
Abstract
When honeybee foragers leave the nest, they receive nectar from nest mates for use as fuel for flight or as binding material to build pollen loads. We examined whether the concentration of nectar carried from the nest changes with the need for sugar. We found that pollen foragers had more-concentrated nectar (61.8 %) than nectar foragers (43.8 %). Further analysis revealed that the sugar concentration of the crop load increased significantly with waggle duration, an indicator of food-source distance, in both groups of foragers. Crop volume also increased with waggle duration. The results support our argument that foragers use concentrated nectar when the need for sugar is high and suggest that they precisely adjust the amount of sugar in the crop by altering both volume and nectar concentrations. We also investigated the impact of the area where foragers receive nectar on the crop load concentration at departure. Although nectar and pollen foragers tend to load nectar at different areas in the nest, area did not have a significant effect on crop load concentration. Departing foragers showed an average of 2.2 momentary (<1 s) begging trophallactic contacts before leaving the nest. They might be rejecting nectar with inappropriate concentrations during these contacts.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Animals usually store energy reserves, such as fat and glycogen, for use when energy requirement exceeds the energy supply from food (Schmidt-Nielsen 1997). Bees, by contrast, store limited reserves in the body and rely largely on food in the crop for energy supply during flight (Panzenböck and Crailsheim 1997). When leaving the nest, honeybees (Apis mellifera) receive food (as nectar) from nest mates through trophallaxis (Beutler 1950; von Frisch 1967), and use it as fuel (but see Núñez 1970). The nectar is held in the crop and sent gradually to the midgut, where sugars are absorbed to generate energy for flight (Crailsheim 1988; Blatt and Roces 2001). It is estimated that the sugar available outside the crop is between 1 and 2 mg (Visscher et al. 1996).
Previous studies showed that the amount of nectar that foragers carry from the nest is adjusted by several factors, including target food type (nectar or pollen), food-source distance, and reward variability (Beutler 1950, 1951; Harano et al. 2013, 2014; Harano and Sasaki 2015; Tan et al. 2015). The frequency of begging trophallactic contacts with nest mates (putative reception of nectar) is also reported to change, depending on food-source profitability and variability (Núñez 1970; Farina 1996; De Marco and Farina 2001, 2003). By contrast, information on the concentration of the crop load upon departing the nest is limited. Nectar in the nest is available at various concentrations. Individual workers collect nectar at concentrations ranging mainly from 20 to 60 % (Seeley 1986), because a colony allocates workers to various food sources to respond to changes in profitability of those food sources (Seeley 1995). The collected nectar is stored after concentrating up to 80 % by in-nest bees (Park 1946). Recently, Tan et al. (2015) reported that Asian honeybee A. cerana foragers carry both more, and more- concentrated nectar as fuel when collecting from variable food sources than from a constant source. The results suggest that foragers can alter the concentration of nectar carried from the nest to adjust the total sugar content in the crop. However, this type of fuel adjustment has not been confirmed in the other contexts.
Honeybee foragers tend to specialize in either nectar or pollen collection (Free 1960; Page 2013; Harano and Sasaki 2015). Pollen foragers not only use the nectar carried from the nest as fuel but also use it as “glue” to build pollen loads (Parker 1926; Hodges 1952). They regurgitate part of the crop content during a foraging trip and mix it with collected pollen to give cohesiveness, which appears to be essential to build pollen loads on corbiculae. Pollen foragers carry more nectar from the nest than nectar foragers because of the need for “glue” (Beutler 1950, 1951; Harano et al. 2013, 2014; Harano and Sasaki 2015). Pollen loads are composed of pollen grains and an almost equal amount of sugar (Roulston et al. 2000). This also indicates the deposition of a large amount of sugar by pollen foragers in the process of load building.
This work examined whether A. mellifera foragers use nectar at different concentrations depending on the necessity for sugar during nectar and pollen collection. First, we investigated the effect of forager subtypes, nectar, or pollen foragers. In addition to need of nectar for glue, pollen foragers may need to carry more fuel from the nest than nectar foragers, because they may not collect nectar that can be used as fuel for the return trip (Harano and Sasaki 2015). Because of their larger sugar requirement, pollen foragers might use more-concentrated nectar than nectar foragers. Second, we investigated the effect of food-source distance on the concentration of nectar carried by foragers from the nest. Foragers increase the amount of nectar carried from the nest (Beutler 1950; Harano et al. 2013, 2014; Harano and Sasaki 2015) with distance to the food source. They might also carry more-concentrated nectar to increase the sugar content of the crop load for visiting distant food sources.
Another reason to expect use of concentrated nectar in bees with a large need for sugar arises from the cost of carrying nectar load. It is known that a large nectar load can become a burden for flight, and increases energy expenditure depending on its mass (Wolf et al. 1989). The amount of dissolved sugar in the crop can be increased by increasing either crop load volume or concentration. When the requirement for sugar increased, if foragers only increase volume of crop load to meet the requirement, the mass of crop load would become so large to cause a considerable cost for flight. Thus, we expect foragers use concentrated nectar to reduce the mass-dependent “carriage cost” when needing to carry a large amount of sugar. Reduction of carriage costs by concentrating collected nectar has been suggested elsewhere for honeybees and other insects (Wittmann and Scholz 1989; Hendrichs et al. 1992; Nicolson and Human 2008).
To determine the mechanism of selective use of nectar at a particular concentration, we also investigated the effect of the location where bees receive food in the nest on the concentration of the crop load at departure. The concentration of nectar available to foragers may differ, depending on the area in the nest. Freshly collected, probably less-concentrated, nectar is unloaded by foragers at the “delivery area” close to the nest entrance (Seeley 1989), whereas concentrated nectar or honey is transferred to and stored in the top part of the nest (Seeley and Morse 1976). Although collected nectar is distributed rapidly through the colony (Seeley 1989; Goyret and Farina 2005), foragers might take advantage of the potentially biased distribution of fresh or concentrated nectar to access nectar at a particular concentration.
Materials and methods
Experiments were carried out for 13 days between 1 July and 24 August 2014. (The average temperature for the 13 days was 26.9 °C and within the range of average temperatures between 22.7 and 30.4 °C, measured at the Fuchu observation site of the Japan Meteorological Agency, approx. 12 km from our study site; Japan Meteorological Agency 2016).
Bees and hives
We used a colony of crossbred race of A. mellifera (Apidae, Apini) which was basically derived from A. mellifera ligustica. The colony consisted of 3000–4000 workers, a single queen, and several tens or hundreds of males. It was housed in a two-frame observation hive (OBS; dimensions, 55 × 55 × 4 cm) in an air-conditioned room at about 26 °C. The hive had an entrance at the bottom connected to the outside via a transparent corridor (9.5 × 33.0 × 3.0 cm) made of polyvinyl chloride except for the wooden floor. Bees were allowed to forage outside freely. The OBS had a wedge-shaped wooden block at the entrance to lead returning foragers to one side of the comb to facilitate the observation of waggle dancers. An area adjacent to the entrance (dance floor) on the side of the comb was covered with nylon mesh (18 × 20 cm) for marking dancing bees (Harano et al. 2013).
Effects of forager type, food-source distance, and nectar reception comb area
The volume and concentration of crop content at nest departure were determined for nectar and pollen foragers. Bees meeting the following two criteria were regarded as nectar foragers: (1) bees returning to the hive without pollen loads; and (2) bees making an offering contact (transferring crop contents through trophallaxis) with nest mates for more than 3 s after returning to the hive. Pollen foragers were identified from pollen loads on their corbiculae at return to the hive. For simplification, only pollen foragers with relatively large pollen loads (larger than pollen basket) were sampled, because such foragers are expected to be more specialized for pollen collection than bees with small pollen loads (Page 2013; Harano and Sasaki 2015). If a pollen forager showed an offering trophallactic contact with nest mates, it was not used for analysis.
We focused primarily on waggle dancers to examine the effect of food-source distance. Nectar and pollen foragers were marked with chalk powder through the nylon screen after performing waggle dances (Harano et al. 2013). The waggle duration indicates the distance to a food source being exploited by dancers (von Frisch 1967). We recorded durations of 3–6 waggle runs for each forager using a digital video camera (GZ-HM450, JVC KENWOOD, Yokohama, Japan) or IC-voice recorder (ICR-PS401RM, Sanyo, Tokyo, Japan; Harano et al. 2013) and averaged times to obtain a representative value. We also sampled non-dancers. Data from non-dancers were used only in the analysis of effects of forager type and loading area.
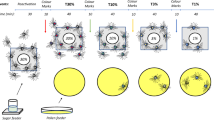
To determine any relationship between loading area and the concentration of crop load at departure, all begging trophallactic contacts (putative food reception) after unloading of collected food were recorded for each forager using a digital video camera or IC-voice recorder. The surface of the lower comb was divided into four areas (Fig. 1), and the area where a forager performed the longest begging contact was recorded as the loading area. Each begging contact was also classified based on its duration into three categories, namely momentary (<1 s), short (1–3 s), and long (>3 s); and the number of begging contacts was counted for each category.

Segmentation of comb in observation hive for determination of nectar loading area
Quantification of volume and concentration of crop contents
Forty-six nectar and 66 pollen foragers were captured in the corridor upon their departure from the hive to determine the volume and concentration of their crop content. They were killed by a cold spray (134a-QREI, Sunhayato Corp., Tokyo, Japan) immediately after capture and kept in a refrigerator at −20 °C until dissection. The crop was dissected out on a dissection dish, and the volume of its contents was measured to the nearest 0.1 µl using a 5- or 50-µl microsyringe (Hamilton, Reno, NE). There may be a small volume underestimate due to possible content remaining in crop tissue. To estimate sugar concentration (w/w), Brix of the crop content was measured to the nearest 0.5 % by a refractometer (low-volume refractometer Nectar 50 and 40–85, Eclipe, Bellingham & Stanley, Hampshire, UK). The Brix value was regarded to show approximated concentration of total sugar, because it represents the concentration of sugar based on reflective index by assuming pure sucrose solution, but nectar in the crop may also contain other sugars, such as glucose and fructose, as well as non-sugar substances. The Brix value is commonly used to estimate the concentration of sugars in nectar and the crop contents of insects (Corbet and Willmer 1980; Leonhardt et al. 2007; Pokorny et al. 2014; Tan et al. 2015).
Statistical analysis
We examined the effects of forager subtype (nectar/pollen forager), waggle duration, and loading area, on the volume and crop content concentration at departure using generalized linear models (GLMs) fitted by R (R Core Team 2015). A normal distribution was assumed as the distribution of the response variable. The significance of each effect was tested by likelihood ratio tests at the 5 % significance level.
GLMs were also used to examine the difference in the number of begging contacts between nectar and pollen foragers. Models were constructed by assuming a Poisson distribution.
The amount of sugar in the crop load was calculated from data on crop volume and concentration. The crop volume was first transformed to weight by multiplying by the density of the liquid, and was then multiplied by the sugar concentration (w/w) of the crop content. Coefficients of determination (R 2) were calculated for waggle duration vs. volume, vs. concentration, and vs. amount of sugar to assess the degree of adjustment for distance (waggle duration) in each variable.
The relationships between crop volume and concentration at departure were examined using Spearman’s rank correlation coefficient, because they did not show a linear relationship. The data were first analyzed separately for nectar and pollen foragers, and then pooled to examine the entire relationship across the two forager subtypes. The relationship between the total amount of dissolved sugar and crop load concentration was similarly analyzed by the same method.
To test whether foragers tend to receive nectar at a particular area on the comb, the proportions of bees for each loading area were compared with proportions expected by chance using a goodness-of-fit test. Whether nectar and pollen foragers tend to receive nectar at different areas was tested by a Chi-squared test of independence. The effect of loading area on the concentration of crop load at departure was tested by GLMs.
Results
Effects of forager subtype and food-source distance
Both the estimated concentration and volume of crop load at departure were significantly larger in pollen foragers than in nectar foragers (Fig. 2a, b; GLM: concentration, N = 36, 66; deviance = 7537.1, P < 0.001; volume, N = 46, 66, deviance = 3829.3, P < 0.001). We could not measure the concentration of crop content in ten nectar foragers because of the small volume (range 0.1–1.3 µl, median = 0.1 µl). The estimated sugar concentration and volume of crop content were positively correlated in nectar foragers (N = 36, r s = 0.41, P < 0.05), but not in pollen foragers (N = 66, r s = 0.04, P = 0.74). A correlation was found when data were pooled (N = 102, r s = 0.67, P < 0.001; Fig. 2c). The estimated sugar concentration also showed a positive correlation with the amount of sugar dissolved in crop contents, whether data were analyzed separately for nectar (N = 36, r s = 0.57, P < 0.001) and pollen foragers (N = 66, r s = 0.43, P < 0.001), or pooled (N = 102, r s = 0.80, P < 0.001; Fig. 2d).

Effect of forager subtype on concentration (a) and volume (b) of crop load carried from hive in Apis mellifera. The relationship between concentration and volume of crop load (c) and between concentration of crop load and amount of dissolved sugar (d). ***P < 0.001 by GLM. r s Spearman’s rank correlation coefficient. Numbers in bars indicate sample sizes. Error bars represent standard errors
The data in Fig. 2 were analyzed in relation to waggle duration. The volume of crop load at departure was significantly increased with waggle duration both in nectar foragers (N = 30, deviance = 46.2, P < 0.001) and pollen foragers (N = 55, deviance = 394.6, P < 0.05) (Fig. 3a). The effect of waggle duration and that of forager subtype on crop volume were significantly interacted (GLM with two explanatory variables “forager subtype” and “waggle duration”, N = 85, deviance = 109.0, P < 0.001), indicating that the slopes of the regression lines were significantly different in nectar and pollen foragers (Fig. 3a).

Effect of food-source distance on volume (a), concentration (b), and amount of sugar (c) in crop load carried from hive in Apis mellifera foragers. Gray line represents estimated sugar consumption for a round trip in (c) (see text for detail). Waggle duration was used as an indicator of food-source distance
The estimated concentration of crop load at departure also increased significantly with waggle durations in the two forager subtypes (Fig. 3b; GLM with two explanatory variables “forager subtype” and “waggle duration”; effect of waggle duration, N = 85, deviance = 973.1, P < 0.001). The models showed significantly higher concentration in pollen foragers than in nectar foragers even when the effect of waggle duration was considered (effect of forager subtype, deviance = 7083.7, P < 0.001). There was no significant interaction between the effect of forager subtype and that of waggle duration on the estimated concentration of crop load at departure (deviance = 85.4, P = 0.14), indicating that there was no significant difference between the slopes of their regression lines (Fig. 3b).
Positive correlations of volume and concentration with waggle duration resulted in a significant increase in the amount of sugar in the crop with waggle durations in both forager subtypes (Fig. 3c; GLM with two explanatory variables “forager subtype” and “waggle duration”; effect of waggle duration, N = 85, deviance = 412.6, P < 0.001). The slope of the regression line was significantly larger in pollen foragers than in nectar foragers (Fig. 3c; interaction between “forager subtype” and “waggle duration”, deviance = 138.5, P < 0.001). The R 2 values showed that waggle duration explains the variation in the amount of sugar better than in the crop volume or in the estimated concentration (Table 1).
Effect of loading area
During the observation of behavior, the observer lost sight temporarily of seven pollen foragers; therefore, they were excluded from the analysis. All remaining 46 nectar and 59 pollen foragers showed more than one begging contact with nest mates before leaving the hive, but except for two pollen foragers, none showed signs of taking honey from cells. The two exceptional pollen foragers inserted their heads into honey cells and remained in this position for 3.2 and 39.6 s, respectively. These bees were also excluded from the analysis, because this type of nectar loading seemed exceptional.
Both nectar and pollen foragers showed a tendency to receive nectar at specific (goodness-of-fit test: nectar forager, d.f. = 3, χ2 = 63.39, P < 0.001; pollen foragers, d.f. = 3, χ2 = 8.06, P < 0.05; Fig. 4a) but different areas (Chi-squared test for independence: d.f. = 4, χ2 = 39.85, P < 0.001; Fig. 4a). The longest begging contacts were most frequent at area 1 in nectar foragers, but at area 2 in pollen foragers (Figs. 1, 4a). However, the estimated concentration of crop load at departure did not depend on the loading area (GLM with two explanatory variables “loading area” and “forager subtype”: effect of loading area, deviance = 63.0, P = 0.90, N.S.; effect of forager subtype, deviance = 4172.6, P < 0.001; interaction, deviance = 22.8, P = 0.95, N.S.; and Fig. 4b).

a Proportion of foragers with respect to area with longest begging contact. ***P < 0.001. b Effect of loading area on concentration of crop load in nectar and pollen foragers departing hive. Numbers in the parentheses indicate sample sizes. Error bars represent standard errors. See Fig. 1 for segmentation of comb
Pollen foragers tended to show longer begging contacts than nectar foragers, reflecting their larger crop load at departure (Table 2). The total number of begging contacts was slightly, but significantly larger in pollen foragers than in nectar foragers. Both forager subtypes showed 2.2 times of momentary contact (<1 s) on average before leaving the hive.
Discussion
Our results show that departing foragers carry nectar at specific concentrations, depending on their foraging role (forager subtype) and food-source distance. These findings support the argument that foragers alter the concentration of nectar carried from the nest to adjust the sugar content in the crop (Tan et al. 2015). Given that more sugar is required by pollen foragers than nectar foragers, and by bees foraging further away than nearby, the present study suggests that bees use more-concentrated nectar when the need for sugar is high. The positive correlation between concentration and amount of sugar (Fig. 3d) also supports this conclusion.
Requirement of fuel and carriage cost
When foragers travel to further food source, larger amount of sugar is required as fuel. Because, the mass of crop load increases the energetic expenditure for flight (carriage cost) (Wolf et al. 1989), foragers should meet the increased requirement for fuel not only by increasing volume but also by increasing concentration of nectar. Our results showed that foragers adjust the amount of sugar by altering both crop volume and concentration. They appear to reduce the mass of the crop load, and hence, the mass-dependent carriage cost using concentrated nectar when they have to carry a large amount of sugar.
The present study also suggests that foragers adjust the amount of sugar for distance more precisely by altering both the volume and concentration of nectar, because the both variables showed a lower R 2 value with waggle duration than the amount of sugar. Foragers might adjust the amount of nectar according to concentration to obtain a certain amount of sugar (taking more nectar if the concentration is low, but taking less if it is high). In accordance with this argument, it is reported that bees can sense the quantity of sugar in consumed nectar based on volume and concentration (Varjú and Núñez 1991; Núñez and Giurfa 1996; Wainselboim and Farina 2000).
Adjustments in nectar foragers
In nectar foragers, the crop is used to carry collected nectar to the nest as well as for carrying fuel from the nest for flight. Any nectar for flight remaining in the crop when arriving at the nectar source reduces the capacity of the crop to carry collected nectar back to the nest. A similar situation is described in water collection (Visscher et al. 1996). However, if no nectar is collected from the nectar source, sufficient fuel must remain to return to the nest.
To examine whether bees can fly back to the nest without refueling (nectar collection), the sugar consumption for a round trip to/from a food source indicated by a waggle dance was estimated using the metabolic rate of flying workers as 100 mg/g/h (Gmeinbauer and Crailsheim 1993), body weight as 87.1 mg (mean, N = 13, Harano unpublished data), flight speed as 29 km/h (von Frisch and Lindauer 1955), and the relationship between the waggle-run duration and distance in the Japanese race of A. mellifera as Y = 1087X − 380, where Y is the distance from the nest to food source (m) and X is the waggle duration (s) (Sasaki 1994). As a result, the amount of sugar in the crop of nectar foragers departing the nest closely matches the estimated round-trip sugar consumption (Fig. 3c, gray line). The result suggests that nectar foragers adjust the amount of sugar in the crop to match the round-trip fuel need on leaving the nest. Some deviations from estimated round-trip sugar consumption could be explained by their responses to the reward variability of food source, because foragers increase fuel for variable food sources, but decrease for constant ones (De Marco and Farina 2003; Tan et al. 2015). Although some bees seemed to have insufficient sugars for a round trip in the crop, they can probably return to the nest using sugars in the midgut and hemolymph. Visscher et al. (1996) estimated that the sugar available outside the crop is between 1 and 2 mg.
Despite the reduced cost of carrying concentrated nectar, it is unclear why nectar foragers use relatively low-concentration nectar as fuel. Perhaps, the colony-level cost of concentrating nectar outweighs the benefit of using very concentrated nectar. Since in-nest bees concentrate nectar using time and energy investments (Park 1925), foragers not only consume sugars in nectar but also the colony’s time and energy investments when using concentrated nectar as fuel, increasing foraging cost at the colony level. It is also possible that they become less selective concerning the concentration of fuel, so that they return to the food source rapidly.
There is another possible explanation; perhaps, during begging contacts, nectar foragers receive both fuel as nectar and information about other food sources being exploited by other foragers. They may learn the characteristics of alternate food sources, such as taste, scent, and nectar concentration, as well as the profitability, and may use it in subsequent decision-making (Núñez 1970; Núñez and Giurfa 1996; Farina 1996; De Marco and Farina 2001, 2003). The nectar information is lost gradually with time after unloading to food receivers, because nectar is mixed with other nectar during concentration. To obtain meaningful information, nectar foragers must receive fresh nectar near the nest entrance. Thus, using freshly collected nectar as an information source might explain the relatively low sugar concentration of crop content in departing nectar foragers. Both energy and information seem important in determining fuel loading behavior.
Adjustments in pollen foragers
Pollen foragers carry nectar as pollen load glue upon leaving the nest. Given that an equal amount of sugar (on a dry-weight basis) is mixed with pollen to build pollen loads (Roulston et al. 2000), the amount of sugar required for building pollen loads is estimated as approx. 7 mg based on dry weights of pollen load as previously reported (14.6 mg, a mean dry weight for a pair of “M” and “L” pollen loads, N = 61, Harano and Sasaki 2015). This amount would partially explain the difference in the amount of sugar carried from the hive between pollen and nectar foragers.
As well as nectar for pollen load glue, they need to carry fuel for flight. Pollen foragers increased the total amount of sugars in the crop with food-source distance as did nectar foragers, but their increase rate of sugar to distance (slop of regression line; Fig. 3c) was more than four times larger than that of nectar foragers. The larger increase rate indicates increased requirement for sugar per unit distance, which may be at least partially due to the large nectar load for pollen glue, because the mass of the crop load increases energy expenditure per unit distance (Wolf et al. 1989). In addition, pollen foragers have higher metabolic rate than nectar foragers even when they are unloaded (Feuerbacher et al. 2003). Another possible factor increasing the need for fuel carried from the nest is the absence of nectar refueling at the food source by pollen foragers. Because of these factors, pollen foragers might need to carry more fuel per unit distance than nectar foragers.
Effects of foraging experience and food-source variability
The present study did not investigate the effect of foraging experience, but it may be an important factor in determining crop load at nest departure. Previous studies show that less-experienced foragers carried more fuel (Brandstetter et al. 1988; Harano et al. 2013) when leaving the nest. Foragers also respond to food-source variability or profitability by changing the frequency and duration of begging contact (Núñez 1970; Farina 1996; De Marco and Farina 2001, 2003). Future studies should investigate how and why these factors affect the crop content at nest departure.
Mechanism for selective use of nectar
What mechanism is used to load nectar at different concentrations by nectar and pollen foragers? We hypothesized that the available nectar concentration depends on comb area, and foragers receive nectar at different sugar concentrations at different areas. However, no effect of comb loading area on sugar concentration of crop content was detected. Similarly, although nectar and pollen foragers have different sucrose response thresholds (Pankiw et al. 2001), this does not explain the difference in concentration at departure, because pollen foragers are more responsive to dilute sugar solution than nectar foragers.
How foragers load nectar at specific concentrations remains unknown. One possible mechanism is selective nectar reception—foragers may reject nectar at low sugar concentrations when the need for sugar is high. We observed that departing foragers showed several momentary (<1 s) begging contacts before leaving the nest, as reported elsewhere (Farina and Wainselboim 2005). During these short contacts, bees were unlikely to receive nectar, but can probably sense the nectar concentration (Farina and Wainselboim 2005). Foragers might refuse offered nectar at these contacts because of inappropriate concentration, although such short contacts may play a role in gathering information about alternate food sources (Farina 1996; Núñez and Giurfa 1996; De Marco and Farina 2001).
In our observations, workers did not leave the nest with nectar at a very low sugar concentration (<20 %). It is logical to avoid using a very low-concentration nectar as fuel. However, Tan et al. (2015) reported considerably lower concentrations (approx. 4–6 %) in departing A. cerana foragers. Whether this is due to species difference or other reasons should be investigated.
In summary, departing A. mellifera foragers alter the concentration of crop load as well as volume to adjust the amount of sugar carried from the nest. Quantitative adjustment of crop content might be accomplished by monitoring the degree of crop expansion using stretch receptors on the crop wall (Brosch and Schneider 1985). However, selecting nectar at a specific concentration requires a different mechanism. This study suggests a complex mechanism involved in the adjustment of crop content in honeybees leaving the nest.
References
Beutler R (1950) Zeit und Raum im Leben der Sammelbiene. Naturwissenschaften 37:102–105
Beutler R (1951) Time and distance in the life of the foraging bee. Bee World 32:25–27
Blatt J, Roces F (2001) Haemolymph sugar levels in foraging honeybees (Apis mellifera carnica): dependence on metabolic rate and in vivo measurement of maximal rates of trehalose synthesis. J Exp Biol 204:2709–2716
Brandstetter M, Crailsheim K, Heran H (1988) Provisioning of food in the honeybee before foraging. BIONA Rep 6:129–148
Brosch U, Schneider L (1985) Fine structure and innervation of the honey stomach (crop) of the honeybee, Apis mellifera L. (Hymenoptera: Apidae). Int J Insect Morphol Embryol 14:335–345
Corbet SA, Willmer P (1980) Pollination of the yellow passionfruit: nectar, pollen and carpenter bees. J Agric Sci 95:655–666
Crailsheim K (1988) Regulation of food passage in the intestine of the honeybee (Apis mellifera L.). J Insect Physiol 34:85–90
De Marco R, Farina W (2001) Changes in food source profitability affect the trophallactic and dance behavior of forager honeybees (Apis mellifera L.). Behav Ecol Sociobiol 50:441–449
De Marco R, Farina W (2003) Trophallaxis in forager honeybees (Apis mellifera): resource uncertainty enhances begging contacts? J Comp Physiol A 189:125–134
Farina WM (1996) Food-exchange by foragers in the hive–a means of communication among honey bees? Behav Ecol Sociobiol 38:59–64
Farina WM, Wainselboim AJ (2005) Trophallaxis within the dancing context: a behavioral and thermographic analysis in honeybees (Apis mellifera). Apidologie 36:43–47
Feuerbacher E, Fewell JH, Roberts SP, Smith EF, Harrison JF (2003) Effects of load type (pollen or nectar) and load mass on hovering metabolic rate and mechanical power output in the honey bee Apis mellifera. J Exp Biol 206:1855–1865
Free J (1960) The behaviour of honeybees visiting flowers of fruit trees. J Anim Ecol 29:385–395
Gmeinbauer R, Crailsheim K (1993) Glucose utilization during flight of honeybee (Apis mellifera) workers, drones and queens. J Insect Physiol 39:959–967
Goyret J, Farina WM (2005) Trophallactic chains in honeybees: a quantitative approach of the nectar circulation amongst workers. Apidologie 36:595–600
Harano K, Sasaki M (2015) Adjustment of honey load by honeybee pollen foragers departing from the hive: the effect of pollen load size. Insect Soc 62:497–505
Harano K, Mitsuhata-Asai A, Konishi T, Suzuki T, Sasaki M (2013) Honeybee foragers adjust crop contents before leaving the hive. Behav Ecol Sociobiol 67:1169–1178
Harano K, Mitsuhata-Asai A, Sasaki M (2014) Honey loading for pollen collection: regulation of crop content in honeybee pollen foragers on leaving hive. Naturwissenschaften 101:595–598
Hendrichs J, Cooley S, Prokopy R (1992) Post-feeding bubbling behaviour in fluid-feeding Diptera: concentration of crop contents by oral evaporation of excess water. Physiol Entomol 17:153–161
Hodges D (1952) The pollen loads of the honeybee: a guide to their identification by colour and form. Bee Research Association, London
Japan Meteorological Agency (2016) Japan Meteorological Agency website http://www.jma.go.jp/jma/index.html. Accessed 8 March 2016
Leonhardt SD, Dworschak K, Eltz T, Blüthgen N (2007) Foraging loads of stingless bees and utilisation of stored nectar for pollen harvesting. Apidologie 38:125–135
Nicolson SW, Human H (2008) Bees get a head start on honey production. Biol Lett 4:299–301
Núñez J (1970) The relationship between sugar flow and foraging and recruiting behaviour of honey bees (Apis mellifera L.). Anim Behav 18:527–538
Núñez JA, Giurfa M (1996) Motivation and regulation of honey bee foraging. Bee World 77:182–196
Page RE Jr (2013) The spirit of the hive. Harvard University Press, Cambridge
Pankiw T, Waddington KD, Page RE (2001) Modulation of sucrose response thresholds in honey bees (Apis mellifera L.): influence of genotype, feeding, and foraging experience. J Comp Physiol A 187:293–301
Panzenböck U, Crailsheim K (1997) Glycogen in honeybee queens, workers and drones (Apis mellifera carnica Pollm.). J Insect Physiol 43:155–165
Park W (1925) The storing and ripening of honey by honeybees. J Econ Entomol 18:405–410
Park OW (1946) Activities of honeybees. In: Grout RA (ed) The hive and the honeybee. Hamilton, Illinois, Dadant & Sons, pp 125–206
Parker RL (1926) The collection and utilization of pollen by the honeybee. Mem Cornell Univ Agric Exp Stn 98:1–55
Pokorny T, Lunau K, Eltz T (2014) Raising the sugar content–orchid bees overcome the constraints of suction feeding through manipulation of nectar and pollen provisions. PLoS ONE 9:e113823
Roulston TH, Cane JH, Buchmann SL (2000) What governs protein content of pollen: pollinator preferences, pollen-pistil interactions, or phylogeny? Ecol Monogr 70:617–643
R Core Team (2015) R: a language and environment for statistical computing. R foundation for statistical computing, Vienna, Austria, http://www.R-project.org/
Sasaki M (1994) Youhou no Kagaku. Science House, Tokyo (in Japanese)
Schmidt-Nielsen K (1997) Animal physiology: adaptation and environment. Cambridge University Press, Cambridge
Seeley TD (1986) Social foraging by honeybees: how colonies allocate foragers among patches of flowers. Behav Ecol Sociobiol 19:343–354
Seeley TD (1989) Social foraging in honey bees: how nectar foragers assess their colony’s nutritional status. Behav Ecol Sociobiol 24:181–199
Seeley TD (1995) The wisdom of the hive. Harvard University Press, Cambridge
Seeley T, Morse R (1976) The nest of the honey bee (Apis mellifera L.). Insect Soc 23:495–512
Tan K, Latty T, Dong S, Liu X, Wang C, Oldroyd BP (2015) Individual honey bee (Apis cerana) foragers adjust their fuel load to match variability in forage reward. Sci Rep 5:16418
Varjú D, Núñez J (1991) What do foraging honeybees optimize? J Comp Physiol 169:729–736
Visscher PK, Crailsheim K, Sherman G (1996) How do honey bees (Apis mellifera) fuel their water foraging flights? J Insect Physiol 42:1089–1094
von Frisch K (1967) The dance language and orientation of bees. Harvard University Press, Cambridge
von Frisch K, Lindauer M (1955) Über die Fluggeschwindigkeit der Bienen und über ihre Richtungsweisung bei Seitenwind. Naturwissenschaften 42:377–385
Wainselboim AJ, Farina WM (2000) Trophallaxis in the honeybee Apis mellifera (L.): the interaction between flow of solution and sucrose concentration of the exploited food sources. Anim Behav 59:1177–1185
Wittmann D, Scholz E (1989) Nectar dehydration by male carpenter bees as preparation for mating flights. Behav Ecol Sociobiol 25:387–391
Wolf TJ, Schmid-Hempel P, Ellington CP, Stevenson RD (1989) Physiological correlates of foraging efforts in honey-bees: oxygen consumption and nectar load. Funct Ecol 3:417–424
Acknowledgments
We thank Mr. M. Hayashi for apiary assistance. We are also grateful to Drs. M. Sasaki, T. Sasaki, and K. Sasaki for stimulating discussion. Two anonymous reviewers improved the manuscript greatly. This study was partly supported by a Kakenhi grant [26850219] to K.H. by the Ministry of Education, Culture, Sport, Science and Technology, Japan.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Harano, Ki., Nakamura, J. Nectar loads as fuel for collecting nectar and pollen in honeybees: adjustment by sugar concentration. J Comp Physiol A 202, 435–443 (2016). https://doi.org/10.1007/s00359-016-1088-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00359-016-1088-x