
Abstract
Universal Adaptive Strategy Theory aims to predict how taxa and assemblages respond to disturbances on the basis of adaptive strategy group (ASG) membership. Here, we test such predictions using the adaptive strategy scheme for reef-building corals developed by Darling et al. (Ecol Lett 15:1378–1386, 2012) and a long-term dataset of coral assemblage structure from inshore reefs on the central Great Barrier Reef. Several disturbances including mass bleaching and tropical storms were recorded in this 15-year interval from 1998 to 2013. ASG membership did not predict how a given taxon responded to disturbance. In fact, all ASGs were on average equally affected by bleaching and a period of multiple disturbances. Furthermore, there were no consistent winners at these sites in response to the 1998 bleaching in contrast to previous work suggesting clear hierarchies in susceptibility to bleaching. In conclusion, while further efforts to re-evaluate the utility of ASGs for reef corals should be encouraged our results and a re-examination of the literature suggests that direct trait-based approaches might prove more useful when exploring how corals respond to disturbance.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Adaptive strategy theory aims to reduce the number of units required to describe assemblages by grouping species based on life history strategies and functional roles. A key utility stemming from such theory is the promise to predict how taxa and assemblages respond to ecological disturbance. For example, the Universal Adaptive Strategy Theory (UAST) of Grime and Pierce (2012) identifies three adaptive groups and makes very specific predictions about the conditions under which each group should dominate and how each should respond to stress and disturbance: competitive species should benefit when disturbance and stress are low; stress-tolerant species should benefit when the intensity of stress is high; and ruderal species should benefit when disturbances are frequent (Grime and Pierce 2012).
The distinction between stress and disturbance is an important aspect of UAST (Grime and Pierce 2012): stress is defined as “the sum of many agents that limit the quantity of living matter created per unit of space and time by constraining its production”; and disturbance as “the sum of the great multiplicity of agents that limit biomass by partly or completely destroying it” (Grime and Pierce 2012). In other words, stress slows production and disturbance destroys production. Under this scheme, bleaching, flooding, and eutrophication can be defined as stresses affecting coral assemblages, whereas severe tropical storms and population outbreaks of corallivorous crown-of-thorns starfish can be defined as disturbances.
Corals often vary in their specific vulnerability to different disturbances and stresses (Van Woesik et al. 1995; Marshall and Baird 2000; Madin et al. 2014; Pratchett et al. 2014; Hughes et al. 2018), such that changing disturbance regimes can modify the species composition of coral assemblages (McWilliam et al. 2020; Pratchett et al. 2020). The adaptive strategy approach has been used to try and explain or predict the response of coral assemblages to stress and disturbance (e. g., Darling et al. 2013; Graham et al. 2014; Sommer et al. 2014). Darling et al. (2013) found that the initial relative abundance of adaptive strategy groups (ASGs) of Darling et al. (2012) could predict change in coral assemblage structure on coastal reefs in Kenya in response to multiple stressors including fishing, which they defined as a disturbance, and a bleaching event, defined as a stress. Similarly, Graham et al. (2014) concluded that the relative abundance of Darling et al.’s (2012) ASGs was useful for distinguishing among reefs with a different disturbance history on mid-shelf reefs in the central region of the Great Barrier Reef (GBR). Specifically, the relative abundance of competitive taxa was higher on undisturbed and recovered reefs than on reefs that had not recovered from a population irruption of crown-of-thorns starfish. Sommer et al. (2014) also used Darling’s scheme to compare the relative abundance of species in each adaptive group among coral assemblages along a high-latitude gradient in south-eastern Australia that included sites at the range limit of most coral species, documenting a decrease in the relative abundance of stress-tolerant species in coral assemblage at higher latitudes.
In this paper, we test whether the groups identified in Darling et al. (2012, 2013) are good predictors of the response of coral taxa to environmental stasis and change. We use a 15-year data set that documents changes in the abundance of coral genera at eight sites in the central GBR. This time-period captures a number of stresses and disturbances, including the 1998 mass-bleaching event, a tropical cyclone, and floods as well as periods of stasis when there were no disturbance events. We document winners and losers through time in order to test three sets of predictions of UAST: (1) that stress-tolerant species are less susceptible to mass bleaching compared with the other adaptive strategies; (2) competitive species increase in abundance during periods of stasis; and (3) weedy species increase in abundance after disturbances, and more-so after repeated disturbances to the same location. In addition, we tested whether the traditional bleaching hierarchies, e.g. the bleaching mortality index (BMI) (McClanahan et al. 2004), were able to reflect the mortality estimates from changes in abundance.
Materials and methods
Study site
Magnetic Island and the Palm Island group are continental islands with extensive fringing reefs located inshore within the central region of the GBR (Fig. 1). The reef environment is characterised by relatively shallow (< 15 m), highly turbid waters, with underwater visibility rarely exceeding five metres. Reefs around these islands are relatively sheltered from oceanic conditions by the expanse of the GBR lagoon, but are exposed to the influence of nearby rivers. Reef development around inshore islands on the GBR is often patchy, giving way to soft sediments as shallow as eight metres on landward reefs. Coral assemblages were monitored at two locations in each island group: Nelly Bay (−19.167, 146.850) and Geoffrey Bay (−19.155, 146.861) at Magnetic Island; and Little Pioneer Bay (−18.594, 146.485) and southeast Pelorus (−18.560, 146.500) at the Palm Islands. Two depths (shallow (S): 2–4 m; deep (D): 5–8 m) were surveyed at each location to give a total of eight sites.

Location of survey sites, including Nelly Bay (NB); Geoffrey Bay (GB); Little Pioneer Bay (LPB); and southeast Pelorus (SEP) in the central GBR region
Survey method
Between six and nine surveys were conducted at each site between 1998 and 2013. The first set of surveys was conducted in February/March 1998 during the mass bleaching event (Baird and Marshall 1998; Marshall and Baird 2000); the second set of surveys was conducted in September/October 1998 by which time the vast majority of the coral had recovered or died (Baird and Marshall 2002). Between four and six replicate 15 m × 0.5 m belt transects were used at each site on each survey. The abundance of all hard and soft corals with a maximum diameter greater than 5 cm within the belt transects was recorded. Hard corals include Scleractinia and Hydrocorallina, whereas soft corals are limited to Lobophytum, Sarcophyton, and Sinularia. All colonies were identified in the field to genus following Veron (2000) for the scleractinian corals and Fabricius and Aldersdale (2001) for the soft corals. Hard coral genera were further classified into ASGs following Darling et al (2012, 2013; Table 1). We used colony abundance instead of the more commonly used metric of coral cover because it provided a much better estimate of population-level mortality. The raw data are accessible from Baird et al. (2020).
Change in coral abundance through time
Change in the mean abundance of corals through time was tested using one-way ANOVA with Tukey post-hoc tests to identify marked temporal changes in coral abundance. Assumptions of normality of residuals and homogeneity of variances were assessed by reviewing plots of residuals against fitted values and Q–Q plots. A log-transformation was applied if violations of assumptions were detected.
Disturbance regime through the course of the study
The period of the study included a number of potential stresses and disturbances, including cyclones, bleaching, floods, and low tide events. These multiple stressors affected each site to a different and often unknown level (Table 2). In order to test the response of the taxa and ASGs, we defined three time-intervals based on the timing of disturbances:
-
1.
Stress—a bleaching event (March 1998);
-
2.
Recovery—no stress or disturbance. There was a brief period free of stress and disturbance on Magnetic Island (October 1998 to April 2000) and in the Palm Islands (December 2001 to March 2005). There were no periods without disturbance at the regional scale;
-
3.
Multiple-disturbances—the total time interval of the study (March 1998 to 2012/13) during which there were multiple disturbance and stress events
The response of taxa in the different time intervals
Changes in the abundance of the taxa during each of the three time-intervals were explored at both the site and regional level (i.e. all sites pooled) using Cohen’s d effect size, which is defined as the difference between two means of each time point, \(\bar{x}_1 \;\text{and} \;\bar{x}_2,\) divided by a pooled standard deviation, s, for the data and was estimated as follows (Cohen 1988):
Cohen’s d takes into account the variance in the data in addition to the difference between the means (Cohen 1988), which addresses potential sampling issues with for taxa with low abundances.
Winners were defined as taxa that increase in abundance in the given time interval and had an effect size > +0.8 which is described in the literature as a very strong effect (Cohen 1988). The strong effect means that colony abundance on 79% of transects sampled in the second time point was higher than the average colony abundance from transects sampled in the first time point. Losers were defined as taxa that decreased in abundance in the given time-interval and had an effect size < −0.8.
Bleaching response index vs. response as estimated by change in effect size
The bleaching mortality index (BMI) was developed by McClanahan et al. (2004) as a metric for comparing bleaching susceptibility among genus based on the categorical bleaching categories of Marshall and Baird (2000). The relationship between BMI and Cohen’s d was tested using linear regression.
Results
Changes in abundance through time
At the regional scale, there have been significant changes in coral abundance over the 15-year time period. The 1998 bleaching caused a 50% reduction in the mean abundance of corals. A gradual increase until 2005 was followed by a decline in abundance towards the lowest coral abundance in the study period in 2012/13 (Fig. 2; Table 3, S1). Seven of the eight sites have experienced significant changes in coral abundance, including at least one period of increase and decrease (Fig. 3; Table 3, S2–S9). Bleaching in 1998 caused significant declines in abundance at six of the eight sites (Fig. 3; Table 3). The only sites unaffected in terms of the overall abundance of coral were the sites at southeast Pelorus. Increases in coral abundance were evident at all sites following the bleaching in 1998. However, recovery at Magnetic Island sites was set back by another bleaching event in 2002 followed by subsequent multiple stressors. This has resulted in significantly fewer corals on Magnetic Island in 2013 compared to 1998, at three of the four sites, except GB-D (Fig. 3; Table 3). Bleaching in 2002 did not appear to affect the sites in Palm Island where coral abundance peaked in 2004/5. Since then, Palm Island sites have experienced multiple stressors, including cyclone Yasi in 2011, leading to significant declines in coral abundance. There were less corals at all sites in the Palm Islands in 2012 compared to 1998 (Fig. 3; Table 3).

Temporal changes in coral colony abundance at regional scale, all sites pooled together, in the central GBR region from 1998 to the last survey in 2013. The five time points are the surveys which were conducted at all eight sites. Arrows indicate disturbances, including two bleaching events (1998 and 2002, white), one low tide event (2005, grey), two flood events (2009, 2010–2011, black). The grey bar indicates tropical cyclone Yasi in 2011. Letters above dots indicate significant groupings by Tukey’s post hoc test at different surveys

Temporal changes in average coral colony abundance (mean ± SE) at 8 sites in the central GBR region from 1998 to 2013. Sites include Nelly Bay 2 m (a) 6 m (b), Geoffrey Bay 2 m (c) 6 m (d), Little Pioneer Bay 2 m (e) 6 m (f), and southeast Pelorus 4 m (g) 6 m (h). Arrows indicated disturbances including two bleaching events (1998 and 2002, white), one low tide event (2005, grey), two flood events (2009, 2010–2011, black). The grey bar indicates tropical cyclone Yasi in 2011. Letters above dots indicate significant groupings by one-way ANOVA, Welch's F test and Tukey’s post hoc test at different surveys
Response of taxa to bleaching in 1998
Most taxa decreased in abundance in response to bleaching. At a regional scale, 37 of the 48 taxa declined in abundance (Table 4, S1). Similarly, at six of the eight sites, most taxa declined in abundance (Table 4, S2–S9). Based on the effect size, losers greatly outnumbered winners at all sites (Table 4, S2–S9).
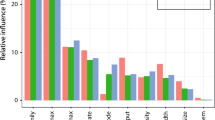
The losers varied greatly among sites (Table 5); however, some taxa were losers at multiple sites. For example, Montipora, Acropora, Cyphastrea, Turbinaria, Porites, Favia, Gonipora, Galaxea, Pocillopora, Sinularia and Montastrea were always among the losers at the five most affected sites (Table 5). Seriatopora and Stylophora were consistent losers at the sites that did not suffer large declines in total coral abundance (i.e. SEP-S & SEP-D). Losers came from all ASGs at both the regional scale (Fig. 4) and at most sites (Fig. 5).

Effect size by taxa for bleaching (a) and multiple disturbances (b) at regional scale. Colours indicate the ASG; competitive (red), generalist (grey), stress-tolerant (blue), weedy (green) and unknown (black)

Effect size by taxa for bleaching event. Sites include Nelly Bay 2 m (a) 6 m (b), Geoffrey Bay 2 m (c) 6 m (d), Little Pioneer Bay 2 m (e) 6 m (f), and southeast Pelorus 4 m (g) 6 m (h). Colours indicate the ASG; competitive (red), generalist (grey), stress-tolerant (blue), weedy (green) and unknown (black)
The winners in response to bleaching were very few (Table 6). At the regional scale there were no winners (Table 6). At the site scale Platygyra and Sarcophyton were winners at two sites and the following taxa were winners at one site; Alveopora, Astreopora, Galaxea, Montipora, Oulophyllia, Pachyseris, Leptastrea and Leptoseris (Table 6). Winners were either stress-tolerant species or generalist taxa (Figs. 4, 5).
Response of taxa to multiple disturbances
The response of taxa to multiple disturbances was very similar to the response to bleaching except there were even fewer winners and more losers, particularly at the sites at southeast Pelorus (Tables 4, 5, 6, S2–S9). At the regional scale, competitive and weedy taxa were more susceptible to multiple disturbances than stress-tolerant and generalist species (Fig. 4, S1). At the site scale, losers came from all ASGs (Fig. 6). At the regional scale there were no winners (Table 4). Winners at the site scale were either stress-tolerant species (Porites, Fungiidae, and Favites) or generalists (Mycedium and Pavona; Figs. 4, 6).

Effect size by taxa for multiple disturbances. Sites include Nelly Bay 2 m (a) 6 m (b), Geoffrey Bay 2 m (c) 6 m (d), Little Pioneer Bay 2 m (e) 6 m (f), and southeast Pelorus 4 m (g) 6 m (h). Colours indicate the ASG; competitive (red), generalist (grey), stress-tolerant (blue), weedy (green) and unknown (black)
Response of taxa to periods of no disturbance or stress
Winners outnumbered losers in the recovery periods at most sites (Table 4, S2–S9). The losing taxa varied greatly among sites with only Montastrea losing at more than one site (Table 5). A number of taxa were consistent winners. In particular, Montipora and Acropora were winners at four sites and Turbinaria, Favia, Favites, Sinularia, Porites and the Fungiidae at two or more (Table 5). Losers were mostly stress-tolerant species and generalist (Fig. 7) but also included weeds and one competitor at one site (Turbinaria at LPB-D). Winners included taxa from all ASGs (Fig. 7).

Effect size by taxa for recover period. Sites include Nelly Bay 2 m (a) 6 m (b), Geoffrey Bay 2 m (c) 6 m (d), Little Pioneer Bay 2 m (e) 6 m (f), and southeast Pelorus 4 m (g) 6 m (h). Colours indicate the ASG; competitive (red), generalist (grey), stress-tolerant (blue), weedy (green) and unknown (black)
Bleaching response index vs response as estimated by change in effect size
There was no correlation between susceptibility to bleaching as determined by the bleaching response index (BMI) and change in abundance as estimated by Cohen’s d effect size (R2 = 0.003, p = 0.362) (Fig. 8).

Bleaching mortality index vs. effect size. Blue line represents the smooth line of LOESS smoother. Red line and the formula above represent the linear regression line and regression result. Colours indicate the ASG; competitive (red), generalist (grey), stress-tolerant (blue), weedy (green) and unknown (black)
Discussion
No taxa were winners during the 15-year period of multiple stressors. At the regional scale, all taxa were less abundant in 2013 than in 1998, which was similar to studies conducted on reefs nearby in the same period (e.g., Torda et al. 2018). Despite these changes and a 50% reduction in coral abundance between 1998 and 2013, there have been no extinctions at the regional scale. Therefore, multiple stressors on inshore reefs on the GBR have resulted in a lower abundance of all corals rather than causing major shifts in assemblage structure. A least one cycle of recovery in abundance has occurred at all sites, with some sites experiencing up to three periods of recovery, suggesting the ecological processes necessary for recovery remain essentially intact. The poor status of these sites at the end of the study is therefore attributable mainly to the recent incidence of disturbances, especially cyclone Yasi (in February 2011), rather than systematic declines in the abundance of corals. Indeed, coral abundance at most sites was increasing until mass bleaching events in March 2016 and 2017 (unpublished data). Nonetheless, recent research suggests that the disturbance regime on reefs has transitioned into an era where climate change and other human-induced changes will predominate over natural disturbances (Hughes et al. 2017; Tan et al. 2018). Furthermore, the intensity of cyclones is predicted to increase in response to ongoing climate change (Knutson et al. 2010). This 15-year period might therefore be a guide to the future status of coral reefs.
Losers greatly outnumbered winners in response to bleaching. This is not surprising because the time interval between censuses was six months and therefore the opportunity to recruit into the sample population (i.e. colonies with a maximum diameter greater than 5 cm) is mostly limited to those species susceptible to fission, such as Platygyra (Babcock 1991) and Sarcophyton. Nonetheless, these results support recent findings that very few taxa are winners when bleaching events are severe (Hughes et al. 2017). In addition, traditional bleaching hierarchies based on a single census of bleaching status within populations during a bleaching event (e.g. Marshall and Baird 2000; McClanahan et al. 2004) do not reflect those based on mortality estimates from changes in abundance (Fig. 8). In particular, a number of taxa that rarely bleach, for example, Cyphastrea spp. and Alveopora spp. suffered high rates of mortality (Marshall and Baird 2000; McClanahan 2004; Fig. 8). These results suggest that some taxa that are susceptible to bleaching do not present with symptoms typical of bleaching, such as loss of symbionts and consequent paling of the colony. Accurate estimates of the effects of thermal anomalies on reefs, therefore, require individuals to be tagged and followed through time (e.g. Baird and Marshall 2002). The fact that very few taxa can cope with thermal stress is probably due to the fact that severe thermal anomalies in the ocean are a relatively recent phenomenon (Spalding and Brown 2015), and therefore, corals have not had the chance to adapt to this form of stress.
Few predictions of UAST with respect to how taxa should respond to stress and disturbance are supported by these data. While the two competitive-taxa, Acropora and Turbinaria, were among the winners at the site level during periods of recovery, winners also included multiple genera from the other groups (Table 6). Indeed, there was a large range of responses among taxa within most adaptive groups. For example, the stress-tolerant taxa Cyphastrea, Favia and Goniastrea were consistently among the biggest losers at many sites following bleaching (Fig. 5). Similarly, the same taxa responded in different ways to the same stress at different sites. For example, Montipora was consistently among the losers in response to bleaching, however, at SEP-D it was a winner (Tables 4, 5). This is evidence of marked response diversity within some species-rich genera (e.g., Montipora), suggesting that higher taxonomic groupings or adaptive groups are inappropriate for representing the vulnerability and responses of corals to disturbances and stresses. Indeed, there are few traits that are similar among species within most genera, in particular species-rich genera like the Acropora, Montipora and Porites (Madin et al. 2016). One prediction of UAST supported by these data is that it is not possible for organisms to adapt to a high frequency of both stress and disturbance (Grime and Pierce 2012). Indeed, there appear to be no true weedy species in the sessile anthozoa on the GBR. In other words, species capable of a rapid increase in abundance in response to disturbance are rare on coral reefs, unlike the numerous species of weeds in terrestrial environments (Grime and Pierce 2012).
The relative abundance of the adaptive strategies in the initial assemblages was not a good predictor of the trajectory of the assemblage in response to stress or multiple disturbances. Indeed, seven of the eight sites were equally degraded in the 15 years of the study despite large differences in initial assemblage structure. In particular, the Acropora dominated assemblages at southeast Pelorus were the least affected by bleaching (Fig. 3), at least with respect to changes in abundance. This is despite very high levels of mortality in tagged colonies of Acropora at southeast Pelorus (Baird and Marshall 2002). This again suggests that there are important differences in the response to bleaching among species within genera and categorising higher taxonomic groups to specific adaptive strategies is inappropriate.
A closer look at previous research also suggests that ASGs rarely behave as predicted (see also Zinke et al. 2018). For example, Darling et al. (2013) tested the response of ASGs to bleaching and fishing. In contrast to predictions, weedy species did not consistently benefit from disturbance, competitive species did not benefit from periods free of disturbance and the responses of stress tolerant species were context dependent (e.g., big declines were observed on unfished reefs, but no declines on fished reefs). The only response consistent with the theory was that competitors were more susceptible to the chronic disturbance in the form of fishing than stress-tolerant and weedy species. However, all the competitive species in this study were either branching Acropora spp. or Pocillopora spp. suggesting that classification of species based on morphology would have been equally as informative. Similarly, Graham et al. (2014) concluded that the relative abundance of ASGs was useful for distinguishing among reefs with a different disturbance history on the GBR. However, the only locally abundant species classified as competitors were Acropora spp. Therefore, these reefs could equally well have been distinguished by classifying taxa as Acropora vs non-Acropora. Sommer et al. (2014) also used an adaptive strategy scheme to compare the relative abundance of species in each group among coral assemblages along a high-latitude gradient in south-eastern Australia. The only clear trend was a decrease in the relative abundance of stress-tolerant species in coral assemblage at higher latitudes (Sommer et al. 2014) in contrast to the predictions of UAST, i.e. that stress-tolerant species should dominate in unproductive habitats, such as these high latitude marginal reefs. This suggested either the current ASGs scheme used on reef corals (e.g. Darling et al. 2012) is inappropriate, or the UAST, developed from plants, is unsuitable for colonel creatures, such as reef corals.
In conclusion, ASGs rarely behaved as predicted in reef corals, which we attribute to marked response diversity within higher taxonomic groups and broadly defined adaptive groups. We suggest a direct trait-based approach (e.g. Mcwilliam et al. 2018) will be more informative to understand differential vulnerabilities of corals to changing disturbance regimes, and to predict potential shifts in species composition.
References
Anthony KRN, Kerswell AP (2007) Coral mortality following extreme low tides and high solar radiation. Mar Biol 151:1623–1631. https://doi.org/10.1007/s00227-006-0573-0
Babcock RC (1991) Comparative demography of three species of scleractinian corals using age- and size-dependent classifications. Ecol Monogr 61:225–244. https://doi.org/10.2307/2937107
Bainbridge ZT, Wolanski E, Álvarez-Romero JG, Lewis SE, Brodie JE (2012) Fine sediment and nutrient dynamics related to particle size and floc formation in a Burdekin River flood plume, Australia. Mar Pollut Bull 65:236–248. https://doi.org/10.1016/j.marpolbul.2012.01.043
Baird AH, Marshall PA (1998) Mass bleaching of corals on the Great Barrier Reef. Coral Reefs 17:376. https://doi.org/10.1007/s003380050142
Baird AH, Marshall PA (2002) Mortality, growth and reproduction in scleractinian corals following bleaching on the Great Barrier Reef. Mar Ecol Prog Ser 237:133–141. https://doi.org/10.3354/meps237133
Baird AH, Marshall PA, Kuo C-Y, Pratchett MS (2020) Coral abundance on the inshore Great Barrier Reef 1998–2013. Zenodo. https://doi.org/10.5281/zenodo.4310394
Berkelmans R, De’ath G, Kininmonth S, Skirving WJ (2004) A comparison of the 1998 and 2002 coral bleaching events on the Great Barrier Reef: spatial correlation, patterns, and predictions. Coral Reefs 23:74–83. https://doi.org/10.1007/s00338-003-0353-y
Cohen J (1988) Statistical power analysis for the behavioral sciences. Lawrence Erlbaum Associates, Hillsdale, New Jersey
Darling ES, Alvarez-Filip L, Oliver TA, McClanahan TR, Côté IM (2012) Evaluating life-history strategies of reef corals from species traits. Ecol Lett 15:1378–1386. https://doi.org/10.1111/j.1461-0248.2012.01861.x
Darling ES, McClanahan TR, Côté IM (2013) Life histories predict coral community disassembly under multiple stressors. Glob Change Biol 19:1930–1940. https://doi.org/10.1111/gcb.12191
Fabricius K, Alderslade P (2001) Soft corals and sea fans: a comprehensive guide to the tropical shallow-water genera of the Central-West Pacific, the Indian Ocean and the Red Sea. Australian Institute of Marine Science, Townsville, Australia
Graham NAJ, Chong-Seng KM, Huchery C, Januchowski-Hartley FA, Nash KL (2014) Coral reef community composition in the context of disturbance history on the Great Barrier Reef, Australia. PLoS ONE 9:e101204. https://doi.org/10.1371/journal.pone.0101204
Grime JP, Pierce S (2012) The evolutionary strategies that shape ecosystems. Wiley-Blackwell, Chichester, West Sussex, UK ; Hoboken, NJ
Haapkylä J, Unsworth RKF, Flavell M, Bourne DG, Schaffelke B, Willis BL (2011) Seasonal rainfall and runoff promote coral disease on an inshore reef. PLoS ONE 6:e16893. https://doi.org/10.1371/journal.pone.0016893
Hughes TP, Kerry J, Álvarez-Noriega M, Álvarez-Romero J, Anderson K, Baird AH, Babcock R, Beger M, Bellwood D, Berkelmans R, Bridge T, Butler I, Byrne M, Cantin N, Comeau S, Connolly S, Cumming G, Dalton S, Diaz-Pulido G, Eakin CM, Figueira W, Gilmour J, Harrison H, Heron S, Hoey AS, Hobbs J-P, Hoogenboom M, Kennedy E, Kuo C-Y, Lough J, Lowe R, Liu G, Malcolm McCulloch HM, McWilliam M, Pandolfi J, Pears R, Pratchett MS, Schoepf V, Simpson T, Skirving W, Sommer B, Torda G, Wachenfeld D, Willis B, Wilson S (2017) Global warming and recurrent mass bleaching of corals. Nature 543:373–378. https://doi.org/10.1038/nature21707
Hughes TP, Kerry JT, Baird AH, Connolly SR, Dietzel A, Eakin CM, Heron SF, Hoey AS, Hoogenboom MO, Liu G, McWilliam MJ, Pears RJ, Pratchett MS, Skirving WJ, Stella JS, Torda G (2018) Global warming transforms coral reef assemblages. Nature 556:492–496. https://doi.org/10.1038/s41586-018-0041-2
Knutson TR, McBride JL, Chan J, Emanuel K, Holland G, Landsea C, Held I, Kossin JP, Srivastava AK, Sugi M (2010) Tropical cyclones and climate change. Nat Geosci 3:157–163. https://doi.org/10.1038/ngeo779
Lukoschek V, Cross P, Torda G, Zimmerman R, Willis BL (2013) The importance of coral larval recruitment for the recovery of reefs impacted by cyclone Yasi in the central Great Barrier Reef. PLoS ONE 8:e65363. https://doi.org/10.1371/journal.pone.0065363
Madin JS, Anderson KD, Andreasen MH, Bridge TCL, Cairns SD, Connolly SR, Darling ES, Diaz M, Falster DS, Franklin EC, Gates RD, Hoogenboom MO, Huang D, Keith SA, Kosnik MA, Kuo C-Y, Lough JM, Lovelock CE, Luiz O, Martinelli J, Mizerek T, Pandolfi JM, Pochon X, Pratchett MS, Putnam HM, Roberts TE, Stat M, Wallace CC, Widman E, Baird AH (2016) The Coral Trait Database, a curated database of trait information for coral species from the global oceans. Sci Data 3:160017. https://doi.org/10.1038/sdata.2016.17
Madin JS, Baird AH, Dornelas M, Connolly SR (2014) Mechanical vulnerability explains size-dependent mortality of reef corals. Ecol Lett 17:1008–1015. https://doi.org/10.1111/ele.12306
Marshall PA, Baird AH (2000) Bleaching of corals on the Great Barrier Reef: differential susceptibilities among taxa. Coral Reefs 19:155–163. https://doi.org/10.1007/s003380000086
McClanahan TR (2004) The relationship between bleaching and mortality of common corals. Mar Biol 144:1239–1245. https://doi.org/10.1007/s00227-003-1271-9
McClanahan TR, Baird AH, Marshall PA, Toscano MA (2004) Comparing bleaching and mortality responses of hard corals between southern Kenya and the Great Barrier Reef, Australia. Mar Pollut Bull 48:327–335. https://doi.org/10.1016/j.marpolbul.2003.08.024
McWilliam M, Hoogenboom MO, Baird AH, Kuo C-Y, Madin JS, Hughes TP (2018) Biogeographical disparity in the functional diversity and redundancy of corals. Proc Natl Acad Sci 115:3084–3089. https://doi.org/10.1073/pnas.1716643115
McWilliam M, Pratchett MS, Hoogenboom MO, Hughes TP (2020) Deficits in functional trait diversity following recovery on coral reefs. Proc R Soc B 287:20192628. https://doi.org/10.1098/rspb.2019.2628
Pratchett MS, Hoey AS, Wilson SK (2014) Reef degradation and the loss of critical ecosystem goods and services provided by coral reef fishes. Curr Opin Environ Sustain 7:37–43. https://doi.org/10.1016/j.cosust.2013.11.022
Pratchett MS, McWilliam MJ, Riegl B (2020) Contrasting shifts in coral assemblages with increasing disturbances. Coral Reefs 39:783–793. https://doi.org/10.1007/s00338-020-01936-4
Sommer B, Harrison PL, Beger M, Pandolfi JM (2014) Trait-mediated environmental filtering drives assembly at biogeographic transition zones. Ecology 95:1000–1009. https://doi.org/10.1890/13-1445.1
Spalding MD, Brown BE (2015) Warm-water coral reefs and climate change. Science 350:769–771. https://doi.org/10.1126/science.aad0349
Tan CH, Pratchett MS, Bay LK, Graham EM, Baird AH (2018) Biennium horribile: very high mortality in the reef coral Acropora millepora on the Great Barrier Reef in 2009 and 2010. Mar Ecol Prog Ser 604:133–142. https://doi.org/10.3354/meps12750
Torda G, Sambrook K, Cross P, Sato Y, Bourne DG, Lukoschek V, Hill T, Torras Jorda G, Moya A, Willis BL (2018) Decadal erosion of coral assemblages by multiple disturbances in the Palm Islands, central Great Barrier Reef. Sci Rep 8:11885. https://doi.org/10.1038/s41598-018-29608-y
Van Woesik R, De Vantier L, Glazebrook J (1995) Effects of cyclone “Joy” on nearshore coral communities of the Great Barrier Reef. Mar Ecol Prog Ser 128:261–270. https://doi.org/10.3354/meps128261
Veron JEN (2000) Corals of the world. Australian Institute of Marine Science, Townsville, Australia
Zinke J, Gilmour JP, Fisher R, Puotinen M, Maina J, Darling E, Stat M, Richards ZT, McClanahan TR, Beger M, Moore C, Graham NAJ, Feng M, Hobbs J-PA, Evans SN, Field S, Shedrawi G, Babcock RC, Wilson SK (2018) Gradients of disturbance and environmental conditions shape coral community structure for south-eastern Indian Ocean reefs. Divers Distrib 24:605–620. https://doi.org/10.1111/ddi.12714
Acknowledgements
The research was funded by Australian Geographic, an Australian Research Council (ARC) Future Fellowship to AHB (FT0990652) and the ARC Centre of Excellence for Coral Reef Studies (CE0561432 & CE140100020). We thank Paul Marshall and numerous volunteers for help in the field.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
On behalf of all authors, the corresponding author states that there is no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kuo, CY., Pratchett, M.S., Madin, J.S. et al. A test of adaptive strategy theory using fifteen years of change in coral abundance. Coral Reefs 42, 951–966 (2023). https://doi.org/10.1007/s00338-023-02399-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00338-023-02399-z