
Abstract
More than 10 years have passed since a number of publications presented evidence for the possible cultivation of wild cereals based on charred remains of potential arable weeds and spikelet bases from late Pleistocene/early Holocene sites in northern Syria. Since then, a number of publications concerning the beginnings of cultivation have appeared which used results from the Syrian sites. This paper was inspired by issues raised in these publications, which will be addressed through an empirical approach, first by discussing the possible techniques and methods used for sowing, harvesting and tilling at sites that produced remains of wild cereals and pulses with reference to experimental cultivation and field observations of wild cereal habitats. Secondly, taking into account the range of possible methods of exploitation demonstrates the complexity of distinguishing between the gathering and the cultivation of morphologically wild cereals and the probability that these two modes were practiced intermittently and interchangeably. In addition to this the inadequacy of evidence obtained from charred plant remains, suggests that the concept of “predomestic” cultivation as used for northern Syria needs to be revised. Thirdly, the causes of the varying proportions of shattering and non-shattering spikelets occurring together are examined and found to be multiple, suggesting that selection rates obtained from ratios of charred spikelet bases need to be reconsidered.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
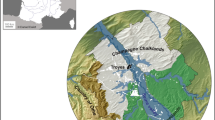
More than 10 years have passed since the publication of three papers in Vegetation History and Archaeobotany and two in Science (Tanno and Willcox 2006, 2012; Willcox et al. 2008; Willcox 2012, 2013), which presented evidence for potential arable weeds, the cultivation of wild cereals and mixed shattering and non-shattering spikelet bases from late Pleistocene/early Holocene sites in northern Syria. Since then, a number of publications concerning the beginnings of cultivation have appeared which were partially inspired by and/or used results from the Syrian sites (Purugganan and Fuller 2011; Allaby et al. 2017, 2022; Fuller and Stevens 2018; Abbo and Gopher 2020; Abbo et al. 2021; Fuller et al. 2022; Weide et al. 2022; Arranz-Otaegui and Roe 2023). In response to issues raised in these publications, an empirical approach to the beginnings of cultivation will be addressed by discussing possible techniques and methods for sowing, harvesting and tilling of cereals and pulses during the initial stages of cultivation. To examine the practicality of these techniques, five questions are asked: (1) How was the sowing of seed adopted? (2) What were the probable methods for harvesting wild cereals? (3) How did selection for non-shattering ears occur when wild cereals were cultivated? (4) How should we interpret mixtures of shattering spikelets mixed with non-shattering spikelet bases on early cultivation sites? (5) What sowing and tilling methods might have been used and what can potential arable weeds tell us about soil preparation? In order to respond to these questions, experimental cultivation of wild einkorn in southern France (Willcox 2007), is reconsidered in the light of new archaeobotanical evidence, taking into account observations of wild cereal and pulse habitats in Turkey, Syria, Jordan and Afghanistan. This will lead to a reassessment of possible modes of cereal and pulse exploitation at sites in northern Syria at the end of the Pleistocene/beginning of the Holocene (Fig. 1).

Approximate location of sites in northern Syria mentioned in the text
Readers are referred to the publications mentioned above and references there within for further chronological, geographical, environmental and archaeological backgrounds of the early-agriculture sites in southwest Asia and for the ongoing debates concerning early cultivation.
Possible techniques and methods for sowing, harvesting and tilling
(1) How was the sowing of seed adopted? The shift from gathering to cultivation has been likened to niche construction, a phenomenon that occurs in varying degrees for most plants and animals (Smith 2007). In our case, niche construction arises through cultural evolution and innovation (Creanza et al. 2017) within the framework of environmental constraints. What concerns us is the innovation of voluntary dispersal of seed. Involuntary dispersal by late Pleistocene gatherers would have been inevitable. During the 19 years of experimental cultivation of wild einkorn at Jalès in southern France (Willcox 1999a, b, 2007), it was observed that, on the path from the cultivated fields leading to and around the processing area, spilled grain germinated the following year into seedlings. This involuntary dispersal of wild einkorn occurred frequently and would have occurred during the late Pleistocene when cereal gatherers in the Near East transported harvests to habitations, processing areas or storage facilities. The easily-shattering spikelets would have spilled accidentally “en route” to and around settlements and would have germinated and developed the following year. This is not an argument in favor of the dump heap theory (Abbo et al. 2005). What needs to be emphasized is that individual members of communities that gathered wild cereals could have imitated involuntary dispersal by purposefully sowing outside wild cereal habitats. This phenomenon may have occurred over a wide geographical area from Anatolia through the northern and southern Levant, possibly extending as far as the Zagros. This broad space-time dimension, may have provided multiple occasions that triggered a conscious decision to sow seed, giving rise to the possibility that sowing was adopted independently by different communities. Plant propagation among hunter-gatherers is well known. Ethnographers during the 19th and first half of the 20th century observed hunter-gatherer groups who had little contact with farmers, such as the Bushmen, indigenous Australians, and native peoples of North America, engaged in burning, watering, burying tubers and seeding useful plants to promote their propagation (Harris 1977; Steensberg 1986; Harlan 1995; Smith 2001). If Epi-Palaeolithic communities carried out similar practices, these would have left little or no archaeobotanical record.
Dispersal of grain outside wild stands was possibly the first way that cereal gatherers embarked on voluntary sowing of cereals in the Near East. In this way natural habitats could have been expanded. Reserves of grain could be sown when wild stands became depleted due to over-exploitation, wildfires, grazing or drought. The sowing or dispersal of seed in its initial stages would have occurred by trial and error, perhaps independently in different geographic regions and was not necessarily enduring. Unfortunately these initial stages would have left little trace in the archaeological record. How sowing techniques may have developed with tilling will be discussed below.
(2) What were the probable methods for harvesting wild cereals? With the onset of cultivation, harvesting and processing procedures did not need to change because crops whether gathered in the wild or harvested in cultivated fields were morphologically identical. So tools, such as hafted flint blades and querns did not necessarily need to be modified. Tools for working the soil probably already existed, for manipulating building earth used in the construction of habitations.
Experimental cultivation (Willcox 2007) and extensive archaeological evidence for glossed flint blades (Maeda et al. 2016) indicate that the most probable harvesting technique for wild cereals in the Near East during the late Pleistocene/early Holocene would have been to harvest with hafted flint blades before shattering, when the spikelets remained intact or partially intact on the ear (Fig. 2). Experiments using methods such as uprooting, shaking into baskets and ground collecting were carried out but were found to be impractical compared to using flint sickles (Kislev et al. 2004; Willcox 2007, Table 12.2); if used, these methods would probably only represent a minor contribution.

Wild two-grained einkorn (Triticum boeoticum ssp. thaoudar) growing on Karaca Dağ in eastern Turkey showing the optimal state of ripeness for harvesting (30 Juin 2007). Note the ear in the upper centre which has started to shatter. On the Feekes scale for wheat ripening the grains correspond to 11.2 Mealy ripe, contents of kernel soft but dry, and 11.3 kernel hard (difficult to divide by thumb-nail)
The question is: Are early harvests before shattering viable? An experiment to test germination rates on wild cereals harvested early or green prior to shattering was carried out some years ago. The following is a more detailed description and explanation than that given in the original publication (Willcox 2007). Populations of wild einkorn and wild barley were broadcast onto tilled soil in the autumn. Then in the first week of July the plots were harvested in a single pass, using flint sickles. The resulting crop was then divided into three stages of ripeness: green, half-green and ripe (Table 1). Germination tests were carried out under laboratory conditions in October on: (a) naked grains, (the glumes having been removed by hand, avoiding damage to the embryo), to ascertain whether the presence of glumes affected dormancy: (b) hulled spikelets which would reflect natural conditions. 100 grains/spikelets were kept humid in the dark at a temperature of 18 °C and observed at 7 and 14 days.
The results (Table 1) of these tests revealed that dehusked grains had higher germination rates than hulled spikelets, confirming the role of the glumes in dormancy. For population No. 122 green spikelets gave 80% germination with hulled spikelets and 82% with dehusked grains. Of the 20 ungerminated spikelets it is possible that some may have germinated at a later date had the tests been carried out under field conditions with spikelets buried in the soil over winter. Half-green spikelets gave lower results, the cause of this is not known. Ripe spikelets for population 122 gave 92 (naked) and 90% (hulled) germination. The single example of wild barley gave a 54% germination rate.
Another experiment was carried out in two parts, first to test the percentage of shattering when the harvest was carried out when the ears were still half green, and second to test the yields on plots during four consecutive years relying only on spontaneously sown spikelets that had fallen to the ground before or during the harvest. Wild einkorn (population No. 122) was broadcast onto tilled soil in five plots in the autumn. The plots were then harvested with flint sickles the following June over a 10-day period. Three humidity tests were carried out during the harvest on spikelets from plots D, E, and H which gave 50.86, 52.68 and 47.5% humidity respectively but some upper spikelets had dried and a few spikelets had shattered before or during the harvest. The ripeness of the crop was approximately equivalent to that shown in Fig. 2. The shattered spikelets which had dispersed to the ground were collected and weighed from a subsample of one-meter squares for each of the five plots. Then the percentage of the total harvests was calculated. Spontaneous shattering before or during sickle-harvesting amounted to between 5 and 20% of the total harvest (Table 2). Not surprisingly, harvests towards the end of the 10-day period were riper and had a higher proportion of shattered spikelets than harvests carried out earlier. Once shattering starts, it proceeds rapidly leaving a narrow window to carry out efficient harvesting. When harvesting wild stands in Turkey, Syria as well as the experimental plots it was not uncommon to miss the optimal stage of ripeness only to find the majority of spikelets had shattered. Ground collection was found to be a very poor substitute compared to cutting with a flint tool.
For the second part of the experiment, plots were sickle-harvested and the crop of threshed spikelets (not dehusked) were weighed to record the yields (Table 3). The first year produced ratios of spikelets sown to spikelets harvested of between 1 to 7.2 and 1 to 2.5. Then for 3 consecutive years, the plots which were left fallow (without annual sowing), were harvested in early July. Yields varied but in general the second year’s crop was so sparse as to make harvesting unproductive. This demonstrates that when the harvest occurs at this stage, annual sowing is necessary to maintain the crop. The diminished yields are to be expected, given that as we have seen, sickle-harvests would have recovered between 95 and 80% percent of seed production leaving insufficient seed in the ground to maintain the crop. Wild cereal stands in their natural habitat, if gathered at this stage of ripeness might also become depleted, if an area was sickle-harvested year after year and particularly if seed loss due to insects, rodents and birds was high. These experiments call for being reproduced on wilds stands in their natural habitat in the Near East before these habitats are disappear.
Harvests consisting of multiple passes have been suggested in the literature (Maeda et al. 2016, p. 234), but the sickle-harvests carried out on the experimental plots and those carried out in wild cereal habitats in Syria and Turkey left only stubble after a single pass, making second passes superfluous. In conclusion, green or half-green harvests are preferable and give a good yield the following year with a single pass, but without additional annual sowing the yields are at risk of diminishing rapidly. However it should be stressed that these experiments may not exactly reflect the original conditions, and would be made more dependable by repetition using wilds stands in their natural habitat in the Near East before they disappear.
(3) How did selection for non-shattering ears occur when wild cereals were cultivated? Two opposing models have been proposed which attempt to explain the early stages of cereal cultivation in the Near East. The first model posits knowledge-based conscious selection leading to a single event in a core area resulting in rapid selection of crops with non-shattering ears (Abbo and Gopher 2020). The second model posits natural selection giving rise to a protracted, non-centered geographically, dispersed system which developed over a period of millennia (Fuller et al. 2012). This model was partly inspired by the evidence for mixtures of shattering and non-shattering spikelets (see below) occurring in the same layers (Tanno and Willcox 2006, 2012), which was interpreted as evidence for low or slow selection rates and hence the slow evolution towards populations with non-shattering ears.
Wheat and barley are predominately self-pollinating, which facilitates selection. Traits such as the non-shattering ear can be fixed in the population more readily than with cross-pollination because populations consist of multiple homozygous pure lines; this allows a single plant to develop into a new population and thus stabilize a desirable genetic strain or trait (Zohary et al. 2012). In addition, self-pollinated grain crops would have been isolated reproductively from their wild progenitors, which, in the case of the Near East, may have been growing not far from cultivated plots (Zohary ibid.).
However, wild wheats and barley are not obligatory selfers. Cross-pollination depends on the timing of the opening of the anthers. Boyeldieu (1980) reported that under increasingly hot conditions einkorn spikelets open prematurely, raising rates of cross-fertilization to 10 or 15%. Cultivated wheats have been shown to exhibit introgression with their wild relatives. Arrigo et al. (2011) and Fricano et al. (2014) suggest that einkorn crossed with T. urartu; they state that “In a germ plasm collection, rare wild T. urartu lines with the presence of T. monococcum alleles were found”. Pajkovic et al. (2014) report “extensive gene flow between wheat (Triticum sp.) and several wild relatives of the genus Aegilops has recently been detected despite notoriously high levels of selfing in these species”. Thus within the habitats of wild progenitors occasional cross-pollination would have provided crops with genetic flexibility and variation while self-fertilization facilitates selection.
Natural selection operated under cultivation whether or not there was conscious selection. Thus we will assess the feasibility of conscious selection for non-shattering ears by early cultivators. Conscious selection can be carried out in two ways: (a) Pure or single line selection: for Lupton (1987) pure line selection was a late development. He states “Little attempt was made to improve the varieties of cereal crops until the early years of the 19th century. The crops grown at this time consisted of a wide range of ‘land races’, each of which had evolved in the area where it was grown, mainly through natural selection”. Pure or single line selection necessitates isolating the chosen line from the main population. It has the disadvantage of reducing genetic diversity. Diversity was necessary in order to have crops with good yield stability (Abbo et al. 2010). In addition it would have been difficult to isolate the single line from the rest of the crop. (b) Mass or recurrent selection: this is the only method known to have been used by subsistence farmers. Mass selection involves choosing grains from plants with desired characteristics for the next generation. They must make up a certain proportion of the population (hence mass selection) in order to continue to have a viable crop. The advantage is that plants do not need to be isolated and so this is the method used by subsistence farmers to select for suitable traits. Mass selection (Redden and Jensen 1974; Hallauer and Darrah 1985; Bos and Caligari 2008; Marais and Botes 2010) requires three prerequisites to be fulfilled: (1) that the trait can be readily recognized in the phenotype, (2) that the trait has a high degree of heritability and is not the result of phenotypic plasticity and (3) that it is desirable. For a trait such as glume color in einkorn, the three prerequisites are fulfilled, so a landrace with this trait could be selected using the mass selection technique. For increased grain size, prerequisite 2 would not be fulfilled because grain size depends mainly on phenotypic plasticity; size is mainly determined by position on the ear, time of harvesting and growing conditions and thus has a very low degree of heritability. Mass selection for plants with non-shattering ears would be problematical because prerequisite 1 would not be fulfilled as this trait would be difficult to recognize, first because harvests would occur before shattering, second because delayed ripening is common, so that some ears remain intact longer than others, only to shatter when dehusked. Prerequisite 2, would be fulfilled because the trait is heritable. Prerequisite 3 would not be fulfilled, because without having experienced harvesting a population of totally non-shattering ears, cultivators could not know that this trait was desirable. Most importantly, if mass selection was attempted for non-shattering ears, a certain proportion of plants with this trait must be already present in the population. Thus natural selection for this trait would have already begun and may have outperformed conscious selection. Finally, early cultivators may not have appreciated that a trait could be inherited and may have perceived diverse traits as arising from environmental factors.
Under natural selection, selective pressures in favor of non-shattering ears would vary depending on harvesting techniques. Efficient harvests prior to shattering could lead to low selection rates (as seen in Table 2), as opposed to late harvests which could lead to high rates. Hillman and Davies (1990) and Zohary et al. (2012, p. 22) argued for rapid selection. Observations during experimental cultivation of wild einkorn suggest that shattering types would compete poorly in the presence of non-shattering types. One of the wild einkorn fields at Jalès was accidentally contaminated with a few non-shattering plants. They multiplied rapidly and became dominant in less than ten years at the expense of the shattering type (Willcox 2007). This suggests that once present under conditions of annual sowing in the same plot, selection for non-shattering ears could progress rapidly.
In conclusion, conscious selection for non-shattering ears in wild wheat and barley by early cultivators is impractical and therefore highly improbable. Under natural selection, if a few non-shattering ears are present in an isolated shattering population which is systematically sown and tilled on an annual basis; the non-shattering types should theoretically be highly competitive.
(4) How should we interpret mixtures of shattering and non-shattering spikelet bases on early cultivation sites? Such mixtures or combinations of spikelet bases appear at Halula (Willcox 1996) and Abu Hureyra (de Moulins 2000) for the Middle PPNB and at other early Neolithic sites in the Near East somewhat earlier, ca. 8500 cal bc. For example, in Aswad levels 2 and 3, non-shattering barley spikelet bases make up 25%, but shattering types persist (Tanno and Willcox 2012; Douché and Willcox 2018, Fig. 5). The proportions vary widely from site to site, which may be due to several factors outlined below.
Mixtures of spikelet bases have been interpreted to indicate low rates of selection (Tanno and Willcox 2006; Purugganan and Fuller 2011). This interpretation has become widely accepted over the last 15 years or so, contradicting earlier estimates by Hillman and Davies (1990) and Zohary et al. (2012, p. 22) who suggested that once cultivation of wild einkorn began, plants with shattering ears would be eliminated within a dozen generations, or up to 200 generations by natural selection. Despite these two opposing interpretations, discussions of the factors that caused these mixtures have been limited. Here five possible explanations for this mixing are presented, being neither mutually exclusive nor exhaustive.
Possibility (a): The mixtures were the result of separate harvests from cultivated fields with non-shattering ears and harvests from wild stands with shattering ears. This would be the case if gathering continued well after the appearance of non-shattering types, the two types becoming mixed on the settlement. Harvests from wild stands to replenish seed stock may have been the only recourse when cultivated crops failed due to drought, disease, pests, wildfires, grazing, spoiled storage or pillage.
Possibility (b): The two types represent two distinct populations of shattering and non-shattering ears growing together in the same cultivated fields. The latter population would compete poorly with the former, but the degree of completion would depend on harvesting methods.
Possibility (c): There were types which had both shattering and non-shattering spikelets on the same ear. It has been observed that the lowest spikelets of wild cereals may remain fused (Kislev 1992), but never make up more than about 10% of the total. Weedy populations of einkorn and barley fall into another category with more spikelets remaining intact. They have been observed for many years (Harlan and Zohary 1966) and can be found in fields in Turkey and Syria (Fig. 3). They adapted to compete under cultivation by possessing spikelets which may shatter before or during the harvest and others which remain intact until threshing. Peleg et al. (2022) described a similar wild population of emmer in which half the ear shatters while the lower half stays intact. Two populations obtained from gene banks of “wild” einkorn grown at Jalès had similar ears.

Weedy form of wild einkorn occuring in wheat fields, southeast of Gaziantep (Turkey). Note the lower spikelets remain attached after the plant has dried
Possibility (d): Identification and taphonomy may affect the proportions of mixtures. A large proportion of archaeological einkorn and emmer spikelet bases remain unidentified due to damage prior to charring (see for example emmer spikelet bases from Aswad, Fig. 7, Douché and Willcox 2018). Experimental dehusking of wild einkorn and emmer has shown that dehusking damages the spikelet bases, producing unidentifiable scars (Tanno and Willcox 2012). Only the infrequent undamaged spikelet bases can be safely used for the distinction between the two types. Bias may occur if smooth wild-type scars are more easily distinguished than the non-shattering scars. Barley spikelet bases recorded by Tanno and Willcox (2012) and Douché and Willcox (2018) were less damaged, perhaps because barley underwent less pounding. Hence there are fewer unidentified specimens, suggesting that barley is more reliable for assessing the proportions of non-shattering to shattering spikelet bases. Other taphonomic factors prior to, during and after charring as well as sampling techniques vary from site to site.
Possiblity (e): Harvesting before shattering results in a mix of brittle and non-brittle spikelets (Peleg et al. 2022, p. 2,005). This possibility is improbable because the separation of the abscission layer (shattering) is brought about by desiccation and not the maturing process. This was demonstrated by experimental harvests of wild einkorn and barley carried out in the fields at Jalès over many years. Intact green ears, containing grains still at the milky stage of development shattered after drying leaving smooth, brittle, shattering-type abscission scars Under the July sun of southern France, drying was achieved rapidly, often within 24 h. As with the other experiments, this would benefit by being repeated using wild emmer.
In conclusion, the mixtures or combinations of shattering and non-shattering abscission scars recovered from archaeological sites could be due to multiple factors and at present there is no reliable way to assess which factors were causing the mixtures. Whatever the case, genetic input from wild populations may have continued independently at geographically widely separated sites, providing variability and allowing cereals to evolve independently in different regions.
(5) What sowing and tilling methods could have been used and what can potential arable weeds tell us about soil preparation? For cereals in the wild and under cultivation, seed loss must be kept to a minimum to maintain a population. Early cultivators through experience would have realized the need to keep seed loss to a minimum. Burying seed protects it from predators and also improves germination rates by keeping seeds humid and avoids desiccation of young seedlings. A possible method for early cultivators might have been to sow in fresh plots every year with minimal preparation, such as the removal of vegetation. This would loosen the soil and so provide a patchy tilth suitable for sowing. This kind of shifting cultivation has two advantages: the soil is not depleted of its nutrients and potential arable weeds would have difficulty in becoming established.
As we have seen above, experiments suggest it was necessary to sow annually if plots were harvested every year. Soil preparation would be desirable to keep seed loss to a minimum. Minimal tilling occurs when seed is dribbled into holes or parallel furrows made at predetermined distances and depths and then covered with earth. These methods can be carried out using digging sticks and create a minimum of soil disturbance. Broadcast sowing, in contrast, requires the preparation of a tilth onto which seed is cast and then buried, typically by raking. This method is better adapted to large-scale cultivation than drilling or dribbling, and it creates more soil disturbance; which brings us to the subject of the adaptation and proliferation of arable weeds. Research into wild/weed taxa or potential arable weeds was pioneered by Colledge (1998). Some taxa which evolved into the first arable weeds in the Near East would have grown and indeed still grow alongside wild cereals in their natural habitats. This has been observed among modern wild cereals in their natural habitats in eastern Turkey and northern and southern Syria and in Israel (Weide et al. 2023). These taxa could have been harvested along with wild cereals by inhabitants of sites where only gathering was practiced. Significantly their presence alongside wild cereals means they could have been brought into cultivation when wild cereals were gathered for sowing.
Weeds evolved under conditions of tilling by adaptations that affected the number of seeds, seed size, dormancy, germination, flowering time and duration (Bourgeois et al. 2019). The progenitor weed species would have been pre-adapted to tilling by already possessing some of the above traits, making them equipped to take advantage of the loose open soils where they could proliferate more rapidly than in their natural habitat. Under cultivation it is well known that weed taxa proliferate. Thus when cultivation started, weed taxa would be expected to develop a higher plant density than in wild cereal stands.
Under annual tilling and sowing, weeds compete with crops for water, light and nutrients, hence it is important to control them in order to produce good harvests. It is probable that early cultivators attempted to reduce weeds by weeding fields and by cleaning crops during processing. Once the weeds were amassed they would have to be disposed of. A possibility might be deliberate burning to destroy them. This would increase the likelihood of potential weed taxa becoming part of the charred assemblage. Two approaches attempting to recognize tilling have been used. Willcox (2012) compared presence/absence of potential weed taxa from late Pleistocene/early Holocene sites and Weide et al. (2022) endeavored to distinguish arable fields from wild cereal habitats by assessing two functional traits, flowering duration and the capacity to regenerate vegetatively (see also Willcox 2023). The results are discussed below.
Fuller and Stevens (2018) expanded the taxonomic approach by adding a wider range of sites and taxa. They conclude that their review gives a positive correlation between weed taxa presence and cereal dependency. They also point out that the original habitats of potential weeds, pre-dating cultivation, either no longer exist in their original form or the ancestral forms of these species have since become extinct. Clearly there is a need for more field studies combined with more work on charred remains with regard to potential arable weeds in order to obtain more reliable evidence for tilling than we have at present.
Reassessing the exploitation of cereals and pulses at late Pleistocene/early Holocene sites in northern Syria
On the basis of the above discussions, cereal and pulse exploitation during the late Pleistocene and early Holocene (Natufian to the middle PPNB, Table 4) in northern Syria will be reassessed taking into account a wider range of possible modes of gathering, sowing and tilling methods than previously considered (Willcox 2007). The evidence for and against cultivation of wild cereals and pulses during the PPNA will also be discussed.
The compositions of the cereal and pulse assemblages are distinct for individual sites during the Epi-Palaeolithic and PPNA periods. This may result from the proximity of sites to wild cereal habitats which vary geographically depending on the species (Willcox 2005). For the Epi-Palaeolithic, at Dederiyeh cave, samples dated to between 11180 and 10950 cal bc (Tanno et al. 2013) were dominated by T. boeoticum/dicoccoides in comparison to Abu Hureyra, dated from 11250 to 10750 cal bc, where wild rye was dominant (Hillman 2000). Both sites have higher frequencies of edible non-founder crops including fruits and nuts (Arranz-Otaegui and Roe 2023). They could be considered primarily as gathering sites with little or no evidence for tilling. But as argued above, cereal gatherers observed involuntary sowing and could practice voluntary sowing if the need arose. The expansion of wild stands through voluntary dispersal by inhabitants is a possibility, and any occasional cultivation would leave little or no trace in the archaeobotanical record.
The PPNA has similar distinct compositions. At Tell Qaramel (Willcox and Herveux 2013) only early PPNA layers were extensively sampled from phase H2, dated to 10200−9300 cal bc (Mazurowski and Kanjou 2012). T. boeoticum was dominant. This site has a fruit assemblage dominated by Pistacia, Amygdalus and Celtis similar to that of Dederiyeh, the two sites being situated in the same vegetation zone which included wild wheats. However, pulses are common at Tell Qaramel and rare at Dederiyeh. Jerf el Ahmar was extensively sampled, covering a period of occupation of over 500 years between approximately 9500 and 8800 cal bc and is distinguished by high frequencies of wild barley followed by wild rye. Wild barley is adapted to higher temperatures than wild rye, which may explain why wild barley became dominant during the global warming which corresponds with the early Holocene. Ubiquity percentages and archaeological evidence leave no doubt that cereals and pulses were the dominant food resource during the PPNA in northern Syria (Willcox et al. 2008; Willcox and Stordeur 2012). Circumstantial evidence for the cultivation of wild progenitors (ibid.) was posited on the basis of six lines of evidence: (1) The apparent location of sites far from wild cereal habitats. (2) A reduction in gathered edible seeds of non-founder plants compared to an increase in founder crops. (3) An increase in the grain size of cereals possibly due to enhanced growing conditions with the adoption of plant husbandry. (4) Large-scale cereal-processing installations. (5) An increase in potential arable weed taxa, similar to those found on later cultivation sites compared to earlier sites. (6) Evidence for extensive rodent activity, in particular the house mouse (ibid.). Individually these lines of evidence do not demonstrate that cultivation was being practiced; however, taken together they correspond to large-scale exploitation of cereals which is reinforced by the ubiquitous use of cereal chaff as tempering material in widely examined building earth (ibid.). Whether this was large-scale exploitation of wild stands or cultivation or both is difficult to disentangle. Systematic regular cultivation at Jerf el Ahmar is improbable, because if it was practiced during the 500 years or more of occupation, we should expect to see, as discussed above, the appearance of non-shattering ears. Despite this, diverse scenarios of sowing, tilling, gathering and harvesting can be envisaged. For example, rye could have been gathered from farther north while barley might have been cultivated. Wild barley, which today grows locally appears to have been a separate crop from rye (Willcox and Stordeur 2012, Fig. 9). Rye is not adapted to the Euphrates valley and today grows only at much higher altitudes, 200 kms farther north. This leaves the possibility of various scenarios for these two cereals: importation for consumption, introduction for sowing, local gathering, local cultivation or combinations of these modes. At late PPNA Dja’de (Douché and Willcox 2018), dated to between 9300 and 8800 cal bc, the dominant crop is lentil. In the wild these pulses grow in small dispersed stands, unlike cereals, making harvesting lentils very time-consuming. Harvesting could have been facilitated by cultivating large dense plots. High levels of dormancy could have been overcome by creating a rich seed bank in the soil or by soaking in warm water (Zohary pers. comm.). A possibility, among others, is that during phase I lentils were a major cultivar and during phases II and III emmer and einkorn (absent from phase I) were introduced for cultivation.
Concerning potential weeds and evidence of tilling during the PPNA, there is an increase in these taxa between Natufian Dederiyeh and PPNA Tell Qaramel located in northwest Syria and between Natufian Abu Hureyra and PPNA Jerf el Ahmar located in the Euphrates valley (Willcox 2012). This increase in the number of potential weed taxa in the PPNA could be interpreted a sign of tilling. However, the lack of contexts with pure crop-processing remains and the fact that samples contain sediments accumulated from multiple activities, preclude the recognition of specific episodes of tilling or gathering. Weide et al. (2022) refer to “low-level disturbance and the possibility that flowering duration is indicative of wild stands at Jerf el Ahmar and Dja’de. However, it is unclear whether low-level disturbance represents low-level soil disturbance i.e. soil preparation or some other activity. The use of flowering duration obtained from identifications of potential arable charred seeds assumes the presence of pure crop-processing by-products which were not found at Jerf el Ahmar and Dja’de, For a criticism of the taphonomic, ecological and taxonomic methods used by Weide et al. (2022) see Willcox (2023). Neither the taxonomic approach nor the functional trait analysis confirms that the wild pulses and cereals which occur throughout the long sequence at Jerf el Ahmar and Dja’de were gathered exclusively from wild stands. The possibility of cultivation should not be overlooked.
In conclusion, during the long occupations at PPNA Jerf el Ahmar 9500−8800 cal bc and Dja’de I 9300−8800 an array of possible modes of cereal and pulse exploitation could have been practiced. If systematic annual cultivation was being carried out over protracted periods, natural selection would have produced non-shattering population, which is not the case. Several lines of evidence for large-scale use of wild cereals at Jerf el Ahmar mark a significant point on the path towards the adoption of cultivation, but the archaeobotanical record based on charred remains does not allow us to distinguish unequivocal episodes of gathering or cultivation during this long period.
Following on from the PPNA, Dja’de II and III represent the early PPNB (8800−8300 cal bc), where we see for the first time in the Euphrates valley the appearance or introduction of emmer, perhaps for cultivation. Unfortunately no representative samples were recovered from Mureybet for this period. After 8300 cal bc we are faced with a hiatus in the archaeobotanical sequence (Fig. 4) until ca. 7700 cal bc, corresponding to the late Middle PPNB levels at Halula and Abu Hureyra. These levels had an economy that resembles mixed farming, with non-shattering emmer and barley and herding of pigs, sheep, goats and cows. The area of occupation of these sites increases dramatically (Goring-Morris and Belfer-Cohen 2008), implying an increase in the human population, which would appear to be a consequence of a production economy, but unfortunately in northern Syria we lack evidence for the cultural and economic transitional developments which led to mixed farming.

14c bc cal date ranges for phases/layers of charred remains from sites in northern Syria mentioned in the text. The hiatuses may be due to lack of sampling and excavation, and/or absence of habitation. The hiatus between the PPNA and the Middle PPNB while apparent in northern Syria does not occur in the southern Levant, including southern Syria. Dates were obtained from Benz (2010)
Conclusions
Following an empirical approach, the replies to the questions asked in this paper suggest that a wide variety of modes of harvesting, tilling and sowing can be envisioned for early Neolithic sites. Thus the initiation of sowing wild seed went from involuntary to diverse possibilities of voluntary sowing. Harvesting of not fully ripe cereals with minimal shattering using hafted flint blades was probably the preferred method for both wild stands and sown plots. Despite the early harvest the seed could be sown the following year to produce a viable population. However, annual gathering in the same area of wild stands might have led to reduced harvests and in the case of cultivated plots annual sowing would be necessary to maintain the crop and this would sooner or later lead to natural selection for non-shattering ears. Conscious selection for non-shattering ears is impractical and in order for it to be set in motion natural selection would have already been operating. The proportions of shattering compared to non-shattering spikelet bases could be due to numerous factors which may differ from sample to sample or site to site. Thus the proportions may not necessarily reflect changing rates of selection. The use of potential weeds to distinguish crops obtained from cultivation as opposed to wild stands is problematical because these plants may occur in both habitats. However, potential weeds would be expected to proliferate more rapidly with tillage compared to wild habitats and their seeds would be more likely to be incorporated into the charred assemblage than those originating in wild stands.
Taking into account the above empirical observations, a reassessment of cereal and pulse exploitation in northern Syria concludes as follows. Natufian settlements appear to have been primarily gathering communities. Despite the lack of evidence, occasional cultivation should not be ruled out. A gap (Fig. 4) in the archaeobotanical record between the end of the Pleistocene (late Natufian) and the beginning of the Holocene (early PPNA) corresponds in part with the Younger Dryas and may represent an interruption in the occupation of the region. With improved climatic conditions some PPNA settlements present evidence of large-scale processing of wild cereals, for example at Jerf el Ahmar and Gobekli (Dietrich et al. 2019). Earlier publications (Willcox 2007) suggested evidence for cultivation of wild cereals at sites in northern Syria. In retrospect the archaeobotanical data does not allow a differentiation between harvests from cultivated plots and those gathered from wild stands because samples with charred remains of crop processing by-products are derived from deposits which correspond to accumulations resulting from multiple activities. These deposits may potentially include remains derived both from wild and/or cultivated harvests. Indeed a probability that different combinations of modes of gathering, harvesting, sowing and tilling may have gone into and out of use during the PPNA is suggested by the augmentation of potential weeds compared to earlier periods.
Between the end of the early PPNB and the Middle PPNB there is another gap in the archaeobotanical record (Fig. 4). At the Middle PPNB sites in northern Syria emmer and barley were cultivated. Unfortunately, for the period leading up to the Middle PPNB with the emergence of mixed farming, evidence is lacking. Elsewhere in southwest Asia non-shattering emmer and barley appear ca. 8300 cal bc (Middle PPNB), leaving no doubt that cultivation was being practiced. Shattering types continue to be present at irregular frequencies during the middle and late PPNB. There are probably several reasons why these types persist, for example if weedy forms were present and also because the input of the non-shattering trait is probable at sites situated within the range of wild cereals.
In the future, excavation of new sites will allow more accurate comparisons of regional differences of charred crops and their byproducts. Further field work on wild cereals and potential weeds is necessary before their habitats are completely destroyed. Finally it is hoped that these discussions will encourage future research that will help to provide more solid evidence for the transition from gathering to cultivation during the late Pleistocene and early Holocene in northern Syria.
References
Abbo S, Gopher A (2020) Plant domestication in the Neolithic Near East: the humans-plants liaison. Quat Sci Rev 242:106,412. https://doi.org/10.1016/j.quascirev.2020.106412
Abbo S, Gopher A, Rubin B, Lev-Yadun S (2005) On the origin of Near Eastern founder crops and the ‘Dump-heap hypothesis’. Genet Resour Crop Evol 52:491–495. https://doi.org/10.1007/s10722-004-7069-x
Abbo S, Lev-Yadun S, Gopher A (2010) Yield stability: an agronomic perspective on the origin of Near Eastern agriculture. Veget Hist Archaeobot 19:143–150
Abbo S, Peleg Z, Lev-Yadun S, Gopher A (2021) Does the proportion of shattering vs. non-shattering cereal remains in archeobotanical assemblages reflect Near Eastern neolithic arable fields? Rev Palaeobot Palynol 284:104,339. https://doi.org/10.1016/j.revpalbo.2020.104339
Allaby RG, Stevens C, Lucas L, Maeda O, Fuller DQ (2017) Geographic mosaics and changing rates of cereal domestication. Philos Trans R Soc B 372:20,160,429. https://doi.org/10.1098/rstb.2016.0429
Allaby RG, Stevens CJ, Kistler L, Fuller DQ (2022) Emerging evidence of plant domestication as a landscape-level process. Trends Ecol Evol 37:268–279
Arranz-Otaegui A, Roe J (2023) Revisiting the concept of the ‘Neolithic founder crops’ in southwest Asia. Veget Hist Archaeobot 32:475–499. https://doi.org/10.1007/s00334-023-00917-1
Arrigo N, Guadagnuolo R, Lappe S, Pasche S, Parisod C, Felber F (2011) Gene flow between wheat and wild relatives: empirical evidence from Aegilops geniculata, Ae. neglecta and Ae. triuncialis. Evol Appl 4:685–695. https://doi.org/10.1111/j.1752-4571.2011.00191
Benz M (2010) Comments on radiocarbon dates of epipalaeolithic and early neolithic sites of the Near East. http://www.exoriente.org/associated_projects/ppnd.php
Bos I, Caligari P (2008) Selection methods in plant breeding. Springer, Dordrecht
Bourgeois B, Munoz F, Fried G et al (2019) What makes a weed a weed? A large-scale evaluation of arable weeds through a functional lens. Am J Bot 106:90–100
Boyeldieu J (1980) Les cultures céréalières (Nouvelle Encyclopédie Des Connaissances Agricoles). Hachette, Paris
Colledge S (1998) Identifying pre-domestication cultivation using multivariate analysis. In: Damania AB, Valkoun J, Willcox G, Qualset CO (eds) The origins of agriculture and crop domestication. ICARDA, Aleppo (Syria), pp 121–131
Creanza N, Kolodny O, Feldman MW (2017) Cultural evolutionary theory: how culture evolves and why it matters. Proc Natl Acad Sci USA 114:7,782–7,789
De Moulins D (2000) The plant food economy Abu Hureyra 1 and 2. Abu Hureyra 2: plant remains from the neolithic. In: Moore AMT, Hillman GC, Legge AJ (eds) Village on the Euphrates: from foraging to farming at Abu Hureyra. Oxford University Press, Oxford, pp 399–416
Dietrich L, Meister J, Dietrich O, Notroff J, Kiep J, Heeb J, Beuger A, Schütt B (2019) Cereal processing at early neolithic Göbekli Tepe, southeastern Turkey. PLoS ONE 14:e0,215,214. https://doi.org/10.1371/journal.pone.0215214
Douché C, Willcox G (2018) New archaeobotanical data from the early neolithic sites of dja’de El-Mughara and tell Aswad (Syria): a comparison between the Northern and the Southern Levant. Paléorient 44:45–57
Fricano A, Brandolini A, Rossini L et al (2014) Crossability of Triticum urartu and Triticum monococcum wheats, homoeologous recombination, and description of a panel of interspecific introgression lines. G3 4:1,931–1,941. https://doi.org/10.1534/g3.114.013623
Fuller DQ, Stevens CJ (2018) The making of the Botanical Battle Ground: domestication and the origins of the worlds’ weed Floras. In: Lightfoot E, Liu X, Fuller DQ (eds) Far from the Hearth: essays in honour of Martin K. Jones. McDonald Institute for Archaeological Research, Cambridge, pp 9–21
Fuller DQ, Willcox G, Allaby RG (2012) Early agricultural pathways moving outside the ‘core area’ hypothesis in Southwest Asia. J Exp Bot 63:617–633
Fuller DQ, Denham T, Kistler L, Stevens C, Larson G, Bogaard A, Allaby R (2022) Progress in domestication research: explaining expanded empirical observations. Quat Sci Rev 296:107,737
Goring-Morris AN, Belfer-Cohen A (2008) A roof over one’s head: developments in Near Eastern residential architecture across the epipalaeolithic–neolithic transition. In: Bocquet-Appel J-P, Bar-Yosef O (eds) The neolithic demographic transition and its consequences. Springer, Dordrecht, pp 239–286. https://doi.org/10.1007/978-1-4020-8539-0_10
Hallauer AR, Darrah LL (1985) Compendium of recurrent selection methods and their application. Crit Rev Plant Sci 3:1–33. https://doi.org/10.1080/07352688509382202
Harlan JR (1995) The living fields: our agricultural heritage. Cambridge University Press, Cambridge
Harlan JR, Zohary D (1966) The distribution of wild wheats and barley. Science 153:1,074–1,080
Harris DR (1977) Alternative pathways toward agriculture. In: Reed CA (ed) Origins of agriculture. Mouton, The Hague, pp 179–243
Hillman GC (2000) The plant food economy Abu Hureyra 1 and 2. Abu Hureyra 1: the Epipalaeolithic. In: Moore AMT, Hillman GC, Legge AJ (eds) Village on the Euphrates: from foraging to farming at Abu Hureyra. Oxford University Press, Oxford, pp 327–399
Hillman GC, Davies MS (1990) Measured domestication rates in wild wheats and barley under primitive cultivation, and their archaeological implications. J World Prehist 4:157–222
Kislev ME (1992) Agriculture in the near east in the 7th millennium B.C. In: Anderson P (ed) Préhistoire De l’agriculture: nouvelles approches expérimentales et ethnographiques. Monographie Du CRA 6. Editions CNRS, Paris, pp 87–94
Kislev ME, Weiss E, Hartmann A (2004) Impetus for sowing and the beginning of agriculture: ground collecting of wild cereals. Proc Natl Acad Sci USA 101(2):692–695
Lupton FGH (1987) History of wheat breeding. In: Lupton FGH (ed) Wheat breeding: its scientific basis. Springer, Dordrecht, pp 51–70. https://doi.org/10.1007/978-94-009-3131-2_3
Maeda O, Lucas L, Silva F, Tanno K-i, Fuller DQ (2016) Narrowing the harvest: increasing sickle investment and the rise of domesticated cereal agriculture in the fertile crescent. Quat Sci Rev 145:226–237. https://doi.org/10.1016/j.quascirev.2016.05.032
Marais GF, Botes WC (2010) Recurrent mass selection for routine improvement of common wheat: a review. In: Lichtfouse E (ed) Organic farming, pest control and remediation of soil pollutants. Springer, Dordrecht, pp 85–105
Mazurowski R, Kanjou Y (eds) (2012) Tell Qaramel 1999–2007: protoneolithic and early pre-pottery neolithic settlement in Northern Syria. Polish Centre of Mediterranean Archaeology, University of Warsaw, Warsaw
Pajkovic M, Lappe S, Barman R et al (2014) Wheat alleles introgress into selfing wild relatives: empirical estimates from approximate Bayesian computation in Aegilops triuncialis. Mol Ecol 23:5,089–5,101. https://doi.org/10.1111/mec.12918
Peleg Z, Abbo S, Gopher A (2022) When half is more than the whole: wheat domestication syndrome reconsidered. Evol Appl 15(2):2,002–2,009. https://doi.org/10.1111/eva.13472
Purugganan MD, Fuller DQ (2011) Archaeological data reveal slow rates of evolution during plant domestication. Evolution 65:171–183
Redden RJ, Jensen NF (1974) Mass selection and mating systems in cereals. Crop Sci 14:345–350. https://doi.org/10.2135/cropsci1974.0011183X001400030001x
Smith BD (2001) Low-level food production. J Archaeol Res 9:1–43
Smith BD (2007) The ultimate ecosystem engineers. Science 315:1,797–1,798
Steensberg A (1986) Man the manipulator: an ethno-archaeological basis for reconstructing the past. National Museum of Denmark, Copenhagen
Tanno K-i, Willcox G (2006) How fast was wild wheat. Domesticated? Sci 311:1,886
Tanno K-i, Willcox G (2012) Distinguishing wild and domestic wheat and barley spikelets from early holocene sites in the Near East. Veget Hist Archaeobot 21:107–115
Tanno K-i, Willcox G, Muhesen S, Nishiaki Y, Kanjou Y, Akazawa T (2013) Preliminary results from analyses of charred plant remains from a Burnt Natufian Building at Dederiyeh Cave in Northwest Syria. In: Bar-Yosef O, Valla FR (eds) Natufian foragers in the Levant: terminal pleistocene social changes in Western Asia, vol 19. International Monographs in Prehistory, Ann Arbor, pp 83–87
Weide A, Green L, Hodgson JG et al (2022) A new functional ecological model reveals the nature of early plant management in southwest Asia. Nat Plants 8:623–634
Weide A, Hodgson JG, Leschner H et al (2023) The Association of Arable Weeds with modern wild cereal habitats: implications for reconstructing the origins of plant cultivation in the Levant. Environ Archaeol 28:296–311. https://doi.org/10.1080/14614103.2021.1882715
Willcox G (1996) Evidence for plant exploitation and vegetation history from three early neolithic pre-pottery sites on the Euphrates (Syria). Veget Hist Archaeobot 5:143–152
Willcox G (1999a) Agrarian change and the beginnings of cultivation in the Near East: evidence from wild progenitors, experimental cultivation and archaeobotanical data. In: Gosden C, Hather JG (eds) Prehistory of food: appetites for change. Routledge, London, pp 479–500
Willcox G (1999b) Archaeobotanical significance of growing Near Eastern progenitors of domestic plants at Jalès (France). In: Anderson PC (ed) Prehistory of agriculture: new experimental and ethnographic approaches. Monograph 40. University of California, Los Angeles, pp 103–118
Willcox G (2005) The distribution, natural habitats and availability of wild cereals in relation to their domestication in the Near East: multiple events, multiple centres. Veget Hist Archaeobot 14:534–541
Willcox G (2007) Agrarian change and the beginnings of cultivation in the Near East: evidence from wild progenitors, experimental cultivation and archaeobotanical data. In: Denham T, White P (eds) The emergence of agriculture: a global view. Routledge, New York, pp 217–241
Willcox G (2012) Searching for the origins of arable weeds in the Near East. Veget Hist Archaeobot 21:163–167
Willcox G (2013) The roots of cultivation in southwestern Asia. Science 341:39–40
Willcox G (2023) Can a functional ecological model reliably reveal the nature of early plant management in southwest Asia? Nat Plants 9:1,962–1,963
Willcox G, Herveux L (2013) Late Pleistocene/early Holocene charred plant remains: preliminary report. In: Mazurowski R, Kanjou Y (eds) Tell Qaramel 1999–2007: protoneolithic and early pre-pottery neolithic settlement in Northern Syria. Polish Centre of Mediterranean Archaeology, University of Warsaw, Warsaw, pp 120–130
Willcox G, Stordeur D (2012) Large-scale cereal processing before domestication during the tenth millennium cal BC in northern Syria. Antiquity 86:99–114
Willcox G, Fornite S, Herveux L (2008) Early holocene cultivation before domestication in northern Syria. Veget Hist Archaeobot 17:313–325
Zohary D, Hopf M, Weiss E (2012) The domestication of plants in the old world: the origin and spread of domesticated plants in Southwest Asia, Europe, and the Mediterranean Basin, 4th edn. Oxford University Press, Oxford
Acknowledgements
I would like to thank the late Gordon Hillman for introducing me to the vegetation of eastern Anatolia, in particular the wild einkorn stands, also Patricia Anderson for setting up the experimental cultivation of wild cereals at Jalès. I am grateful to Sue Colledge who pioneered the study of wild/weed taxa on early Neolithic sites. My appreciation goes to several members of the International Center for Agricultural Research in the Dry Areas, formerly based in Aleppo, who guided me to numerous wild cereal and pulse habitats in northern and southern Syria. During on-site field work in eastern Anatolia, northern and southern Syria, Tajikistan, Afghanistan and Armenia, supportive excavation directors provided transport and precious time for me to sample a wide range of vegetation types. My thanks also go to Dr Jean-Paul Mandin, formerly of the Lycée Agricole Oliver de Serres who with his students carried out the germination tests.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by A. Fairbairn.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Willcox, G. Sowing, harvesting and tilling at the end of the Pleistocene/beginning of the Holocene in northern Syria: a reassessment of cereal and pulse exploitation. Veget Hist Archaeobot (2024). https://doi.org/10.1007/s00334-023-00984-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00334-023-00984-4