
Abstract
Lotus corniculatus is exposed to many forms of biotic stresses, including pathogen and herbivore attack. Jasmonates (JAs) are lipid-based signaling molecules that play important roles in mediating secondary metabolite biosynthesis and plant defense responses. Here, we first characterized LcJAZ1 and LcMYC2, two key components of the JAs signaling pathway. Both LcJAZ1 and LcMYC2 mRNA accumulated quickly in response to JAs. Subcellular localization analysis showed that both LcJAZ1 and LcMYC2 are nuclear proteins. In addition, LcJAZ1 protein was degraded after JAs treatment, but LcMYC2 is not. Yeast two-hybrid, pull-down, and bimolecular fluorescence complementation assays demonstrated that LcJAZ1 interacts with LcMYC2 and AtMYC2. In addition, LcMYC2 interacts with AtJAZ1. The Arabidopsis protoplast transactivation assay revealed that LcJAZ1 acts as repressors of LcMYC2- or AtMYC2-mediated transcription. Our data indicate that the conserved JAZ-MYC complex contributes to the modulation of the JAs signaling pathway.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Lotus corniculatus L. (Bird’s-foot trefoil), a member of Fabaceae, is considered the most agriculturally important forage plant because of the main agricultural advantages, such as the anti-bloating properties due to tannin content, ability to grow in low-fertility, acidic, and high-salinity soils (Sun and others 2014). Although L. corniculatus is generally abiotic stress tolerant, it can and does succumb to diseases caused by a wide range of biotic stresses including insects and pathogens. To defend itself against different types of attackers, the plant has developed an array of constitutive and induced defensive strategies. Jasmonic acid (JA) and derived jasmonates (JAs) are lipid-based signaling molecules that activate plant defenses against many forms of biotic and abiotic stresses and regulate diverse developmental processes (Fonseca and others 2009). In the JAs signal perception and transduction pathway, several components have been identified. CORONATINE INSENSITIVE1 (COI1) protein is an F-box protein that forms part of a putative E3 ubiquitin ligase complex of the Skp1/Cullin/F-box (SCFCOI1) type (Pieterse and others 2009). The importance of COI1 in the perception of bioactive JAs (that is, jasmonoyl-l-isoleucine or JA-Ile) was revealed with the discovery of the JA-Ile co-receptor Jasmonate ZIM-motif (JAZ) proteins, which act as repressors of JAs-responsive transcription (Chini and others 2007; Thines and others 2007; Yan and others 2009; Sheard and others 2010). The basic helix-loop-helix (bHLH) transcription factors (TFs) MYC2, MYC3, and MYC4 were identified as JAZ protein-binding partners (Chini and others 2007; Cheng and others 2011; Fernández-Calvo and others 2011; Niu and others 2011). More recently, the bHLH-related TFs GL3, EGL3 and TT8 (Qi and others 2011), bHLH3 (JAM3), bHLH13 (JAM2), and bHLH14 and bHLH17 (JAM1) (Nakata and others 2013; Sasaki-Sekimoto and others 2013; Song and others 2013) also have been shown to interact with JAZ repressors. In response to JA-Ile, JAZ proteins are rapidly degraded (Chini and others 2007; Thines and others 2007), which lead to de-repression of bHLHs and to the activation of plant defenses against wounding and insect herbivory.
Currently, completion of high-quality sequencing of the L. corniculatus var. japonicus genome has provided an excellent opportunity for searching key components of the JAs signaling pathway in Lotus (Sato and others 2008). In this study, we focus on the analysis of AtJAZ1 and AtMYC2 orthologous genes through bioinformatics analysis from the available genome database from L. corniculatus var. japonicus (Sato and others 2008). Yeast two-hybrid, pull-down, and bimolecular fluorescence complementation (BiFC) assays showed that the LcJAZ1 protein interacted with LcMYC2 and AtMYC2; interestingly, LcMYC2 also interacted with AtJAZ1. In the Arabidopsis protoplast transactivation assays, both LcJAZ1 and AtJAZ1 act as repressors of LcMYC2 or AtMYC2. Our results indicate that LcJAZ-LcMYC interaction is an evolutionarily conserved component of the JAs signaling pathway in L. corniculatus.
Materials and Methods
Searching for LcMYC2 and LcJAZ1 Genes
Protein sequences of Arabidopsis AtMYC2 (At1g32640) and AtJAZ1(At1g19180) were used as queries to search against the protein database of Lotus Japonicus (http://www.kazusa.or.jp/lotus/) and the National Centre for Biotechnology Information (NCBI; http://www.ncbi.nlm.nih.gov/). All sequences with an E value <1e-6 were selected for manual inspection.
Plant Materials and Hormone Treatment
Lotus corniculatus cv Leo seeds were sterilized and germinated on the Murashige–Skoog (MS) agar medium in a growth chamber with 16 h of light/8 h of dark; 30-day-old seedlings were used to examine the expression patterns of LcJAZ1 and LcMYC2 genes under 50 μM methyl jasmonate (MeJA) treatment. The seedlings were harvested at 0.5, 2, 6, 12, and 24 h after treatment. Protoplasts were treated for 4 h with 50 μM MeJA or 50 μM MG132 dissolved in DMSO at a final concentration of 0.1 %. All samples were frozen in liquid nitrogen and stored at −80 °C until use.
Quantitative Real-Time PCR Analysis (qRT-PCR)
Total RNA was isolated from leaves using an RNAprep pure Plant Kit, following the manufacturer’s instructions and treated with RNase-free DNase. The quality of RNA was checked on agarose gel electrophoresis. Two µg of RNA of each sample was reverse transcribed into cDNA using TransScript cDNA synthesis supermix (TransGen Biotech.). The list of gene-specific primers used for qRT-PCR is given in Supplementary Table S1. PCR reactions were carried out as described previously (Zhou and others 2011; Sun and others 2014). At least three biological and three technical replicates were performed for each reaction.
Arabidopsis Protoplast Transformation and Microscopic Analysis
The list of gene-specific primers used for PCR is given in Supplementary Table S1. The AtMYC2, AtJAZ1, LcMYC2, and LcJAZ1 full-length fragments were cloned in the pRTL2 derivatives or p163-GFP (Bracha-Drori and others 2004). Co-transformation or transformation with 10 µg each of plasmids carrying N-terminal YFP-fused protein and C-terminal YFP-fused protein or GFP and GFP-fused protein were introduced by PEG-mediated transfection as previously described into Arabidopsis protoplasts (Schirawski and others 2000). Images of transfected protoplasts were acquired with a Leica DM IRBE confocal laser scanning microscope.
Yeast Two-Hybrid (Y2H) Assays
Full-length AtMYC2, AtMYC3, AtMYC4, and LcMYC2 cloned in pACT2 were co-transformed with empty pAS2.1, pAS2.1-AtJAZ1, and pAS2.1-LcJAZ1, respectively, to yeast strain PJ64-4A (James and others 1996). The list of gene-specific primers used for PCR is given in Supplementary Table S1. Interaction assays were performed by co-transformation of bait and prey plasmids into yeast strain PJ64-4A according to a yeast transformation protocol modified from Gietz and others (1992), and plated on SD-LW medium. As controls, empty pAS2.1 and pACT2 were used. Transformants were allowed to grow for 4–5 days. Subsequently, cells were incubated for 16 h in liquid SD-LW, and 10 µL of 10 and 100-fold dilutions were spotted on solid SD-LWH supplemented with increasing 3-AT concentrations ranging from 0 to 20 mM. Yeast cells were allowed to grow for 7 days at 30 °C.
In Vitro Pull-Down Assay
To produce HA-tagged proteins, LcJAZ1 and AtJAZ1 were amplified and cloned in pASK-IBA45plus. To produce Strep-tagged proteins, LcMYC2 and AtMYC2 were amplified and cloned in pASK-IBA45plus. The list of gene-specific primers used for PCR is given in Supplementary Table S1. These constructs were transformed into E. coli strain BL21 (DE3) pLysS selecting on 50 μg/mL chloramphenicol and 200 μg/mL carbenicillin, and protein expression was induced by 0.2 μg/mL anhydrotetracycline. The soluble Strep fusion proteins were extracted and immobilized onto Strep-tactin Sepharose (IBA), whereas the soluble HA-fusion proteins were extracted [bacterial protein extraction reagent (B-PER)] and immobilized onto anti-HA agarose beads (Thermo Scientific).
For pull-down assays, 2 µg HA-JAZ1 and/or HA-AtJAZ1 was incubated with the immobilized Strep-LcMYC2 and or Strep-AtMYC2 fusion proteins at 4 °C for 1 h. Proteins retained on the beads were subsequently resolved by SDS-PAGE and detected with anti-HA antibody (Roche). Detection was carried out by incubating the blots in 6 mL of the western lightning ultra reagent (Perkin-Elmer) and exposure to X-ray films (Fuji).
Arabidopsis Protoplast Transactivation Assays
The 1472 bp fragment upstream of the putative translation start site (ATG) of the VSP2 (At5g24770) gene was PCR-amplified and cloned in plasmid GusXX as a reporter (Pasquali and others 1994). Co-transformation with plasmids carrying VSP2-promoter-GUS and effector plasmids carrying AtMYC2 and LcMYC2 fused to the CaMV 35S promoter in pRT101 were carried out. The list of gene-specific primers used for PCR is given in Supplementary Table S1. To study a possible effect of JAZ interaction with MYC2, a ratio of 2:2:2 (μg GUS:AtMYC2/LcMYC2:AtJAZ1/LcJAZ1) was chosen. As controls, co-transformations of VSP2-promoter-GUS with the empty pRT101 expression vector were used. Plasmid DNA was introduced into protoplasts by PEG-mediated transfection as previously described (Schirawski and others 2000). GUS activity assays were performed as described previously (van der Fits and Memelink 1997).
Plants Protein Extraction and Immunoblot Analysis
Protein extracts (extraction buffer: 25 mM Na-phosphate buffer pH 7.5, 1 mM EDTA, 7 mM β-mercaptoethanol, 1 % triton X-100, and 10 % glycerol) were separated on SDS-PAA gels and transferred to protran nitrocellulose by semidry blotting. After blocking 1 h in TBST with 5 % non-fat dry milk at room temperature, the Western blots were incubated overnight with HA or GFP peroxidase antibodies (1:2000; Roche) in TBST with 5 % non-fat milk. After 1 h incubation at room temperature, the blots were washed ×4 with TBST. Detection was carried out by incubating the blots in 6 mL of the western lightning ultra reagent (Perkin-Elmer) and exposure to X-ray films (Fuji).
Generation of Transgenic Plants
For the construction of transgenic lines constitutively overexpressing LcJAZ1-HA or LcMYC2-HA, the Cauliflower Mosaic Virus (CaMV) 35S cassette containing the LcJAZ1-HA or LcMYC2-HA ORF was cloned in pCAMBIA1300. The binary vector pCAMBIA1300-LcJAZ1-HA or LcMYC2-HA was introduced into Agrobacterium tumefaciens strain LBA4404. Arabidopsis plants were transformed using the floral dip method. Transgenic plants were selected on solid MA medium containing 100 mg/L timentin and 50 mg/L hygromycin. Transgenic plants from T2 generations were selected on MA medium containing only 50 mg/L hygromycin. Positive lines were tested for the protein levels of LcJAZ1-HA or LcMYC2-HA by Western blot.
Anthocyanin Determination
For anthocyanin measurement, 2-week-old Arabidopsis seedlings grown on MS medium with 50 µM MeJA were measured as described by Swain and Hillis (Swain and Hillis 1959). The anthocyanin content is presented as (A535–A650)/g fresh weight. The experiment was repeated three biological times.
Results
Characterization of LcJAZ1 and LcMYC2
Through a systematic computational analysis, 1 AtJAZ1 (At1g19180) (chr6.CM0013.820.r2.d) and 1 AtMYC2 (At1g32640) (chr2.CM0177.810.r2.m) putative orthologous genes were identified from the current L. corniculatus var. japonicus genome database (detailed information about these genes is described in Supporting Information Document 1). The full-length cDNA sequences of the chr6.CM0013.820.r2.d gene (designated LcJAZ1) and chr2.CM0177.810.r2.m (designated LcMYC2) were isolated from L. corniculatus total RNA. Localization of LcJAZ1 and LcMYC2 fused to the green fluorescent protein (GFP) was analyzed in Arabidopsis cell suspension protoplasts. As shown in Fig. 1, both LcJAZ1- and LcMYC2-fused GFP were found in the nucleus of Arabidopsis protoplasts, whereas GFP alone was expressed, the protein was localized in the cytoplasm and nucleus.

Subcellular localization of LcJAZ1 and LcMYC2. Transiently expressed LcJAZ1-GFP, LcMYC2-GFP, or GFP in Arabidopsis protoplasts. Scale bar 20 µm
JAZ repressors act as switches in JAs-responsive gene expression due to their JAs-stimulated degradation via the action of the JAs-receptor COI1 (Chini and others 2007; Thines and others 2007). Therefore, we investigated LcJAZ1 gene expression and protein stability in response to JA. As shown in Fig. 2, both LcJAZ1 and LcMYC2 mRNA accumulated quickly in response to JAs and reached its maximal level at 0.5 h. To study protein stability, plasmids expressing GFP-tagged LcJAZ1 proteins were co-expressed in Arabidopsis protoplasts with GFP, and the proteins were extracted after treatments with JA, MG132, or the solvent DMSO. Fluorescence microscopy showed that MG132, but not DMSO and JA, increased the abundance of LcJAZ1-GFP in protoplasts (Fig. 3a). As shown in Fig. 3b, MG132 treatment drastically increased LcJAZ1-GFP accumulation in protoplasts, indicating that LcJAZ1 protein is subject to 26S proteasome-mediated degradation. However, LcMYC2 protein amount was not affected by JA or MG132 treatment (data not shown).

The expression of LcJAZ1 and LcMYC2 in response to MeJA as assessed by qRT-PCR. The data were normalized to wild-type (set at 1) using the comparative threshold cycle method. Error bars represent the standard deviation of triplicate runs for qRT-PCR

LcJAZ1 is degraded by the 26S proteasome. a Confocal laser scanning microscopy images of Arabidopsis protoplasts expressing LcJAZ1-GFP or GFP. Eighteen hours after transformation, protoplasts were treated for 4 h with 50 μM MeJA and/or 50 μM MG132 or 0.1 % DMSO. b Transiently expressed LcJAZ1-GFP and GFP in Arabidopsis protoplasts. The proteins were extracted 18 h after transformation of protoplasts and detected with anti-GFP antibodies
LcJAZ1 Interacts with LcMYC2
Most AtJAZ proteins were shown to interact in vitro and in yeast with AtMYC2 (Chini and others 2007, 2009; Chung and Howe 2009). Because LcMYC2, a bHLH TF, acts as an orthologous gene of AtMYC2, we performed a yeast two-hybrid assay to test whether LcJAZ1 protein could interact with LcMYC2 protein. We used AtMYC2 as a positive control. As shown in Fig. 4, AtMYC2 interacted with AtJAZ1 consistent with previous reports (Chini and others 2009; Fernández-Calvo and others 2011). We found that LcJAZ1 interacted with LcMYC2. Yeast cells co-expressing LcJAZ1 and LcMYC2 were able to sustain growth at 3-AT concentrations up to 10 mM on selective medium. These interactions are considered significant because background auto-activation was undetectable.

MYC2 interacts with JAZ1 in yeast. Yeast cells expressing LcJAZ1 and AtJAZ1 proteins fused to the GAL4 BD and AtMYC2 and LcMYC2 fused to the GAL4 AD were spotted on SD/LW (-2) to select for the plasmids and on SD/-LWH with 10 mM 3-AT (-3) to select for transcriptional activation of the His3 gene. Growth was monitored after 7 days. Yeast cells transformed with the empty plasmids pAS2.1 and pACT2, expressing GAL4 BD and AD, respectively, were used as controls
To confirm the interaction of LcJAZ1 with LcMYC2 proteins in planta, a BiFC assay was employed. The N-terminal (YN) or C-terminal (YC) parts of the yellow fluorescent protein (YFP) were fused either N-terminally or C-terminally with LcJAZ1 and LcMYC2. The constructs were transiently co-expressed in Arabidopsis suspension cell protoplasts in all possible combinations of YN and YC fusion pairs with the unfused YFP parts as negative controls. Reconstitution of a fluorescing YFP chromophore occurred only upon co-expression of certain combinations of fusion proteins. The YFP signal was detected in the nucleus of Arabidopsis protoplasts co-transformed with YN-LcJAZ1 and LcMYC2-YC (Fig. 5). A similar result was observed upon co-expression of AtMYC2-YC with YN-AtJAZ1 as a positive control. Cells transfected with single plasmids and any combination of empty YFP vectors produced no or only background fluorescence. These results demonstrate that LcJAZ1 can interact with LcMYC2 in the nucleus of plant cells.

MYC2 interacts with JAZ1 assayed by BiFC in planta. YFP fluorescence images alone or merged with bright field images of Arabidopsis cell suspension protoplasts co-transformed with constructs encoding the indicated fusion proteins with YFP at the C-terminus (YC) or the N-terminus (YN). Scale bar 20 μm
Conserved JAZ-MYC Interaction
The highest amino acid identity of LcJAZ1 or LcMYC2 is shared with AtJAZ1 or AtMYC2, respectively, suggested that LcJAZ1 or LcMYC2 could interact with AtMYC2 or AtJAZ1 and indeed this was confirmed by Y2H assays (Fig. 4). In addition, LcJAZ1 also interacted with AtMYC3 and AtMYC4 in yeasts (Supporting Information Figure S2). To confirm the interaction of LcJAZ1 or LcMYC2 with AtMYC2 or AtJAZ1 proteins in planta, a BiFC assay was employed. As shown in Fig. 5, the YFP signal was detected in the nucleus of Arabidopsis protoplasts co-transformed with YN-LcJAZ1 and AtMYC2-YC (Fig. 5). A similar result was observed upon co-expression of LcMYC2-YC with YN-AtJAZ1 (Fig. 5). Our results revealed that the JAZ-MYC2 interaction is conserved in plants.
To verify the interaction of protein–protein in vitro, we performed pull-down assays using HA- and Strep-tagged proteins (Fig. 6). As shown in Fig. 6, HA-LcJAZ1 interacted with either Strep-LcMYC2 or Strep-AtMYC2. The positive control showed interaction between HA-AtJAZ1 and Strep-AtMYC2. These data are consistent with the results from yeast two-hybrid and BiFC assays.

MYC2 interacts with JAZ1 assayed by pull-down in vitro. a In vitro interaction between HA-LcJAZ1 or HA-AtJAZ1 and Strep-AtMYC2. b In vitro interaction between HA-LcJAZ1 or HA-AtJAZ1 and Strep-LcMYC2. HA-LcJAZ1 or HA-AtJAZ1 protein was incubated with immobilized Strep-AtMYC2 or Strep-LcMYC2. The immuno-precipitated fractions were detected with anti-HA antibody. Input was detected with anti-Strep antibody. Asterisks denote bands from HA antibody
LcJAZ1 and LcMYC2 Mediated JA Responses
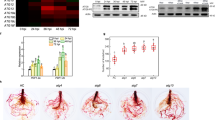
To functionally test the interaction between LcJAZ1/AtJAZ1 and AtMYC2/LcMYC2, transactivation assays were performed. Co-transformation of Arabidopsis cell suspension protoplasts with a VSP2 promoter-GUS reporter construct and an effector plasmid carrying AtMYC2 or LcMYC2 fused to the CaMV 35S promoter resulted in strong activation of around 20-fold or 15-fold, respectively (Fig. 7a). Combinations with effector plasmids carrying LcJAZ1 or AtJAZ1 caused repression of AtMYC2 or LcMYC2 activity (Fig. 7a). Thus, in agreement with previous studies, the activity of MYC2 was inhibited by JAZ1 (Hou and others 2010). Our results from transactivation assays indicated that LcJAZ1 is a negative regulator of AtMYC2. To investigate whether LcJAZ1 protein regulates VSP2 expression in plants, Arabidopsis plants constitutively overexpressing LcJAZ1-HA were analyzed (Supporting Information Figure S2). qRT-PCR performed with two-week-old seedlings showed that the VSP2 gene was significantly repressed in LcJAZ1-HA overexpressing plants compared to wild-type (WT) plants (Fig. 7b). Additionally, the VSP2 gene was highly induced in LcMYC2-HA overexpressing plants compared to WT plants (Fig. 7b). To determine whether modulation of LcJAZ1 or LcMYC2 expression levels affects JA responses, JA-induced anthocyanin accumulation tests were performed for all genotypes (Fig. 7c). As shown in Fig. 7c, the LcJAZ1-HA overexpressing plants exhibited reduced JA responses compared with wild-type, as indicated by reduction in JA-induced anthocyanin accumulation. However, the LcMYC2-HA overexpressing plants showed enhanced JA responses. Taken together with data of the genetic and physiological analysis on the overexpression transgenic lines, we demonstrated that the LcJAZ1 and LcMYC2 probably function as negative and positive regulators of JA responses, respectively.

LcJAZ1 and LcMYC2 mediated JA responses. a JAZ1 represses MYC2-activated VSP2pro-GUS activity. Arabidopsis cell suspension protoplasts were co-transformed with plasmids carrying VSP2pro-GUS (2 μg) and/or overexpression vectors containing 35S:MYC2 and/or 35S:JAZ1 (2 μg), as indicated. Protein concentrations were used to correct for differences in protein extraction efficiencies. Error bars represent the standard deviation of three independent measurements. b Expression of VSP2 in all genotypes as assessed by qRT-PCR. c Anthocyanin contents of the 2-week-old seedlings in all genotypes grown on MS medium containing 50 μM MeJA. WT, wild-type; LcMYC2 OX, LcMYC2-HA overexpressing plants; LcJAZ1 OX, LcJAZ1-HA overexpressing plants; FW fresh weight. The mean value is from three independent transgenic lines, and error bars indicate the standard deviation. Asterisks denote Student’s t test significance: **P < 0.01, *P < 0.05
Discussion
JAs regulate two distinct and antagonistic branches of plant defenses, against herbivorous insects and against necrotrophic pathogens (Pieterse and others 2009). Within the signaling cascades that are triggered by JAs, the JAZ repressor proteins play a central role. The expression of the JAZ genes in Arabidopsis is induced by JA (Thines and others 2007; Yan and others 2007) and is controlled by MYC2 (Chini and others 2007) and MYC3 and MYC4 (Fernández-Calvo and others 2011; Niu and others 2011). MYC and JAZ proteins form a JAs-responsive oscillator, where JAZ proteins negatively regulate MYC activity at the protein level, JAs cause JAZ degradation and MYC activation, and MYC switches on the expression of JAZ repressors at the gene level. Here, we identified LcJAZ1 and LcMYC2 from L. corniculatus. LcJAZ1 probably functions as negative regulator to repress JA responses, whereas LcMYC2 functions as a positive one. Similar to AtJAZ1, LcJAZ1 was induced by JAs at the gene expression level and was degraded by JAs at the protein level (Fig. 2, 3). AtJAZ proteins contain, in addition to the highly conserved central ZIM domain, a highly conserved C-terminal Jas domain and a less conserved N-terminal region. The Jas domain of the AtJAZ1 is responsible for the interaction with MYC factors, COI1, and several other R2R3-MYB TFs (Chini and others 2007; Pauwels and Goossens 2011). It has been reported that the ZIM domain mediates homo- and heterodimerization of AtJAZ proteins (Chini and others 2009; Chung and Howe 2009). The ZIM domain of JAZ also contributes to the interaction with the NOVEL INTERACTOR OF JAZ (NINJA), which consists of the ERF-associated amphiphilic repression (EAR) domain bridging the JAZ with the co-repressors TOPLESS (TPL) and TPL-related proteins (TPRs) (Pauwels and others 2010). Additionally, MYC2 interacts with JAZ depending on its conserved JAZ interaction domain (JID) (Fernández-Calvo and others 2011). The JID is also essential for the interaction with the R2R3-MYB proteins involved in the biosynthesis of glucosinolates in Arabidopsis (Schweizer and others 2013). The highest amino acid identity of the LcJAZ1 Jas domain or LcMYC2 JID domain is shared with the AtJAZ1 Jas domain or AtMYC2 JID domain, with an overall identity of 81 or 67 %, respectively (data not shown). Here, we showed that LcJAZ1 could interact with both LcMYC2 and AtMYC2 proteins, whereas LcMYC2 interacts with both LcJAZ1 and AtJAZ1 (Figs. 4, 5, 6). The conserved Jas domains of tomato JAZ1 (Katsir and others 2008) and AtJAZ1 (Melotto and others 2008) were shown to be necessary for binding to COI1 in a JA-Ile or coronatine-dependent manner. These results revealed that both the Jas domain and JID domain are very conserved in the plant JAs signaling pathway. Overall, our results highlight that the conserved JAZ-MYC2 complex contributes to the function of JAs in plants. Further research is required to identify and characterize the proteins that interact with LcJAZ1, which will aid in better understanding of the mechanism and networking of JAs hormone signaling in Lotus.
References
Bracha-Drori K, Shichrur K, Katz A, Oliva M, Angelovici R, Yalovsky S, Ohad N (2004) Detection of protein–protein interactions in plants using bimolecular fluorescence complementation. Plant J 40:419–427
Cheng Z, Sun L, Qi T, Zhang B, Peng W, Liu Y, Xie D (2011) The bHLH transcription factor MYC3 interacts with the jasmonate ZIM-domain proteins to mediate jasmonate response in Arabidopsis. Mol Plant 4:279–288
Chini A, Fonseca S, Fernández G, Adie B, Chico JM, Lorenzo O, García-Casado G, López-Vidriero I, Lozano FM, Ponce MR, Micol JL, Solano R (2007) The JAZ family of repressors is the missing link in jasmonate signalling. Nature 448:666–671
Chini A, Fonseca S, Chico JM, Fernández-Calvo P, Solano R (2009) The ZIM domain mediates homo- and heteromeric interactions between Arabidopsis JAZ proteins. Plant J 59:77–87
Chung HS, Howe GA (2009) A critical role for the TIFY motif in repression of jasmonate signaling by a stabilized splice variant of the JASMONATE ZIM-domain protein JAZ10 in Arabidopsis. Plant Cell 21:131–145
Fernández-Calvo P, Chini A, Fernández-Barbero G, Chico JM, Gimenez-Ibanez S, Geerinck J, Eeckhout D, Schweizer F, Godoy M, Franco-Zorrilla JM, Pauwels L, Witters E, Puga MI, Paz-Ares J, Goossens A, Reymond P, De Jaeger G, Solano R (2011) The Arabidopsis bHLH transcription factors MYC3 and MYC4 are targets of JAZ repressors and act additively with MYC2 in the activation of jasmonate responses. Plant Cell 23:701–715
Fonseca S, Chico JM, Solano R (2009) The jasmonate pathway: the ligand, the receptor and the core signalling module. Curr Opin Plant Biol 12:539–554
Gietz D, St Jean A, Woods RA, Schiestl RH (1992) Improved method for high efficiency transformation of intact yeast cells. Nucleic Acids Res 20:1425
Hou X, Lee LYC, Xia K, Yan Y, Yu H (2010) DELLAs modulate jasmonate signaling via competitive binding to JAZs. Dev Cell 19:884–894
James P, Halladay J, Craig EA (1996) Genomic libraries and a host strain designed for highly efficient two-hybrid selection in yeast. Genetics 144:1425–1436
Katsir L, Schilmiller AL, Staswick PE, He SY, Howe GA (2008) COI1 is a critical component of a receptor for jasmonate and the bacterial virulence factor coronatine. Proc Natl Acad Sci USA 105:7100–7105
Melotto M, Mecey C, Niu Y, Chung HS, Katsir L, Yao J, Zeng W, Thines B, Staswick P, Browse J, Howe GA, He SY (2008) A critical role of two positively charged amino acids in the Jas motif of Arabidopsis JAZ proteins in mediating coronatine- and jasmonoyl isoleucine-dependent interactions with the COI1 F-box protein. Plant J 55:979–988
Nakata M, Mitsuda N, Herde M, Koo AJ, Moreno JE, Suzuki K, Howe GA, Ohme-Takagi M (2013) A bHLH-type transcription factor, ABA-INDUCIBLE BHLH-TYPE TRANSCRIPTION FACTOR/JA-ASSOCIATED MYC2-LIKE1, acts as a repressor to negatively regulate jasmonate signaling in Arabidopsis. Plant Cell 25:1641–1656
Niu Y, Figueroa P, Browse J (2011) Characterization of JAZ-interacting bHLH transcription factors that regulate jasmonate responses in Arabidopsis. J Exp Bot 62:2143–2154
Pasquali G, Ouwerkerk PBF, Memelink J (1994) Versatile transformation vectors to assay the promoter activity of DNA elements in plants. Gene 149:373–374
Pauwels L, Goossens A (2011) The JAZ proteins: a crucial interface in the jasmonate signaling cascade. Plant Cell 23:3089–3100
Pauwels L, Barbero GF, Geerinck J, Tilleman S, Grunewald W, Cuéllar Pérez A, Chico JM, Vanden Bossche R, Sewell J, Gil E, García-Casado G, Witters E, Inzé D, Long JA, De Jaeger G, Solano R, Goossens A (2010) NINJAconnects the corepressor TOPLESS to jasmonate signalling. Nature 464:788–791
Pieterse CMJ, Leon-Reyes A, Van der Ent S, Van Wees SCM (2009) Networking by small-molecule hormones in plant immunity. Nat Chem Biol 5:308–316
Qi T, Song S, Ren Q, Wu D, Huang H, Chen Y, Fan M, Peng W, Ren C, Xie D (2011) The jasmonate-ZIM-domain proteins interact with the WD repeat/bHLH/MYB complexes to regulate jasmonate-mediated anthocyanin accumulation and trichome initiation in Arabidopsis thaliana. Plant Cell 23:1795–1814
Sasaki-Sekimoto Y, Jikumaru Y, Obayashi T, Saito H, Masuda S, Kamiya Y, Ohta H, Shirasu K (2013) Basic helix-loop-helix transcription factors JASMONATE-ASSOCIATED MYC2-LIKE1 (JAM1), JAM2, and JAM3 are negative regulators of jasmonate responses in Arabidopsis. Plant Physiol 163:291–304
Sato S, Nakamura Y, Kaneko T, Asamizu E, Kato T, Nakao M, Sasamoto S, Watanabe A, Ono A, Kawashima K, Fujishiro T, Katoh M, Kohara M, Kishida Y, Minami C, Nakayama S, Nakazaki N, Shimizu Y, Shinpo S, Takahashi C, Wada T, Yamada M, Ohmido N, Hayashi M, Fukui K, Baba T, Nakamichi T, Mori H, Tabata S (2008) Genome structure of the legume, Lotus japonicus. DNA Res 15:227–239
Schirawski J, Planchais S, Haenni AL (2000) An improved protocol for the preparation of protoplasts from an established Arabidopsis thaliana cell suspension culture and infection with RNA of turnip yellow mosaic tymovirus: a simple and reliable method. J Virol Methods 86:85–94
Schweizer F, Fernández-Calvo P, Zander M, Diez-Diaz M, Fonseca S, Glauser G, LewseyMG Ecker JR, Solano R, Reymond P (2013) Arabidopsis basic helix-loop-helix transcription factors MYC2, MYC3, and MYC4 regulate glucosinolate biosynthesis, insect performance, and feeding behavior. Plant Cell 25:3117–3132
Sheard LB, Tan X, Mao H, Withers J, Ben-Nissan G, Hinds TR, Kobayashi Y, Hsu FF, Sharon M, Browse J, He SY, Rizo J, Howe GA, Zheng N (2010) Jasmonate perception by inositol-phosphate-potentiated COI1-JAZ co-receptor. Nature 468:400–405
Song S, Qi T, Fan M, Zhang X, Gao H, Huang H, Wu D, Guo H, Xie D (2013) The bHLH subgroup IIId factors negatively regulate jasmonate-mediated plant defense and development. PLoS Genet 9(7):e1003653
Sun ZM, Zhou ML, Xiao XG, Tang YX, Wu YM (2014) Genome-wide analysis of AP2/ERF family genes from Lotus corniculatus shows LcERF054 enhances salt tolerance. Funct Integr Genom 14:453–466
Swain T, Hillis WE (1959) The phenolic constituents of Prunus domestica. I: the quantitative analysis of phenolic constituents. J Agric Food Sci 10:63–68
Thines B, Katsir L, Melotto M, Niu Y, Mandaokar A, Liu G, Nomura K, He SY, Howe GA, Browse J (2007) JAZ repressor proteins are targets of the SCFCOI1 complex during jasmonate signalling. Nature 448:661–665
van der Fits L, Memelink J (1997) Comparison of the activities of CaMV 35S and FMV 34S promoter derivatives in Catharanthus roseus cells transiently and stably transformed by particle bombardment. Plant Mol Biol 33:943–946
Yan Y, Stolz S, Chételat A, Reymond P, Pagni M, Dubugnon L, Farmer EE (2007) A downstream mediator in the growth repression limb of the jasmonate pathway. Plant Cell 19:2470–2483
Yan J, Zhang C, Gu M, Bai Z, Zhang W, Qi T, Cheng Z, Peng W, Luo H, Nan F, Wang Z, Xie D (2009) The Arabidopsis CORONATINE INSENSITIVE1 protein is a jasmonate receptor. Plant Cell 21:2220–2236
Zhou ML, Hou HL, Zhu XM, Shao JR, Wu YM, Tang YX (2011) Soybean transcription factor GmMYBZ2 represses catharanthine biosynthesis in hairy roots of Catharanthus roseus. Appl Microbiol Biotechnol 91:1095–1105
Acknowledgments
This work was supported by the National Natural Science Foundation of China (Grant No. 31372361) and National Program on Key Basic Research Project (973 Program) (Grant No. 2014CB138701).
Author information
Authors and Affiliations
Corresponding authors
Additional information
Meiliang Zhou and Zhanmin Sun have contributed equally to this work.
Electronic supplementary material
Below is the link to the electronic supplementary material.
344_2015_9551_MOESM2_ESM.pdf
Supplementary material 2 . Supporting Information Figure S1: AtMYC3 and AtMYC4 interact with LcJAZ1 and AtJAZ1 in yeast. Yeast cells expressing LcJAZ1 and AtJAZ1 proteins fused to the GAL4 BD and AtMYC3 and AtMYC4 fused to the GAL4 AD were spotted on SD/LW (-2) to select for the plasmids and on SD/-LWH with 10 mM 3-AT (-3) to select for transcriptional activation of the His3 gene. Growth was monitored after 7 days. Yeast cells transformed with the empty plasmids pAS2.1 and pACT2, expressing GAL4 BD and AD, respectively, were used as controls. (PDF 41 kb)
344_2015_9551_MOESM3_ESM.pdf
Supplementary material 3. Supporting Information Figure S2: Western blot using anti-HA to identify the positive transgenic plants. W: wild-type plants. (PDF 49 kb)
Rights and permissions
About this article

Cite this article
Zhou, M., Sun, Z., Li, J. et al. Identification of JAZ1-MYC2 Complex in Lotus corniculatus . J Plant Growth Regul 35, 440–448 (2016). https://doi.org/10.1007/s00344-015-9551-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00344-015-9551-4