound is likely influenced by the selective pressures on each species as well as by the physical integration of energy in the auditory system. Such species differences in function are expected to be reflected in the physiology of their auditory systems.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Echolocating bats are remarkable for their use of brief echolocation calls (0.2 to 10 ms) for orientation and prey capture (e.g., Hill and Smith 1984). In addition, their communication calls are nearly as brief—about 6–25 ms for the species in this report (e.g., Gould et al. 1973; Funakoshi et al. 1995; Carter et al. 2012). The use of such brief signals suggests that bats might be more sensitive to brief sounds than most mammals. Indeed, neurons tuned to short durations are more common in bats and respond to much shorter durations than neurons in other vertebrates (e.g., Covey and Casseday 1999; Brand et al. 2000). Yet, as the duration of a tone is shortened below a certain limit, greater signal amplitude is required for its detection. This is based at least in part on the integration of energy over time (e.g., Watson and Gengel 1969). For these reasons, the ability of bats to detect short-duration sounds, especially at the frequencies of their echolocation calls, is of interest.
For about 20 years, we were involved in determining pure-tone thresholds (audiograms) and sound-localization abilities in six species of bats (e.g., Koay et al. 1997; Heffner et al. 2015). When time permitted and healthy individuals were available, the ability to detect short-duration tones was also investigated. The initial expectation was that echolocating bats might be specialized for the detection of short-duration tones in the frequency range of their echolocation calls and would have smaller threshold shifts at short durations for those frequencies, compared to tones outside the range of their echolocation calls.
We present here the results for four species of bats in addition to results for two laboratory rats and a human observer that were obtained for comparison. Contrary to expectation, none of the three echolocating bats we tested were unusually sensitive to brief tones in the range of their echolocation calls. Instead, all three echolocating species, as well as a non-echolocating fruit bat, had unusually large threshold shifts to low-frequency brief tones (i.e., 10 kHz and lower).
Materials and methods
Tone detection thresholds were obtained for signals ranging from 2 ms (0 ms peak, 1 ms rise/fall) to 400 ms duration (398 ms peak, 1 ms rise/fall) for four species of bats and, for comparison, two hooded Norway rats and a human observer. The sample of bats included both suborders of bats (Yinpterochiroptera and Yangochiroptera), three different families, Old and New World lineages, echolocating and non-echolocating species, use and non-use of time cues for passive localization, diets ranging from fruit to meat/insects to blood, and sizes ranging from 20 to 180 g. Although few individuals of each species were available for testing, a range of lineages and lifestyles is represented. Frequencies were selected to sample the hearing range of each species. A conditioned suppression/avoidance procedure was used for the animal subjects (Heffner and Heffner 1995; Heffner et al. 2006b). Audiograms and sound-localization thresholds of the same individuals had been previously obtained so that all were trained and experienced observers by the beginning of the tests using short-duration tones. To ensure that the bats’ thresholds had not changed, test frequencies were rechecked using the 400-ms duration, but with a 1-ms rise/fall time. Comparing their thresholds with those previously obtained with the 400-ms duration and 10-ms rise/fall showed that the animals were still attending to the tones and gave no indication of any hearing loss or improvement as a result of using a 1-ms rise/fall time.
Subjects
The four species of bats are listed here in the order in which they were tested, with more detailed testing carried out on the first species examined. Because these exploratory tests took place over many years, not all durations were tested with each species.
Egyptian fruit bat (Rousettus aegyptiacus), one male and one female of unknown age. These megachiropteran bats echolocate using brief tongue-clicks (50–100 µs) to orient in caves, rather than the laryngeal pulses used by microchiropteran species. The clicks range in frequency from about 14 to 60 kHz, with peak energy around 25 kHz to 30 kHz (Yovel et al. 2011). At a level of 60 dB sound pressure level (SPL), their hearing range extends from 2.25 to 64 kHz (Koay et al. 1998).
Greater spear-nosed bat (Phyllostomus hastatus), one female approximately 5-years-old. This species uses frequency-modulated echolocation calls in the range of 25–100 kHz, with durations of about 1 ms (Bellwood 1986). At a level of 60 dB SPL, its hearing range extends from 1.8 to 105 kHz (Koay et al. 2002).
Dog-faced fruit bat (Cynopterus brachyotis), one male approximately 2-years-old. This is a small megachiropteran fruit bat that does not echolocate. At a level of 60 dB SPL, its hearing extends from 2.6 to 70 kHz (Heffner et al. 2006a).
Common vampire bat (Desmodus rotundus), two males approximately 4-years-old. This species uses frequency-modulated echolocation calls of about 1–2 ms in the range of 45 kHz to 100 kHz (Pye 1980; Bellwood 1986; Greenhall and Schmidt 1988). At a level of 60 dB SPL, its hearing extends from 700 Hz to 113 kHz (Heffner et al. 2013).
Hooded Norway rats (Rattus norvegicus), two males, both young adults. The rats served as non-echolocating comparisons to bats. Rats were chosen because their hearing range is comparable to that of Egyptian fruit bats; at a level of 60 dB SPL, their hearing extends from 530 Hz to 68 kHz (Heffner et al. 1994).
Human (Homo sapiens), a 27-years-old woman with normal hearing. She served as a control to ensure that the unusually large threshold shifts observed in the bats for frequencies of 10 kHz and lower were not due to some artifact in the acoustic stimuli. In addition, threshold shifts were determined for short durations at frequencies up to 18 kHz, higher than previously tested in humans. At a level of 60 dB SPL, this subject’s hearing range extended from 31 Hz to 17.6 kHz (e.g., Jackson et al. 1999).
Behavioral apparatus and procedure
The test environment and behavioral procedures have been described in detail for each species (Koay et al. 1998, 2002; Heffner et al. 2006a, 2013). The test room was a double-walled sound-proof chamber (Industrial Acoustics model 1204, 2.55 × 2.75 × 2.05 m) lined with carpeting and eggcrate foam to reduce reflections. The test cage was mounted on a tripod in the center of the room, approximately 1 m away from other surfaces to avoid sound reflections. A small reward spout was located at a comfortable height below the level of an animal’s ears while feeding (for a drawing of the test cage, see Koay et al. 2002).
The wire mesh test cage was custom built for each species to limit its movement in the sound field and to reduce sound reflections. A small spout was located in the front of the cage through which was dispensed a slow trickle of food reward via a syringe pump (fruit juice for R. aegyptiacus, C. brachyotis, and P. hastatus, and blood for D. rotundus), or, for rats, water. Reward delivery was controlled by an electrical contact switch that activated the syringe pump when an animal licked the reward spout. The syringe pump dispensed sufficient reward in one or two test sessions per day for the animals to maintain healthy body weights, yet at a rate slow enough to sustain feeding for at least 20–40 min per test session. A brief, mild constant-current shock could be delivered between the cage floor and the food/water spout. The shock was avoidable and was adjusted for each individual to the lowest intensity that elicited a reliable avoidance response—it was defined as mild because the animals never developed a fear of the reward spout and readily returned to it following trials in which they received a shock.
The important features of the behavioral procedure were the following. The animals were trained to eat or drink continuously from the spout until they heard a test tone, which signaled impending shock. Following the onset of four tone pulses, the animals had 2.0 s to break contact with the spout to avoid the shock (classified as a hit) (D. rotundus was given 0.5 s additional time to respond because the viscous blood it was drinking required more time to break electrical contact). Each shock was accompanied by a 0.3-s light flash from a 25 W bulb to signal its presence and termination (the broad-spectrum light is detectable by bats; cf., Baron et al. 1996). The shock duration was 0.3 s so the animals had to forego feeding only briefly.
For testing, an animal was placed in the test cage and allowed to obtain a food or water reward. Testing consisted of presenting a single frequency and duration for the entire session, varying only the amplitude of the tone to determine a threshold for that stimulus configuration. Testing alternated between low and high frequencies as follows: Egyptian fruit bat (R. aegyptiacus) 10 kHz, 45 kHz, 32 kHz, 56 kHz, 25 kHz, 20 kHz, 5.6 kHz; Dog-faced fruit bat (C. brachyotis) 50 kHz, 10 kHz, 20 kHz, 5.6 kHz, 10 kHz, 25 kHz; Common vampire bat (D. rotundus) 20 kHz, 5.5 kHz, 63 kHz, 4 kHz; Greater spear-nosed bat (P. hastatus) 64 kHz, 10 kHz, 50 kHz, 5.6 kHz, 8 kHz, 32 kHz, 20 kHz; Norway rat (R. norvegicus) 5.6 kHz, 10 kHz, 20 kHz, 32 kHz, 56 kHz, 64 kHz, 2 kHz, 16 kHz, 25 kHz, 45 kHz, 32 kHz. Testing focused on sampling frequencies in the echolocation and communication range of the subjects. It was not possible to test all possible frequencies and durations on each species due to time limitations and the exploratory nature of the studies.
Detection thresholds were determined by reducing the sound level in 5–10 dB steps until an animal no longer responded above chance (i.e., hit rate did not differ reliably from the false alarm rate, p > 0.01). Threshold was defined as the sound level at which the corrected hit rate equaled 0.5 (Hit rate − (hit rate × false alarm rate)). Approximately fifty to sixty tone trials were typically obtained in one session, enough to determine threshold for at least one stimulus configuration. Thresholds for each individual were replicated in subsequent sessions until they stabilized within a range of 3 dB, which took one to three additional sessions (details of the test procedure can be found in Heffner and Heffner 1995).
The human observer was tested with the same acoustical apparatus and procedure except that she sat in a chair facing the speaker with her head in the same position as the animal subjects. She pressed a hand-held button when she heard a sound, and the light flash following a sound trial provided feedback and confirmation that a sound had been presented. The order in which the frequencies were tested was 5.6 kHz, 10 kHz, 16 kHz, 18 kHz, 4 kHz, 2 kHz.
Acoustical apparatus and procedure
Pure tones were generated by a Krohn-Hite 2400 AM/FM Phase Lock Generator (R. aegyptiacus, humans, and Norway rats) or Zonic A&D 3525 signal generator (D. rotundus, C. brachyotis, P. hastatus) and pulsed on and off four times in a 2.0 s trial (five times in a 2.5 s trial for D. rotundus). To compensate for the different tone durations of 2–400 ms, the time between tone pulses varied from 498 to 100 ms. A rise/fall of 1 ms (Coulbourn S84-04, cosine gating) was used for all frequencies and durations. For the 2-ms duration, this meant that full amplitude was not sustained (0 ms plateau); this was the shortest stimulus configuration. For tone durations of 5 ms (3 ms peak, 1 ms rise/fall), the plateau amplitude was sustained for at least 6 cycles (at 2 kHz, the lowest frequency tested). All higher frequencies or longer durations contained a proportionally greater number of cycles.
The electrical signal was band pass filtered (Krohn-Hite 3202, 24-dB/octave roll-off, one-third octave above and below the center frequency), attenuated (Hewlett Packard 350D) and amplified (Crown D75) before being sent to either a ribbon tweeter (Panasonic EAS-10TH400C) or a piezoelectric tweeter (Motorola KSN1005). The electrical signal to the loudspeaker was also observed on an oscilloscope and the voltage adjusted as needed to keep the peak voltage the same for all durations of a tone. For human testing below 5 kHz, a 6-in. woofer (Infinity RS2000) was used. A speaker was placed approximately 1 m in front of the subject at the level of the ears.
Sound levels in the position of an animal feeding from the spout were calibrated daily using a 1/4-in. (0.64 cm) microphone (Brüel and Kjaer 4939), corrected for free-field with the protection grid on. The output of a preamplifier (Brüel and Kjaer 2669), and measuring amplifier (Brüel and Kjaer 2608) were then routed to a spectrum analyzer (Zonic A&D 3525) to continuously monitor the speaker output for harmonics or distortion.
The maximum stimulus amplitude of a tone pip was determined at the beginning of each test session by observing the output of the Brüel and Kjaer measuring system on an oscilloscope to determine whether the maximum amplitude of the tone at short-durations was the same as it was for long-durations. The measurements showed that the maximum peak-to-peak amplitudes of the tones dropped less than 1 dB at short durations—and when that occurred the voltage was increased to keep the amplitude of the sine wave emitted by the loudspeaker constant. Note that all durations used the same 1-ms rise/fall time so they all contained the same spectral splattering. Thus, the various durations of a particular frequency differed only in the duration of the tone.
Figure 1 shows the spectrum of an 8-kHz tone with a 1-ms rise/fall, 0-ms peak amplitude. The electronic signal was band passed filtered from 6.3 to 10 kHz, amplified by the Crown D75 amplifier, sent to the Motorola KSN1005 piezo speaker, measured using the previously described Brüel and Kjaer equipment and routed to the Stanford Research Systems SR770 FFT network analyzer. Measurement of the signal was synchronized with the signal pulse and averaged using a uniform window. Whereas a continuous tone would be shown as a single line at 8 kHz, the fast rise/fall of the tone produces an FFT that shows spectral spread containing energy between 7 and 9 kHz.

Spectrum of a 2-ms tone at 8 kHz with a 1-ms rise/fall time, 0-ms peak. The spectrum of a continuous tone would be a vertical line at 8 kHz
Results
Egyptian fruit bat, R. aegyptiacus
Thresholds for 400-ms duration tones using 1-ms rise/fall times were determined at the beginning of testing for 5.6, 10, 20, and 25 kHz and were within 3 dB of the previously determined thresholds of these individuals based on 10-ms rise/fall time (Koay et al. 1998).
Figure 2 shows that shortening the duration of a tone caused greater threshold shifts (reduced sensitivity) at low frequencies than at high frequencies. This effect of frequency was particularly strong at the two lowest frequencies, 5.6 kHz and 10 kHz, reaching more than 40 dB at 2 ms duration. The magnitudes of the threshold shifts for the two individuals were in good agreement, with a mean difference of 2.4 dB among all frequencies (range 0–7 dB).

Threshold shift for tones as a function of signal duration for four species of bats. The graphs are arranged in ascending order of the threshold shifts for 10-ms, low-frequency tones. All of these bats, regardless of whether they used echolocation, showed much greater threshold shifts than other mammals for short-duration tones of 10 kHz and lower frequencies
Because the threshold shifts at lower frequencies were far greater than typically observed in non-bat species (see below), including humans (e.g., Watson and Gengel 1969), we immediately began testing a human observer and two laboratory rats along with the Egyptian fruit bats to explore the possibility that the Egyptian fruit bats might have been responding on the basis of some anomaly in the stimuli or procedures.
Human
The human observer was tested using the same acoustical equipment used to test the Egyptian fruit bat, including presenting the stimuli via a loudspeaker rather than through headphones as is common in human studies. The threshold shifts shown in Fig. 3 are greater for the lower frequencies than for higher frequencies, a frequency effect that has been observed in humans by others (e.g., Plomp and Bouman 1959; Watson and Gengel 1969; Pedersen and Elberling 1972). Moreover, the magnitude of the human threshold shifts is similar to that found by Plomp and Bouman for 5-ms durations. However, the threshold shifts at 5.6 and 10 kHz for the 5-ms duration are far smaller than those found for the Egyptian fruit bats. Whereas the bats’ threshold shifts were 40 and 42 dB at 5 and 10 kHz, the human threshold shifts were only 15 and 10 dB, respectively. These results suggest that the large threshold shifts for the Egyptian fruit bats for short duration tones at 5.6 kHz and 10 kHz were not due to acoustic anomalies since the human observer showed typical threshold shifts at those frequencies when tested with the same acoustic apparatus.

Threshold shift as a function of signal duration for a human observer and for two hooded Norway rats. Neither showed the large threshold shifts at short durations seen for the bats at frequencies of 10 kHz and below. Note that whereas the rats showed a general frequency effect, their threshold shift for 32 kHz (asterisk) was as large as those for 10 kHz and lower frequencies
Hooded Norway rat, R. norvegicus
Thresholds for the 400-ms duration were determined for frequencies from 2 kHz to 64 kHz at the beginning of testing and agreed with previously published thresholds for rats (Heffner et al. 1994). The threshold shifts resulting from shorter durations are shown in Fig. 3. As with the Egyptian fruit bats and humans, there was a frequency effect in which the magnitude of the threshold shifts was greater for lower frequencies, reaching 26 dB at 2 kHz and 2 ms duration (Fig. 3); the individual rats were in close agreement, with a 2.8-dB mean difference (range 1.5–8.5 dB). As with the human subject, the rats did not show elevations of thresholds comparable to those of bats at frequencies of 10 kHz and lower. Because the rats and humans were tested with the same apparatus as the bats, we interpret the large low-frequency threshold shifts of the bats to reflect a true species difference.
Greater spear-nosed bat, P. hastatus
Thresholds for 400-ms duration tones using 1-ms rise/fall times were checked at the beginning of testing for 5.6, 10, 20, and 25 kHz. They were found to be within 2 dB of the previously determined thresholds of these individuals that used a 10-ms rise/fall time (Koay et al. 2002). As shown in Fig. 2, the Greater spear-nosed bat showed the largest threshold shifts of any of the bats tested—72 dB for 10-ms 5.6-kHz tones. The threshold shift would likely have been even greater at low frequencies had it been possible to produce an undistorted signal at 5.6 kHz of sufficient intensity to test the animal at shorter durations.
At the higher frequencies in the range of its echolocation calls, this species had the smallest threshold shift, 0.5 dB for the 50-kHz tone at 10 ms duration. Whether this bat is exceptional at 50 kHz, however, would require testing more than one individual.
Dog-faced fruit bat, C. brachyotis
Thresholds for 400-ms duration tones using 1-ms rise/fall times were rechecked at the beginning of testing for 5.6, 10, 20, 32 and 50 kHz. The replicated thresholds were within 2.5 dB of the previously determined thresholds of these individuals using a 10-ms rise/fall time (Heffner et al. 2006a).
Like the previous two species, the Dog-faced fruit bat showed large threshold shifts for 2-ms low-frequency tones, with maximum shifts of 41 and 44 dB for 10 kHz and 5.6 kHz, respectively (Fig. 2).
Common vampire bat, D. rotundus
Thresholds for 400-ms duration tones using 1-ms rise/fall times were checked at the beginning of testing for 4, 20, and 64 kHz; these thresholds were within 4 dB of the previously determined thresholds of these individuals for tones with the 10-ms rise/fall time (Heffner et al. 2013).
Because of their slightly better sensitivity to lower frequencies, we were able to test D. rotundus at 4 kHz, as well as up to 64 kHz. As can be seen in Fig. 2, this species also showed large threshold increases at low frequencies. The magnitudes of the threshold shifts for the two individuals were in good agreement (mean difference of 2.2 dB, range 1–4.5 dB). The Common vampire bats were tested with 10-ms tones for which they showed threshold shifts of 39 and 42 dB at 4 and 5.5 kHz, respectively. These shifts were larger than those shown by both the Dog-faced fruit bat and the Egyptian fruit bats at that duration. The Common vampire bats were not tested at shorter durations because it was not possible to generate shorter tones at sufficient intensities at low frequencies without distortion.
Summary of results
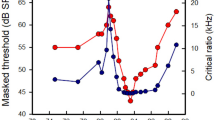
Figure 4 summarizes the results for the four species of bats. Absolute thresholds for the brief tone pips (10-ms duration, 1-ms rise/fall) are shown along with the published audiograms that had been obtained using long-duration tones (400-ms duration, 10-ms rise/fall). Comparing these thresholds for long- versus short-duration tones shows that it is detection in the lower frequency range of the audiogram that is strongly compromised by brief signals. Moreover, the lower frequencies that were affected most by short durations are those reportedly used in communication (thin bars along the X axis); frequencies used in echolocation (bold bars along the X axis) are only mildly affected.

Summary of results. Shown are the previously published audiograms obtained using 400-ms tones with 10-ms rise/fall times (gray lines), thresholds for the 400-ms tones with 1-ms rise/fall (open circles), and thresholds for the10-ms tones with 1-ms rise/fall (indicated by 10’s). Horizontal lines along the X axes indicate the range of the echolocation calls (bold bars) and communication calls (thin bars). Thresholds for the brief 10-ms tone pips reveal the large elevations of thresholds at frequencies of 10 kHz and below that are used primarily for communication, compared to the much smaller threshold elevations at the higher frequencies used for echolocation. Audiograms from Koay et al. (1998, 2002), Heffner et al. (2006a, 2013). Communication and echolocation frequencies: Phyllostomus hastatus from Belwood (Bellwood 1986), Boughman and Wilkinson (1998), Desmodus rotundus from Schmidt (1972), Carter et al. (2012), San Pedro and Allendes (2017), Rousettus aegyptiacus from Pye (1980), Holland et al. (2004), Cynopterus brachyotis (communication only) from Funakoshi et al. (1995)
Also shown in Fig. 4 are the replications for the 400-ms tones using the shorter 1-ms rise/fall time (open circles). The close agreement between the replicated thresholds and original thresholds indicates that the bats remained motivated and had suffered no hearing loss that might affect their thresholds for brief sounds. It also shows that the very short rise/fall used in these tests neither improved detection nor compromised it.
Discussion
Rather than examining the physical energy integration in the ear, this report has taken a functional approach to gather data on whether species differ in their ability to detect brief sounds and, if so, what that might imply about specializations possible in the auditory system and perhaps the selective pressures behind those specializations.
The primary finding is that all four species in this sample of bats, both echolocators and non-echolocators, showed a very large frequency effect, with threshold shifts of more than 30 dB for brief tones at 10 kHz and below. Whereas the maximum threshold shifts for the human and rats tested here never noticeably exceeded 25 dB, even at the 2-ms duration, the threshold shifts for bats at 10 kHz and lower ranged from 40 to 75 dB. Indeed, the threshold shift for the Greater spear-nosed bat for 5.6 kHz at 10 ms was so large (72 dB) that we were unable to generate a signal of sufficient amplitude to determine its threshold for a 2-ms signal. Nor is it always necessary to test at such short durations to show that bats have greater threshold shifts at low frequencies. In the case of the Egyptian fruit bat, the species tested first and most extensively, the difference between low and high frequencies begins to emerge at 50-ms durations. For the Greater spear-nosed bat a difference is suggested at 100 ms (Fig. 2).
At higher frequencies, the bats’ threshold shifts are similar to those of non-bat species in the upper portions of their hearing ranges. For example, with 10-ms signals, the bats’ threshold shifts for 20 kHz and higher ranged between about 2 and 15 dB, comparable to the 6–10 dB shifts seen for the human with 5-ms signals at 10 to 18 kHz. Other species showed similar threshold shifts at 20 kHz and higher, as demonstrated by their performances with the 10-ms duration (Fig. 5). Thus, the echolocating bats did not show exceptional sensitivity to brief signals in the frequency range of their echolocation calls despite their reliance on very brief signals for echolocation.

Threshold shift as a function of frequency for 10-ms duration tone pulses for the animals tested in this study and five additional species for which similar data are available (data for humans tested with 10-ms tones (H1) are included for comparison to the human 2 tested with 5-ms durations for this report (H2)). Bats have an unusually large threshold shift for frequencies of 10 kHz and lower, but are similar to other mammals for frequencies of 20 kHz and higher. The difference between the bats and non-bats for frequencies of 10 kHz and below is statistically significant, p < 0.005 (see text). Seal, Phoca vitulina (Kastelein et al. 2010a); H1 (Plomp and Bouman 1959); House mouse, Mus musculus (Ehret 1976), Harbor porpoise, Phoca vitulina (Kastelein et al. 2010b), and Dolphin, Tursiops truncatus (Johnson 1968). The range of threshold shifts among non-bats is shaded for emphasis
Although threshold shifts were generally greater for lower frequencies than for higher frequencies in all six species tested here (Figs. 2 and 3), the relationship was sometimes non-monotonic. Occasionally a high frequency elicited a greater threshold shift than some lower frequencies. A clear example of this is the rat’s threshold shift at 32 kHz (Fig. 3). This suggests that simple physics may not be the sole determiner of the detection of brief sounds and that biological factors also play a role. With purely physical determinants, threshold shifts would be expected to change progressively with frequency. However, it is likely that a species’ sensory abilities reflect the selective pressures acting on it, and these appear to differ between species.
Regardless of small fluctuations, the key result is that in four different species of bats representing different lineages, lifestyles, and echolocation, the threshold shifts for short-duration tones at 10 kHz and lower are far larger than those at higher frequencies. They are also larger than threshold shifts so far found in other mammals (Fig. 5). Indeed, the difference between bats and other mammals in Fig. 5 is statistically significant. Using the Mann–Whitney U test, the probability that the absence of overlap of the four bat species with the six non-bat species at 10 kHz or at 5.6 kHz is due to chance is p < 0.005 (Siegel 1956, for n1 = 4, n2 = 6, U = 0). Understanding the anatomical and physiological differences underlying this dichotomy may provide insight into more general hearing mechanisms.
Comparison with other bats
The unusually large threshold shifts for short-duration tones of 10 kHz and lower has not been previously observed in bats. However, there have been only two studies of temporal summation in bats, both of which looked at only one frequency.
The first study tested an Egyptian fruit bat (R. aegyptiacus) and reported that its ability to detect a 12-kHz tone increased by only 6 dB for durations from 100 to 2 ms (Suthers and Summers 1980). That 6-dB shift is much smaller than the 38-dB shift we found when going from 100 to 2 ms for the same species, at 10 kHz (Fig. 1). However, Suthers and Summers expressed reservations about their behavioral method as they were only able to train one of their six bats. Moreover, their thresholds for long-durations tones in the region of 10 and 12 kHz were more than 40 dB higher than later found for this species (Koay et al. 1998). Thus, their methods may not have been optimal for determining thresholds.
In a second study, Schmidt and Thaller (1994) tested six Mexican free-tailed bats (Tadarida brasiliensis) at 40 kHz, with durations from 500 to 2 ms and in two different levels of masking noise. The threshold shifts they found in the presence of their low-level masking noise were 23 dB (2 ms), 17 dB (5 ms), and 12 dB (10 ms). The threshold shifts we found for bats in that frequency range (32–50 kHz) were similar. Thus, the threshold shifts of Mexican free-tailed bats are consistent with our finding that bats are not unusually sensitive to brief sounds at high frequencies.
Temporal summation in other mammals
Bats show a consistent pattern of a large threshold shift for brief sounds at 10 kHz and lower frequencies. That pattern has not yet been observed in other mammals. However, exact comparisons with other species are hampered by differences in test procedures, stimuli, and the durations examined. Moreover, many were tested over only a portion of their hearing range. The following is a brief description of results obtained at multiple frequencies and at durations of 15 ms or shorter.
Rhesus macaques (Macaca mulatta) were tested in an early comparative study (Clack 1966). Durations as short as 15 ms were presented at frequencies from 250 Hz to 1 kHz using headphones and from 2 to 8 kHz using loudspeakers. An effect of frequency was evident in which the greatest threshold shift of about 39 dB occurred at the lowest frequency, 250 Hz. The magnitude of this threshold shift was comparable to that observed in the megachiropteran bats, but the effect of frequency was gradual, unlike the abrupt increase that occurred at 10 kHz seen in bats. A later study of macaques (O’Connor et al. 1999) examined durations only as short as 25 ms and found no effect of frequency.
Eight domestic House mice (Mus musculus) were tested at twelve frequencies ranging from 1 kHz to 120 kHz and durations from 1 to 500 ms (Ehret 1976). As can be seen in Fig. 5, the results revealed no effect of frequency. At a duration of 10 ms, the largest threshold shift was 11 dB, occurring at 60 kHz and 20 kHz. At 1 ms, the shortest duration examined, threshold shifts reached as high as 19 dB (20 kHz). Neither duration produced threshold shifts comparable to those observed in bats. This lack of an effect of frequency and overall smaller threshold shifts is especially noteworthy since the hearing range of mice is comparable to that of bats.
Three species have been tested under water and despite the different acoustics, their threshold shifts were comparable to those of other non-bat species that hear in air. Underwater testing at 10-ms durations was carried out with two Harbor seals (Phoca vitulina) at six frequencies from 1 kHz to 40 kHz (Kastelein et al. 2010a). As shown in Fig. 5, threshold shifts were greater for low frequencies (20 dB) than for high frequencies (7 dB). Threshold shifts of the Harbor seals at high frequencies were comparable to those of bats but remained smaller than the threshold shifts shown by bats at low frequencies.
A Bottlenose dolphin (Tursiops truncatus) was tested at 1 kHz to 100 kHz at a duration of 10 ms. Threshold shifts ranged from 6 dB at 45 kHz to only 13 dB at 1 kHz (Fig. 5). Although there was a small effect of frequency, that effect may have been confounded with absolute threshold (Johnson 1968). The absence of an abrupt increase in threshold shifts at low frequencies in this echolocating mammal supports the inference that the results reported here for bats may not be an adaptation for echolocation.
A single Harbor porpoise (Phocoena phocoena) was tested at a duration of 10 ms using frequency-modulated tones from 500 Hz to 150 kHz (frequency range ± 1% around center frequency, Kastelein et al. 2010b). The results showed no consistent frequency effect (Fig. 3). This echolocating porpoise also did not show the dramatic threshold shifts seen in bats, again suggesting that the phenomenon may be specific to bats, both echolocating and non-echolocating, and not attributable (at least so far) to a general requirement of echolocation.
Two conclusions can be drawn from the available data comparing temporal summation in bats to that reported for other mammals as summarized in Fig. 5. First, the effect of frequency varies in magnitude with non-bat species, including echolocating Cetacea, showing at most a relatively small and gradual increase in threshold for brief sounds progressing from high to low frequencies. Second, the threshold shifts found in bats are similar to those of other species at frequencies of 20 kHz and higher, but become markedly greater below 20 kHz.
Frequency effect or threshold effect?
All of the subjects examined for this report—bats, rats, and human—showed threshold shifts that increased as the frequency of brief tones became lower. However, it has been suggested that what we are seeing may not be an effect of frequency, but rather an effect of absolute threshold (Clack 1966). This is because, beginning at 4 kHz (the frequency to which humans are most sensitive), absolute thresholds gradually increase toward lower frequencies (e.g., Jackson et al. 1999), as do threshold shifts for brief sounds, leading to the possibility that the magnitude of the threshold shifts is related to the sensitivity of the ear rather than to frequency. Although humans have rarely been tested at the upper end of their hearing range where absolute thresholds also increase, the results for the human tested for this report suggest that threshold shifts do not increase with short durations at frequencies near the upper limit of human hearing.
To determine whether the threshold shifts for bats show an effect of frequency and/or absolute threshold, we examined the relationship between the magnitude of the 10-ms threshold shifts to both frequency and absolute threshold for the Norway rats and three species of bats tested for this report (the vampire bat was not included because too few frequencies were tested for a statistical analysis; the human data do not sample a broad enough frequency range to dissociate frequency from overall sensitivity).
Multiple regression analyses and subsequent partial regression plots (Fig. 6) reveal that, for the bats, frequency remains a reliable predictor of threshold shift when the effect of absolute threshold is removed (p values between 0.003 and 0.018), but absolute threshold is not a reliable predictor if the effect of frequency is removed (p values between 0.07 and 0.438). Rats show relatively small threshold shifts and neither frequency nor absolute threshold is a significant predictor of threshold shift when the effect of the other is removed. Thus, the large threshold shifts at lower frequencies in bats cannot be attributed to differences in absolute thresholds at these frequencies. Instead there is a frequency effect in which the magnitude of the threshold shifts for short-duration tones increases as frequency decreases, independent of absolute sensitivity.

Partial regression plots showing the relationship between threshold shift (for 10-ms tones), signal frequency (in log scale), and absolute threshold (for long-duration tones). Note that the values for both x- and y-axes are unstandardized residuals, and each plot represents the correlation between threshold shift and each independent variable, with the effect of the other independent variable removed from both. Signal frequency (upper row) is a reliable predictor of threshold shift for bats even when the contribution of absolute threshold is removed from both variables. In contrast, absolute threshold (lower row) is not a reliable predictor of threshold shift for bats when the contribution of frequency is removed. Neither frequency nor absolute threshold is a reliable predictor of threshold shift in rats. These plots were obtained in SPSS by selecting “Regression > Linear > Method: Enter”, followed by selecting the Dependent (Threshold shift) and Independent (Absolute threshold and Frequency) variables, and then selecting the “Plots > Produce all partial plots” option
Effect of the 1-ms rise/fall time on the detectability of a tone
The spectrum of a continuous pure tone (sine wave) is a single line at the frequency of the tone. However, turning a pure tone on and off with a fast rise/fall time of 1 ms results in a spread of energy into adjacent frequencies (Fig. 1), referred to as “spectral spatter” or “spectral spread”. For this reason, the original audiograms of the bats used a 10-ms onset and offset to reduce or eliminate spectral spread for the 400-ms tones. Thus, the question arises as to whether the 1-ms rise/fall time used in this study gave rise to spectral splatter that affected the detectability of the signals.
There are several reasons for believing that spectral splatter played no significant role in the detectability of the tone pips used here. First, the greatest amplitude of these signals was at the frequency of the pure tone and that amplitude was kept constant for all durations. Although it has been suggested that the spread of energy into adjacent frequencies reduces the amplitude of the tone (e.g., Watson and Gengel 1969; Yost 2007), we did not find that to be the case with the equipment used here. In short, the 1-ms rise/fall time did not reduce the amplitude of the tone, but resulted in energy of much lower amplitude at adjacent frequencies (see Fig. 1). Moreover, the bats’ thresholds for the 400-ms tones obtained with 1-ms versus 10-ms rise/fall times show no significant differences (Fig. 4) suggesting that the spectral splatter resulting from the 1-ms rise/fall had little effect on the detectability of the tones. Finally, Dallos and Johnson (1966) demonstrated that the detectability of a tone is determined by its duration, not by its rise/fall time; they found that thresholds for 1-kHz tones with rise/fall times as short as 0 ms were higher than those for longer rise times, indicating that spectral splatter played little if any role in the detection of the 1-kH tone used in their tests. In summary, although long rise/fall times are typically used when determining audiograms, that may not be a crucial factor especially with modern signal generators and loudspeakers.
Physiological implications
Proposals for the physiological basis of temporal summation vary. Some localize it to the cochlea, whereas others place it in the central auditory system. Descriptions of these can be found in Watson and Gengel (1969), Klump and Maier (1990), and Formby et al. (2002). The unusually large frequency effect in bats seen at frequencies of 10 kHz and lower may lead to insight into the mechanisms underlying temporal summation. This will require comparing auditory anatomy and physiology of bats with that of non-bat species that have similar hearing ranges.
As one example, the current results may be relevant for some aspects of duration selectivity seen in the midbrain and auditory cortex of bats. Neurons selective for short durations have been found in several species of bats (e.g., Casseday et al. 1994; Covey and Casseday 1999), as well as to a lesser extent in other species (Aubie et al. 2012). However, neurons tuned to frequencies below 20 kHz are rarely reported in bats, and those tuned below 10 kHz seem not to exist (e.g., Wu and Jen 2008; Morrison et al. 2014). In part, this may be because some investigations focused on echolocation frequencies. Yet responses to frequencies in the lower portion of the hearing range seem rare even when surveys include broad frequency ranges and responses to somewhat longer durations (greater than 50 ms). In rats, however, although duration-selective neurons are not as common, they are found throughout the hearing range (Perez-Gonzalez et al. 2006); this is consistent with the relatively small threshold shifts in the lower portion of the hearing range reported here for rats. The rarity of responses to lower frequencies in bats is consistent with the possibility that detection of lower frequencies may be suppressed for tones shorter than about 25 ms.
Evolutionary implications
The evolutionary history of bats remains uncertain. Arguments are made for the common ancestor of bats to have had both echolocation and flight, with laryngeal echolocation subsequently lost by species in the family Pteropodidae. Yet it cannot be ruled out that the common ancestor of bats was capable of flight, but that echolocation evolved multiple times in different lineages (for a review, see Teeling et al. 2016). Our results suggest another auditory feature that might be exclusive to bats, perhaps lending support to the single origin hypothesis.
As noted in “Materials and methods”, this initial sample of four species of bats is broad in that it includes echolocators and non-echolocators, and a wide range of body sizes, habitats, and diets. All show unusually large threshold shifts for brief sounds in the lower portion of their hearing ranges (10 kHz and below) that is so far unknown in any other mammal, suggesting that this sensory function might be a distinguishing feature of the bat lineage. It is not associated with echolocation since it also appears in the non-echolocating Dog-faced fruit bat, but it is not present in echolocating cetaceans. This result was unexpected. Because bats rely on extremely short signals for echolocation, it was thought that, if they differed from other mammals, it would take the form of better sensitivity to short sounds, but that was not observed. It would be of value to examine temporal summation in species that rely on long constant-frequency echolocation calls for comparison to help determine whether this is an auditory feature of all bats or if it differs in that specialized branch of Yinpterochiroptera.
The unusual insensitivity to short-duration tones at frequencies below the echolocation frequencies, but within the communication range leads us to wonder what might underlie its presence in bats. What benefit might be gained by reducing sensitivity to brief signals at communication frequencies? Is this a loss of function at low frequencies, a sacrifice that accompanied the use of echolocation, but has not yet been lost in non-echolocating bats? The fact that the non-echolocating Dog-faced fruit bat shows the smallest effect, and the non-laryngeal echolocating Egyptian fruit bat shows the second smallest effect, supports this possibility, but additional species should be examined. It is also possible that this is a gain of function that allows bats to suppress detection of lower frequencies in circumstances where they might interfere with detection of more important signals. Behavioral tests that explore the masking effects of lower frequencies in species representing additional families and echolocation styles may answer these questions.
References
Aubie B, Sayegh R, Faure PA (2012) Duration tuning across vertebrates. J Neurosci 32:6373–6390
Baron G, Stephan H, Frahm HD (1996) Comparative neurobiology in chiroptera. Birkhauser Verlag, Berlin
Bellwood JJ (1986) Foraging behavior, prey selection, and echolocation in phyllostomid bats (Phyllostomidae). In: Nachtigall PE, Moore PWB (eds) Animal sonar processes and performance. Plenum Press, New York, pp 601–605
Boughman JW, Wilkinson GS (1998) Greater spear-nosed bats discriminate group mates by vocalizations. Animal Behav 55:1717–1732
Brand A, Urban A, Grothe B (2000) Duration tuning in the mouse auditory midbrain. J Neurophysiol 84:1790–1799
Carter GG, Logsdon R, Arnold BD, Menchaca A, Medellin RA (2012) Adult vampire bats produce contact calls when isolated: acoustic variation by species, population, colony, and individual. PLoS ONE 7:338791. https://doi.org/10.1371/journal.pone.0038791
Casseday JH, Ehrlich D, Covey E (1994) Neural tuning for sound duration: role of inhibitory mechanisms in the inferior colliculus. Science 264:847–850
Clack D (1966) Effect of signal duration on the auditory sensitivity of humans and monkeys (Macaca mulatta). J Acoust Soc Am 40:1140–1146
Covey E, Casseday JH (1999) Timing in the auditory system of the bat. Ann Rev Physiol 61:457–476
Dallos PJ, Johnson KR (1966) Influence of rise/fall time upon short-tone threshold. J Acoust Soc Am 40:1160–1163
Ehret G (1976) Temporal auditory summation for pure tones and white noise in the house mouse (Mus musculus). J Acoust Soc Am 59:1421–1427
Formby C, Heinz MG, Aleksandrovsky IV (2002) Temporal integration of sinusoidal increments in the absence of absolute energy cues. J Speech Lang Hear R 45:1285–1296
Funakoshi K, Zubaid A, Matsumura S (1995) Regular pulse emission in some megachiropteran bats. Zool Sci 12:503–505
Gould E, Woolf NK, Turner DC (1973) Double-note communication calls in bats: occurrence in three families. J Mammal 54:998–1001
Greenhall AM, Schmidt U (1988) Natural history of vampire bats. CRC Press, Boca Raton
Heffner HE, Heffner RS (1995) Conditioned avoidance. In: Klump GM, Dooling RJ, Fay RR, Stebbins WC (eds) Methods in comparative psychoacoustics. Birkhäuser, Basel, pp 73–87
Heffner HE, Heffner RS, Contos C, Ott T (1994) Audiogram of the hooded Norway rat. Hear Res 73:244–248
Heffner HE, Koay G, Heffner RS (2006a) Behavioral assessment of hearing in mice—conditioned suppression. In: Crowley J et al (eds) Current protocols in neuroscience. Wiley, New York, pp 8.21D.1–8.21D.15
Heffner RS, Koay G, Heffner HE (2006b) Hearing in large (Eidolon helvum) and small (Cynopterus brachyotis) non-echolocating fruit bats. Hear Res 221:17–25
Heffner RS, Koay G, Heffner HE (2013) Hearing in American leaf-nosed bats. IV: the common vampire bat, Desmodus rotundus. Hear Res 296:42–50
Heffner RS, Koay G, Heffner HE (2015) Sound localization in common vampire bats: acuity and use of the binaural time cue by a small mammal. J Acoust Soc Am 137:42–52
Hill JE, Smith JD (1984) Bats: a natural history. University of Texas Press, Austin
Holland RA, Waters DA, Rayner JMV (2004) Echolocation signal structure in the megachiropteran bat Rousettus aegyptiacus. J Exp Biol 207:4361–4369
Jackson LL, Heffner RS, Heffner HE (1999) Free-field audiogram of the Japanese macaque (Macaca fuscata). J Acoust Soc Am 106:3017–3023
Johnson CS (1968) Relation between absolute threshold and duration-of-tone pulses in the bottlenosed dolphin. J Acoust Soc Am 43:757–763
Kastelein RA, Hoek L, Wensveen PJ, Terhune JM, deJong CAF (2010a) The effect of signal duration on the underwater hearing thresholds of two harbor seals (Phoca vitulina) for single tonal signals between 0.2 and 40 kHz. J Acoust Soc Am 127:1135–1145
Kastelein RA, Hoek L, Terhune JM, de Jong CAF, Wensveen PJ (2010b) The effect of signal duration on the underwater detection thresholds of a harbor porpoise (Phocoena phocoena) for single frequency-modulated tonal signals between 0.25 and 160 kHz. J Acoust Soc Am 128:3211–3222
Klump GM, Maier EH (1990) Temporal summation in the European starling (Sturnus vulgaris). J Comp Psychol 104:94–100
Koay G, Heffner HE, Heffner RS (1997) Audiogram of the big brown bat (Eptesicus fuscus). Hear Res 105:202–210
Koay G, Heffner RS, Heffner HE (1998) Hearing in a megachiropteran fruit bat, Rousettus aegyptiacus. J Comp Psychol 112:371–382
Koay G, Bitter KS, Heffner HE, Heffner RS (2002) Hearing in American leaf-nosed bats. I: Phyllostomus hastatus. Hear Res 171:96–102
Morrison JA, Farzan F, Fremouw T, Sayegh R, Covey E, Faure PA (2014) Organization and trade-off of spectro-temporal tuning properties of duration-tuned neurons in the mammalian inferior colliculus. J Neurophysiol 111:2047–2060
O’Connor KN, Barruel P, Hajalilou R, Sutter ML (1999) Auditory temporal integration in the rhesus macaque (Macaca mulatta). J Acoust Soc Am 106:954–965
Pedersen CB, Elberling C (1972) Temporal integration of acoustic energy in normal hearing persons. Acta Otolaryng 74:398–405
Perez-Gonzalez D, Malmierca MS, Moore JM, Hernandez O, Covey E (2006) Duration selective neurons in the inferior colliculus of the rat: topographic distribution and relation of duration sensitivity to other response properties. J Neurophysiol 95:823–836
Plomp R, Bouman MA (1959) Relation between hearing threshold and duration for tone pulses. J Acoust Soc Am 31:749–758
Pye JD (1980) Adaptiveness of echolocation signals in bats: flexibility and evolution. Trends Neurosci 10:232–235
San Pedro AR, Allendes JL (2017) Echolocation calls of free-flying common vampire bats Desmodus rotundus (Chiroptera: Phyllostomidae) in Chile. Bioacoustics 26:153–160
Schmidt U (1972) Social calls of juvenile vampire bats (Desmodus rotundus) and their mothers. Zool Beitr 4:310–316
Schmidt S, Thaller J (1994) Temporal auditory summation in the echolocating bat, Tadarida brasiliensis. Hear Res 77:125–134
Siegel S (1956) Nonparametric statistics. McGraw-Hill, New York
Suthers RA, Summers CA (1980) Behavioral audiogram and masked thresholds of the megachiropteran echolocating bat, Rousettus. J Comp Physiol 136:227–233
Teeling EC, Jones G, Rossiter SJ (2016) Phylogeny, genes, and hearing: Implications for the evolution of echolocation in bats. In: Fenton MB, Grinnell A, Popper AN, Fay RR (eds) Bat bioacoustics: Springer handbook of auditory research. Springer, New York, pp 25–54
Watson CS, Gengel RW (1969) Signal duration and signal frequency in relation to auditory sensitivity. J Acoust Soc Am 46:989–997
Wu CH, Jen PH-S (2008) Echo frequency selectivity of duration-tuned inferior collicular neurons of the big brown bat, Eptesicus fuscus, determined with pulse-echo pairs. Neuroscience 156:1028–1038
Yost WA (2007) Fundamentals of hearing. Academic Press, San Diego
Yovel Y, Geva-Sagiv M, Ulanovsky N (2011) Click-based echolocation in bats: not so primitive after all. J Comp Physiol A 197:515–530
Acknowledgements
Supported by National Institutes of Health Grants R01-DC02960 and R15-DC009321. We thank Dr. Greg Meyer for his help in understanding the details of partial regression procedures in SPSS. We thank the Smithsonian's National Zoo, the Lubee Bat Conservancy, and the Milwaukee County Zoo for the loan of bats.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no competing interests.
Human and animal rights
All animals were cared for following the Guide for the Care and Use of Animals. All procedures were conducted with the approval of the institutional committees overseeing research with humans and with animal subjects, and were in accord with the ethical standards of the institutional research committee and with the 1964 Helsinki declaration and its later amendments.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Heffner, R.S., Koay, G. & Heffner, H.E. Bats are unusually insensitive to brief low-frequency tones. J Comp Physiol A 205, 583–594 (2019). https://doi.org/10.1007/s00359-019-01349-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00359-019-01349-x