
Abstract
Mesophotic coral ecosystems (MCEs) represent the lowest depth distribution inhabited by many coral reef-associated organisms. Research on fishes associated with MCEs is sparse, leading to a critical lack of knowledge of how reef fish found at mesophotic depths may vary from their shallow reef conspecifics. We investigated intraspecific variability in body condition and growth of three Hawaiian endemics collected from shallow, photic reefs (5–33 m deep) and MCEs (40–75 m) throughout the Hawaiian Archipelago and Johnston Atoll: the detritivorous goldring surgeonfish, Ctenochaetus strigosus, and the planktivorous threespot chromis, Chromis verater, and Hawaiian dascyllus, Dascyllus albisella. Estimates of body condition and size-at-age varied between shallow and mesophotic depths; however, these demographic differences were outweighed by the magnitude of variability found across the latitudinal gradient of locations sampled within the Central Pacific. Body condition and maximum body size were lowest in samples collected from shallow and mesophotic Johnston Atoll sites, with no difference occurring between depths. Samples from the Northwestern Hawaiian Islands tended to have the highest body condition and reached the largest body sizes, with differences between shallow and mesophotic sites highly variable among species. The findings of this study support newly emerging research demonstrating intraspecific variability in the life history of coral-reef fish species whose distributions span shallow and mesophotic reefs. This suggests not only that the conservation and fisheries management should take into consideration differences in the life histories of reef-fish populations across spatial scales, but also that information derived from studies of shallow fishes be applied with caution to conspecific populations in mesophotic coral environments.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Coral-reef research has been historically focused on the organisms and processes in shallow water habitats accessible to snorkelers and SCUBA divers. Beyond these depths, mesophotic coral ecosystems (MCEs) are prevalent in tropical and subtropical waters worldwide, ranging from 30 to 150 m or deeper (Hinderstein et al. 2010). These light-dependent coral-reef communities are composed of a variety of corals, algae, and sponges in benthic communities supporting reef fishes and mobile invertebrates (Hinderstein et al. 2010). Compared to shallow coral reefs, MCEs experience lower light, seawater temperature, and wave stress (Kahng et al. 2010; Rooney et al. 2010; Franklin et al. 2013). Despite the differences in environmental conditions between shallow and mesophotic reefs, many of the flora and fauna that characterize shallow reefs are also found inhabiting the upper reaches of MCEs (Rooney et al. 2010; Bridge et al. 2011; Wagner et al. 2014). While changes in species composition from shallow reefs to MCEs have been characterized in several locations globally (Baker et al. 2016), there has been little examination of intraspecific traits that might vary across the range of environmental conditions experienced along this depth gradient, and how these traits of the species at mesophotic depths may vary across populations.
As shallow coral reefs continue to decline globally due to the effects of climate change (increasing seawater temperatures and ocean acidification) and other anthropogenic activities (e.g., overfishing and pollution) (Kleypas et al. 1999; Hoegh-Guldberg et al. 2007; De’ath et al. 2009), a better understanding of the similarities and differences in ecological structure and function between MCEs and shallow coral reefs has become increasingly important. The “deep-reef refugia hypothesis”, first introduced by Glynn (1996), suggests that MCEs are more stable environments than shallow coral reefs and may become vital refuges from environmental stressors as well as sources of replenishment for shallow coral-reef species (Bongaerts et al. 2010). Due to the presence of shallow coral-reef species at mesophotic depths, a level of biological and physical connectivity between these two ecosystems can be postulated (Hinderstein et al. 2010). However, it is unclear whether or how individual species may differ biologically between shallow and mesophotic reefs. Increasing the understanding of population ecology within MCEs by studying the life-history traits of organisms that inhabit these remote environments has been emphasized as a research priority (Puglise et al. 2009; Kahng et al. 2010).
Factors such as temperature, habitat, prey availability, and predation intensity influence intraspecific variability in the life-history characteristics of many reef fishes (Gust et al. 2002; Ruttenberg et al. 2005). For reef-fish species with wide ranges, prior life-history investigations have examined geographic variability among shallow reefs but typically neglected to address intraspecific variability across broad depth gradients that include MCEs. In shallow ecosystems, reef fishes have been shown to exhibit significant plasticity in demographic traits across scales ranging from tens to hundreds of kilometers (Gust et al. 2002; Ruttenberg et al. 2005; Trip et al. 2008). Therefore, intraspecific variability is also likely to be observed not only between shallow and mesophotic populations, but also across mesophotic populations that are spatially separate. The few studies that have examined the effects of depth on life history have shown that there are differences in life-history traits among populations of fishes across depth ranges in overall shallow environments (<10–39 m) (Hoey et al. 2007; Srinivasan 2003) and across a depth range encompassing both shallow and mesophotic environments (<10–70 m) (Goldstein et al. 2016). The only existing study to compare the life-history traits of a tropical coral-reef fish between shallow and mesophotic environments (Goldstein et al. 2016) suggested that species inhabiting both shallow coral reefs and MCEs may potentially exhibit intraspecific variability in their life-history traits, thereby acclimating and/or adapting to inherently different environments. Reef fish exhibit intraspecific phenotypic plasticity in visual acuity, with individuals at lower limits of their depth range able to adapt to lower light levels (Brokovich et al. 2010), which may influence their persistence at these depths, and potentially, intraspecific variability in life-history traits. However, given that the work by Goldstein et al. (2016) comparing shallow and mesophotic populations only focused on a single species and was conducted within a roughly 200 km radius, further work is warranted to explore how the life-history traits of other tropical coral-reef fish with broad depth distributions vary.
To examine intraspecific variability of fishes across their depth range, we investigated the life-history traits of three species of coral-reef fishes that are endemic to the Hawaiian Archipelago and Johnston Atoll, all three with broad depth distributions and varying diets: Ctenochaetus strigosus, Chromis verater and Dascyllus albisella. While Ct. strigosus grazes on benthic detritus, diatoms, and bacteria (Jones 1968) and is commonly found in shallow reef areas, this species has also been observed at a depth of 113 m (Randall and Clements 2001). The damselfishes Ch. verater and D. albisella are found in both shallow coral reefs and deeper mesophotic habitats (≥55 m deep) (Stevenson 1963; Mundy 2005). Chromis verater and D. albisella are both planktivorous species, feeding primarily on copepods (Stevenson 1963; Swerdloff 1970). In this study, we tested the hypothesis that there would be intraspecific variability reflecting environmental gradients across depths by comparing age-based life-history traits and estimates of body condition of the three reef-fish species between shallow and mesophotic depths across the extent of their geographic ranges.
Materials and methods
Study site and sample collections
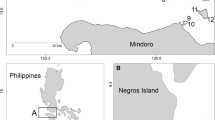
A total of 169 Ch. verater (Jordan and Metz, 1912), 282 Ct. strigosus (Bennett, 1828), and 162 D. albisella (Gill, 1862) were collected on research cruises throughout the Northwestern Hawaiian Islands (NWHI), Main Hawaiian Islands (MHI), and Johnston Atoll from September 2012 to November 2015 (Fig. 1). Shallow and mesophotic (hereafter referred to as “deep”) samples were collected by divers using pole spears or hand nets. Divers collecting in shallow sites (<40 m) used open-circuit breathing systems, while divers collecting in deep sites (≥40 m deep) used closed-circuit rebreather systems. Sampling effort was considerably higher within shallow sites due to logistical limitations of rebreather diving that restrict bottom time. To reduce bias in the collection, divers endeavored to collect representative samples across the observed size range per species. The length of Ch. verater specimens ranged from 68 to 156 mm standard length (SL) in shallow locations and 41 to 152 mm SL in deep locations (Table 1). Ctenochaetus strigosus specimens ranged from 65 to 176 mm in shallow locations and 60 to 165 mm in deep locations (Table 1). Dascyllus albisella specimens ranged from 39 to 98 mm in shallow locations and 14 to 101 mm in deep locations (Table 1). All fish were stored frozen until dissections were performed.

Collection sites of Chromis verater, Ctenochaetus strigosus, and Dascyllus albisella in the Hawaiian Archipelago
Dissections and otolith preparation
Specimens whose otoliths were damaged by spearing were not used for analysis. Morphometric measurements were taken for 158, 275, and 155 specimens of Ch. verater, Ct. strigosus, and D. albisella, respectively, which were then dissected and aged (Table 1). SL was measured to the nearest mm, and whole body weights were recorded to the nearest g of each thawed specimen. Sagittal otoliths were removed, cleaned of residual material, rinsed in 95% ethanol, and air-dried. Once dry, otoliths were weighed (to 0.1 mg), photographed, and stored dry for aging later.
The sagittal otoliths were prepared following the methodology of Choat et al. (2003). Unless missing or damaged, the left otolith of each specimen was processed. Each otolith was mounted on the edge of a glass microscopy slide with thermoplastic glue (CrystalBond) and grounded to the nucleus with a 1200 grit diamond lap on a CrystalMaster 8 grinding wheel. The otolith was then remounted so that the ground side of the otolith was flush with the slide, and the other side of the otolith was ground down until a thin transverse section was attained. The resulting section was finally hand-polished using 3–9-μm grit aluminum oxide lapping film and coated with a layer of CrystalBond for improved clarity of growth increments.
Each sectioned otolith was examined under transmitted light with a low-power dissecting microscope (10–40× magnification). Growth increments have been shown to be deposited annually for several tropical pomacentrids (e.g., Meekan et al. 1999) as well as acanthurids (e.g., Choat and Axe 1996), so they were assumed to be deposited annually for Ch. verater, Ct. strigosus, and D. albisella. Pairs of dark opaque and light translucent bands assumed to be the annual growth increments were counted along a consistent axis by two independent observers (MW and BT). If the two counts varied, each observer independently recounted until an exact agreement was made. If the observers could not reach a consensus after each counting three times, that otolith was excluded from analysis. Otoliths of individuals less than 1 yr old were subsequently reground until daily growth increments were visible and reanalyzed to estimate daily ages. Daily ages were converted into fractions of a year to be incorporated into growth curves.
As an evaluation of aging estimates, the otolith weight was regressed on estimated age using an ordinary linear regression, given that a positive relationship would indicate continual growth of otoliths through the accretion of calcium carbonate during a fish’s lifespan (Fowler and Doherty 1992; Choat and Axe 1996).
Analysis of life-history traits
Body condition
Fulton’s body condition index (also known as Fulton’s K) was calculated for individual samples using the following equation:
where W is total weight (g), and L is standard length (cm). Fulton’s K provides a morphometric index of fish “bulk,” or body condition (Fulton 1904; Ricker 1975). For each species, a two-way analysis of variance (ANOVA) of body condition as a function of region, depth, and their interaction was performed. Assumptions of ANOVAs were evaluated for each test using standard diagnostic practices.
Growth
The von Bertalanffy growth function (VBGF), a conventional method of describing growth in fishes (von Bertalanffy 1938), was fit to size-at-age data for each species using nonlinear least squares techniques:
where \( L_{t} \) is length at age t (yr), \( L_{\infty } \) is the mean asymptotic length, K is the growth coefficient, and L 0 is the length at age zero (i.e., size at settlement). For each species, multiple VBGFs were fit to compare the asymptotic length and growth coefficient between shallow and deep samples across regions. However, due to the low sample size of Ct. strigosus and D. albisella (two and seven specimens, respectively) from the deep MHI, there were no growth curves fit for these species at this region and depth. Growth curves were fit by constraining the y-intercept (i.e., \( L_{0} \)) to a common length at settlement using the original form of the von Bertalanffy equation (von Bertalanffy 1938) for each species to increase the accuracy of VBGF parameter estimates (Kritzer et al. 2001). Length at settlement of Ct. strigosus and D. albisella was obtained from the existing literature and set as 28 mm (Randall 1955; Sancho et al. 1997) and 10 mm (Booth 1992), respectively. Settlement size for Ch. verater has not been published; therefore, the length at settlement of 19 mm for the congeneric Ch. multilineata (Wellington and Robertson 2001) was used, given that these two species attain similar maximum lengths. Estimates of the VBGF parameters were obtained from nonlinear least squares regression of the best-fit models and used to generate growth curves for each species. The age at which the species has zero length, t 0, was estimated for each VBGF set of \( L_{\infty } \) and K parameters by substituting into the more common form of the equation (Ricker 1975):
To visually compare parameter estimates, 95% confidence ellipses were constructed around maximum likelihood estimates of \( L_{\infty } \) and K following methods of Kimura (1980). Likelihood ratio tests were used to test for differences in \( L_{\infty } \) and K between depths within regions for each species using a procedure similar to that outlined in Kimura (1980) and Haddon (2011). For comparisons across species–region data sets, maximum likelihood estimates of common or different \( L_{\infty } \) and K for each combination were calculated with a likelihood ratio test used to test the null hypotheses for each set of parameter constraints. All data manipulation and statistical analyses described in this section were conducted using R v3.1.1 (R Development Core Team 2009).
Results
Body condition
Body condition (Fulton’s K) differed significantly between depths (p < 0.01) and among regions (p < 0.05) for Ct. strigosus, with a significant interaction effect (p < 0.05). For Ch. verater and D. albisella, body condition was not related to depth, but was significantly different among regions (p < 0.001) (Electronic supplementary material, ESM, Table S1). For each species, body condition was significantly lowest in samples collected at Johnston Atoll (Fig. 2). In fishes collected from shallow locations, the body condition of each species generally appeared to be lowest at Johnston Atoll and highest in either the NWHI or MHI. Body condition in fishes from deep locations was variable among species, with no clear emerging trend (Fig. 2).

Comparison of Fulton’s condition index (K, g m−3) for Chromis verater (a), Ctenochaetus strigosus (b), and Dascyllus albisella (c) in shallow coral reefs (light blue) and mesophotic coral ecosystems (dark blue) at Johnston Atoll (JA), the Main Hawaiian Islands (MHI), and the Northwestern Hawaiian Islands (NWHI). Boxes indicate the interquartile range of Fulton’s condition index, with the median shown by horizontal lines, minimum and maximum values shown by whiskers, and points representing outliers
Chromis verater from the shallow MHI had higher body condition than conspecifics collected from deep Johnston Atoll (Fig. 2a). Body condition was higher in fishes from both the shallow and deep NWHI than in fishes from both shallow and deep Johnston Atoll sites, and in fishes collected from the deep MHI. Body condition did not differ between depths within regions.
The body condition of Ct. strigosus was highest in fishes from the shallow NWHI (Fig. 2b). Even though Ct. strigosus from the deep NWHI had lower body condition than shallow conspecifics from the NWHI, there was no difference in body condition between fishes from shallow and deep sites at Johnston Atoll. The low sample size of Ct. strigosus from the deep MHI did not allow for comparisons at this location for this species.
Dascyllus albisella from the shallow MHI had higher body condition than conspecifics from shallow and deep sites in Johnston Atoll, and from the deep NWHI (Fig. 2c). Body condition did not differ between fishes from shallow and deep sites within any region, but the low sample size of D. albisella from the deep MHI did not permit this location to be incorporated into the comparisons.
Growth
The growth curves for all three species were asymptotic, with initial rapid growth followed by slower growth, typical of fishes. As a validation of age estimates, otolith weight was a good predictor of age for Ch. verater (r 2 = 0.70, F 1,150 = 341.1, p < 0.001), Ct. strigosus (r 2 = 0.76, F 1,237 = 750.9, p < 0.001), and D. albisella (r 2 = 0.79, F 1,144 = 538.4, p < 0.001) (ESM Fig. S1). Ctenochaetus strigosus was the longest-lived species in this study; the oldest individual, at 39 yr, surpassed previous age estimations for this species (Langston et al. 2009). Length-at-age relationships were highly variable due to the observed variability in growth among individuals at each location (Table 2; Figs. 3, 4). Intraspecific variability in growth, with trends unique to each species, occurred between locations.

Von Bertalanffy growth functions fitted to Chromis verater (a, b, c), Ctenochaetus strigosus (d, e, f), and Dascyllus albisella (g, h, i) size-at-age data from samples collected from shallow coral reefs (light blue) and mesophotic coral ecosystems (dark blue) at Johnston Atoll (a, d, g), the Main Hawaiian Islands (b, e, h), and the Northwestern Hawaiian Islands (c, f, i)

Plots of 95% confidence ellipses for the mean asymptotic length (\( L_{\infty } \)) and the growth coefficient (k) by location and depth for Chromis verater (a), Ctenochaetus strigosus (b), and Dascyllus albisella (c)
Chromis verater from shallow habitats at Johnston Atoll had faster juvenile growth and reached a smaller asymptotic length than individuals from deep habitats at Johnston Atoll, but a similar pattern was not observed in the Hawaiian Archipelago (Fig. 3a–c). The largest estimated asymptotic length of Ch. verater was in fishes from the shallow NWHI (Table 2). Chromis verater sampled in the MHI and NWHI from deep sites grew more quickly at younger ages than their conspecifics sampled from shallow sites (Fig. 3a–c). However, the growth model for the shallow MHI fishes may not accurately represent this population because of the lack of young individuals (<3 yr old) collected. The 95% confidence ellipses surrounding the VBGF parameters \( L_{\infty } \) and K did not overlap between estimates obtained for the shallow and deep NWHI fishes, which suggests growth patterns in the NWHI differed significantly between shallow and deep populations (Table 2; Fig. 4a). Likelihood ratio tests identified significant differences between \( L_{\infty } \) and K for Ch. verater between shallow and deep habitats in NWHI (ESM Table S2). While the confidence ellipses for the shallow and deep Johnston Atoll fishes overlapped, these ellipses were separate from all others constructed for the NWHI and MHI fishes (Fig. 4a). Thus, both shallow and deep Ch. verater from Johnston Atoll reached a significantly smaller asymptotic length at a generally faster rate than most fishes from the Hawaiian Archipelago.
Deep Ct. strigosus from the NWHI reached the largest asymptotic length (Table 2), yet individuals older than 10 yr were rare at this location. The smallest asymptotic length estimated for Ct. strigosus was from shallow Johnston Atoll sites (Table 2). The non-overlapping 95% confidence ellipses surrounding \( L_{\infty } \) and K for shallow and deep NWHI fishes demonstrated a significant difference in their growth patterns (Fig. 4b). Likelihood ratio tests identified significant differences in \( L_{\infty } \) for Ct. strigosus between shallow and deep habitats in NWHI (ESM Table S2). The confidence ellipses constructed for fishes from shallow sites did not overlap, revealing that fish from the MHI reached the largest asymptotic length while fish from Johnston Atoll reached the smallest. The confidence ellipses of shallow and deep Ct. strigosus from Johnston Atoll did overlap, largely due to their similarity in \( L_{\infty } \), but these growth patterns were distinct from the rest of the Hawaiian Archipelago fishes.
There were no significant differences in the growth patterns of D. albisella sampled from shallow and deep sites at Johnston Atoll (Table 2; Figs. 3g, 4c). Dascyllus albisella from the shallow NWHI had more rapid juvenile growth and reached a larger asymptotic length than those from the deep NWHI (Table 2; Fig. 3i), and the non-overlapping confidence ellipses provided evidence that these growth patterns were distinct (Fig. 4c). Likelihood ratio tests identified significant differences between \( L_{\infty } \) and K for D. albisella between shallow and deep habitats in NWHI (ESM Table S2). Unlike Ch. verater and Ct. strigosus, the growth patterns of D. albisella from Johnston Atoll were not distinct from those collected from the Hawaiian Archipelago. The largest mean asymptotic sizes were found for individuals of D. albisella from the shallow locations at the NWHI and Johnston Atoll (Table 2). While the smallest size was found for D. albisella from the shallow MHI, all the fishes from this location were found to reach an age of 5 yr or less.
Discussion
Prior studies have investigated intraspecific variability in the demographic traits of coral-reef fishes, but they primarily examined differences across geographic scales at shallow depths (Gust et al. 2002; Ruttenberg et al. 2005). The present study not only presents patterns of intraspecific variability in the growth and condition of Ch. verater, Ct. strigosus, and D. albisella across the extent of their geographic ranges, but also demonstrates previously undocumented differences between populations inhabiting shallow coral reefs and MCEs. This study demonstrates that there is considerable variability in patterns of life history among species; therefore, caution should be applied in generalizing these patterns to other reef fishes with broad depth ranges. These results emphasize the need to further investigate variability in the life history of fishes whose distributions span shallow coral reefs and MCEs.
The variability in life-history traits across space and depth is likely influenced by multiple environmental and ecological factors. Given that reef fish exhibit significant intraspecific plasticity in demographic traits across broad spatial scales (Gust et al. 2002; Ruttenberg et al. 2005; Trip et al. 2008), conclusions drawn from comparisons between pooled shallow and deep samples may not accurately represent populations in different regions. Quantifying the differences in life-history traits between shallow and mesophotic reef-fish populations based purely on pooled results requires broad generalizations; examining variability in life history at a finer scale is warranted. Moreover, there were unique differences in life-history patterns among species, indicative of an interaction of species-specific drivers (such as habitat preference, behavior, and morphology) with broad-scale environmental factors.
Geographic variability
Several trends in life-history characteristics demonstrated differences within and among the three geographic regions. Body condition generally increased with latitude, with the lowest body condition for all species found at Johnston Atoll and the highest body condition found in the NWHI (Ct. strigosus and Ch. verater) or the MHI (D. albisella). Body condition, estimated using Fulton’s K, is defined as a measure of energy reserves relative to body size that represents a measure of overall well-being. Body condition can represent a proxy for how favorable the surrounding environment is for an organism. A habitat with low food availability, high competition for limiting resources, and/or high predation risk is innately unfavorable; thus, body condition is significantly related to these environmental factors (Page et al. 2007; Donelson et al. 2008). The observed increase in body condition with increasing latitude may be associated with the increase in productivity along this gradient. Mean annual primary productivity, using surface chlorophyll-a concentrations as a proxy, was higher in the NWHI (average of 0.1 mg m−2) than the MHI and Johnston Atoll (average of 0.08 and 0.05 mg m−2, respectively) (Gove et al. 2013). Low levels of primary productivity directly influence feeding resources for many coral-reef fish species, and food availability is positively related to body condition (Page et al. 2007; Donelson et al. 2008). Furthermore, under conditions with limited food, decreased body condition in females results largely from increased energy allocation to reproduction. Females under conditions with abundant food sources do not sacrifice their body condition during reproductive periods and also reproduce more frequently with larger and more numerous eggs that lead to larger juveniles on average. Moreover, juvenile fishes in better condition are reported to have higher levels of survivorship on coral reefs (Booth and Hixon 1999; Booth and Alquezar 2002).
Maximum length increased with latitude in Ct. strigosus and Ch. verater, though no consistent trends in growth rate emerged across this gradient. Colder waters associated with increases in latitude often result in increased body sizes of tropical and subtropical reef fishes (e.g., Luckhurst et al. 2000; Choat and Robertson 2002). While sea surface temperatures can drop as low as 16 °C at certain locations in the NWHI (Kane et al. 2014), Johnston Atoll experiences sea surface temperatures of 25–27 °C with little seasonal variability due to its proximity to the equator (Ralston et al. 1986; Boehlert et al. 1992). For all species, growth patterns did not differ between shallow and deep sites at Johnston Atoll. This result may similarly be driven by temperature; while the thermocline is found in waters 60 m or shallower in certain locations in the NWHI (Grigg et al. 2008), further south the thermocline becomes deeper, ranging from 100 to 400 m at Johnston Atoll (US Army Corps of Engineers 1983). At such a depth at Johnston Atoll, MCEs are less likely to experience substantially lower temperatures than found in shallow coral reefs. Hence, the temperature difference between MCEs and shallow coral reefs at Johnston Atoll may not be strong enough to drive variability in the growth rate of reef fishes found at both depths.
While the maximum length of Ct. strigosus and Ch. verater increased along the latitudinal gradient from Johnston Atoll to the NWHI, D. albisella exhibited a different trend in body size. Shallow samples of D. albisella were found to obtain larger body sizes in Johnston Atoll and the NWHI, where the percentage of hard coral cover and substrate height is higher than the MHI (Williams et al. 2015). This trend may be associated with this species’ site-attached nature, with larger body sizes reached in locations with higher shelter availability. Larval D. albisella are known to settle primarily on coral heads (Stevenson 1963; Groll 1984; Booth 1992). Even the distribution of juvenile D. albisella depends on the distribution of corals (Stevenson 1963), and in conspecific groups, this species remains closely associated with the benthos until becoming sexually mature (Booth 1991, 1992). As adults, D. albisella swim higher in the water column feeding on plankton, yet individuals of all ages will retreat into coral heads when they become alarmed (Stevenson 1963). The benthic surroundings of D. albisella that occupy habitats with greater potential refuge availability may allow them to avoid predation and facilitate growth to larger body sizes.
Species-specific variability
The NWHI was the only region examined in this study where significantly different life-history characteristics were found between shallow and mesophotic populations. While intraspecific variability in growth and condition occurred between depths for Ct. strigosus, Ch. verater, and D. albisella, differences in life-history traits were also evident among the three species.
Ctenochaetus strigosus attained larger body sizes at mesophotic depths in the NWHI, but had slower growth and lower body condition than their conspecifics at shallow depths in the NWHI. It is unlikely that this variability is driven by competition for limited resources, given the low density of benthic grazers found in MCEs and potentially similar food availability between depths. In the NWHI, the density of benthic grazers peaks at depths less than 10 m (Fukunaga et al. 2016), resulting in fewer potential competitors of Ct. strigosus in deeper habitats. Despite the decrease in light irradiance with depth, upwelling of cold, nutrient-rich waters to MCEs can support diverse and productive algae and phytoplankton communities at these depths (Leichter and Genovese 2006; Lesser 2006; Spalding 2012), thereby enhancing primary productivity and increasing resource availability. Low competition for abundant resources, along with colder water temperatures, may facilitate the growth of Ct. strigosus to larger maximum sizes in the MCEs of the NWHI. The substantially colder water temperatures found at mesophotic depths here may also affect the growth rate and body condition of Ct. strigosus. Decreasing temperatures reduce growth rates (Atkinson and Sibly 1997; Pauly 1998), and in these cold waters, Ct. strigosus may be approaching their thermal tolerance limit, lowering their body condition.
Mesophotic samples of D. albisella grew more slowly and attained smaller body sizes than shallow samples in the NWHI, which may be driven by lower shelter availability and heightened predation intensity. In the NWHI, the abundance of scleractinian corals decreases with depth (C Chang pers. comm.), while the density of piscivores peaks between 50 and 60 m depth (Fukunaga et al. 2016). Given that D. albisella also reached smaller sizes in regions with lower coral cover, the difference between shallow and deep samples in the NWHI is consistent with the hypothesis that shelter availability is a principal driver of the life history of this site-attached species. Dascyllus albisella is more vulnerable to predation in mesophotic depths in the NWHI not only because there is less shelter available, but also due to the increase in the abundance of piscivores. Increased vigilance for predators slows growth (Holbrook and Schmitt 1988; Werner et al. 1983), and decreased asymptotic size may be driven by the increased risk of mortality due to predation (Choat and Robertson 2002).
Unlike Ct. strigosus and D. albisella, shallow and deep Ch. verater samples from the NWHI attained similar maximum sizes, but deep samples exhibited more rapid initial growth than shallow samples. Though defined as depth generalists, Ch. verater are typically sparse in shallow waters and are more abundant at depths >18 m (Swerdloff 1970; Randall 1998), dominating deep reefs in the North Central Pacific (Brock and Chamberlain 1968; Wagner et al. 2014; Fukunaga et al. 2016). Moreover, species can adapt to colder temperatures by increasing their growth rates (Conover and Present 1990). Given that fast growth rates may enhance the survival of newly settled reef fishes, Ch. verater may be better adapted to life in the mesophotic zone than Ct. strigosus and D. albisella.
While Ch. verater and D. albisella both feed on zooplankton, their patterns of life history differ. Not only are Ch. verater more abundant in MCEs than D. albisella (which are most common at depths from 15 to 24 m; Stevenson 1963), these species have different morphologies that may influence their success at relative depths. Regardless of depth, both species feed in the water column, where there is a high flow rate of plankton. However, fishes swimming through the water column are exposed and vulnerable to predation. In the low-light environment of MCEs where there are high densities of piscivores, Ch. verater may have an advantage over D. albisella due to their differing caudal fin morphologies. Chromis verater have a forked tail with a high aspect ratio, which facilitates rapid acceleration, while D. albisella have a rounded tail with a low aspect ratio, which serves to improve maneuverability (Bridge et al. 2016). Chromis verater are likely to be able to flee faster from predators, while D. albisella can better maneuver into tight refuge spaces. However, in certain mesophotic environments where shelter availability is lower, such as the NWHI, improved maneuverability may not be as beneficial to D. albisella as faster acceleration is to Ch. verater. Therefore, the variability in life-history characteristics and preferred depth distributions of these planktivorous species may be related to differing morphological traits.
Beyond factors such as diet, morphology, and behavior that influence differences in life-history patterns among species, the myriad of varying environmental factors between shallow and mesophotic depths and along the latitudinal gradient may have influenced intraspecific variability in the life-history traits of the coral-reef fishes examined in this study. Given the confounding nature of many of these factors, future work should incorporate accurate measurements of reef productivity and examine the processes of competition and predation on mesophotic reefs. Moreover, this study presents results for only three of the many coral-reef fishes that have broad depth distributions. Given the logistical difficulty in collecting fish at mesophotic depths, the study provides a preliminary examination of differences in life-history traits between shallow and deep reef-fish populations based on limited sample sizes. The results of this study contrast slightly with the results of the only other previous work on the influence of depth (encompassing shallow and mesophotic reefs) on life-history traits, where individuals of the damselfish Stegastes partitus on the Florida Shelf attained larger maximum body sizes and older ages in deeper, mesophotic habitats than their shallow water conspecifics (Goldstein et al. 2016). Further studies examining the life history of other coral-reef fish species at the locations investigated in this study, and in MCEs worldwide, are warranted. Variability in reproduction between shallow and mesophotic reef-fish populations should also be investigated to understand energetic trade-offs in life-history traits.
This study provides life-history information for three coral-reef fish species endemic to the Hawaiian Archipelago and Johnston Atoll that have been the subject of few age-based demographic studies. Not only do endemic species hold high cultural importance, they also serve an important function in the origin of biodiversity on coral reefs (Bowen et al. 2013). However, endemic species are also more susceptible to ill consequences that can arise from declines in habitat quality due to their restricted geographic ranges (Bonn et al. 2002). The vulnerability of endemic species emphasizes a need to properly understand the habitat occupied at all depths of their distributions. Given that reef-fish assemblages at mesophotic depths are dominated by endemic species (Kane et al. 2014; Kosaki et al. 2016), MCEs may act as hotspots for marine biodiversity that require special management and conservation consideration.
With the continued global loss of coral reefs (Wilkinson 2008), generating information on MCEs is critical for conserving and managing environments that may eventually serve as refugia for coral-reef species to rejuvenate shallow systems. The sustainable management of MCE fish communities can contribute to maintaining healthy fisheries and local and regional biodiversity in coral-reef ecosystems (Riegl and Piller 2003). Determining whether and how the life-history strategies of mesophotic fishes vary from their shallow water conspecifics is critical in the effective assessment and management of MCE populations because the establishment of accurate demographic parameters is essential for informing sound decision-making by fisheries managers (Ault et al. 2014; Nadon et al. 2015). This study addresses a gap in knowledge of understudied mesophotic environments by further documenting how life-history traits change with depth. The intraspecific variability observed between shallow and mesophotic coral-reef fishes indicates that MCEs harbor organisms with life-history patterns distinct from their shallow water complements, which must be considered in future management decisions to protect these valuable environments.
References
Atkinson D, Sibly RM (1997) Why are organisms bigger in colder environments? Making sense of a life history puzzle. Trends Ecol Evol 12:235–239
Ault JS, Smith SG, Browder JA, Nuttle W, Franklin EC, Luo J, DiNardo GT, Bohnsack JA (2014) Indicators for assessing the ecological dynamics and sustainability of southern Florida’s coral reef and coastal fisheries. Ecol Indic 44:164–172
Baker EK, Puglise KA, Harris PT (2016) Mesophotic coral ecosystems—a lifeboat for coral reefs?. UNEP and GRID-Arendal, Nairobi and Arendal, p 98
Boehlert GW, Watson W, Sun LC (1992) Horizontal and vertical distributions of larval fishes around an isolated oceanic island in the tropical Pacific. Deep Sea Res A 39:439–466
Bongaerts P, Ridgway T, Sampayo EM, Hoegh-Guldberg O (2010) Assessing the “deep reef refugia” hypothesis: focus on Caribbean reefs. Coral Reefs 29:309–327
Bonn A, Rodrigues ASL, Gaston KJ (2002) Threatened and endemic species: are they good indicators of patterns of biodiversity on a national scale? Ecol Lett 5:733–741
Booth DJ (1991) Larval settlement and juvenile group dynamics in the domino damselfish (Dascyllus albisella). Ph.D. thesis, Oregon State University, p 193
Booth DJ (1992) Larval settlement patterns and preferences by domino damselfish Dascyllus albisella Gill. J Exp Mar Bio Ecol 155:85–104
Booth DJ, Hixon MA (1999) Food ration and condition affect early survival of the coral reef damselfish, Stegastes partitus. Oecologia 121:364–368
Booth DJ, Alquezar RA (2002) Food supplementation increases larval growth, condition and survival of the brooding damselfish Acanthochromis polyacanthus (Pomacentridae). J Fish Biol 60:1126–1133
Bowen BW, Rocha LA, Toonen RJ, Karl SA (2013) The origins of tropical marine biodiversity. Trends Ecol Evol 28:359–366
Bridge TCL, Luiz OJ, Coleman RR, Kane C, Kosaki R (2016) Ecological and morphological traits predict depth-generalist fishes on coral reefs. Proc R Soc Lond B Biol Sci 283:20152332
Bridge TCL, Done TJ, Friedman A, Beaman RJ, Williams SB, Pizarro O, Webster JM (2011) Variability in mesophotic coral reef communities along the Great Barrier Reef, Australia. Mar Ecol Prog Ser 428:63–75
Brock VE, Chamberlain TC (1968) A geological and ecological reconnaissance off western Oahu, Hawaii, principally by means of the research submarine “Asherah”. Pacific Science 22:373–394
Brokovich E, Ben-Ari T, Kark S, Kiflawi M, DIshon G, Iluz D, Shashar N (2010) Functional changes of the visual system of the damselfish Dascyllus marginatus along its bathymetric range. Physiol Behav 101:413–421
Choat JH, Axe LM (1996) Growth and longevity in acanthurid fishes; an analysis of otolith increments. Mar Ecol Prog Ser 134:15–26
Choat JH, Robertson DR (2002) Age-based studies on coral reef fishes. In: Sale PF (ed) Coral reef fishes: dynamics and diversity in a complex ecosystem. Academic Press, San Diego, CA, pp 57–80
Choat JH, Robertson DR, Ackerman JL, Posada JM (2003) An age-based demographic analysis of the Caribbean stoplight parrotfish Sparisoma viride. Mar Ecol Prog Ser 246:265–277
Conover DO, Present TMC (1990) Countergradient variation in growth rate: compensation for length of the growing season among Atlantic silversides from different latitudes. Oecologia 83:316–324
De’ath G, Lough JM, Fabricius KE (2009) Declining coral calcification on the Great Barrier Reef. Science 323:116–119
Donelson JM, McCormick MI, Munday PL (2008) Parental condition affects early life history of a coral-reef fish. J Exp Mar Bio Ecol 360:109–116
Fowler AJ, Doherty PJ (1992) Validation of annual growth increments in the otoliths of two species of damselfish from the southern Great Barrier Reef. Marine and Freshwater Research 43:1057–1068
Franklin EC, Jokiel PL, Donahue MJ (2013) Predictive modeling of coral distribution and abundance in the Hawaiian Islands. Mar Ecol Prog Ser 481:121–132
Fukunaga A, Kosaki RK, Wagner D, Kane C (2016) Structure of mesophotic reef fish assemblages in the Northwestern Hawaiian Islands. PLoS One 11:e0157861
Fulton TW (1904) The rate of growth of fishes. Fisheries Board of Scotland Annual Report 22, Edinburgh, UK, pp 141–241
Glynn PW (1996) Coral reef bleaching: facts, hypotheses and implications. Glob Chang Biol 2:495–509
Goldstein ED, D’Alessandro EK, Sponaugle S (2016) Demographic and reproductive plasticity across the depth distribution of a coral reef fish. Sci Rep 6:34077
Gove JM, Williams GJ, McManus MA, Heron SF, Sandin SA, Vetter OJ, Foley DG (2013) Quantifying climatological ranges and anomalies for Pacific coral-reef ecosystems. PLoS One 8:e61974
Grigg RW, Plovina J, Friedlander AM, Rohmann SO (2008) Biology of coral reefs in the Northwestern Hawaiian Islands. In: Riegl B, Dodge RE (eds) Coral reefs of the USA. Springer, Florida, pp 573–594
Groll P (1984) Recruitment and growth of juvenile Dascyllus albisella. M.Sc. thesis, University of Hawaii, p 62
Gust N, Choat JH, Ackerman JL (2002) Demographic plasticity in tropical reef fishes. Mar Biol 140:1039–1051
Haddon M (2011) Modelling and quantitative methods in fisheries, 2nd edn. Taylor & Francis, Boca Raton, Florida
Hinderstein LM, Marr JCA, Martinez FA, Dowgiallo MJ, Puglise KA, Pyle RL, Zawada DG, Appeldoorn R (2010) Introduction to mesophotic coral ecosystems: characterization, ecology, and management. Coral Reefs 29:247–251
Hoegh-Guldberg O, Mumby PJ, Hooten AJ, Steneck RS, Greenfield P, Gomez E, Harvell CD, Sale PF, Edwards AJ, Caldeira K, Knowlton N, Eakin CM, Iglesias-Prieto R, Muthiga N, Bradbury RH, Dubi A, Hatziolos ME (2007) Coral reefs under rapid climate change and ocean acidification. Science 318:1737–1742
Hoey J, McCormick MI, Hoey AS (2007) Influence of depth on sex-specific energy allocation patterns in a tropical reef fish. Coral Reefs 26:603–613
Holbrook SJ, Schmitt RJ (1988) Effects of predation risk on foraging behavior: mechanisms altering patch choice. J Exp Mar Bio Ecol 121:151–163
Jones RS (1968) Ecological relationships in Hawaiian and Johnston Island Acanthuridae (surgeonfishes). Micronesica 4:309–361
Kahng SE, Garcia-Sais JR, Spalding HL, Brokovich E, Wagner D, Weil E, Hinderstein L, Toonen RJ (2010) Community ecology of mesophotic coral reef ecosystems. Coral Reefs 29:255–275
Kane C, Kosaki RK, Wagner D (2014) High levels of mesophotic reef fish endemism in the Northwestern Hawaiian Islands. Bull Mar Sci 90:693–703
Kimura DK (1980) Likelihood methods for the von Bertalanffy growth curve. Fishery Bulletin 77:765–776
Kleypas JA, McManus JW, Meñez LAB (1999) Environmental limits to coral reef development: where do we draw the line? Am Zool 39:146–159
Kosaki R, Pyle RL, Leonard JA, Hauk BB, Whitton RK, Wagner D (2016) 100% endemism in mesophotic reef fish assemblages at Kure Atoll, Hawaiian Islands. Marine Biodiversity. doi:10.1007/s12526-016-0510-5
Kritzer JP, Davies CR, Mapstone BD (2001) Characterizing fish populations: effects of sample size and population structure on the precision of demographic parameter estimates. Can J Fish Aquat Sci 58:1557–1568
Langston R, Longenecker K, Claisse J (2009) Reproduction, growth, and mortality of kole, Ctenochaetus strigosus. Final report. Division of Aquatic Resources, Honolulu, Hawaii, pp 1–25
Leichter JJ, Genovese SJ (2006) Intermittent upwelling and subsidized growth of the scleractinian coral Madracis mirabilis on the deep fore-reef slope of Discovery Bay, Jamaica. Mar Ecol Prog Ser 316:95–103
Lesser MP (2006) Benthic–pelagic coupling on coral reefs: feeding and growth of Caribbean sponges. J Exp Mar Bio Ecol 328:277–288
Luckhurst BE, Dean JM, Reichert M (2000) Age, growth and reproduction of the lane snapper Lujanus synagris (Pisces: Lutjanidae) at Bermuda. Mar Ecol Prog Ser 203:255–261
Meekan MG, Wellington GM, Axe L (1999) El Niño-Southern Oscillation events produce checks in the otoliths of coral-reef fishes in the Galapagos Archipelago. Bull Mar Sci 64:383–390
Mundy BC (2005) Checklist of the fishes of the Hawaiian archipelago. Bishop Museum Press, Honolulu, Hawaii, p 704
Nadon MO, Ault JS, Williams ID, Smith SG, DiNardo GT (2015) Length-based assessment of coral reef fish populations in the main and northwestern Hawaiian Islands. PLoS One 10:e0133960
Page HM, Dugan JE, Schroeder DM, Nishimoto MM, Love MS, Hoesterey JC (2007) Trophic links and condition of a temperate reef fish: comparisons among offshore oil platform and natural reef habitats. Mar Ecol Prog Ser 344:245–256
Pauly D (1998) Tropical fishes: patterns and propensities. J Fish Biol 53:1–17
Puglise KA, Hinderstein LM, Marr JCA, Dowgiallo MJ, Martinez FA (2009) Mesophotic coral ecosystems research strategy: international workshop to prioritize research and management needs for mesophotic coral ecosystems, Jupiter, Florida, 12–15 July 2008. NOAA Technical Memorandum NOS NCCOS 98 and OAR OER 2, National Oceanographic and Atmospheric Administration, Silver Spring, MD, p 24
R Development Core Team (2009) A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria
Ralston S, Gooding RM, Ludwig GM (1986) An ecological survey and comparison of bottom fish resource assessments (submersible versus handline fishing) at Johnston Atoll. Fishery Bulletin 84:141–155
Randall JE (1955) A revision of the surgeon fish genus Ctenochaetus, family Acanthuridae, with descriptions of five new species. Zoologica 40:149–166
Randall JE (1998) Shore fishes of Hawai’i. University of Hawai’i Press, Honolulu, p 216
Randall JE, Clements KD (2001) Second revision of the surgeonfish genus Ctenochaetus (Perciformes: Acanthuridae), with descriptions of two new species. Indo-Pacific Fishes 32, Bishop Museum, Honolulu, Hawaii, 33 pp
Ricker W (1975) Computation and interpretation of biological statistics of fish populations. Bulletin of the Fisheries Research Board of Canada 191:1–382
Riegl B, Piller WE (2003) Possible refugia for reefs in times of environmental stress. Int J Earth Sci 92:520–531
Rooney J, Donham E, Montgomery A, Spalding H, Parrish F, Boland R, Fenner D, Gove J, Vetter O (2010) Mesophotic coral ecosystems in the Hawaiian archipelago. Coral Reefs 29:361–367
Ruttenberg BI, Haupt AJ, Chiriboga AI, Warner RR (2005) Patterns, causes and consequences of regional variation in the ecology and life history of a reef fish. Oecologia 145:394–403
Sancho G, Ma D, Lobel PS (1997) Behavioral observations of an upcurrent reef colonization event by larval surgeonfish Ctenochaetus strigosus (Acanthuridae). Mar Ecol Prog Ser 153:311–315
Spalding H (2012) Ecology of mesophotic macroalgae and Halimeda kanaloana meadows in the Main Hawaiian Islands. Ph.D. thesis, University of Hawaii, p 199
Srinivasan M (2003) Depth distributions of coral reef fishes: the influence of microhabitat structure, settlement, and post-settlement processes. Oecologia 137:76–84
Stevenson RAJ (1963) Life history and behavior of Dascyllus albisella Gill, a pomacentrid reef fish. Ph.D. thesis, University of Hawaii, p 221
Swerdloff SN (1970) The comparative biology of two Hawaiian species of the damselfish genus Chromis (Pomacentridae). Ph.D. thesis, University of Hawaii, p 192
Trip EL, Choat JH, Wilson DT, Robertson DR (2008) Inter-oceanic analysis of demographic variation in a widely distributed Indo-Pacific coral-reef fish. Mar Ecol Prog Ser 373:97–109
US Army Corps of Engineers (1983) Johnston Atoll chemical agent disposal system (JACADS): environmental impact statement. US Army Corps of Engineers, Hawaii, p 82
von Bertalanffy L (1938) A quantitative theory of organic growth (inquiries on growth laws. II). Human Biology 10:181–213
Wagner D, Kosaki RK, Spalding HL, Whitton RK, Pyle RL, Sherwood AR, Tsuda RT, Calcinai B (2014) Mesophotic surveys of the flora and fauna at Johnston Atoll, Central Pacific Ocean. Mar Biodivers Rec 7:e68
Wellington GM, Robertson DR (2001) Variation in larval life-history traits among reef fishes across the Isthmus of Panama. Mar Biol 138:11–22
Werner EE, Gilliam JF, Hall DJ, Mittelbach GG (1983) An experimental test of the effects of predation risk on habitat use in fish. Ecology 64:1540–1548
Wilkinson C (2008) Status of coral reefs of the world: 2008. Global Coral Reef Monitoring Network and Reef and Rainforest Research Centre, Townsville, Australia, p 296
Williams ID, Baum JK, Heenan A, Hanson KM, Nadon MO, Brainard RE (2015) Human, oceanographic and habitat drivers of Central and Western Pacific coral-reef fish assemblages. PLoS One 10:e0120516
Acknowledgements
We thank the captain and crew of the NOAA ship Hi’ialakai and J. Burns, D. Coffey, R. Coleman, J. Copus, B. Hauk, I. Fernandez, L. Giuseffi, R. Kosaki, J. Leonard, K. Longenecker, Y. Papastamatiou, D. Pence, R. Pyle, M. Royer, J. Troller, D. Wagner, R. Whitton, and K. Williams and the ToBo Lab at HIMB for their assistance in procuring samples for this study. M. Hixon and I. Williams provided valuable guidance and review of this work. Z. Oyafuso and M. Kapur assisted with statistical analysis and R programming. This work was conducted under permits # PMNM-2012-0450A1, PMNM-2014-031, PMNM-2015-020, PMNM-2015-030, PMNM-2015-029, PMNM-2015-021. Funding support was NOAA award #NA10NMF4520163 (ECF). This is SOEST contribution 9848 and HIMB contribution 1670.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Biology Editor Dr. Andrew Hoey
Electronic supplementary material
Below is the link to the electronic supplementary material.
Fig. S1
Relationship between age and otolith weight (g) for Chromis verater (a), Ctenochaetus strigosus (b), and Dascyllus albisella (c) in the North Central Pacific (EPS 863 kb)
Rights and permissions
About this article

Cite this article
Winston, M.S., Taylor, B.M. & Franklin, E.C. Intraspecific variability in the life histories of endemic coral-reef fishes between photic and mesophotic depths across the Central Pacific Ocean. Coral Reefs 36, 663–674 (2017). https://doi.org/10.1007/s00338-017-1559-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00338-017-1559-8