
Abstract
Purpose
BAG3 is overexpressed in several malignancies and mediates a non-canonical, selective form of (macro)autophagy. By stabilizing pro-survival Bcl-2 proteins in complex with HSP70, BAG3 can also exert an apoptosis-antagonizing function. ABT-737 is a high affinity Bcl-2 inhibitor that fails to target Mcl-1. This failure may confer resistance in various cancers.
Methods
Urothelial cancer cells were treated with the BH3 mimetics ABT-737 and (−)-gossypol, a pan-Bcl-2 inhibitor which inhibits also Mcl-1. To clarify the importance of the core autophagy regulator ATG5 and BAG3 in ABT-737 treatment, cell lines carrying a stable lentiviral knockdown of ATG5 and BAG3 were created. The synergistic effect of ABT-737 and pharmaceutical inhibition of BAG3 with the HSF1 inhibitor KRIBB11 or sorafenib was also evaluated. Total cell death and apoptosis were quantified by FACS analysis of propidium iodide, annexin. Target protein analysis was conducted by Western blotting.
Results
Knockdown of BAG3 significantly downregulated Mcl-1 protein levels and sensitized urothelial cancer cells to apoptotic cell death induced by ABT-737, while inhibition of bulk autophagy through depletion of ATG5 had no discernible effect on cell death. Similar to knockdown of BAG3, pharmacological targeting of the BAG3/Mcl-1 pathway with KRIBB11 was capable to sensitize both cell lines to treatment with ABT-737.
Conclusion
Our results show that BAG3, but not bulk autophagy has a major role in the response of bladder cancer cells to BH3 mimetics. They also suggest that BAG3 is a suitable target for combined therapies aimed at synergistically inducing apoptosis in bladder cancer.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Bladder cancer is the second most common malignancy of the urinary tract. The prognosis of patients with metastatic bladder cancer is extremely poor in spite of various chemotherapeutic options. Malignant progression is associated with alterations in numerous cellular processes, including those that lead to apoptosis, which is dysregulated in numerous cancers [1]. This process is regulated by several pro- and anti-apoptotic genes, among them are Bcl-2 and its homologues. Consistently, many investigators have demonstrated that overexpression of Bcl-2 correlates with the progression and chemotherapeutic response of various human malignancies, including bladder cancer [2].
Drug development attempts to disrupt the interaction between pro-apoptotic and anti-apoptotic proteins of the Bcl-2 family yielded the Bcl-2 homology domain 3 (BH3) mimetic class of drugs [3]. The BH3-mimetic ABT-737 is a small molecule that binds with high affinity to Bcl-2, Bcl-xL, and Bcl-w, but not to Mcl-1 [4]. This failure to target Mcl-1 may confer ABT-737 resistance in various cancers with higher expression of pro-survival Bcl-2 family members [5]. In contrast (−)-gossypol, a natural product from cottonseed, has been identified as a small molecule pan-Bcl-2 inhibitor also targeting Mcl-1 [6]. By displacing the pro-autophagic BH3-only protein Beclin-1 from the pro-survival Bcl-2 family members, BH3 mimetics can also activate autophagy [4].
Autophagy itself is a catabolic process, in which portions of the cytoplasm or complete organelles are delivered to the autophagosomal–lysosomal pathway for bulk degradation [7]. This type of autophagy is termed macroautophagy. Distinct types of autophagy are microautophagy and chaperone-mediated autophagy (CMA) [8]. In regard to cell death/survival decisions, the role of macroautophagy (hereafter called autophagy) is highly contextual. In general, autophagy acts as a pro-survival stress response; there is also evidence that overactivation of autophagy can act as an alternative cell death pathway [9]. The execution of autophagy involves multiple conserved autophagy-related genes (ATG). Among these gene products, ATG5 is critically involved in the early stages of autophagosome formation [10].
The HSP70 co-chaperone Bcl-2-associated athanogene 3 (BAG3) mediates a selective form of autophagy for the degradation of damaged client proteins known to accumulate under conditions of cellular stress, thereby consolidating cells by recruitment of the macroautophagy pathway [11]. Furthermore, BAG3 was shown to sustain cell survival and underlie resistance to chemotherapy in several human neoplasms [12]. Overexpression of BAG3 has been observed in different types of tumors and may stabilize pro-survival Bcl-2 family members via preventing their degradation in the proteasomal pathway, thereby supporting their anti-apoptotic activity [13].
Here, we investigated the role of autophagy-regulating proteins in bladder cancer cells to treatment with ABT-737. Our data show that depletion of BAG3 through RNA silencing augmented ABT-737-mediated cell death, whereas inhibition of bulk autophagy via knockdown of ATG5 had no significant effect on cell survival. Also, the pro-apoptotic effect of ABT-737 can be significantly increased by combination with drugs that target upstream signaling mechanisms to inhibit the expression of BAG3 and Mcl-1.
Materials and methods
Materials
(−)-Gossypol (>98 % purity) was acquired from Tocris (Bristol, UK). Z–Val-Ala-DL-Asp(OMe)-fluoromethylketone (z-VAD) was purchased from Bachem (Weil am Rhein, Germany). KRIBB11 was from Calbiochem (Darmstadt, Germany). Sorafenib and ABT-737 were purchased from Santa Cruz (Heidelberg, Germany). Staurosporine (STS) was obtained from Alexis Biochemicals (San Diego, CA, USA). All other chemicals were used in analytic grade purity from Sigma-Aldrich (Munich, Germany).
Cell lines and culture
RT-112, 5637 (ATCC/LGC Promochem GmbH, Wesel, Germany), and TCC-SUP (DSMZ, Braunschweig, Germany) bladder carcinoma cells were grown and subcultured in IMDM, 10 % fetal calf serum, 2 % glutamin, and 1 % penicillin/streptomycin (all: Gibco/Invitrogen, Karlsruhe, Germany) cultivated in a humidified incubator at 37 °C and 5 % CO2. Human urothelial epithelial cells (HUEPC) were obtained from Provitro GmbH (Berlin, Germany). Cells were grown in Provitro urothelial growth medium, which is a sterile liquid medium. The Provitro essential supplement kit components were added to the basal growth medium. HUEPC have to be cultivated in a humidified incubator at 37 °C and 5 % CO2.
Lentiviral transduction
Lentiviral vector stocks specific for ATG5 (SHVRS-NM_004849; Sigma-Aldrich) and BAG3 (SHCLNV-NM_004281; Sigma-Aldrich) were used for transduction of bladder cancer cells. The target sets included five sequences for different small hairpins. The pLKO.1-puro control transduction particles (SHC001 V) did not contain a hairpin insert and were used as a negative control. Transduction and selection of stable cells were performed as described previously [9].
SDS-PAGE and Western blotting
Western blots of whole-cell lysates were prepared as described previously [9]. The resulting blots were probed with a mouse monoclonal anti-Bcl-2 antibody diluted 1:1000 (Santa Cruz Biotechnology), a rabbit polyclonal anti-Bcl-xL antibody diluted 1:1000 (Cell Signaling, Leiden, The Netherlands), a rabbit polyclonal anti-Mcl-1 antibody diluted 1:1000 (Cell Signaling), a rabbit polyclonal anti-Bcl-w diluted 1:1000 (Cell Signaling), a rabbit polyclonal anti-ATG5 antibody (Cell Signaling) diluted 1:1000, an anti-BAG3 antibody diluted 1:3000 von (Biozol/Abnova, Eching, Germany), a mouse monoclonal anti-LC3 antibody (Sigma-Aldrich) diluted 1:1000, and a mouse monoclonal anti-glyceraldehyde-3-phosphate dehydrogenase (GAPDH) antibody (Calbiochem) diluted 1:10,000.
Flow cytometry
Cell death was detected by flow cytometry after annexin V-fluos/propidium iodide (PI) double staining [Roche Applied Science (Penzberg, Germany); Sigma-Aldrich] as described before [9]. Flow cytometric analyses were done on a FACS-Canto II (BD Biosciences, Heidelberg, Germany) followed by analysis using FACSDiva software (BD Biosciences).
Statistics
Data are given as mean ± SEM. For statistical comparison, one-way ANOVA followed by Tukey’s test was used using SPSS software (SPSS GmbH Software). P values <0.05 were considered to be statistically significant.
Results
ABT-737 moderately induces a caspase-dependent apoptotic type of cell death in bladder cancer cells
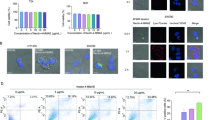
Previous studies have suggested that the mode and extent of cell death induced by BH3 mimetics depend on the level of Bcl-2 [9, 14]. We first analyzed the expression levels of pro-survival Bcl-2 family members in a series of bladder cancer cell lines by Western blot (Fig. 1a). Cell line TCC-SUP has higher levels of Bcl-2 and Mcl-1 than 5637 and RT112, but exhibits lower expression of Bcl-w. We next compared their responses to ABT-737 treatment, which has been described to be capable of inhibiting Bcl-2, Bcl-xL, and Bcl-w, but not Mcl-1 [4], by FACS analysis of annexin V binding and propidium iodide uptake to determine both early apoptosis and total cell death (Fig. 1b). The extent of ABT-737-induced cell death was correlated with the expression of Bcl-2 proteins: 5637 cells with the lowest level of Bcl-2 and Mcl-1 expression displayed the highest sensitivity to treatment with 10 µM ABT-737 for 48 h with ~40 % total cell death followed by RT112 with ~26 % and TCC-SUP with ~20 % total cell death. Pre-treatment with z-VAD, a cell-permeant pan-caspase inhibitor that binds irreversibly to the catalytic site of caspase proteases, significantly decreased cell death in all cell lines compared to ABT-737 treatment alone, indicating that ABT-737 had triggered apoptosis. In comparison with ABT-737, the natural BH3 mimetic (−)-gossypol which has been described as a pan-Bcl-2 inhibitor targeting Bcl-2, Bcl-xL, Bcl-w, and Mcl-1 [4, 9] induced higher levels of total cell death in 5637 with ~73 %, in RT112 with ~42 % and TCC-SUP with ~35 % total cell death. As seen in the case of ABT-737 pre-treatment, treatment with z-VAD significantly decreased cell death in all cell lines compared to (−)-gossypol treatment alone.

Western blot analysis of the expression of pro-survival Bcl-2 family members in 5637, RT112, and TCC-SUP cells. GAPDH served as a loading control (a). Cells were pre-treated with z-VAD (100 µM) for 1 h followed by treatment with 10 µM ABT-737 (b) and 15 µM (−)-gossypol (c) for 48 h. Total cell death (Q1, 2, 4) and apoptotic cell death (Q4) was quantified by flow cytometric analysis. *P < 0.05 versus DMSO. # P < 0.05 versus ±zVAD. Representative data from the experiment in 5637, RT112, and TCC-SUP shown in (b) and (c) exhibited as FACS dot plot profiles (d, e)
Knockdown of BAG3, but not ATG5 sensitizes bladder cancer cells to cell death induced by ABT-737
BAG3 overexpression has been previously reported in several malignancies [13]. BAG3 protein levels in our panel of bladder cancer cells in comparison with non-transformed HUEPC were determined by Western blot (Fig. 2a). HUEPC hardly showed any BAG3 expression, while RT112 cells showed the highest BAG3 protein level among the investigated bladder cancer cell lines. All other cell lines had a nearly even level of BAG3. In contrast to BAG3, ATG5 is a key player involved in canonical bulk, i.e., non-selective macroautophagy and ATG5-deficient cells exhibit a significantly diminished formation of autophagic vesicles [7]. Western blot analysis revealed that ATG5 displays a robust expression at an even level in the four evaluated cell lines (Fig. 2a).

Western blot analysis of the expression of BAG3 and ATG5 in human urothelial epithelial cells (HUEPC), 5637, RT112, and TCC-SUP bladder cancer cells. GAPDH served as a loading control (a). Stable knockdown of BAG3 in 5637 and RT112 cells as shown by Western blot analysis with a BAG3 antibody. Whole-cell lysates of 5637 and RT112 control cells transfected with empty vector (∅-vec), and BAG3-KD cells were probed and analyzed with an antibody against BAG3. Western blot analysis of the expression of Mcl-1 in the 5637 BAG3-KD and RT112 BAG3-KD cells. GAPDH served as a loading control (b). Stable knockdown of ATG5 in 5637 and RT112 cells as shown by Western blot analysis with an ATG5 antibody. Whole-cell lysates of 5637 and RT112 control cells transfected with empty vector (∅-vec), and ATG5-KD cells were probed and analyzed with an antibody against ATG5. Western blot analysis of the expression of Mcl-1 in the 5637 ATG5-KD and RT112 ATG5-KD cells. GAPDH served as a loading control (c). Western blot analysis of the expression of the Bcl-2 proteins, LC3-I, and LC3-II in 5637 ∅-vec, BAG3-KD, and ATG5-KD cells. The cells were treated with 10 µM ABT-737 and 15 µM (−)-gossypol for 48 h. GAPDH served as a loading control (d)
To investigate and compare the potential role of BAG3 and bulk autophagy in modulating cell death induced by BH3 mimetics, lentiviral knockdowns of BAG3 and ATG5 were established in cell lines RT112 and 5637 (Fig. 2b, c). Empty vector-transduced cells were used as controls. It was previously shown that BAG3 stabilizes Mcl-1 and thus promotes its anti-apoptotic effects [13]. In line with these findings, 5637 BAG3-KD cells exhibited barely detectable levels of Mcl-1 (Fig. 2b), whereas the Mcl-1 level remained unaltered in the ATG5-KD cells (Fig. 2c). As expected, analysis of LC3 conversion indicated a decreased amount of LC3-II after treatment with the potent autophagy inducer (−)-gossypol in the ATG5-KD cells, but not in BAG3-KD cells (Fig. 2d). On a closer look, we also observed that treatment with (−)-gossypol, but not with ABT-737 leads to markedly decreased protein levels of Mcl-1, Bcl-2, and Bcl-w, and to a lesser extent of Bcl-xL in all cell lines (Fig. 2d).
In subsequent experiments, an annexin V/PI FACS analysis was performed to detect early apoptotic cells and overall cell death after treatment with ABT-737 and (−)-gossypol. Staurosporine was used as a positive control for induction of apoptotic cell death (Fig. 3a). In 5637 cells after 48 h ABT-737 and (−)-gossypol treatment, the annexin V-positive/PI-negative fraction representing the early apoptotic cells and overall cell death was significantly higher in BAG3-KD cells compared to the controls. This cell death-enhancing effect of the BAG3 knockdown was particularly evident after treatment with ABT-737. In contrast, neither the annexin V-positive/PI-negative fraction nor overall cell death was significantly altered in ATG5-KD cells compared to controls (Fig. 3b, c). When treating 5637 BAG3-KD cells with ABT–737, the total cell death level (~80 %) exceeded that of (−)-gossypol treatment in 5637 cells (~74 %). This phenomenon could also be detected in the RT112 triplet, where the early apoptotic cells and overall cell death were significantly higher in BAG3-KD cells compared to the controls and ATG5-KD cells. The fact that 5637 BAG3-KD cells are slightly more sensitive to (−)-gossypol than 5637 cells can be explained by the fact that pro-survival Bcl-2 proteins are not the only anti-apoptotic targets of BAG3 [15]. In comparison with ABT-737, the potentiating effect of the BAG3 knockdown on (−)-gossypol-induced cell death is minor, clearly demonstrating that in bladder cancer cells, the apoptosis-sensitizing effects of the BAG3 knockdown are primarily mediated through suppression of pro-survival Bcl-2 proteins.

Quantification of total cell death and apoptotic cell death by FACS cytometry (annexin V/PI). Empty vector-transfected 5637 (∅-vec) and 5637 BAG3-KD/ATG5-KD cells were treated with 3 µM STS (a) for 6 h, 15 µM (−)-gossypol (b) and 10 µM ABT-737 (c) for 48 h. Empty vector-transfected RT112 (∅-vec) and RT112 BAG3-KD/ATG5-KD cells were also treated with 10 µM ABT-737 for 48 h (d). *P < 0.05 versus DMSO. # P < 0.05 compared with controls (∅-vec) with the same respective treatment. Representative data from the experiment (b) and (c) in 5637 ∅-vec, BAG3-KD, and ATG5-KD exhibited as FACS dot plot profiles (e)
Targeting the HSF1/Mcl-1 pathway sensitizes bladder cancer cells to treatment with ABT-737
Our results obtained with the knockdown models so far indicated that BAG3 may be crucially implicated in supporting the anti-apoptotic function of Mcl-1 in bladder cancer cells. In a pharmacological approach, we aimed at further investigating this hypothesis by modulating the expression of BAG3 (and its interactor HSP70) via application of the HSF1 inhibitor KRIBB11 [16] and the multikinase inhibitor sorafenib in the parental cells. Indeed, Western blot analysis after KRIBB11 treatment of 5637 cells showed decreased levels of HSF1, HSP70, BAG3, and Mcl-1 as outlined in Fig. 4a. FACS analysis confirmed that ABT-737 alone had only moderate effects on cell death of 5637 cells. Combined treatment with ABT-737 and KRIBB11, however, evoked a significant increase in early apoptosis and overall cell death compared to single treatment with ABT-737 and KRIBB11. The combination of ABT-737 with sorafenib, a drug which is known to downregulate Mcl-1 [17], also induced a significant increase in total cell death, albeit to a lesser degree than KRIBB11 (Fig. 4c, d). Western blot analysis showed strongly reduced BAG3 levels after the ABT-737/sorafenib treatment, while the ABT-737/KRIBB11 combination evoked total depletion of BAG3 (Fig. 4b). These results demonstrate that Mcl-1-dependent resistance to ABT-737 can be overcome through combination with drugs targeting expression/stability of Mcl-1 and BAG3.

Western blot analysis of the expression of HSF1, HSP70, BAG3, and Mcl-1 in 5637 bladder cancer cells after treatment with 10 µM KRIBB11 for 12 h. GAPDH served as a loading control (a). Western blot analysis of the expression of Mcl-1 and BAG3 in 5637 cells. The cells were treated with 10 µM ABT-737, 5 µM sorafenib, and 10 µM KRIBB11. GAPDH served as a loading control (b). Quantification of total cell death (Q1, 2, 4) and apoptotic cell death (Q4) (c) by FACS cytometry (annexin V/PI). 5637 cells were treated with 10 µM ABT-737, 5 µM sorafenib, 10 µM KRIBB11, and the combination of ABT-737 with KRIBB11 and sorafenib for 12 h. *P < 0.05 versus DMSO. # P < 0.05 compared with ABT-737 single treatment. Representative data from the experiment shown in (c) exhibited as FACS dot plot profiles (d)
Discussion
Apoptosis is characterized by a cascade of molecular events that are initiated by distinct upstream signals and culminate in the activation of effector caspases, the major executors of apoptotic cell death [18]. Proteins of the Bcl-2 family are key regulators of apoptosis, but also represent important inhibitors or inducers of autophagy [19]. Bcl-2 family proteins play a pivotal role in the intrinsic (mitochondrial) pathway of apoptosis, which triggers increased mitochondrial permeability and the release of pro-apoptotic molecules into the cytoplasm.
The Bcl-2 family consists of anti-apoptotic members such as Bcl-2, Bcl-xL, Bcl-w, and Mcl-1 which are highly overexpressed in many malignancies, e.g., bladder cancer [2], the pro-apoptotic molecules and pore-forming multidomain proteins Bax and Bak, as well as the BH3-only proteins which couple diverse stress signals to the intrinsic apoptosis pathway. Upon induction of the intrinsic pathway by BH3-only proteins, Bax and Bak are inserted into the outer mitochondrial membrane, thus promoting outer mitochondrial membrane permeabilisation (MOMP) and cytochrome c release [20]. In contrast, the anti-apoptotic Bcl-2 family members block apoptosis by preventing Bax and Bak activation and MOMP [21].
Autophagy is a regulated degradation pathway for the removal of intracellular organelles and long-lived proteins, and depending on the respective cellular context and its extent/duration, autophagy can have either pro-survival or pro-death functions [9]. The anti-apoptotic members of the Bcl-2 family also function as anti-autophagic regulators via their inhibitory interaction with the core autophagy factor and non-apoptotic BH3-only protein Beclin 1, which participates in autophagosome formation [22].
BH3 mimetics can induce apoptotic or non-apoptotic cell death, and this apparent dichotomy appears to be correlated with the expression levels of pro-survival Bcl-2 family members [14]. The role of autophagy in modulation of BH3 mimetic-induced cell death is currently controversial. Previous studies have shown a cell type- and context-dependent autophagy after treatment with ABT-737 and (−)-gossypol, which play either a death-promoting [9] or cytoprotective role [23].
In this study, we wanted to identify the specific roles of BAG3 and bulk autophagy in the response of bladder cancer cells to ABT-737. To this end, we established stable knockdowns of BAG3, a protein that mediates a non-canonical, selective form of autophagy to limit proteotoxicity [24], but also acts in an anti-apoptotic manner through stabilizing anti-apoptotic Bcl-2 proteins such as Bcl-xL, and Mcl-1 [13]. In a comparative approach, we also blocked the non-selective macroautophagy pathway by knockdown of ATG5.
Overexpression of anti-apoptotic Bcl-2 proteins is commonly observed in a variety of cancers and associated with resistance to conventional chemotherapeutic drugs. Our expression profiling and cell death/survival assays showed that ABT-737 and (−)-gossypol induce an apoptotic cell death in RT112 and 5637 bladder cancer cells which is correlated with the expression levels of Bcl-2. The pan-Bcl-2 inhibitor (−)-gossypol induced higher levels of cell death in comparison with ABT-737, likely due to the failure of ABT-737 to target Mcl-1 [9]. In our experimental setting, the inhibition of bulk autophagy via ATG5 knockdown evoked no significant changes in early apoptosis and total cell death after treatment with the apoptosis inducers staurosporine, ABT-737, and (−)-gossypol in bladder cancer cells.
Similar to Bcl-2, Mcl-1 overexpression has been observed in a variety of human tumors such as colorectal cancer [25]. The co-chaperone BAG3 binds to Mcl-1 preventing its proteasomal degradation and thus promoting the anti-apoptotic effect [13]. In line with our findings, a substantial expression of BAG3 has been reported in several types of human cancers, such as glioblastoma, pancreatic carcinoma, leukemia, and thyroid carcinoma, compared with very low basal levels of BAG3 in non-malignant cells [26]. In our experiments, knockdown of BAG3 significantly decreased the protein levels of Mcl-1 and potently increased apoptotic cell death after treatment with ABT-737 and to a minor extent after treatment with (−)-gossypol.
BAG3 was shown to interfere with the HSP70-mediated delivery of anti-apoptotic proteins such as Mcl-1, Bcl-2, and Bcl-xL to the proteasomal pathway by competing with BAG1, a protein that also functions as co-chaperone of HSP70 [26]. In the case of Mcl-1, the BAG3 interaction with Mcl-1 and HSP70 may prevent or destabilize the binding of Mcl-1 to HSP70, reducing the delivery of Mcl-1 to the proteasome and supporting the anti-apoptotic activity of Mcl-1 [13]. In line with this hypothesis and previously published findings [27], silencing of Mcl-1 was required to efficiently trigger ABT-737-dependent cell death, suggesting that stabilization of Mcl-1 is a key mechanism of drug resistance induced by BAG3. Under conditions of cellular stress as induced by ABT-737 and (−)-gossypol, damaged proteins are known to accumulate, a condition known to trigger BAG3-dependent autophagy of client proteins [11]. Therefore, it is likely that BAG3-dependent recruitment of the macroautophagy pathway and consolidation of cells via decreased proteotoxicity may additionally protect them against apoptosis.
Consistent with our data obtained here, single-agent treatment with ABT-737 displayed only moderate cytotoxic effects in different types of cancer with high Mcl-1 expression, including glioblastoma [9]. One promising approach to enhancing the cellular sensitivity to ABT-737 is the (pharmacological) neutralization of BAG3 to increase Mcl-1 degradation via the proteasome [13]. In order to overcome this Mcl-1-driven resistance, we pharmacologically targeted the BAG3/HSP70/Mcl-1 signaling axis by modulating upstream activation of Mcl-1 with the small molecule inhibitor KRIBB11. KRIBB11 was found to selectively inhibit the transcriptional activity of HSF1 [28], which is required for expression of both BAG3 [29] and its interactor HSP70 [16]. Indeed, we could observe significant synergistic effects on cell death in the combination therapy with ABT-737 and KRIBB11. Similar, albeit weaker enhancing effects were observed after treatment with sorafenib, a multikinase inhibitor, whose pro-apoptotic effect was previously correlated with downregulation of Mcl-1 expression in multiple tumors [30]. According to the results of our preclinical study, we propose that a combination therapy of the orally applicable ABT-737 derivative ABT-263 with the multikinase inhibitor sorafenib represents a promising approach for the treatment of bladder cancer which warrants further investigation in the preclinical and clinical settings.
Conclusion
Here, we demonstrate that BAG3 plays a major role for the cell death resistance of bladder cancer cells and their response to BH3 mimetics. Our data suggest that BAG3 is a suitable target for combined therapies aimed at synergistically inducing apoptosis in advanced bladder cancer and may aid the design of new strategies to overcome the resistance to current bladder cancer therapy.
Abbreviations
- CMA:
-
Chaperone-mediated autophagy
- ATG:
-
Autophagy-related genes
- z-VAD:
-
Z-Val-Ala-DL-Asp(OMe)-fluoromethylketone
- STS:
-
Staurosporine
- PI:
-
Propidium iodide
References
Hanahan D, Weinberg RA (2011) Hallmarks of cancer: the next generation. Cell 144(5):646–674. doi:10.1016/j.cell.2011.02.013
Karam JA, Lotan Y, Karakiewicz PI, Ashfaq R, Sagalowsky AI, Roehrborn CG, Shariat SF (2007) Use of combined apoptosis biomarkers for prediction of bladder cancer recurrence and mortality after radical cystectomy. Lancet Oncol 8(2):128–136. doi:10.1016/S1470-2045(07)70002-5
Ni Chonghaile T, Letai A (2008) Mimicking the BH3 domain to kill cancer cells. Oncogene 27(Suppl 1):S149–S157. doi:10.1038/onc.2009.52
Lessene G, Czabotar PE, Colman PM (2008) BCL-2 family antagonists for cancer therapy. Nat Rev Drug Discov 7(12):989–1000. doi:10.1038/nrd2658
Oltersdorf T, Elmore SW, Shoemaker AR, Armstrong RC, Augeri DJ, Belli BA, Bruncko M, Deckwerth TL, Dinges J, Hajduk PJ, Joseph MK, Kitada S, Korsmeyer SJ, Kunzer AR, Letai A, Li C, Mitten MJ, Nettesheim DG, Ng S, Nimmer PM, O’Connor JM, Oleksijew A, Petros AM, Reed JC, Shen W, Tahir SK, Thompson CB, Tomaselli KJ, Wang B, Wendt MD, Zhang H, Fesik SW, Rosenberg SH (2005) An inhibitor of Bcl-2 family proteins induces regression of solid tumours. Nature 435(7042):677–681. doi:10.1038/nature03579
Kang MH, Reynolds CP (2009) Bcl-2 inhibitors: targeting mitochondrial apoptotic pathways in cancer therapy. Clin Cancer Res 15(4):1126–1132. doi:10.1158/1078-0432.CCR-08-0144
Mizushima N, Levine B, Cuervo AM, Klionsky DJ (2008) Autophagy fights disease through cellular self-digestion. Nature 451(7182):1069–1075. doi:10.1038/nature06639
Dice JF (2007) Chaperone-mediated autophagy. Autophagy 3(4):295–299
Voss V, Senft C, Lang V, Ronellenfitsch MW, Steinbach JP, Seifert V, Kogel D (2010) The pan-Bcl-2 inhibitor (−)-gossypol triggers autophagic cell death in malignant glioma. Mol Cancer Res 8(7):1002–1016. doi:10.1158/1541-7786.MCR-09-0562
Mizushima N, Noda T, Yoshimori T, Tanaka Y, Ishii T, George MD, Klionsky DJ, Ohsumi M, Ohsumi Y (1998) A protein conjugation system essential for autophagy. Nature 395(6700):395–398. doi:10.1038/26506
Gamerdinger M, Hajieva P, Kaya AM, Wolfrum U, Hartl FU, Behl C (2009) Protein quality control during aging involves recruitment of the macroautophagy pathway by BAG3. EMBO J 28(7):889–901. doi:10.1038/emboj.2009.29
Liu S, Kulp SK, Sugimoto Y, Jiang J, Chang HL, Dowd MK, Wan P, Lin YC (2002) The (−)-enantiomer of gossypol possesses higher anticancer potency than racemic gossypol in human breast cancer. Anticancer Res 22(1A):33–38
Boiani M, Daniel C, Liu X, Hogarty MD, Marnett LJ (2013) The stress protein BAG3 stabilizes Mcl-1 protein and promotes survival of cancer cells and resistance to antagonist ABT-737. J Biol Chem 288(10):6980–6990. doi:10.1074/jbc.M112.414177
Lian J, Wu X, He F, Karnak D, Tang W, Meng Y, Xiang D, Ji M, Lawrence TS, Xu L (2011) A natural BH3 mimetic induces autophagy in apoptosis-resistant prostate cancer via modulating Bcl-2–Beclin1 interaction at endoplasmic reticulum. Cell Death Differ 18(1):60–71. doi:10.1038/cdd.2010.74
Colvin TA, Gabai VL, Gong J, Calderwood SK, Li H, Gummuluru S, Matchuk ON, Smirnova SG, Orlova NV, Zamulaeva IA, Garcia-Marcos M, Li X, Young ZT, Rauch JN, Gestwicki JE, Takayama S, Sherman MY (2014) Hsp70-Bag3 interactions regulate cancer-related signaling networks. Cancer Res 74(17):4731–4740. doi:10.1158/0008-5472.CAN-14-0747
Yoon YJ, Kim JA, Shin KD, Shin DS, Han YM, Lee YJ, Lee JS, Kwon BM, Han DC (2011) KRIBB11 inhibits HSP70 synthesis through inhibition of heat shock factor 1 function by impairing the recruitment of positive transcription elongation factor b to the hsp70 promoter. J Biol Chem 286(3):1737–1747. doi:10.1074/jbc.M110.179440
Huber S, Oelsner M, Decker T, zum Buschenfelde CM, Wagner M, Lutzny G, Kuhnt T, Schmidt B, Oostendorp RA, Peschel C, Ringshausen I (2011) Sorafenib induces cell death in chronic lymphocytic leukemia by translational downregulation of Mcl-1. Leukemia 25(5):838–847. doi:10.1038/leu.2011.2
Slee EA, Adrain C, Martin SJ (2001) Executioner caspase-3, -6, and -7 perform distinct, non-redundant roles during the demolition phase of apoptosis. J Biol Chem 276(10):7320–7326. doi:10.1074/jbc.M008363200
Pattingre S, Tassa A, Qu X, Garuti R, Liang XH, Mizushima N, Packer M, Schneider MD, Levine B (2005) Bcl-2 antiapoptotic proteins inhibit Beclin 1-dependent autophagy. Cell 122(6):927–939. doi:10.1016/j.cell.2005.07.002
Cheng EH, Wei MC, Weiler S, Flavell RA, Mak TW, Lindsten T, Korsmeyer SJ (2001) BCL-2, BCL-X(L) sequester BH3 domain-only molecules preventing BAX- and BAK-mediated mitochondrial apoptosis. Mol Cell 8(3):705–711
Debatin KM (2004) Apoptosis pathways in cancer and cancer therapy. Cancer Immunol Immunother 53(3):153–159. doi:10.1007/s00262-003-0474-8
Zhou F, Yang Y, Xing D (2011) Bcl-2 and Bcl-xL play important roles in the crosstalk between autophagy and apoptosis. FEBS J 278(3):403–413. doi:10.1111/j.1742-4658.2010.07965.x
Gao P, Bauvy C, Souquere S, Tonelli G, Liu L, Zhu Y, Qiao Z, Bakula D, Proikas-Cezanne T, Pierron G, Codogno P, Chen Q, Mehrpour M (2010) The Bcl-2 homology domain 3 mimetic gossypol induces both Beclin 1-dependent and Beclin 1-independent cytoprotective autophagy in cancer cells. J Biol Chem 285(33):25570–25581. doi:10.1074/jbc.M110.118125
Rapino F, Jung M, Fulda S (2014) BAG3 induction is required to mitigate proteotoxicity via selective autophagy following inhibition of constitutive protein degradation pathways. Oncogene 33(13):1713–1724. doi:10.1038/onc.2013.110
Koehler BC, Scherr AL, Lorenz S, Elssner C, Kautz N, Welte S, Jaeger D, Urbanik T, Schulze-Bergkamen H (2014) Pan-Bcl-2 inhibitor obatoclax delays cell cycle progression and blocks migration of colorectal cancer cells. PLoS ONE 9(9):e106571. doi:10.1371/journal.pone.0106571
Festa M, Del Valle L, Khalili K, Franco R, Scognamiglio G, Graziano V, De Laurenzi V, Turco MC, Rosati A (2011) BAG3 protein is overexpressed in human glioblastoma and is a potential target for therapy. Am J Pathol 178(6):2504–2512. doi:10.1016/j.ajpath.2011.02.002
Tagscherer KE, Fassl A, Campos B, Farhadi M, Kraemer A, Bock BC, Macher-Goeppinger S, Radlwimmer B, Wiestler OD, Herold-Mende C, Roth W (2008) Apoptosis-based treatment of glioblastomas with ABT-737, a novel small molecule inhibitor of Bcl-2 family proteins. Oncogene 27(52):6646–6656. doi:10.1038/onc.2008.259
Dai C, Whitesell L, Rogers AB, Lindquist S (2007) Heat shock factor 1 is a powerful multifaceted modifier of carcinogenesis. Cell 130(6):1005–1018. doi:10.1016/j.cell.2007.07.020
Jacobs AT, Marnett LJ (2009) HSF1-mediated BAG3 expression attenuates apoptosis in 4-hydroxynonenal-treated colon cancer cells via stabilization of anti-apoptotic Bcl-2 proteins. J Biol Chem 284(14):9176–9183. doi:10.1074/jbc.M808656200
Carra E, Barbieri F, Marubbi D, Pattarozzi A, Favoni RE, Florio T, Daga A (2013) Sorafenib selectively depletes human glioblastoma tumor-initiating cells from primary cultures. Cell Cycle 12(3):491–500. doi:10.4161/cc.23372
Acknowledgments
The authors would like to thank Gabriele Köpf for excellent technical assistance. This study was supported by a grant of the Medical Faculty, Goethe University Frankfurt (Frankfurter Forschungsförderung 2012), to JM.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
No potential conflicts of interest were disclosed.
Ethical standard
The manuscript does not contain clinical studies or patient data.
Rights and permissions
About this article

Cite this article
Mani, J., Antonietti, P., Rakel, S. et al. Knockdown of BAG3 sensitizes bladder cancer cells to treatment with the BH3 mimetic ABT-737. World J Urol 34, 197–205 (2016). https://doi.org/10.1007/s00345-015-1616-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00345-015-1616-2