
Abstract
Web-building spiders construct their own vibratory signaling environments. Web architecture should affect signal design, and vice versa, such that vibratory signals are transmitted with a minimum of attenuation and degradation. However, the web is the medium through which a spider senses both vibratory signals from courting males and cues produced by captured prey. Moreover, webs function not only in vibration transmission, but also in defense from predators and the elements. These multiple functions may impose conflicting selection pressures on web design. We investigated vibration transmission efficiency and accuracy through two web types with contrasting architectures: sheet webs of Eratigena agrestis (Agelenidae) and tangle webs of Latrodectus hesperus (Theridiidae). We measured vibration transmission efficiencies by playing frequency sweeps through webs with a piezoelectric vibrator and a loudspeaker, recording the resulting web vibrations at several locations on each web using a laser Doppler vibrometer. Transmission efficiencies through both web types were highly variable, with within-web variation greater than among-web variation. There was little difference in transmission efficiencies of longitudinal and transverse vibrations. The inconsistent transmission of specific frequencies through webs suggests that parameters other than frequency are most important in allowing these spiders to distinguish between vibrations of prey and courting males.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Communication signals of animals are transmitted through a signaling medium to reach the intended receiver. Ideally, the signals reach the receiver with a minimum loss of energy and without attenuation or degradation. Any signal alterations can be kept to a minimum by matching the properties of the signal with the properties of the signaling medium (Endler 1992).
Web-building spiders build their own signaling environment and thus provide a fascinating system to study interactions between signals and the signaling medium (Witt 1975; Maklakov et al. 2003; Vollrath and Selden 2007). Being able to alter the signaling medium, web-building spiders may be able to optimize signal transmission. Yet, spider webs provide not only a medium for sexual signaling, with males courting on females’ webs, they also serve for prey detection and prey capture, and as shelter from the elements and predators. In response to different evolutionary pressures, spiders have evolved to produce very different types of webs, and to use their webs in different ways (Vollrath and Selden 2007).
Using their webs as an extension of their sensory systems, spiders are quickly alerted to the presence of any prey, predator, competitor or prospective mate that contacts the web and thus causes web vibrations. However, only orb webs have been studied in detail for vibration transmission (Masters 1984; Landolfa and Barth 1996), so variation in transmission properties across contrasting web architectures is not known. Here, we propose to study sheet and tangle webs for their signal transmission characteristics. This study will advance our understanding as to how web architecture affects the transmission of vibrations.
Our study species are the hobo spider, Eratigena agrestis (Agelenidae), which builds a sheet or funnel web (Fig. 1a), and the western black widow spider, Latrodectus hesperus (Theridiidae), which builds a tangle web (Fig. 1b). The mostly two-dimensional web of E. agrestis consists of a funnel-shaped retreat from which extends a dense sheet of multiple layers of silk threads. The three-dimensional tangle web of L. hesperus consists of loose threads that are suspended above substrate from supporting threads, with sticky capture threads connecting the tangle to the ground.

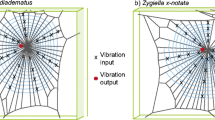
Web structures of Eratigena agrestis and Latrodectus hesperus. Schematic drawing illustrating a the sheet web of Eratigena agrestis, and b the tangle web of Latrodectus hesperus. Webs of E. agrestis consist of a two-dimensional sheet of silk with a funnel at one end serving as a retreat. Webs of L. hesperus consist of a dense three-dimensional tangle of threads. Glue-coated capture threads extend from the tangle to the ground. The stars indicate the locations where the spiders typically reside while waiting for prey. The position of the spiders indicates the location of their retreats. Illustrations reproduced from Vibert et al. (2014)
These two types of web are highly variable depending on the age and size of the spider, her nutritional state, the web age and the geometry of the surrounding micro-habitat (Blackledge and Zevenbergen 2007; S.V. pers. obs.). Parameters such as web size, thread density, number of web attachment points, and overall web shape, all vary. The high degree of variability of these web parameters invokes doubts whether sheet and tangle webs can serve as a medium for reliable and consistent transmission of vibratory signals or cues.
Our objectives were: (1) to characterize longitudinal and transverse vibration transmission of sheet and tangle webs, and (2) to investigate the consistency of frequency transmission both within and between webs.
Methods
Study spiders
We collected juvenile black widow and hobo spiders from Island View Beach, on the Saanich peninsula of Vancouver Island, British Columbia, Canada. We housed spiders singly in large Petri dishes (2.5 × 15 cm) under a photoperiod of 12L:12D. We fed juvenile spiders twice a week, and adults once a week, either crickets (Acheta domesticus) or house flies (Musca domestica), providing water ad libitum. To facilitate web-spinning, we confined randomly selected virgin females, 10–21 days post maturity, inside wood-framed boxes (20 × 20 × 15 cm for hobo spiders; 30 × 30 × 20 cm for black widow spiders) for 10–15 days. The mean (SD) masses of adult female hobo spiders and black widow spiders were 186 (37) mg and 226 (41) mg, respectively.
General procedures
Recordings
We recorded web vibrations inside a sound-dampened room. To keep extraneous vibratory noise to a minimum (see Fig. S1 for recordings of background noise), we also placed the entire recording apparatus on a concrete table covered with a thick rubber mat. We recorded web vibrations using a laser Doppler vibrometer (LDV) (Polytec OFV-2500 with OFV-534 sensor head; Polytec GmbH, Waldbronn, Germany), the associated Polytec data acquisition system VIB-E-220, and the VibSoft 4.8 data acquisition software. In sequence, we recorded vibrations at bandwidths of 0–1000 Hz (objective 1) and 0–500 Hz (objective 2), using a frequency resolution of 0.08 Hz, and a vibrometer range of (10 mm/s)/V. All individual recordings lasted 12.8 s, the longest possible data acquisition period.
Frequency sweeps
We used software developed with LabView Graphical Programming for Instrumentation (National Instruments Corporation) to generate constant-amplitude frequency sweeps of 1–1000 Hz (for the piezoelectric vibrator) and 1–500 Hz (for the loudspeaker), each 12.8 s in duration. The computer generating the sweeps was connected to an amplifier (Creek OBH-21SE) and thence to the vibrating element. The volume of both the computer and the amplifier was set to maximum during playback.
Vibration playback devices
The piezoelectric vibrator consisted of a piezoelectric diaphragm 44 mm in diameter and 0.23 mm thick (part no. CEB-44D06, CUI Inc., Tualitin, OR, USA) that we cut into a pointed wedge shape and attached to a metal dowel. The resonant frequency of the piezoelectric element was 0.6 kHz and the resonant impedance was 1000 Ω. We clamped the dowel to an adjustable stand such that the piezoelectric vibrator was suspended pointing down. The vibrator was oriented in the same way for recording of both longitudinal and transverse vibration of the webs. Because many individual silk threads interconnect in the sheet of an E. agrestis web and in the upper tangle of a black widow webs, vibrations of the piezo in a single direction (always perpendicular to the sheet or upper tangle) produced vibrations that propagated in multiple orientations through the web, including those that we measured (i.e., longitudinal and transverse). Here and throughout the paper, when we refer to longitudinal and transverse vibrations, we refer to vibrations of the sheet or tangle web as a whole, rather than of individual threads.
Struggling prey or courting males do not generate vibrations >200 Hz on either E. agrestis or L. hesperus webs (Vibert et al. 2014). We therefore decided to restrict subsequent frequency sweeps to a biologically relevant range of <500 Hz. As the piezoelectric vibrator produced low-magnitude vibrations below 200 Hz (maximal velocity: 1.1 μm/s on E. agrestis webs and 1.9 μm/s on L. hesperus webs; Fig. 2a–d), we further decided to deploy a vibrator that produced high-magnitude vibrations at low frequencies (maximal velocity: 2.85 mm/s on E. agrestis webs and 4.57 mm/s on L. hesperus webs; Fig. 2e–f). The vibration-inducing apparatus consisted of a modified unenclosed loudspeaker (12 Ω; diameter 14 cm), with its cone removed and a metal rod (1 mm thick × 18 mm long) attached to the center of the dust cap. We attached the loudspeaker to an adjustable stand so that it could be suspended over the webs.
We used the adjustable stand to position the vibration playback device such that the tip of the piezo element, or the vibrating rod, was facing downward, 1 cm above the topmost threads. Using a small jack, we then raised the box containing the web until the tip of the vibrator was in contact with the web causing a slight tensing of the threads. We positioned the tip of the playback device at the entrance to the funnel for E. agrestis, and at the top of the tangle in the middle of the web for L. hesperus; these are the positions where spiders await prey during their active hunting period.
Recording protocol
Prior to recordings, we removed each female spider from her 10- to 21-day-old web. If a web was damaged during the removal of the female, we returned her to the web for another 1–2 days to effect repairs. Onto each empty web, we positioned small squares [1 mm2, mass (mean ± SE): 0.9 ± 0.3 mg] of reflective tape (Polytec Retroreflective Sheeting; Polytec GmbH, Waldbronn, Germany). When using the piezoelectric vibrator, we placed three reflective squares in a straight line at a distance of 1, 20, and 70 mm, respectively, from the tip of the vibrator. When using the loudspeaker, we placed five reflective squares as follows: one square (#0 in Fig. 3) just 1 mm from the tip of the rod and two squares each at a distance of 20 mm (#1 and #2) and 70 mm (#3 and #4) from the tip of the vibrator, with squares #0, #1, and #3 forming an approximately straight line and squares #0, #2, and #4 forming another (Fig. 3). The orientation of these lines varied somewhat from web to web and was guided by the shape of the web, the location of the retreat, and the areas of the web that were most dense (to facilitate placement of reflective squares). Because E. agrestis webs are sheet-like, we attached reflective squares at the intersection of many individual threads, whereas on L. hesperus webs we placed squares at an intersection of three or more threads. We then placed the web-containing box on the vibration-proof table and focussed the LDV beam on the piece of reflective tape closest (1 mm) to the vibrator.

Representative piezo sweeps (a–d) and loudspeaker sweeps (e, f) at 1 mm from the input vibration. Frequency sweeps were generated by a piezoelectric disk or modified loudspeaker and recorded on webs of Eratigena agrestis (a, c, e) and Latrodectus hesperus (b, d, f) by a laser Doppler vibrometer 1 mm away from the source of the input vibration (see Fig. 4). The graphs show the velocity of vibrations (mm/s) over frequency (Hz). The beam of the vibrometer was either parallel or perpendicular to the plane of the webs to capture longitudinal vibrations (a, b) or transverse vibrations (c–f), respectively. Note the different scales of the y axes: μm/s in a–d and mm/s in e and f

Experimental design for frequency sweeps. A 12-s frequency sweep from 1 to 500 Hz was transferred onto the web by a modified loudspeaker (S). The resulting transverse vibration was recorded by a laser Doppler vibrometer (LDV). The laser beam was set perpendicular to the plane of the web and focused on small squares (1 mm2) of reflective tape that were placed 1 mm (#0), 20 mm (#1 and #2), and 70 mm (#3 and #4) from the tip of the vibrating rod. Frequency sweeps were recorded seven times in a fixed sequence (see methods for details). Illustration modified from Vibert et al. (2014)
For recordings using the piezoelectric vibrator, we started the 1–1000 Hz frequency sweep and the LDV recording of the resultant web vibration, simultaneously. Once the sweep was completed, we repeated the procedure focussing on the pieces of reflective tape 20 and 70 mm from the piezoelectric vibrator. For 28 E. agrestis webs and 28 L. hesperus webs, we obtained three recordings (one each at a distance of 1, 20, and 70 mm from the piezoelectric vibrator), with 14 webs assigned to recordings of longitudinal vibrations and 14 webs assigned to recordings of transverse vibrations. To record transverse vibrations, we oriented the LDV beam at a 90° angle to the “plane” of the web (i.e., beam approximately vertical). To record longitudinal vibrations, we oriented the LDV beam as parallel as possible to the “plane” of the web (i.e., beam approximately horizontal and pointing toward the vibrator). For these recordings, the squares of reflective tape were also oriented at approximately 90° to the plane of the web.
We did not measure transverse vibrations in the plane of the webs (lateral vibrations) because we assumed the results would have been similar to those we obtained with longitudinal vibration measurements. In the upper tangle of the widow web, silk threads of all orientations are interconnected in a three-dimensional “sheet” approximately parallel to the substrate, and in the sheet of the hobo web, silk threads are oriented in all directions within a roughly two-dimensional “plane”. Any measurement taken in the “plane” of an E. agrestis web, or parallel to the three-dimensional upper tangle of a L. hesperus web, would include both lateral and longitudinal vibrations, thus we expected that taking a second set of measurements would have yielded similar results.
For recordings using the loudspeaker, we recorded only transverse vibrations, with the beam of the LDV oriented vertically. We simultaneously triggered the 1–500 Hz frequency sweep and the LDV recording of the resultant web vibration. Once we had completed a sweep, we focussed the laser on the next square. From each web, we obtained seven recordings, focussing the beam of the LDV in a fixed sequence on the following reflective squares: #0 (1 mm), #1 (20 mm), #3 (70 mm), #0 (1 mm), #2 (20 mm), #4 (70 mm), and again #0 (1 mm). If the vibration profile at 1 mm differed among subsequent recordings on the same web, we began a new series of recordings. We then used the mean of the three recordings as a baseline for calibrating the 20 and 70 mm recordings as described below. We used a total of 16 individual webs of each species. Here, we recorded only transverse vibrations because the transmission of longitudinal and transverse vibrations was similar when using the piezoelectric vibrator.
Analysis
Characterizing longitudinal and transverse vibration transmission through webs
From the VibSoft 4.8 data acquisition software, we obtained a measure of a web’s velocity (measured in mm/s) at 1 Hz intervals in response to the 1–1000 Hz sweep played through the piezoelectric vibrator. For each web, we used the vibration measurement curve obtained at the 1 mm recording point as the baseline against which we measured the vibration transmission efficiency of the web. For the 20 and 70 mm recording positions, we calculated the transmission efficiency curve by taking the log10 of the ratio of velocity at the measurement point to the reference velocity at that point.
Consistency of vibration frequency transmission between and within webs
From the 1–500 Hz sweep data, we obtained a measure of transmission consistency. For each web, we averaged the data of the three square #0-recordings and used these mean data as the baseline against which we estimated the transmission efficiency of the web. For recordings at positions #1–#4 we calculated transmission with reference to the average velocity at position #0 using the same formula as for the 1–1000 Hz sweep. We then calculated the repeatability of each transmission efficiency measurement using within-web and among-web variance components (σ2 and σ 2 A ) (Nosil and Crespi 2006). We calculated \(s_{A}^{2}\), an estimate of \(\sigma_{A}^{2}\), using the group mean square (MS) from the ANOVA results. In the equation below, n is the number of measurements taken on each web (here, n = 2)
Repeatability values can vary between 0 and 1. A repeatability value near 0 indicates that nearly all the variance in transmission efficiency results from differences between separate measurements taken on the same web. A repeatability value near 1 indicates that repeated measurements on the same web result in very similar transmission efficiency profiles. We calculated repeatability values at 10 Hz intervals between 10 and 200 Hz, the range of dominant frequencies produced by prey and males (Vibert et al. 2014). We calculated the repeatability for the distances 20 mm (squares #1 and #2) and 70 mm (squares #3 and #4) from the point of the input vibration (Fig. 3).
Results
Longitudinal and transverse vibration transmissions of webs
Webs of both E. agrestis and L. hesperus exhibited similar transmission efficiencies of longitudinal and transverse vibrations (Fig. 4a–d). Longitudinal vibrations tended to be more greatly attenuated than transverse vibrations, except at 70 mm in E. agrestis webs, where the reverse was true. The transmission efficiency tended to be greater at 20 mm than at 70 mm in both web types, but the difference was much more pronounced in E. agrestis webs (Fig. 5a–d), indicating that vibrations attenuate differently over distance in E. agrestis and L. hesperus webs. Furthermore, in both web types there was a tendency for transmission loss to increase with frequency up to about 300 Hz, after which it became relatively consistent. There are large differences in transmission efficiency among webs of either species (Supplementary material, Table S1), as indicated by the large standard deviations of the data. It seems that the highly variable and complex web structure leads to great differences in vibration transmission profiles. On average, input vibrations of <50 Hz cause the least mean attenuation. In both species, this low-frequency range overlaps with that of vibrations produced by struggling prey and courting males that generate vibrations mostly between 10 and 75 Hz (Vibert et al. 2014).

Comparison of transmission efficiencies for longitudinal and transverse web vibrations. Transmission efficiency of longitudinal and transverse vibrations were measured on webs of Eratigena agrestis (a, b) and Latrodectus hesperus (c, d) (n = 14 each) at distances of 20 and 70 mm from a piezoelectric vibrator generating the vibration. Mean transmission efficiency (solid line) and 95 % confidence intervals (shaded band) are shown. Transmission efficiency of vibrations (dB) was measured by comparing vibrations recorded at a distance of 20 and 70 mm from the vibrator with those obtained just 1 mm away from it

Comparison of transmission efficiencies of longitudinal and transverse vibrations measured at two distances from the point of vibration input. Longitudinal transmission efficiencies of vibrations were measured on webs of Eratigena agrestis (a, b) and Latrodectus hesperus (c, d) (n = 14 each) at distances of 20 and 70 mm from a piezoelectric vibrator generating the vibration. Mean transmission efficiency (solid lines) and 95 % confidence intervals (shaded bands) are shown. Transmission efficiency of vibrations (dB) was measured by comparing vibrations recorded at a distance of 20 and 70 mm from the vibrator with those obtained just 1 mm away from it
Consistency of vibration frequency transmission between and within webs
The transmission efficiency of transverse vibrations caused by 1–500 Hz frequency sweeps was highly variable in both E. agrestis (Fig. 6a–d) and L. hesperus (Fig. 6e–h). To determine whether this variability was driven mostly by between-web or within-web variation, we calculated the repeatability of measurements at 10 Hz intervals between 10 and 200 Hz (Supplementary material, Table S2). For webs of E. agrestis, the repeatability was on average 0.23 and 0.26 for data collected at a distance of 20 and 70 mm, respectively, from the input vibration. For 87.5 % of all frequencies tested, the repeatability of measurements was <0.5 (Fig. 7a). For L. hesperus webs, the repeatability was on average 0.25 and 0.39 for data collected at a distance of 20 and 70 mm, respectively, from the input vibration. For 85 % of all vibration frequencies tested, the repeatability of measurements was <0.5 (Fig. 7b). These results show that most of the total variance in transmission efficiency resulted from variation within the same web.

Comparison of transmission efficiencies of transverse vibrations measured at two locations equidistant to the point of vibration input. Mean transmission efficiencies (solid line) and standard deviations (gray band) were measured on webs of Eratigena agrestis (a–d) and Latrodectus hesperus (e–h) (n = 16 each). The transmission efficiencies of vibrations (dB) were measured by comparing vibrations recorded at a distance of 20 and 70 mm from the loudspeaker vibrator with those just 1 mm away from it

Repeatability of transmission efficiency measurements of transverse vibrations. The repeatability of transmission efficiency measurements of transverse vibrations was calculated for recordings taken at a distance of 20 mm (black squares) and 70 mm (gray circles) from the point of the vibration input, using webs of Eratigena agrestis (a) and Latrodectus hesperus (b). Vibrations were caused by frequency sweeps ranging between 1 and 500 Hz. Repeatability values were calculated at 10 Hz intervals from 0 to 200 Hz, the range of dominant frequencies produced by prey and males. Values can vary between 0 and 1. A value near 0 indicates that most of the variance in transmission efficiency results from variations between the two measurements taken on the same web
Discussion
Our data can be summarized as follows: (1) the sheet webs of E. agrestis and tangle webs of L. hesperus revealed large variability in transmission efficiency across the frequency range of the input vibrations; (2) transmission loss is quite variable even within a single web, with the result that transmission profiles obtained at different locations equidistant from the point of the input vibration can be very dissimilar; (3) within-web rather than between-web variation plays a larger role in explaining the variability among transverse vibrations (suggesting that the same is likely true for longitudinal vibrations); and (4) there is little difference in transmission efficiency between longitudinal and transverse vibrations.
Our findings that both sheet and tangle webs have similar transmission efficiencies for longitudinal and transverse vibrations contrast with findings reported for orb webs. This is in part because we measured longitudinal and transverse vibrations of entire webs rather than through individual silk threads, as has been done for orb webs. In webs of Nuctenea sclopetaria, longitudinal vibrations attenuate less than transverse ones (Masters and Markl 1981). The same is true in webs of Nephila clavipes: longitudinal vibrations attenuate five times less than transverse ones (Landolfa and Barth 1996). That sheet and tangle webs have similar transmission efficiencies for longitudinal and transverse vibrations is likely due to the architecture of these types of webs. The tangle of a tangle web is formed by silk threads deposited in all directions, from vertical to horizontal orientations, with multiple points of contact between threads. A sheet web is composed of mostly horizontal threads criss-crossing each other and deposited on top of each other until a dense mat is formed. Given the density of nodes, or points of contact, in this type of web, longitudinal and transverse web vibrations probably combine in a complex manner. This may explain why on E. agrestis webs longitudinal vibrations were more greatly attenuated than transverse vibrations at 20 mm, but the opposite was true at 70 mm (Fig. 4a, b).
Vibrations can be characterized by envelope (amplitude modulation), spectral (frequency), and temporal patterns (duration, periodicity of repeating elements). Information can be conveyed by one or all of these parameters (Bradbury and Vehrencamp 1998). Frequency has been shown to be of significance in birds (Podos 2001), amphibians (Ryan et al. 1990), and insects (Rebar et al. 2009). For E. agrestis and L. hesperus, specific frequencies are probably not of great relevance, as they are not transmitted consistently through the web. However, lower frequencies in the range that includes the dominant frequencies of male and prey vibrations (Vibert et al. 2014) tended to be transmitted with the least attenuation through both web types. Thus, frequency may be important to these spiders in that it must fall into a certain range to transmit information. In L. hesperus, the amplitude of a male courtship vibration determines whether or not a female will respond with aggression (Vibert et al. 2014). Temporal patterns may also play an important role, as they do in other spider systems in which spiders communicate via substrate-borne vibrations. For instance, both spectral and temporal properties of male signals affect responses of female spiders in the Ctenidae (Schüch and Barth 1990; Schmitt et al. 1994), Lycosidae (Gibson and Uetz 2008) and Salticidae (Sivalinghem et al. 2010).
The sheet and tangle webs we tested here exhibit a striking degree of variability in the transmission efficiency of vibratory cues or signals, and both types of webs are apparently inferior media for consistent transmission of specific frequencies. Our input vibrations were greatly attenuated and large differences in transmission frequency profiles were noticeable both within and between webs. These phenomena reflect the complexity and variability of web architecture. Contrary to orb webs, E. agrestis sheet webs and L. hesperus tangle webs are not uniform structures. The density of threads and their orientation, degree of tension, number of connections, and distance to anchor points all vary greatly from one area of the web to another, and undoubtedly affect a web’s transmission characteristics. Tangle and sheet webs may be the product of potentially conflicting adaptive imperatives. Superior transmission characteristics of web vibrations may have been compromised in favor of prey capture. For a hunting spider to decide whether to treat the source of a web vibration as prey, she may need to know only the amplitude of that vibration. To, then, effectively capture the prey, the spider may need to know only the location of the prey on the web. Both types of information are likely sensed by E. agrestis and L. hesperus, even if some vibration characteristics are transmitted inconsistently.
Frequency sweeps are useful to obtain an overall view of the transmission properties of a medium (Masters 1984; Landolfa and Barth 1996) based on controlled input vibrations. Greater insight into the relevance of these properties for the spider receiving the vibrations transmitted through the web could be gained by examining the transmission of specific vibrations such as those generated by courting males, struggling prey, wind, or debris hitting the web. One could study, in particular, the parameters of courtship signals or prey cues that are transmitted with the highest fidelity, and whether certain areas of the web transmit the information that is relevant to the receiving spider better than others.
References
Blackledge TA, Zevenbergen JM (2007) Condition-dependent spider web architecture in the western black widow, Latrodectus hesperus. Anim Behav 73:855–864. doi:10.1016/j.anbehav.2006.10.014
Bradbury JW, Vehrencamp L (1998) Principles of animal communication. Sinauer Associates, Sunderland
Endler JA (1992) Signals, signal conditions, and the direction of evolution. Am Nat 139:S125–S153 http://www.jstor.org/stable/2462431. Accessed 30 Jan 2011
Gibson JS, Uetz GW (2008) Seismic communication and mate choice in wolf spiders: components of male seismic signals and mating success. Anim Behav 75:1253–1262. doi:10.1016/j.anbehav.2007.09.026
Landolfa MA, Barth FG (1996) Vibrations in the orb web of the spider Nephila clavipes: cues for discrimination and orientation. J Comp Physiol A 179:493–508. doi:10.1007/BF00192316
Maklakov AA, Bilde T, Lubin Y (2003) Vibratory courtship in a web-building spider: signalling quality or stimulating the female? Anim Behav 66:623–630. doi:10.1006/anbe.2003.2245
Masters WM (1984) Vibrations in the orbwebs of Nuctenea sclopetaria (Araneidae). Behav Ecol Sociobiol 15:207–215. doi:10.1007/BF00292977
Masters WM, Markl H (1981) Vibration signal transmission in spider orb webs. Science 213:363–365. doi:10.1126/science.213.4505.363
Nosil P, Crespi BJ (2006) Experimental evidence that predation promotes divergence in adaptive radiation. Proc Nat Acad Sci USA 103:9090–9095. doi:10.1073/pnas.0601575103
Podos J (2001) Correlated evolution of morphology and vocal signal structure in Darwin’s finches. Nature 409:185–188. doi:10.1038/35051570
Rebar D, Bailey NW, Zuk M (2009) Courtship song’s role during female mate choice in the field cricket Teleogryllus oceanicus. Behav Ecol 20:1307–1314. doi:10.1093/beheco/arp143
Ryan MJ, Fox JH, Wilczynski W, Rand AS (1990) Sexual selection for sensory exploitation in the frog Physalaemus pustulosus. Nature 343:66–67. doi:10.1038/343066a0
Schmitt A, Schuster M, Barth FG (1994) Vibratory communication in a wandering spider, Cupiennius getazi: female and male preferences for features of the conspecific male’s releaser. Anim Behav 48:1155–1171. doi:10.1006/anbe.1994.1348
Schüch W, Barth FG (1990) Vibratory communication in a spider: female responses to synthetic male vibrations. J Comp Physiol A 166:817–826. doi:10.1007/BF00187328
Sivalinghem S, Kasumovic MM, Mason AC, Andrade MCB, Elias DO (2010) Vibratory communication in the jumping spider Phidippus clarus: polyandry, male courtship signals, and mating success. Behav Ecol. doi:10.1093/beheco/arq150
Vibert S, Scott C, Gries G (2014) A meal or a male: the ‘whispers’ of black widow males do not trigger a predatory response in females. Front Zool 11:4. doi:10.1186/1742-9994-11-4
Vollrath F, Selden P (2007) The role of behavior in the evolution of spiders, silks, and webs. Annu Rev Ecol Evol Syst 38:819–846. http://www.jstor.org/stable/30033881. Accessed 11 Oct 2013
Witt PN (1975) The web as a means of communication. Biosci Commun 1:7–23
Acknowledgments
We thank Stephen Takács for making the frequency sweeps, Stephen DeMuth for some illustrations, and Friedrich Barth and two anonymous reviewers for meticulous reviews and constructive comments. Funding was provided by a Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery Grant and by an NSERC Industrial Research Chair to G.G., with Scotts Canada Ltd. as the industrial sponsor.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Ethical approval
All applicable international, national, and institutional guidelines for the care and use of animals were followed.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Vibert, S., Scott, C. & Gries, G. Vibration transmission through sheet webs of hobo spiders (Eratigena agrestis) and tangle webs of western black widow spiders (Latrodectus hesperus). J Comp Physiol A 202, 749–758 (2016). https://doi.org/10.1007/s00359-016-1113-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00359-016-1113-0